Exploring the interactions between microbes
and plants in rhizosphere of arsenic
hyperaccumulators to improve arsenic
phytoremediation
著者
Yang Chongyang
学位授与機関
Tohoku University
学位授与番号
11301甲第19381号
Summary
Exploring the interactions between microbes and plants in
rhizosphere of arsenic hyperaccumulators to improve arsenic
phytoremediation
(ファイトレメディエーション効果改善のためのヒ素高蓄
積植物根圏における微生物-植物間相互作用の探究)
Chongyang Yang
Graduate School of Environmental Studies
Tohoku University
2019/03/05
Sendai, Japan
Graduate School of Environmental Studies
Examination Committee
Prof. Chihiro INOUE
(Chairman)
Prof. Keiko YAMAJI
(Member)
Prof. Yuyou LI
(Member)
Assist. Prof. Mei-Fang CHIEN
(Member)
1. Introduction
Phytoremediation is a promising eco-friendly technique to remove As contamination that employs plants to extract As from contaminated soil via their roots (Hettick et al., 2015). This process takes more time compared to other conventional techniques (e.g. soil excavation and disposal in a landfill), while it is effective when treating low As concentration or continuously As releasing by natural processes (Ghosh et al., 2011). Pteris vittata and Pteris multifida are two well known As hyperaccumulators that can accumulate up to 22,630 mg As kg -1 in dry biomass (Ma et al., 2001) and more than 1000
mg As kg -1 in fronds (Du et al., 2005), respectively.
Aside from plant, bacteria also play important role in As phytoremediation through changing its chemical form. Specifically, inorganic As transformations between As(V) and As(III) are commonly mediated by microbes mainly for energy generation and detoxification (Oremland and Stolz, 2003). Bacteria not only develop As resistance mechanisms but also utilize As as electron donors for physiological processes such as respiration (Paez-Espino et al., 2009). In aerobic soils, As(V) is usually present in very low concentrations in the soil solution because of strong adsorption by oxides/hydroxides of iron and aluminum (Zhao et al., 2010). On the other hand, As(III) is more soluble and generally considered to be more bioavailable than As(V). In a study conducted by Huang et al. (2011), P. vittata was reported to accumulate As(V) more efficiently than As(III). Given that two As species exist in soil and are absorbed by plants, intensification of As(III) oxidation and suppression of As(V) chelation in the rhizosphere could enhance As
removal by P. vittata. Beside As speciation, bacteria can provide specific compounds which promote growth of the plant. Specifically, plant-associated bacteria can produce the phytohormone indole acetic acid (IAA), which can promote plant growth, contributing to the enhancement of phytoremediation (Patten and Glick, 2002). Also, rhizobacteria can synthesize siderophores that can solubilize As adsorbed on Fe-oxides, releasing As for plant to absorb (Rajkumar et al., 2012). Better understanding of these processes has led to the development of conventional phytoremediation strategies for the cleanup of As. However, little information is available in the literature regarding how these bacteria work in the rhizosphere.
Understanding how plants affect rhizobacteria and vice versa is of paramount importance especially since interactions between the two have evolved adaptive and complicated strategies to resist and metabolize As (Mesa et al., 2017). One means of interaction is through the plant’s root exudates. Root exudates are assumed to be the major nutrient source for rhizobacteria that induce them to colonize near plant roots (Lugtenberg et al., 1999). Successful colonization is believed to be the key step to maximum their function. Moreover, from the view of plants, it can shape the soil microflora by secreting exudation into the rhizosphere which might supply carbon and nitrogen substrates for microbial growth (Hu et al., 2018). Root exudates have a multitude of functions on rhizosphere microbes by acting not only as signaling molecules, attractants, stimulants, but also as inhibitors or repellents (Baetz and Martinoia, 2014). Thus, root exudate metabolites may recruit functional rhizobacteria to take part in phytoremediation
synergistically. However, there is very limited information about how P. vittata grown on As polluted soils respond to shaping rhizobacteria and assisting As phytoremediation.
The objectives of this study are (1) to isolate and characterize rhizobacteria of As hyperaccumulators to find candidate strains which facilitate As phytoremediation; (2) to evaluate the long-term effectiveness of microbe assisted phytoremediation on As phytoremediation.
2. Materials and methods
2.1 As-resistant rhizobacteria isolation and identification
As-resistant bacteria were isolated from the rhizosphere of P. vittata and P. multifida cultivated in abandoned mine soil (As content exceeding 60 mg kg-1). The rhizospheric
soil of P. vittata and P. multifida was diluted appropriately with TE buffer which was consequently spread on 0.2X trypticase soy broth (TSB) agar plates containing with 1 mM NaAsO2 (As(III)) or 10 mM Na2HAsO4 (As(V)) at 30°C overnight. Colonies with
different morphotypes were selected and re-cultured in the 0.2X TSB medium. Genomic DNA was extracted and purified with the Wizard Genomic DNA Purification Kit (Promega, Madison, WI, USA). Isolates identification was carried out through PCR amplification of 16S rRNA gene by primers 8F with 1492R (Weisburg et al., 1991). The PCR product of 16S rDNA was sequenced and then submitted to GenBank. The accession numbers of isolates from P. vittata and P. multifida were MK934360-MK934408 and MK934428–MK934462, respectively.
The minimum inhibitory concentrations (MICs) of As(III) and As(V) was detected. The OD600nm of the initial stage and 48 h after incubation by 0.2X TSB medium contained
no As (negative control) or As(III) (1, 2, 4, 8, or 16 mM) or As(V) (1, 10, 80, 160, or 320 mM) were measured by a N60 Nanophotometer (IMPLEN, Munich, Germany). The MIC for each strain was defined as the lowest As concentration which showed no visible increase at OD600nm.
A primer pair aroA95F and aroA599R, described by Hamamura et al. (2019) were used to amplify the aioA-like genes. The isolated strains showed aioA-like genes amplicons were selected for investigations of As(III) oxidation. Selected isolates were incubated at a uniform concentration (OD600nm = 1) in 0.2X TSB medium containing 0.1
mM As(III) at 30°C with shaking at 200 rpm. The media inoculated without bacteria were used as controls. The bacterial suspensions were transferred into new tubes with 24 h of incubation. Arsenic species in the solutions were determined using As-Speciation Cartridges (Waters Co., Ltd, Milford, MA, USA), which adsorbed As(V) and excluded As(III). Arsenic concentration was measured by the NexION 2000 inductively coupled plasma mass spectrometer (ICP-MS; PerkinElmer, Waltham, MA, USA).
For determination of IAA production, selected isolates were cultured in 0.2X TSB medium with 500 μg mL-1 tryptophan. The strains were incubated at 30°C for 24 h with
shaking at 200 rpm. After that, 1.0 mL of the cell-free suspension was transferred into a tube and mixed vigorously with 0.5 mL Salkowski’s reagent (Patten and Glick, 2002). A pink color developed after 40 min of incubation at room temperature (RT, 25°C). The
IAA concentration was measured by absorbance at 530 nm. To standardize the curve, 3-Indoleacetic acid (Kanto Chemical CO., INC. Tokyo, Japan) was purchased and used. The production of siderophores was determined according to the chrome azurol-S (CAS) analytical method (Schwyn and Neilands, 1987). The uninoculated supernatant was used as a reference (Ar). After 3 h of incubation in the dark at RT, the decrease of sample absorbance at 630 nm was recorded (As). The siderophore units are defined as [(Ar-As)/Ar]*100 (Machuca and Milagres, 2003). The siderophore production was evaluated as the percentage of siderophore units in liquid medium.
