1
題 目
ERK
( extracellular signal-regulated kinase)標 的 遺伝子欠損マウスを用いた、麻酔薬の作用機序 における ERK 経路の役割に関する研究
あ ら き よ し ゆ き
荒 木 義 之
(麻酔・蘇生医学専攻)
防 衛 医 科 大 学 校
平成25年度
2 目 次
第1章 緒言 1 頁
第2章 ERK1とERK2の全身麻酔薬に対する影響の検討 3 頁
第1節 背景及び目的 3 頁
第2節 実験材料と方法 4 頁
1、実験動物 4 頁
1)Erk1標的遺伝子欠損マウス
2)Erk2条件的標的遺伝子欠損マウス
3)コントロールマウス
2、使用薬剤 5 頁
1)プロポフォール
2)ペントバルビタール
3)ケタミン
4)セボフルラン
3、実験方法 6 頁
正向反射消失時間 最少肺胞内濃度
4、統計学的解析 7 頁
第3節 結果 8 頁
第4節 小括 10 頁
第3章 MEK阻害薬による解析 11 頁
第1節 背景及び目的 11 頁
第2節 実験材料と方法 11 頁
1、実験動物 12 頁
2、使用薬剤 12 頁
MEK阻害薬
3、実験方法 12 頁
第3節 結果 12 頁
第4節 小括 13 頁
3
第4章 Erk2 CKOマウスの脳・脊髄GABA濃度及び脳スライス切片漏出GABA濃度の測定 14 頁
第1節 背景及び目的 14 頁
第2節 実験材料と方法 15 頁
1、実験動物 15 頁
2、実験方法 15 頁
1)脳及び脊髄GABA濃度の測定
2)脳スライス切片漏出GABA濃度の測定
第3節 結果 16 頁
第4節 小括 16 頁
第5章 考察と結論 17 頁
謝辞 23 頁
引用文献 24 頁
図表 32 頁
4 第 1 章 緒言
全身麻酔は意識・記憶・痛覚を抑制するがその構造薬理作用ともに多種多様である。現在臨床
的に広く用いられている全身麻酔薬は静脈麻酔薬と吸入麻酔薬に分類される。麻酔薬は多くのチャ
ネルやレセプタに作用してその効果を発揮すると考えられており、麻酔薬の主な分子標的として、
細胞膜上に存在するγ-アミノ酪酸(γ-aminobutyric acid:GABA)A受容体、N-メチル-D-アスパラ
ギン酸(N-methyl-D-aspartic acid:NMDA)受容体が示唆されている1-5)。これまでの研究で、NMDA
受容体機能に対する ERK の直接的な作用の報告はないものの、GABA 受容体機能では細胞外シグナル
調節キナーゼ(extracellular signal- regulated kinase: ERK)経路がネガディブモジュレーター
として機能しており6)、ERK が直接 GABAA受容体α1 サブユニットの T375 をリン酸化して GABAA受容
体を抑制しているとの報告があり7)、麻酔薬の作用は ERK と関連している可能性が考えられた。 し
かしながら、麻酔薬の結合した受容体がさらにどのようなメカニズムで細胞応答をもたらすのかに
ついては未解明な点が多く、本研究では、神経細胞において重要な役割を示唆されている、分裂促
進因子活性化タンパク質キナーゼ(mitogen-activated protein kinase: MAPK)ファミリーに属す
る ERK に着目し、麻酔作用における ERK 経路の役割を解析することを目的とした。
ERK は、細胞膜の増殖因子受容体にリガンド(増殖因子)が結合し 2 量体化すると、アダプター
分子、低分子量 G 蛋白質 Ras を経由して、Raf→MEK→ERK とリン酸化反応する MAPK 経路(MAPK カス
ケード)に位置し、活性化した ERK は様々なシグナル伝達や細胞増殖や分化の遺伝子発現に関与し
ている8)。これまでの ERK 研究では主に MEK 阻害薬を用いた方法がとられていたが、MEK 阻害薬は ERK のアイソフォームである ERK1 と ERK2 の両方の活性化を阻害するため、各個の役割については
評価できなかった。今回、麻酔作用における ERK の役割を生体において直接的に解明するため、ERK1
5