2.3 Colonization of selected strains to P. vittata
For strain r507 colonization test to P. vittata, acclimated plants were rinsed with Milli-Q water and then immersed in 50 mL of strain r507 resuspended solution (OD600nm=1.0) or sterilized Milli-Q water as negative control for 30 min. Then the plants were transferred to 250 mL of 0.2X Hoagland solution (HS) with 1.1 g L-1 pyruvic acid. After 0, 24, and 48 h of cultivation, the roots of plants were washed with sterilized Milli-Q water three times. The final rinsed water was collected and used for DNA extraction by Genomic DNA Purification Kit (Promega, Promega, Madison, WI, USA). The roots were surface-sterilized by immersion in 1.2% sodium hypochlorite for 1 min and then washed in sterile Milli-Q water several times to remove surface sterilization agents. The sterilized plant samples were ground in a sterile mortar with a sterile pestle. The DNA of tissue was extracted by PowerSoil DNA Isolation Kit (MoBio Laboratories Inc., Carlsbad, CA, USA). The abundances of aioA-like genes of strain r507 were
determined by quantitative real-time PCR (qPCR) performed on a CFX ConnectTM Thermocycler (BioRad, USA). A primer set specific to aioA-like genes of strain r507 was designed and defined as r507aioA239F (5’-ATGACCAACGTGATAACGGC-3’) and r507aioA373R (5’-GTCCGGCGTGTACATGTATG-3’). The qPCR reaction conditions used were as follows; 95°C for 5 min followed by 40 cycles of 95°C for 45 sec, 60°C for 20 sec, and 72°C for 30 sec.
For colonization test to strain m318 to P. vittata, the similarly acclimated ferns were rinsed with Milli-Q water and immersed in sterilized medium (0.2X TSB) with or without the strain m318 culture for 30 min (OD600 nm =1.0). The plants were then moved into 250
mL 0.2X HS with 1.1 g L-1 pyruvic acid and grown for 2 days. Root samples were
retrieved every day after gently rinsing with sterilized Milli-Q water several times. The genomic DNA of the rhizosphere was extracted using the PowerSoil DNA Isolation Kit (MoBio Laboratories Inc., Carlsbad, CA, USA). Colonization effects were evaluated by the abundance of 16S rRNA and m318 aioA genes in the rhizosphere using a CFX ConnectTM Thermocycler (BioRad, USA). The qPCR primers for 16S rRNA were Eu341F and Eu518R (Arfken et al., 2017; Shigematsu et al., 2006). The primers for aioA-like genes of strain m318 were designed and defined as m318aioAF
(5’-ATAGAGCGCCAGAGCATCAT-3’) and m318aioAR
(5’-GGACAAACAGTGCGACGTTA -3’). The qPCR program for the 16S rRNA genes was 95°C for 30 s followed by 40 cycles at 95°C for 5 s, 55°C for 30 s. The qPCR program for the m318 aioA-like genes was 95°C for 30 s followed by 40 cycles of 95°C for 45 s,
55.5°C for 20 s, and 72°C for 30 s.
2.4 Inoculation of candidate strains with As hyperaccumulators in the field
Field trial site was located in Sendai city, Miyagi, Japan, which is in northeast area of Japan (38°17'23.4"N, 140°46'52.2"E). A comparative evaluation on the effect of strain r507 to P. vittata and strain m318 to P. vittata and P. multifida on As phytoremediation in field trials in 2015 and 2016, respectively. The treatment to fern was set as non-inoculation (PvN/PmN, control) and non-inoculation (PvI/PmI, ferns got inoculated before planting). Seedlings of P. multifida and P. vittata with 6-9 fronds were supplied by Fujita Co., Ltd (Tokyo, Japan). The bacterial strains were stored in nutrient broth with 25% glycerol at -80°C prior to testing. For this study, selected strains cultures were prepared in 20% strength trypticase soy broth (TSB) medium and growing on a rotary shaker (200 rpm min−1; 30°C) overnight. Cultures were inoculated to root of plants for one day before
transplantation. Plant samples with nylon mesh bags were collected almost every month and three biological replicates were performed. The harvested plants were washed and then air-dried for 3 days and then oven-dried at 50°C for one week. The dried plants were separated into shoot and root parts. Afterward, their dry weight biomass was measured and recorded. The total biomass of each plant was calculated. The dried materials (shoot or root parts) were ground (<1 mm) and digested by HNO3 (Wako, Japan). Arsenic
concentration was analyzed by the NexION 2000 inductively coupled plasma mass spectrometer (ICP-MS; PerkinElmer). The total As accumulation in each plant was calculated.
Rhizospheric soil samples were taken from nylon mesh bags with the plant. Soil tightly attached on the root surface of 0.2-0.5 g was carefully collected. Genomic DNA of soil was extracted by the PowerSoil DNA Isolation Kit (MoBio Laboratories Inc., Carlsbad, CA, USA). Microbial analysis was conducted to the abundance of total bacteria and bacteria harboring aioA-like genes in the rhizosphere by quantitative polymerase chain reaction (qPCR) of 16S rRNA and arsenite oxidase (aioA-like) genes using a CFX ConnectTM Thermocycler (BioRad, USA). Standards were prepared through a serial dilution of plasmids carrying the target genes. The primers Eu341F (5’-CCTACGGGAGGCAGCAG-3’) and Eu518R (5’-GTATTACCGCGGCTGCTGG-3’) were used to generate 178 bp amplicons of bacterial 16S rRNA genes (Arfken et al., 2017; Nyambura Ngamau, 2012). The qPCR primers of the bacterial aioA-like gene were
aroA95F (5′-TGYCABTWCTGCAIYGYIGG) and aroA599R
(5′-TCDGARTTGTASGCIGGICKRTT) (Hamamura et al., 2009). The ratio of aioA-like genes to 16S rRNA genes was calculated represented the proportion of bacteria containing aioA-like genes to total bacteria.
2.5 Statistical analysis
All experiments were conducted in triplicate for each treatment. The data were presented as the mean values of the repetitions with standard error. Statistical analyses were performed using the t-test and ANOVA by Tukey HSD.
3 Results and discussion
multifida
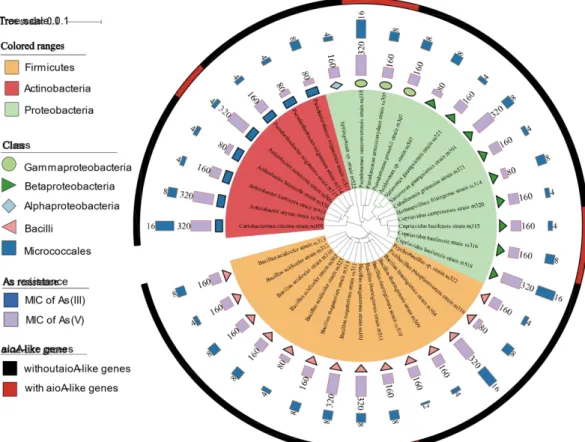
A total of 49 arsenic resistant rhizobacteria strains were isolated from rhizosphere samples of P. vittata. The isolates were numbered after a letter “r” or “v” which showed the source of samples as field trials or pot experiments, respectively. Phylogenetic analysis of the isolates was based on the alignment of 16S rRNA gene sequences and the construction of the phylogenetic tree using the neighbor-joining method (Fig. 1). At the phylum level, isolated rhizobacteria were mainly affiliated with the phyla Proteobacteria, Firmicutes, and Actinobacteria. Proteobacteria was the dominant phylum in the isolated rhizobacteria, representing up to 49% of total isolates, which was similar to the results of Han et al. (2017a). In the class levels, Bacilli occupied the largest proportion of the isolates, covering 37%. Several genera, such as Acinetobacter, Bacillus, Ralstonia, and Cupriavidus, have been reported as metal resistant rhizobacteria (Mondal et al., 2008; Rajbanshi, 2008; von Rozycki and Nies, 2009). The results of our previous study which analyzed the rhizosphere of P. vittata through culture-independent method showed that α-Proteobacteria was the most dominant community in the rhizosphere of P. vittata (Chien et al., 2015). The difference could be due to the bias of cultivation. Nonetheless, the culture-independent method could present a comprehensive picture of the microbiome while isolation could help to obtain useful bacteria.
A total of 35 As resistant rhizobacteria strains were isolated from the rhizosphere of P. multifida (Fig. 2). Phylogenetic analysis of the isolates suggested that these isolates were close to the phylum of Actinobacteria, Proteobacteria, and Firmicutes by
approximately 40%, 37%, and 23%, respectively. Among the class levels, the culturable rhizobacteria were predominantly represented by Bacilli, Micrococcales, Beta-Proteobacteria, Gamma-Proteobacteria. The main genera were Bacillus, Pseudomonas, Cupriavidus, and Arthrobacter. The compositions of the cultivable bacterial communities were similar to that of culturable rhizobacteria in P. vittata (Han et al., 2017a). This may be because both ferns belonging to Pteris L. of Pteridaceae (Du et al., 2005), which secrete similar substances to mediate the rhizosphere community. Moreover, plant species, growth stages and soil properties could influence how microflora is shaped in the rhizosphere (Pii et al., 2016; Schlemper et al., 2017). Among the cultured bacteria, many common soil bacterial genera such as Bacillus, Pseudomonas, Cupriavidus, and Arthrobacter have been reported for their metal resistance (Monsieurs et al., 2011; Barzanti et al., 2007; Achour et al., 2007).
Fig. 1 Diversity and characteristics of cultivable isolates from P. vittata. Color range identifies phyla within the tree. Different colorful domains stand for the class level of isolates. Purple bars and yellow bars represent the MICs of As(V) and As(III). The color strip is used for identification of aioA-like genes.
Fig. 2 Diversity and characteristics of cultivable isolates from P. multifida. Color range identify phyla within the tree. Differently colored domains indicate the class levels of the isolates. Bars represent the MICs of As(V) and As(III). Color strips are used for identification of aioA-like genes.
3.2 Arsenic tolerance of isolated strains from P. vittata and P. multifida
Arsenic resistance is the first criterion used to evaluate the isolates’ overall potential in improving As phytoremediation. MIC is the lowest concentration at which the isolate’s growth is completely suppressed. All 49 rhizobacteria isolates from P. vittata were evaluated for their potentially useful features (Fig. 1), including arsenic resistance and aioA-like gene presence. The majority of isolates were resistant to As(III) and As(V) at concentrations exceeding 8 mM and 160 mM, respectively. Ten strains showed resistance higher than 16 mM As(III) and 320 mM As(V) (r301, r302, r305, r306, r308, r502, r505,
r506, v309 and v513). The strains demonstrated flexible resistance to both As(III) and As(V) which incredibly assist the survival of these rhizobacteria when exposed to various arsenic species conditions.
Similarly, the MICs for As(V) and As(III) were examined for evaluating the As tolerance of the isolated strains from P. multifida (Fig. 2). The isolates were found to be tolerant of different As(V) and As(III) concentrations, and most of the isolates were resistant to As(V) and As(III) concurrently. Among all the strains, 47% were simultaneously tolerant to As(V) in excess of 160mM and As(III) in excess of 80 mM. Strains m308, m318, m504, and m518 showed the highest tolerance to both As(V) and As(III) that could reach up to 320 mM and 16 mM respectively. In this study, MICs for the isolates were found to be higher for As(V) than As(III), which suggested that As(III) is more toxic than As(V) to bacteria. Despite this, there are many bacteria that have high tolerance for both As(V) and As(III), which greatly facilitates their survival when exposed to different As species under varying ambient conditions (Wang et al., 2012).
3.3 The aioA-like genes identification on isolated strains from P. vittata and P. multifida The aioA-like gene is the most important gene associated with arsenite oxidation by bacteria (Inskeep et al., 2007). Among 49 isolates from P. vittata, aioA-like genes were detected in 7 isolates, in which 5 strains belonged to Proteobacteria and 2 strains belonged to Firmicutes (Fig. 1). It is conceivable that specific arsenic oxidizing rhizobacteria play dominant roles in transformation of arsenic in rhizosphere of P. vittata (Wang et al., 2012). Although a wide range of As-oxidizing isolates has been reported, cultivable rhizobacteria
with aioA-like genes isolated from the rhizosphere of P. vittata were limited (Das et al., 2014; Ghosh et al., 2011). These genes were also identified in six isolates from P. multifida that were used for preliminary screening of the As-oxidizing rhizobacteria (Fig.
2).
3.4 Rapid As(III) oxidation ability determination on isolated strains from P. vittata and P. multifida
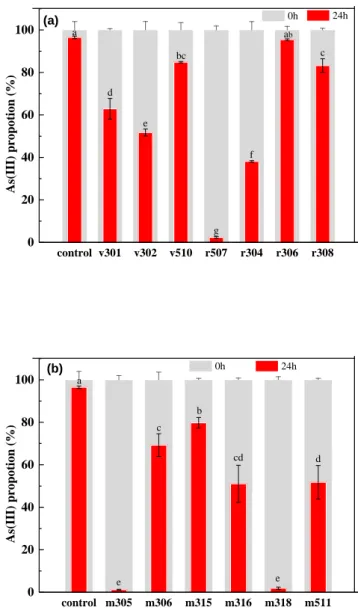
To determine arsenic redox ability, 7 isolates from P. vittata harboring aioA-like genes were incubated in 0.2X TSB medium spiked with 0.1 mM of As(III) or As(V). Fig.
3(a) shows the As(III) conversion with bacterial growth. As demonstrated by the
comparison of the proportion of remaining As(III) after 24 h incubation, strain r507 exhibited extraordinarily rapid As(III) oxidation ability, with 98% of As(III) oxidized to As(V), while the average oxidation by other isolates was 5–62%. Among them, the lowest As(III) oxidation was observed in strain r306. Arsenite oxidation is considered a detoxification pathway in microorganisms. Because arsenite is more toxic than arsenate, some bacteria oxidize arsenite to arsenate by a specific enzyme, arsenite oxidase, to reduce the toxicity of arsenite (Stolz et al., 2002). Previous studies showed the efflux of arsenite to the external environment from the P. vittata roots to lessen arsenic stress, but the presence of arsenate inside the plant lead to the identification of arsenite oxidizers in the rhizosphere of P. vittata, indicating that bacterial arsenite oxidation greatly facilitated As hyperaccumulation in the plant (Chen et al., 2016; Wang et al., 2012). Considering the bacterial traits needed in phytoextraction of arsenic with P. vittata, accelerated arsenite
oxidation ability is crucial especially in the mobilization of insoluble arsenite to arsenate and subsequently efficient uptake by P. vittata.
control v301 v302 v510 r507 r304 r306 r308 0 20 40 60 80 100 g f e d c bc ab As (III ) p ropotion (%) 0h a 24h (a) control m305 m306 m315 m316 m318 m511 0 20 40 60 80 100 e c cd e b d As (III ) p ropotion (%) 0h a (b) 24h
Fig. 3 Arsenic transformation of 7 isolates from P. vittata (a) and P. multifida (b) with aioA-like genes in 0.2X TSB medium spike with 0.1 mM As(III).