又は ERK2 が欠損したマウス(ノックアウトマウス)を用いて解析することとした。
第 2 章では、各ノックアウトマウスに全身麻酔薬を投与して、麻酔作用に対する ERK1 と ERK2 の役
割の違いをそれぞれ検討した。続いて第 3 章では MEK 阻害薬で ERK の活性を薬理学的に阻害した場合
の麻酔作用に対する検討、第 4 章では脳および脊髄組織や脳スライス切片を用いた GABA 濃度に関し
て解析を行った。最後に第1章から第 4 章までの内容をまとめて第 5 章で考察した。
本研究は、防衛医科大学校学内動物実験倫理委員会の承認を受けて行った。(倫理委員会受付番号
12082)
6 第 2 章 ERK1 と ERK2 の全身麻酔薬に対する影響の検討
第 1 節 背景及び目的
MEK 阻害剤を用いた研究から ERK1/2 は記憶や神経可塑性に関与し9)、さらには海馬 LTP では ERK2
の方が ERK1 よりもより顕著に活性化される10)ことから、ERK1 と ERK2 はその機能に違いが存在する
ことが明らかとなった。しかし、MEK 阻害薬を用いた手法では ERK1 と ERK2 の 2 つのアイソフォーム
の違いを解析することはできなかった。昨今、ERK1 あるいは ERK2 のいずれか一方のみを欠損したマ
ウスが作出され、Erk1 KO マウスは正常に発育するのに対して、Erk2 KO マウスは胎生致死であるこ
とから発生の初期段階における ERK1 と ERK2 の役割の違いが明らかにされた。さらに、Erk1 KO マウ
スは脳を含めて胸腺以外に形態的な異常は観察されず 11)、海馬 LTP や海馬依存性記憶についての異
常はないと報告12)されていることから、海馬において ERK1 は重要でないか ERK2 により補償される
可能性が考えられている。一方で、Erk1 KO マウスでは側座核において新皮質−側座核間の神経伝達
に著名な増強が観察されることから、野生型マウスでは ERK1 が神経伝達を抑制している可能性が報
告13)されている。また、Erk2 KO マウスの胎生致死を回避するために作成されたErk2 KD マウスは野
生型マウスの 60〜80%の ERK2 を有しており、形態や情動・短期記憶などの異常は観察されなかった
が、長期記憶の障害を示し14)、 ERK2 の減少が長期記憶に影響を及ぼすことを示唆された。さらに、
条件的Erk2 KO マウスを用いた研究から、ERK2 は ERK1 で補償できない生理機能を有することが明ら
かにされている14-17)。このように、遺伝子改変マウスを用いて ERK1 と ERK2 の生体内での生理機能
の差異がより詳細に明らかになってきており、麻酔薬の作用においても遺伝子改変動物を利用するこ
7
とで、ERK1 と ERK2 の生体内での生理機能の差異について明らかにすることが重要であると考えた。
全身麻酔薬の多くは興奮性受容体である NMDA 受容体に対する拮抗作用、または抑制性受容体であ
る GABAA受容体に対する機能亢進作用が主な作用機序とされている。臨床的に広く用いられている全
身麻酔薬のなかで、静脈麻酔薬のケタミン(NMDA 受容体拮抗作用)とペントバルビタール、プロポ
フォール(GABAA受容体機能亢進作用)および吸入麻酔薬のセボフルラン(GABAA受容体機能亢進作用)
を用いて、麻酔作用に対する ERK1 および ERK2 の関与について遺伝子改変マウスを用いて解析した。
第 2 節 実験材料と方法
1、実験動物
1)Erk1標的遺伝子欠損マウス:Erk1 KO マウス
全身でERK1が欠損しているマウスは共同研究者(Gilles Pagès、Jacques Pouysségur)から譲り
うけた18)。
2)Erk2条件的標的遺伝子欠損マウス:Erk2 CKO マウス
通常の方法により作出したErk2標的遺伝子欠損マウスは胎生致死であるため19)、ERK2 の欠損部
位を限局することによって致死性を回避した、条件的遺伝子欠損マウス(コンディショナルノック
アウトマウス)を用いた。中枢神経特異的Erk2標的遺伝子欠損マウスは、ERK2 を loxP 配列で挟み
込んだErk2 flox マウス(ERK2 のエクソン 2 とエクソン 3 の部位に、loxP 配列で挟み込んだマウ
ス)と、中枢神経特異的に Cre を発現するようにしたマウスを掛け合わせることで共同研究者(Satoh
8
Yasushi)が作出したものである(図 2A、図 2B)19)。Erk2 (flox/+)マウスは、バッククロス(戻
し交配)を 10 世代以上行ったものを用いている。
3)コントロールマウス:
コントロールマウスとして、同腹のその他のマウス(Erk2 (flox/+), Cre-、Erk2 (+/+), Cre-
、Erk2 (flox/flox), Cre-、Erk2 (+/+), Cre+)を使用した(図2B)。
4)本研究では8から10週齢のマウスを用いた。
2、使用薬剤
1)プロポフォール
1%ディプリバン注(アストラゼネカ株式会社、大阪)をマウスの腹腔内へ 100 mg/kg を投与した。
2)ペントバルビタール
ペントバルビタールナトリウム(大日本製薬株式会社、大阪)を生理食塩水で希釈して、5 mg/ml
溶液を作成し、マウスの腹腔内へ50 mg/kgを投与した。
3)ケタミン
ケタラール静注用 200 ㎎(第一三共株式会社、東京)をマウスの腹腔内へ 200 mg/kg を投与した。
4)セボフルラン
セボフレン吸入麻酔薬 (丸石製薬株式会社、大阪)をバポール気化器(ドレーゲルメディカルジャ
パン、東京)で調整して投与した。
9 3、実験方法
静脈麻酔薬の作用は、薬剤腹腔内投与後の正向反射消失 (Loss Of Righting Reflex: LORR)到達時
間(Latency to LORR)と LORR 持続時間(Duration of LORR)を指標とした。正向反射は頭部を重力
の方向に対して正常の位置に回復するようにする反射運動であり、背臥位にしたマウスが 10 秒以内
に腹側位に回復することが反射陽性と定義されている20)。LORR 到達時間は、薬剤腹腔内投与時点か
ら背臥位にしたマウスが 10 秒間連続して腹側位に回復しなかった時点(LORR 到達時点)までの時間
とした。また、LORR 持続時間は、LORR 到達時点からマウスが腹側位に回復した時点までの時間とし
た。
吸入麻酔薬の麻酔作用の指標としては、LORR 時点の吸入麻酔薬濃度のほかに最小肺胞内濃度
(Minimum Alveolar Concentration: MAC,最大刺激に対して動物の 50%を不動化させるのに必要な
肺胞吸入麻酔薬濃度)も測定した。吸入麻酔薬を使用した実験では、適度に加湿・加温(室内温度
38℃ ± 1℃)したグローブボックスに 2 l/min の 25%酸素ガスをキャリーガスとしてセボフルラ
ンを供給した。酸素濃度と吸入麻酔薬濃度はカプノマックウルティマガス分析装置(デタックスオメ
ダ、東京)を用いて調整した。グローブボックス内の吸入麻酔薬濃度が定常状態となった時点でマウ
スを投入し、吸入麻酔薬濃度を変化させる場合は 0.1%ずつ変化させ、30 分間持続させて肺胞気と
中枢神経間の濃度平衡状態を確立した。MAC 測定法は Eger らの方法21)に従い、濃度平衡状態におい
てアリゲータークリップを用いたテイルクランプ法(最大刺激としてマウスの尾近位側 1 cm をアリ
ゲータークリップで 10 秒間クランプしたときの体動の有無を観察し、調査群の 50%で最大刺激時の
体動が消失した濃度を MAC として記録する方法)にて測定した。
10
1)Erk1 KO マウスにおける静脈麻酔薬投与後の LORR 到達時間と持続時間
①Erk1 KO マウス群(n = 15)とコントロールマウス群(n = 16)に、ペントバルビタール(50 mg/kg)
を腹腔内投与し、LORR 到達時間と持続時間を測定した。②Erk1 KO マウス群(n = 15)とコントロー
ルマウス群(n = 15)に、プロポフォール(100 mg/kg) を腹腔内投与し、LORR 到達時間と持続時間