Rapid As(III) oxidation was also observed to these 6 strains from P. multifida. Isolates can oxidize 20–98% of 0.1mM As(III) in 200 μL within 24 h. Strains m305 and m318
demonstrated extraordinary As(III) oxidation ability (Fig. 3(b)). It is known from literature that As(V) uptake by P. vittata is more efficient than As(III) (Huang et al., 2011). Thus, microbial rapid oxidation of As(III) to As(V) can enhance phytoremediation by transforming the more mobile and toxic As(III) to the less toxic As(V), which is absorbed more easily by P. vittata (Wang et al., 2012). Han et al. (2017b) found that aioA-like genes were 50 times more abundant than arsenate reductase (arsC) genes in the rhizosphere of P. vittata, which implied that As-oxidizing bacteria play a more active role in the host accumulation of As by transforming As(III) to As(V). Furthermore, Han et al. (2016) reported that inoculation of an As-oxidizer resulted to the highest accumulated As concentration in P. vittata.It hinted that increasing the abundance of aioA-like genes in the rhizosphere might promote As phytoremediation. The aioA-like genes provide a detoxification system in microorganisms that promotes As(III) oxidation to As(V) (Suhadolnik et al., 2017). However, only a small proportion of the isolates harboring the aioA-like gene were detected in our study. This indicated that culturable bacteria with the ability to detoxify As(V) (via reducing to As(III)) could be the predominant species in the rhizosphere soil (Ghosh et al., 2011).
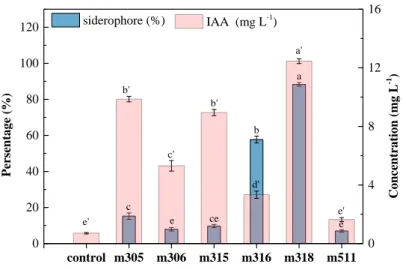
control m305 m306 m315 m316 m318 m511 0 20 40 60 80 100 120 e' e' d' c' b' b' a' e e ce c b siderophore (%) Pers entage (%) a 0 4 8 12 16 IAA (mg L-1) Concentration (mg L -1 )
Fig. 4 Siderophore and IAA secretion of selected isolates from P. multifida.
3.5 IAA and siderophore secretion of isolated strains with aioA-like genes from P. multifida
Aside from As speciation, plant growth promoting traits of rhizobacteria can boost plant growth hindered by a stressed environment. In relation to this, IAA and siderophore secretions of the above six isolates were detected (Fig. 4). Isolated strains had the capacity to produce IAA at levels of 1.6–12.45 mg L-1, while the siderophore units were 7–88%.
Strain m318 exhibited a unique superiority in IAA production (12.45 mg L-1) and high
quantities of siderophore units (88%) among all the strains (Fig. 4). In soil, As often binds with metals (i.e. Ca, Al, and Fe), thus making them unavailable for plant uptake (Liu et al., 2016). Siderophores produced by rhizobacteria could release the As adsorbed on Fe-oxides, thus making both As and Fe absorption easier for plants (Jeong et al., 2014). This means that siderophores not only improve As phytoextraction by release of As but also
indirectly promote the growth of plant biomass by supplying trace elements such as Fe (Rajkumar et al., 2012). IAA is a major endogenous auxin of the higher plants that facilitates plant growth (Duca et al., 2014). Recent studies found that most As-resistant endophytic bacteria from P. vittata and P. multifida can synthesize IAA and IAA levels of isolates from P. vittata was much higher than that from P. multifida (Zhu et al., 2014; Xu et al., 2016). In this study, it was found that the IAA levels of As-resistant rhizobacteria were slightly lower than that of the reported As-resistant endophytic bacteria from these two As hyperaccumulators (Zhu et al., 2014; Xu et al., 2016). Thus, it is worth exploring how the different production levels of IAA by rhizobacteria and endophyte can ultimately affect plant growth during phytoremediation. On the basis of rapid As oxidation ability and multiple plant growth promoting traits, strain m318 demonstrated the most positive effect and was thus selected for the subsequent experiments.
3.6 Colonization of strain r507 and strain m318 to P. vittata
strain r507 was inoculated to P. vittata, and the abundance of r507 specific aioA genes in the surface and interior of the root was determined (Fig. 5). In the root surface, the abundance of r507 specific aioA after 48 h of inoculation was 6.74 × 108 copies μg-1 DNA,
which was significantly higher than that of without inoculation (1.59 × 108 copies μg-1
DNA). Similar results were observed in the interior of the root, which was 9.78 × 108
copies μg-1 DNA for with inoculation against 1.45 × 108 copies μg-1 DNA in
non-inoculated samples. Strain r507 was isolated from rhizosphere of P. vittata, hence the exhibited affinity to root of P. vittata with or without inoculation. However, the results
stressed that the strain r507 was not only able to successfully colonize the rhizosphere of P. vittata, but also was able to enter and proliferate inside the root.
Fig 5 The abundance of r507aioA genes in surface and interior of roots with inoculation of strain r507.
Colonization of the root system is required to maximize the beneficial effect of the inoculated bacteria. However, microbial motility alone does not suffice to allow successful colonization of the root. Some leguminous plants and microbes have evolved complex signal exchange mechanisms that allow a specific bacterial species to induce its host plant to form invasion structures through which the bacteria can enter the plant root (Jones et al., 2007). The species Cupriavidus basilensis is a Gram-negative bacterium which have been reported to utilize root exudates and organic compounds released by living plants’ roots in their surrounding environment (Koopman et al., 2010; Mesa et al., 2017). The strain r507 easily colonized and persisted in both the surface and interior of P. vittata’s root. These abilities raise the possibility of its increased survival in the
0 2 4 6 8 10 uninoculated d' d' c'd' b'c' b' a' d d c c b inoculated inoculated Root interior r507 aioA gene c opies (10 8 g -1 DNA ) 0h 24h 48h Root surface uninoculated a
environment as it can readily seek shelter in P. vittata’s roots. More interestingly, since P. vittata absorbs mainly arsenate, the presence of an arsenic oxidizer r507 in its root could be a foundation for a plant-microbe co-cultivation system with a more efficient arsenic phytoextraction. Colonization effects were evaluated by the abundance of 16S rRNA and m318aioA genes after inoculation with strain m318 for 2 days. The absolute copy numbers of m318aioA genes were normalized with those of the 16S rRNA gene copies (Fig. 6). It was noted that the inoculated treatments increased m318aioA gene frequencies to 16S rRNA gene. Effective colonization of the root system by isolates is crucial for enabling the beneficial effects of inoculants. Root exudates play a vital role in stimulating the microorganisms present in the rhizosphere by supplying nutrients (Bacilio-Jiménez et al., 2003). In this research, strain m318 successfully colonized the rhizosphere of P. vittata (Fig. 6). 0.00 0.05 0.10 0.15 d d c c b a Uninoculated Inoculated Copy ratio of m 318 aio A gene c opies to 16S rRNA gene 0h 24h 48h
Fig. 6 The ratio of m318aioA genes to 16S rRNA genes with the inoculation of strain m318 in 48 h.
3.7 The effect of strain r507 on As phytoremediation by P. vittata
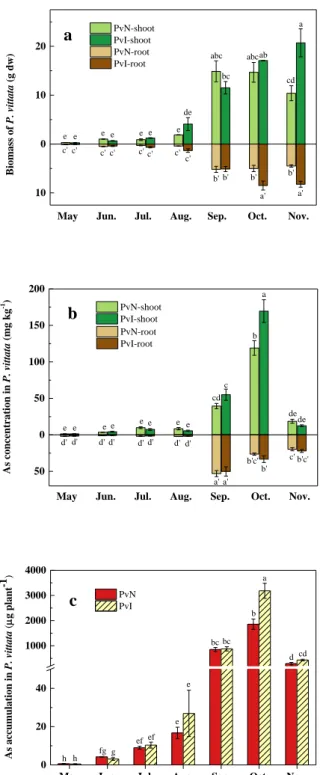
The effect of co-cultivation with Cupriavidus basilensis strain r507 and P. vittata was investigated. The field trial was conducted from May to November 2015, during which P. vittata was cultivated with or without inoculation of strain r507 (PvI or PvN). The change of biomass in PvI and PvN was shown in Fig. 7(a). There was almost no marked change observed in the biomass for the first four months. However, a significant increase was observed from September, calculated as nearly 48-fold increase in shoot of PvI against the initial state. In addition, the shoot and root biomass of PvN decreased from October to November, while PvI revealed a gradual upward trend. Although the shoot of PvN exhibited better growth than that of PvI in September, the shoot of PvI increased in October and November, which was 1.14-fold and 1.99-fold of that of PvN, respectively. Similarly, the root biomass of PvI showed satisfactory growth in the last two months, with amounts of 1.69-fold and 1.83-fold of that of PvN, respectively.