を測定した。③Erk1 KO マウス群(n = 9)とコントロールマウス群(n = 10)に、ケタミン (200 mg/kg)
を腹腔内投与し、LORR 到達時間と持続時間を測定した。
2)Erk2 CKO マウスにおける静脈麻酔薬投与後の LORR 到達時間と持続時間
①Erk2 CKO マウス群(n = 17)とコントロールマウス群(n = 19)に、ペントバルビタール(50 mg/kg)
を腹腔内投与し、LORR 到達時間と持続時間を測定した。②Erk2 CKO マウス群(n = 14)とコントロー
ルマウス群(n = 16)に、プロポフォール(100 mg/kg)を腹腔内投与し、LORR 到達時間と持続時間を
測定した。③Erk2 CKO マウス群(n = 23)とコントロールマウス群(n = 22)に、ケタミン (200 mg/kg)
を腹腔内投与し、LORR 到達時間と持続時間を測定した。
3)Erk1 KO マウスにおけるセボフルラン投与時の LORR 濃度及び MAC
Erk1 KO マウス群(n = 14)とコントロールマウス群(n = 15)にセボフルラン麻酔を行い、LORR
濃度と MAC を測定した。
4)Erk2 CKO マウスにおけるセボフルラン投与時の LORR 濃度及び MAC
Erk2 CKO マウス群(n = 20)とコントロールマウス群(n = 20)にセボフルラン麻酔を行い、LORR
濃度と MAC を測定した。
4、統計学的解析
11
Student's t-testを用いて検定し、平均値 ± 標準誤差で表した。
第 3 節 結果
1、Erk1 KO マウスにおける静脈麻酔薬投与後の LORR 到達時間と持続時間 (図 3)
1)ペントバルビタール (50 mg/kg) の腹腔内投与による LORR 到達時間と持続時間はコントロールマ
ウスと変化なかった。(Latency to LORR: Erk1 KO マウス群、230.0 ± 28.3 秒、n = 15 対 コン
トロールマウス群、217.2 ± 24.6 秒、n = 16; t = 0.34, P = 0.73)(Duration of LORR: Erk1 KO
マウス群、59.92 ± 5.88 分、n = 15 対 コントロールマウス群、59.75 ± 3.20 分、n = 16; t =
1.02, P = 0.32)。
2)プロポフォール (100 mg/kg)の腹腔内投与による LORR 到達時間と持続時間はコントロールマウス
と変化なかった。(Latency to LORR: Erk1 KO マウス群、251.0 ± 28.1 秒、n = 15 対 コントロー
ルマウス群、239.0 ± 21.5 秒、n = 15; t = 0.34, P = 0.74)(Duration of LORR: Erk1 KO マウ
ス群、22.17 ± 3.76 分、n = 15 対 コントロールマウス群、20.07 ± 3.70 分、n = 15; t = 0.40,
P = 0.69)。
3)ケタミン (200 mg/kg)の腹腔内投与による LORR 到達時間と持続時間はコントロールマウスと変化
なかった。(Latency to LORR: Erk1 KO マウス群、110.6 ± 3.8 秒、n = 9 対 コントロールマウ
ス群、128.5 ± 8.2 秒、n = 10; t = 1.99, P = 0.07)(Duration of LORR: Erk1 KO マウス群、77.97
± 5.72 分、n = 8 対 コントロールマウス群、68.63 ± 4.39 分、n = 9; t = 1.31, P = 0.21)。
12
2、Erk2 CKO マウスにおける静脈麻酔薬投与後の LORR 到達時間と持続時間(図 4)
1)ペントバルビタール (50 mg/kg) の腹腔内投与による LORR 到達時間はコントロール群と比較して
変化しなかったが、持続時間は延長した。(Latency to LORR: Erk2 CKO マウス群、223.5 ± 26.09 秒、