Arsenic concentration in P. vittata shoots and roots was shown in Fig. 7(b). Arsenic concentration in plant shoots and roots visibly increased from September and peaked in October and September, respectively. Arsenic concentration in shoots of PvI was higher than that in PvN, up to 170 mg kg-1 in October, whereas the arsenic concentration in both
PvI and PvN exhibited a sharp decline in November. Arsenic concentration in the roots peaked in September, slightly earlier than in the shoots. However, arsenic concentration in the roots was much lower than that in the shoot, which indicated that arsenic is predominantly accumulated in the shoots of P. vittata, and the roots mainly function to
Fig. 7 Field-scale evaluation of As phytoextraction in P. vittata. (a) Biomass of P. vittata (g dw); (b) As concentration (mg kg-1 dw) in P. vittata; (c) As accumulation (μg plant-1) in P. vittata.
absorb and translocate arsenic to the fronds (Tu et al., 2002). A calculated As
10 0 10 20 PvN-root PvI-root Biomass of P. vitt ata (g d w)
May Jun. Jul. Aug. Sep. Oct. Nov. PvN-shoot PvI-shoot a c' c' c' c' c' c' c' c' b' b' b' b' a' a' e e e e e e e de cd bc abc abc ab a 50 0 50 100 150 200 d' d' d' d' d' d' d' d' c' b'c' b'c' b' a' a' e e e e e e e e de de cd c b PvN-root PvI-root As conce nt ration i n P. vitt ata ( mg kg -1)
May Jun. Jul. Aug. Sep. Oct. Nov. PvN-shoot
PvI-shoot
a b
May Jun. Jul. Aug. Sep. Oct. Nov.
0 20 40 1000 2000 3000 4000 h h fg g ef ef e e d cd bc bc b As acc um ulat ion i n P. vitt ata ( g plan t -1) PvN PvI a c
accumulation in each plant based on biomass and As concentration was shown in Fig.
7(c). The maximum amount of As accumulation in shoots and roots was observed in
October and September, respectively. The highest amount of arsenic accumulation in shoots of PvI and PvN was 3,184 μg plant-1 and 1,858 μg plant-1, respectively. This
significant difference suggested that strain r507 efficiently promoted the accumulation of As by P. vittata.
P. vittata, as a tropical-zone fern, usually grows in the tropical or subtropical zone and cannot tolerate extreme cold such that the germination and growth rates of P. vittata were limited even near 25°C (Wan et al., 2010). In this study, P. vittata acclimated to the field for the first four months, and the biomass significantly increased from September along with the temperature (maximum temperature: 24.4°C). However, the increase in biomass was halted in October and November, which may be due to the decrease in temperature (minimum temperature: 11.6°C to 7.7°C). Nonetheless, the results showed that inoculation with strain r507 lessened the stress of low temperature to P. vittata. In addition, inoculation of strain r507 facilitated As uptake by P. vittata and resulted in a high As concentration in PvI. Though the As concentration in P. vittata greatly declined in November because of the decrease in temperature, the effect of strain r507 inoculation on improvement of As accumulation was noteworthy. This can be improved further by optimizing the time frame of phytoremediation or employing cold-tolerant hyperaccumulators.
aioA-like genes are important genes associated with arsenite oxidation needed for arsenic accumulation by P. vittata. The copy ratio of aioA-like genes to 16S rRNA genes in rhizosphere was shown in Fig. 8. The aioA-like genes occupy 62.7% of 16S rRNA genes in the rhizosphere of PvI, which was significantly higher than that of in PvN (16.7%) at initial stage. These phenomena can be accounted to the inoculated strain r507 which might have dominated the bacterial composition of PvI rhizosphere. Further, the copy ratio of aioA-like genes in rhizosphere of PvI was continuously higher than that of PvN in the following four months (June, July, August and September (no statistical difference)). The inoculation treatments increased the ratio of aioA-like genes by 6%-46% (ratio of PvI-ratio of PvN) from May to September. This trend elevated ratio of aioA to 16S rRNA genes) was lost in October and November, in which the rhizosphere might be affected by the low temperature.
The rhizospheric microbial community is more diverse than that of the surrounding bulk soil, which suggests the possibility for utilization of rhizospheric microorganisms to address challenges in soil ecology. Improving the efficiency of phytoremediation by inoculation may cause a transient perturbation of the equilibrium of soil microbial communities (Trabelsi and Mhamdi, 2013). However, bacterial acclimation may cause a new flora to be formed in root surroundings and no loss of function happens because of bacterial redundancy. In this study, the strain r507 successfully colonized the rhizosphere, which may have evolved strategies to increase the dominancy of aioA-like genes in the rhizosphere, consequently conferring the higher rate of arsenite oxidation. The above shift
resulted in an increment of arsenic accumulation in inoculated plants. A time lag between the dominant existence of aioA containing bacteria and the arsenic accumulation by P. vittata was observed, which might be due to the vegetation characteristics of P. viitata. Meanwhile, plant-soil-biota interactions may induce the bacterial community structure to restore to its original status, which may be the reason why aioA-like genes in the last two months showed no obvious increase with inoculation (Fig. 8). It is noticeable that the ratio of aioA-like genes in the rhizosphere
Fig. 8 The ratio of aioA-like genes to 16S rRNA genes in rhizosphere of P. vittata.
of PvN was higher than that of PvI in October, which can be accounted to the higher concentration of As(III) in rhizosphere of PvN than PvI (Fig. 3.3b). Enriched arsenite may have recruited more rhizobacteria with the capacity of detoxifying arsenite,through aioA-like genes resulting to their dominance in the rhizosphere soil.
May Jun. Jul. Aug. Sep. Oct. Nov. 0.0 0.2 0.4 0.6 ns * * *** * Ratio of aioA-lik e genes to 16S rRNA genes PvN PvI *** ns
In this study, an arsenite oxidizing rhizobacterium named Cupriavidus basilensis strain r507, which was isolated from a rhizospheric sample of P. vittata and displayed promising levels of root colonization. In the field-scale evaluation trial, P. vittata inoculated with strain r507 exhibited a notable increase in As accumulation. Our research showed a successful combination of P. vittata and rhizobacteria as an efficient method of arsenic phytoextraction through co-cultivation. Further studies exploring rhizobacteria and plant-microbe interactions at the laboratory- and field-scale are needed to optimize the efficacy of this method.