n = 17 対 コントロールマウス群、219.7 ± 24.03 秒、n = 19; t = 0.11, P = 0.92)(Duration
of LORR: Erk2 CKO マウス群、75.31 ± 4.00 分、n = 17 対 コントロールマウス群、58.66 ± 3.84
分、n = 19; t = 3.00, P<0.01)。
2)プロポフォール (100 mg/kg)の腹腔内投与による LORR 到達時間はコントロール群と比較して変化
しなかったが、持続時間は延長した。(Latency to LORR: Erk2 CKO マウス群、202.9 ± 43.98 秒、
n = 14 対 コントロールマウス群、195.9 ± 10.1 秒、n = 16; t = 0.15, P = 0.88)(Duration of
LORR: Erk2 CKO マウス群、40.46 ± 5.39 分、n = 14 対 コントロールマウス群、24.44 ± 3.93
分、n = 16; t = 2.44, P<0.05)。
3)ケタミン (200 mg/kg)の腹腔内投与による LORR 到達時間はコントロール群と比較して変化しなか
ったが、持続時間は延長した。(Latency to LORR: Erk2 CKO マウス群、109.6 ± 11.3 秒、n = 23
対 コントロールマウス群、109.6 ± 11.3 秒 n = 22; t = 1.85, P = 0.07)(Duration of LORR: Erk2
CKO マウス群、74.93 ± 3.08 分、n = 23 対 コントロールマウス群、63.57 ± 2.65 分、n = 22;
t = 2.79, P<0.01)。
3、Erk1 KO マウスにおけるセボフルラン投与時の LORR 濃度及び MAC(図 5)
13
Erk1 KO マウスでは、セボフルラン投与による LORR 濃度と MAC ともにコントロールマウスと比較
して有意差を認めなかった。(LORR: Erk1 KO マウス群、EC50 = 1.342%、n = 14 対 コントロール
マウス群、EC50 = 1.367%、n = 15; P = 0.49)(MAC: Erk1 KO マウス群、EC50 = 2.747%、n = 14
対 コントロールマウス群、EC50 = 2.779%、 n = 15; P = 0.45)。
4、Erk2 CKO マウスにおけるセボフルラン投与時の LORR 濃度及び MAC(図 6)
Erk2 CKO マウスでは、セボフルラン投与による LORR 濃度と MAC ともにコントロールマウスと比較
して有意差を認めなかった。(LORR:Erk2 CKO マウス群、EC50 = 1.322%、n = 20 対 コントロール
マウス群、EC50 = 1.367%、n = 20; P = 0.11)(MAC: Erk2 CKO マウス群、EC50 = 2.724%、n = 20
対 コントロールマウス群、EC50 = 2.782%、n = 20; P = 0.07)。
第 4 節 小括
Erk1 KO マウスではプロポフォール、 ペントバルビタール、 ケタミンの腹腔内投与による LORR
到達時間と持続時間はコントロールマウスと比較して差を認めなかった。Erk2 CKO マウスではプロ
ポフォール、 ペントバルビタール、 ケタミンの腹腔内投与により、LORR 到達時間には差を認めな
かったが、LORR 持続時間の延長を認めた。セボフルラン麻酔では、Erk1 KO マウス、Erk2 CKO マウ
スともに LORR 濃度と MAC に差を認めなかった。
ERK1 と ERK2 のアミノ酸配列は約 83%が同じであるにもかかわらず、静脈麻酔薬の作用には ERK2
が重要な役割を担っていることが示唆された。一方、吸入麻酔薬であるセボフルランの作用には ERK1
と ERK2 ともに関与していない可能性が示唆された。
14 第 3 章 MEK 阻害薬による解析
第1節 背景及び目的
ERK2 は静脈麻酔薬のケタミン(NMDA 受容体拮抗作用)とペントバルビタール、プロポフォール