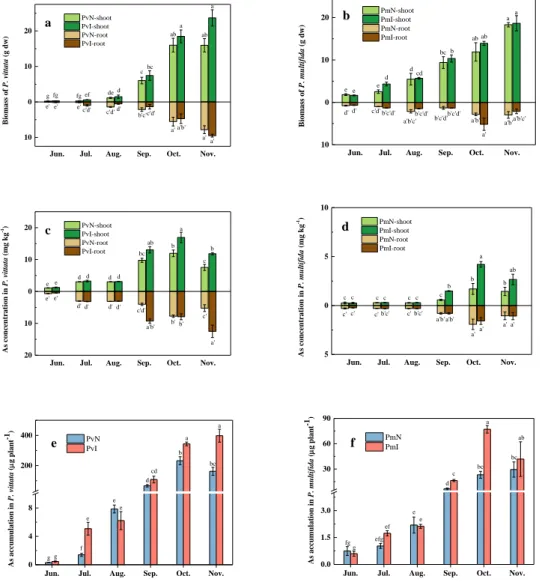
3.8 The effect of strain m318 on As phytoremediation by P. vittata and P. multifida The effects of strain m318 on plant biomass, As concentration, and As accumulation for P. vittata and P. multifida are shown in Fig. 9. Biomass is a factor of great practical relevance for evaluation of the effect of biological inoculum. In this study, noticeable effects were not observed during the first 3 months, but dramatic increases on the biomass of the shoots, As uptake, and As accumulation from September onwards were recorded. The shoot biomass of the ferns in November reached maximum with 23.70 g dw (PvI11-shoot) and 18.66 g dw (PmI11-(PvI11-shoot). There was no statistically significant difference between the inoculated and uninoculated treatments. Similarly, the root biomass of both ferns achieved significant growth from September onwards. The roots biomass of P. vittata achieved maximum in November with 9.51 g dw (PvI11-root), whereas the largest root biomass of P. multifida was recorded in October with 5.19 g dw (PmI10-root). Similar to this trend, As concentration in the shoots of both P. vittata and P. multifida
showed little increase in the first 3 months but significantly increased from September. The maximum values of As concentration for the two ferns were in October (PvI10-shoot: 16.93 mg kg-1, PmI10-shoot: 4.20 mg kg-1). Subsequently, As concentration declined in
November in the shoots of the two ferns. On the basis of the dry weight biomass and As concentrations of the plants,As accumulation for each of the plants is shown in Fig. 9(e) and Fig. 9(f). As accumulation displayed a gradual increase from June to October. P. vittata exhibited the highest As accumulation in October with 397.99μg plant-1 for PvI10,
whereas P. multifida was 77.07 μg plant-1 for PmI10. It was found that As accumulation
in P. vittata was more than that in P. multifida, and inoculated treatments showed greater amount than that of uninoculated treatments.
The abundance of strain m318 and rhizobacteria with aioA-like genes with reference to the total rhizobacterial population in the two ferns was calculated, and the absolute copy numbers of aioA-like genes were normalized with that of the 16S rRNA genes (Fig 10). In both ferns, the copy ratios of m318aioA genes to 16S rRNA genes exhibited significant increase in inoculated treatments during the initial period. Subsequent inoculum amendment did not show any significant difference in the relative copy numbers of the m318aioA genes with non-inoculation in both ferns. The copy ratio of the universal aioA-like genes to 16S rRNA genes presented similar enhancement with inoculation at the startup phase, whereas only marginal increase was found in subsequent sampling points with no statistically significant differences in both ferns.
20 10 0 10 20 PvN-root PvI-root As conce nt ration i n P. vitt ata ( mg kg -1)
Jun. Jul. Aug. Sep. Oct. Nov.
PvN-shoot PvI-shoot c d' d' d' b' e d d d b e' d' c'd' b' a'b' c' a' ab b a bc c d e e' 5 0 5 10 PmN-root PmI-root As conce nt ration i n P. m ult ifi da ( mg kg -1)
Jun. Jul. Aug. Sep. Oct. Nov.
PmN-shoot PmI-shoot d c' c' b'c' c' a'b' a' a' a' a' c c c c c c b b c' a'b' b'c' ab b a c
Jun. Jul. Aug. Sep. Oct. Nov.
0 4 8 200 400 g g f e e e d cd bc b a PvN PvI As acc um ulat ion i n P. vitt ata ( g plan t -1) a e
Jun. Jul. Aug. Sep. Oct. Nov.
0.0 1.5 3.0 30 60 90 bc fg g efg ef e e d c bc ab PmN PmI As acc um ulat ion i n P. m ult ifi da ( g plan t -1) a f
Fig. 9 Field-scale evaluation of As phytoremediation in P. vittata and P. multifida: (a) biomass of
P. vittata (g dw); (b) biomass of P. multifida (g dw); (c) As concentration (mg kg-1 dw) in P. vittata;
(d) As concentration (mg kg-1 dw) in P. multifida; (e) As accumulation (μg plant-1) in P. vittata. (f)
As accumulation (μg plant-1) in P. multifida.
The strain m318 was isolated from P. multifida while the results of this study suggested that P. vittata, the most well studied As hyperaccumulator, is also a good host plant for this strain. The effect of phytoextraction was inconspicuous in the first 3 months because of the acclimatization and vegetative stages of the plants. Subsequently, the plant biomass
10 0 10 20 PvN-root PvI-root Biomass of P. vitt ata (g d w)
Jun. Jul. Aug. Sep. Oct. Nov.
PvN-shoot PvI-shoot a e' e' d' c'd' c'd' c'd' b'c' a'b' a' a' a' ab ab a a bc c d de ef fg fg e' g 10 0 10 20 PmN-root PmI-root Biomass of P. m ult ifi da (g d w)
Jun. Jul. Aug. Sep. Oct. Nov.
PmN-shoot PmI-shoot b d b'c'd' b'c'd' b'c'd' a'b'c' a'b' c'd' d' b'c'd' a'b'c' a'b' a' ab ab a a bc b cd d e e d' e
displayed an insignificant difference between the inoculated and uninoculated treatments (Fig. 9(a) and 9(b)). However, inoculation increased As uptake by plants, which could be accounted to the strain m318 exerting its potential functions in the rhizosphere. It has been demonstrated that microthermal climates affect plant transpiration, growth, and metabolism, and therefore both uptake and elimination efficiency of pollutants are affected (Yu et al., 2007). Hence, As efflux may occur in November (mean temperature: 8.6 °C), leading to the decrease in As concentration in both plants. In fact, once As(V) has been taken up, it is reduced rapidly to As(III) in the rhizomes and then translocated to the fronds of P. vittata, which is considered to be a critical step for As(V) detoxification in plants (Han et al., 2017c). However, As(III) efflux may occur in the root under high As and other extreme environmental stresses (Han et al., 2016). To counter this, As-oxidizing rhizobacteria may circularly mediate As(III) oxidation and consequent As(V) uptake by plants. This phenomenon stresses may prompt the optimization of harvest timing of plants used in phytoremediation.
Jun. Jul. Aug. Sep. Oct. Nov. 0.0 0.2 0.4 0.6 * * ns ** ns PvN PvI Ratio of aioA -li ke genes to 16S rRNA genes **
a
Jun. Jul. Aug. Sep. Oct. Nov. 0.0 0.2 0.4 0.6 * ** ** ns * PmN PmI Ratio of aioA -li ke genes to 16S rRNA genes **
b
Fig. 10 Copy ratio of aioA-like genes to 16S rRNA genes in rhizospheres of (a) P. vittata and (b)
P. multifida in field-scale evaluations.
In terms of rhizosphere bacterial community (Fig. 10), it was found that the proportion of bacteria harboring aioA-like genes was increased by the inoculation, which might have recruited bacteria with aioA-like genes to aggregate in the rhizosphere, and have exerted synergetic As(III) oxidation, finally resulting to an increase in As absorption
by plants. Microbial motility has evolved with strategies to help microorganisms respond to external environmental circumstances, such as temporal and spatial changes in the stimuli (Yazdi and Ardekani, 2012). Hence, microorganisms in the natural environment often exist as multicellular aggregates. It should be noted that bacterial aggregation was selectively formed in the rhizospheres of our plants, which may account for the improvement of As phytoextraction by the inoculated treatments. This approach harnesses the potential of multifunctional rhizobacteria in conjunction with As hyperaccumulators that further broadens the possibilities of As phytoextraction, while a long-term field trails verification experiment is necessary.