(GABAA受容体機能亢進作用)の麻酔持続時間に影響をえることが示唆された。さらに詳細に解明す
るため、昨今頻用される静脈麻酔薬のプロポフォールと ERK2 の関係について研究を進めた。
過去に NMDA 受容体ε1 サブユニットノックアウトマウスを用いてケタミンの麻酔作用を調べた研
究では当初、ε1 ノックアウトマウスはコントロールマウスに比べてケタミンに対して抵抗性を示し
たと報告された 22)。これと時を同じくして、ノックアウトマウスを用いた実験結果の解釈には慎重
性を求める考え方も示され 23, 24)、後にε1 サブユニットノックアウトマウスのケタミンに対しての
抵抗性は、必ずしも麻酔薬の標的がノックアウトされていることによる直接的な影響とは解釈できな
いことが明らかとなった。現在では、このノックアウトマウスは NMDA 受容体の機能低下に加えて、
ドパミン、セロトニン、GABA などによるシナプス伝達の機能変化が認められ、その結果として運動
性の充進などを呈することが明らかとなり25)、NMDA 受容体以外の機能変化が二次的に麻酔薬感受性
の違いに関与していると考えられている。特定の遺伝子欠損マウスで認められた麻酔作用の変化が、
特定の遺伝子欠損に由来しない可能性が指摘されているため22-24)、MEK 阻害薬(SL-327)で ERK の活性
を薬理学的に阻害したマウスを用いて、プロポフォールの麻酔作用について実験を行った。
第 2 節 実験材料と方法
15 1、実験動物
コントロールマウス
2、使用薬剤
MEK 阻害薬(SL-327, ENZO Life Sciences 社, Farmingdale, NY, USA)はα
-[amino[(4-aminophenyl)thio]methylene]-2-(trifluoromethyl)benzeneacetonitrile を、溶媒であ
るジ メ チ ル ス ル ホ キ シ ド ( DMSO)で溶解し、マウスの腹腔内に 50 mg/kg 投与した。
3、実験方法
コントロールマウスに、MEK 阻害薬 50 ㎎/㎏を腹腔内投与し、その 30 分後にプロポフォール (100
mg/kg)を腹腔内投与した群(n = 17)と、Vehicle 100 μl を腹腔内投与し、その 30 分後にプロポ
フォール (100 mg/kg)を腹腔内投与した群(n = 15)の 2 群に分けて LORR 到達時間と持続時間を測
定した。
4、統計学的解析
Student's t-test を用いて検定し、平均値 ± 標準誤差で表した。
第 3 節 結果
ERK 活性化阻害薬である MEK 阻害薬(SL-327)を用いた群では、Vehicle 群と比較してプロポフォ
ール (100 mg/kg)の腹腔内投与後の LORR 到達時間には差を認めなかったが、LORR 持続時間は延長し
16
た。(Latency to LORR: MEK 阻害薬群、271.2 ± 11.85 秒、n = 17 対 Vehicle 群、289.2 ± 14.50
秒、n = 15; t = 0.97, P= 0.34)(Duration of LORR: MEK 阻害薬群、48.32 ± 2.83 分、n = 17 対
Vehicle 群、34.49 ± 2.51 分、n = 15; t = 3.61, P<0.01)(図 7)。
第 4 節 小括
プロポフォール腹腔内投与後の LORR 到達時間は MEK 阻害薬投与群と Vehicle 群で差を認めなかっ
た。一方、プロポフォール腹腔内投与後の LORR 持続時間は、MEK 阻害薬投与により ERK を阻害した
ところ Vehicle 群と比較して延長を認め、第 2 章のErk2 CKO マウス群で認められたプロポフォール
腹腔内投与後の LORR 持続時間の延長と同様の結果であった。遺伝子工学的および薬理学的実験で同
様な結果が得られたことから、プロポフォール投与による LORR 持続時間の延長は、ERK2 の作用に
よる可能性が示唆された。
17