4. Conclusion
In this study, Cupriavidus basilensis strain r507 and Pseudomonas vancouverensis strain m318 strains was successfully isolated from P. vittata and P. multifida, which showed several promising potential traits that may facilitate As phytoremediation, such as higher As tolerance, with aioA-genes, As oxidation, siderophore and IAA secretion, high affinity to P. vittata. Long-term field trials demonstrated that inoculation with strain r507 and strain m318 increased the As(III)-oxidase gene in the rhizospheres of As hyperaccumulators resulting to an improved As phytoextraction. This research sheds light on the combination of As hyperaccumulators and rhizobacteria as the efficient methods of arsenic phytoextraction through co-cultivation. Further studies exploring efficient rhizobacteria and plant-microbe interactions at the laboratory- and field-scale are needed
Reference
Achour, A.R., Bauda, P., Billard, P., 2007. Diversity of arsenite transporter genes from arsenic-resistant soil bacteria. Res. Microbiol. 158, 128–137.
Arfken, A., Song, B., Bowman, J.S., Piehler, M., 2017. Denitrification potential of the eastern oyster microbiome using a 16S rRNA gene based metabolic inference approach. PLoS One 12, e0185071. Bacilio-Jiménez, M., Aguilar-Flores, S., Ventura-Zapata, E., Pérez-Campos, E., Bouquelet, S., Zenteno, E.,
2003. Chemical characterization of root exudates from rice (Oryza sativa) and their effects on the
chemotactic response of endophytic bacteria. Plant and Soil, 249, 271–277.
https://doi.org/10.1023/A:1022888900465
Barzanti, R., Ozino, F., Bazzicalupo, M., Gabbrielli, R., Galardi, F., Gonnelli, C., Mengoni, A., 2007. Isolation and characterization of endophytic bacteria from the nickel hyperaccumulator plant Alyssum bertolonii. Microb. Ecol. 53, 306–316.
Baetz, U., Martinoia, E., 2014. Root exudates: the hidden part of plant defense. Trends Plant Sci. 19, 90– 98. https://doi.org/10.1016/j.tplants.2013.11.006
Chen, Y., Fu, J. W., Han, Y. H., Rathinasabapathi, B., & Ma, L. Q., 2016. High As exposure induced substantial arsenite efflux in As-hyperaccumulator Pteris vittata. Chemosphere. 144, 2189-2194. Chien, M. F., F., Makita, R., Sugawara, K., & Inoue, C., Study on As uptake and rhizobacteria of two as
hyperaccumulators forward to As phytoremediation. Advanced Materials Research, Vol. 1130. Trans Tech Publ, 2015, pp. 568-571.
Das, S.,Jean, J. S., Kar, S., Chou, M. L., & Chen, C. Y., 2014. Screening of plant growth-promoting traits in arsenic-resistant bacteria isolated from agricultural soil and their potential implication for arsenic bioremediation. Journal of hazardous materials. 272, 112-120.
Du, W., Li, Z., Zou, B., Peng, S., 2005. Pteris multifida Poir., a new arsenic hyperaccumulator: characteristics and potential. Int. J. Environ. Pollut. 23, 388–396.
Duca, D., Lorv, J., Patten, C.L., Rose, D., Glick, B.R., 2014. Indole-3-acetic acid in plant–microbe interactions. Antonie Van Leeuwenhoek 106, 85–125. https://doi.org/10.1007/s10482-013-0095-y Ghosh, P, Rathinasabapathi, B., Ma, L.Q., 2011. Arsenic-resistant bacteria solubilized arsenic in the growth
media and increased growth of arsenic hyperaccumulator Pteris vittata L. Bioresour. Technol. 102, 8756–8761. https://doi.org/10.1016/j.biortech.2011.07.064
Hamamura, N., Macur, R.E., Korf, S., Ackerman, G., Taylor, W.P., Kozubal, M., Reysenbach, A.-L., Inskeep, W.P., 2009. Linking microbial oxidation of arsenic with detection and phylogenetic analysis of arsenite oxidase genes in diverse geothermal environments. Environ. Microbiol. 11, 421–431.
https://doi.org/10.1111/j.1462-2920.2008.01781.x
Han, Y.-H., Fu, J.-W., Chen, Y., Rathinasabapathi, B., Ma, L.Q., 2016. Arsenic uptake, arsenite efflux and plant growth in hyperaccumulator Pteris vittata: Role of arsenic-resistant bacteria. Chemosphere 144, 1937–1942. https://doi.org/10.1016/J.CHEMOSPHERE.2015.10.096
Han, Y.-H., Jia, M.-R., Liu, X., Zhu, Y., Cao, Y., Chen, D.-L., Chen, Y., Ma, L.Q., 2017a. Bacteria from the rhizosphere and tissues of As-hyperaccumulator Pteris vittata and their role in arsenic transformation. Chemosphere 186, 599–606.
Han, Y.H., Fu, J.W., Xiang, P., Cao, Y., Rathinasabapathi, B., Chen, Y.S., Ma, L.Q., 2017b. Arsenic and phosphate rock impacted the abundance and diversity of bacterial arsenic oxidase and reductase genes
in rhizosphere of As-hyperaccumulator Pteris vittata. J. Hazard. Mater. 321, 146–153.
https://doi.org/10.1016/j.jhazmat.2016.08.079
Han, Y.-H., Liu, X., Rathinasabapathi, B., Li, H.-B., Chen, Y., Ma, L.Q., 2017c. Mechanisms of efficient As solubilization in soils and As accumulation by As-hyperaccumulator Pteris vittata. Environ. Pollut. 227, 569–577. https://doi.org/10.1016/J.ENVPOL.2017.05.001
Hettick, B.E., Cañas-Carrell, J.E., French, A.D., Klein, D.M., 2015. Arsenic: A Review of the element’s toxicity, plant interactions, and potential methods of remediation. J. Agric. Food Chem. 63, 7097– 7107. https://doi.org/10.1021/acs.jafc.5b02487
Hu, L., Robert, C.A.M., Cadot, S., Zhang, X., Ye, M., Li, B., Manzo, D., Chervet, N., Steinger, T., Van Der Heijden, M.G.A., Schlaeppi, K., Erb, M., 2018. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 9, 1–13. https://doi.org/10.1038/s41467-018-05122-7
Huang, Y., Hatayama, M., Inoue, C., 2011. Characterization of As efflux from the roots of As hyperaccumulator Pteris vittata L. Planta. 234, 1275–1284. https://doi.org/10.1007/s00425-011-1480-2
Inskeep, W. P., Macur, R. E., Hamamura, N., Warelow, T. P., Ward, S. A., & Santini, J. M., 2007. Detection, diversity and expression of aerobic bacterial arsenite oxidase genes. Environmental Microbiology. 9, 934-943.
Jeong, S., Moon, H.S., Nam, K., 2014. Enhanced uptake and translocation of arsenic in Cretan brake fern (Pteris cretica L.) through siderophorearsenic complex formation with an aid of rhizospheric bacterial activity. J. Hazard. Mater. 280, 536–543. https://doi.org/10.1016/j.jhazmat.2014.08.057 Jones, K. M., Kobayashi, H., Davies, B. W., Taga, M. E., & Walker, G. C., 2007. How rhizobial symbionts
invade plants: the Sinorhizobium–Medicago model. Nature Reviews Microbiology. 5, 619. Koopman, F., Wierckx, N., de Winde, J. H., & Ruijssenaars, H. J., 2010. Identification and characterization
of the furfural and 5-(hydroxymethyl) furfural degradation pathways of Cupriavidus basilensis HMF14. Proceedings of the National Academy of Sciences. 107, 4919–4924.
Liu, X., Fu, J.-W., Guan, D.-X., Cao, Y., Luo, J., Rathinasabapathi, B., Chen, Y., Ma, L.Q., 2016. Arsenic induced phytate exudation, and promoted FeAsO4 dissolution and plant growth in
As-Hyperaccumulator Pteris vittata. Environ. Sci. Technol. 50, 9070–9077.
https://doi.org/10.1021/acs.est.6b00668
Lugtenberg, B.J., Kravchenko, L. V., Simons, M., 1999. Tomato seed and root exudate sugars: composition, utilization by Pseudomonas biocontrol strains and role in rhizosphere colonization. Environ. Microbiol. 1, 439–446. https://doi.org/10.1046/j.1462-2920.1999.00054.x
Ma, L.Q., Komar, K.M., Tu, C., Zhang, W., Cai, Y., Kennelley, E.D., 2001. A fern that hyperaccumulates arsenic. Nature 409, 579. https://doi.org/10.1038/35054664
Machuca, A., Milagres, A.M.F., 2003. Use of CAS‐agar plate modified to study the effect of different variables on the siderophore production by Aspergillus. Lett. Appl. Microbiol. 36, 177–181. Mesa, V., Navazas, A., González-Gil, R., González, A., Weyens, N., Lauga, B., Gallego, J.L.R., Sánchez,
J., Peláez, A.I., 2017. Use of endophytic and rhizosphere bacteria to improve phytoremediation of arsenic-contaminated industrial soils by autochthonous Betula celtiberica. Appl. Environ. Microbiol. 83, e03411-16.
reactor by using Ralstonia eutropha MTCC 2487 and granular activated carbon. Journal of hazardous materials. 153, 588-599.
Monsieurs, P., Moors, H., Van Houdt, R., Janssen, P.J., Janssen, A., Coninx, I., Mergeay, M., Leys, N., 2011. Heavy metal resistance in Cupriavidus metallidurans CH34 is governed by an intricate transcriptional network. Biometals 24, 1133–1151.
Nyambura Ngamau, C., 2012. Isolation and identification of endophytic bacteria of bananas (Musa spp.) in Kenya and their potential as biofertilizers for sustainable banana production. African J. Microbiol. Res. 6, 6414–6422. https://doi.org/10.5897/ajmr12.1170
Oremland, R.S., Stolz, J.F., 2003. The ecology of arsenic. Science (80). 300, 939–944.
Paez-Espino, D., Tamames, J., de Lorenzo, V., Canovas, D., 2009. Microbial responses to environmental arsenic. Biometals 22, 117–130. https://doi.org/10.1007/s10534-008-9195-y
Patten, C.L., Glick, B.R., 2002. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl. Environ. Microbiol. 68, 3795–3801. https://doi.org/10.1128/aem.68.8.3795-3801.2002
Pii, Y., Borruso, L., Brusetti, L., Crecchio, C., Cesco, S., Mimmo, T., 2016. The interaction between iron nutrition, plant species and soil type shapes the rhizosphere microbiome. Plant Physiol. Biochem. 99, 39–48. https://doi.org/10.1016/J.PLAPHY.2015.12.002
Rajbanshi, A., 2008. Study on heavy metal resistant bacteria in Guheswori sewage treatment plant. Our Nature. 6, 52-57.
Rajkumar, M., Sandhya, S., Prasad, M.N. V, Freitas, H., 2012. Perspectives of plant-associated microbes
in heavy metal phytoremediation. Biotechnol. Adv. 30, 1562–1574.
https://doi.org/https://doi.org/10.1016/j.biotechadv.2012.04.011
Schlemper, T.R., Leite, M.F.A., Lucheta, A.R., Shimels, M., Bouwmeester, H.J., van Veen, J.A., Kuramae, E.E., 2017. Rhizobacterial community structure differences among sorghum cultivars in different growth stages and soils. FEMS Microbiol. Ecol. 93. https://doi.org/10.1093/femsec/fix096
Schwyn, B., Neilands, J.B., 1987. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 160, 47–56. https://doi.org/https://doi.org/10.1016/0003-2697(87)90612-9
Stolz, J., Basu, P., & Oremland, R., 2002. Microbial transformation of elements: the case of arsenic and selenium. International Microbiology. 5, 201-207.
Suhadolnik, M.L.S., Salgado, A.P.C., Scholte, L.L.S., Bleicher, L., Costa, P.S., Reis, M.P., Dias, M.F., Ávila, M.P., Barbosa, F.A.R., Chartone-Souza, E., Nascimento, A.M.A., 2017. Novel arsenic-transforming bacteria and the diversity of their arsenic-related genes and enzymes arising from arsenic-polluted freshwater sediment. Sci. Rep. 7, 11231. https://doi.org/10.1038/s41598-017-11548-8
Trabelsi, D., Mhamdi, R., 2013. Microbial inoculants and their impact on soil microbial communities: a review. Biomed Res. Int. 2013.
Tu, C., Ma, L. Q., & Bondada, B., 2002. Arsenic accumulation in the hyperaccumulator Chinese brake and its utilization potential for phytoremediation. Journal of environmental quality. 31, 1671-1675. von Rozycki, T., Nies, D. H., 2009. Cupriavidus metallidurans: evolution of a metal-resistant bacterium.
Antonie Van Leeuwenhoek. 96, 115.
influenced by pH, calcium, and temperature. Int. J. Phytoremediation 12, 85–95.
https://doi.org/10.1080/15226510902767148
Wang, X., Rathinasabapathi, B., Oliveira, L.M. de, Guilherme, L.R.G., Ma, L.Q., 2012. Bacteria-mediated arsenic oxidation and reduction in the growth media of arsenic hyperaccumulator Pteris vittata. Environ. Sci. Technol. 46, 11259–11266. https://doi.org/10.1021/es300454b
Xu, J.-Y., Han, Y.-H., Chen, Y., Zhu, L.-J., Ma, L.Q., 2016. Arsenic transformation and plant growth promotion characteristics of As-resistant endophytic bacteria from As-hyperaccumulator Pteris vittata. Chemosphere 144, 1233–1240. https://doi.org/10.1016/J.CHEMOSPHERE.2015.09.102 Weisburg, W.G., Barns, S.M., Pelletier, D.A., Lane, D.J., 1991. 16S ribosomal DNA amplification for
phylogenetic study. J. Bacteriol. 173, 697 LP – 703.
Yazdi, S., Ardekani, A.M., 2012. Bacterial aggregation and biofilm formation in a vortical flow. Biomicrofluidics 6. https://doi.org/10.1063/1.4771407
Yu, X.Z., Trapp, S., Zhou, P.H., Chen, L., 2007. Effect of temperature on the uptake and metabolism of
cyanide by weeping willows. Int. J. Phytoremediation 9, 243–255.
https://doi.org/10.1080/15226510701376141
Zhao, F.J., McGrath, S.P., Meharg, A.A., 2010. Arsenic as a food chain contaminant: mechanisms of plant uptake and metabolism and mitigation strategies. Annual Review of Plant Biology, Vol 61. Annual Reviews, Palo Alto, pp. 535–559. https://doi.org/10.1146/annurev-arplant-042809-112152
Zhu, L.-J., Guan, D.-X., Luo, J., Rathinasabapathi, B., Ma, L.Q., 2014. Characterization of arsenic-resistant endophytic bacteria from hyperaccumulators Pteris vittata and Pteris multifida. Chemosphere 113, 9– 16. https://doi.org/10.1016/J.CHEMOSPHERE.2014.03.081