第 4 章 Erk2 CKO マウスの脳・脊髄組織 GABA 濃度及び脳スライス切片漏出 GABA 濃度の測定
第 1 節 背景及び目的
近年、麻酔薬は神経細胞表面の受容体蛋白に作用することが明らかになってきている。中枢神経系
におけるニューロン間の神経伝達は、シナプス前ニューロンから放出される神経伝達物質が、シナプ
ス後ニューロンの受容体を活性化し、細胞膜に活動電位を発生させることにより行われる。
神経伝達物質である GABA はグルタミン酸脱炭酸酵素(glutamic acid decarboxylase:GAD)によ
り触媒される反応により、L-グルタミン酸から合成され、小胞膜 GABA トランスポーター(VGAT)に
よりシナプス小胞に貯蓄されている26)。GABA はシナプス後膜 GABA 受容体と神経細胞体樹状突起や軸
索上に広範囲に分布するシナプス外 GABA 受容体に作用しており27)、GABAA受容体を介した 2 種類の
抑制性作用を呈する。それらは、シナプス後膜受容体での二次的な抑制作用(Phasic inhibition)
とシナプス外 GABA 受容体での持続的な抑制作用(Tonic inhibition)であり、ともに麻酔薬の作用
メカニズムに大きく関与している可能性が示唆されている 28)。Phasic inhibition には、比較的高
濃度の GABA が必要とされる29)が、Tonic inhibition はシナプス外に漏出してきた低濃度の GABA に
よる、GABA 親和性が高い GABA 受容体の活性化によると考えられている30)。以上から、脳 GABA 濃度
や VGAT 漏出 GABA 濃度は麻酔作用に大きく影響すると考えられ本実験でもその測定を行った。
また、脳だけでなく脊髄における全身麻酔薬の作用も以前から指摘されており 31, 32)、マウスの脊
髄後角膠様(substantia gelatinosa: SG)細胞の一部にも GABAA受容体介在性の Tonic inhibition
が存在し、プロポフォールによって SG 細胞の Tonic inhibition が増強さることから、脊髄レベルに
おいても Tonic inhibition が麻酔薬の作用メカニズムに重要であることが示唆されている33)。その
18
ため、本実験でも脊髄における GABA 濃度の測定も行った。
第 2 節 実験材料と方法
1、実験動物
Erk2 CKO マウス及びコントロールマウス
2、実験方法
1)Erk2 CKO マウス 6 匹及びコントロールマウス 9 匹から脳および脊髄をとりだし重さを測定した。
脳には 2.5 ml、脊髄には 0.6 ml の 0.2%プロテアーゼインヒビター入りリン酸緩衝生理食塩水を加
えて、ポリトロン(Kinematica, Littau-Lucerne, Switzerland)にてホモジナイズ(24,000 rpm、
15 秒、3 回)した。組織片を除去したのち、遠心(3000 rpm、20 分、4℃)し上澄みを 500 μl 回
収した。株式会社エスアールエル(東京)に依頼して高速液体クロマトグラフィー(High performance
liquid chromatography: HPLC)法により GABA 濃度を測定し、組織重量あたりの GABA の量を算出し
た。
2)Erk2 CKO マウス 4 匹及びコントロールマウス 7 匹から脳スライス切片(大脳皮質部位)を作成し、
人工脳脊髄液(Artificial cerebrospinal fluid:ACSF)中で 60 分間安定化した。その後、切片を
別の ACSF1.5 ml に移動し 60 分間培養したのち、ACSF を 120 μl を回収し HPLC 法により GABA 濃度
を測定した。回収した脳スライス切片のタンパク量を Branford 法により計測し、単位タンパク当た
19
りの漏出 GABA の量を算出した。HPLC 法の測定は 2 回行い、2 回の平均値を用いた。
3、統計学的解析
Student's t-test を用いて検定し、平均値 ± 標準誤差で表した。
第 3 節 結果
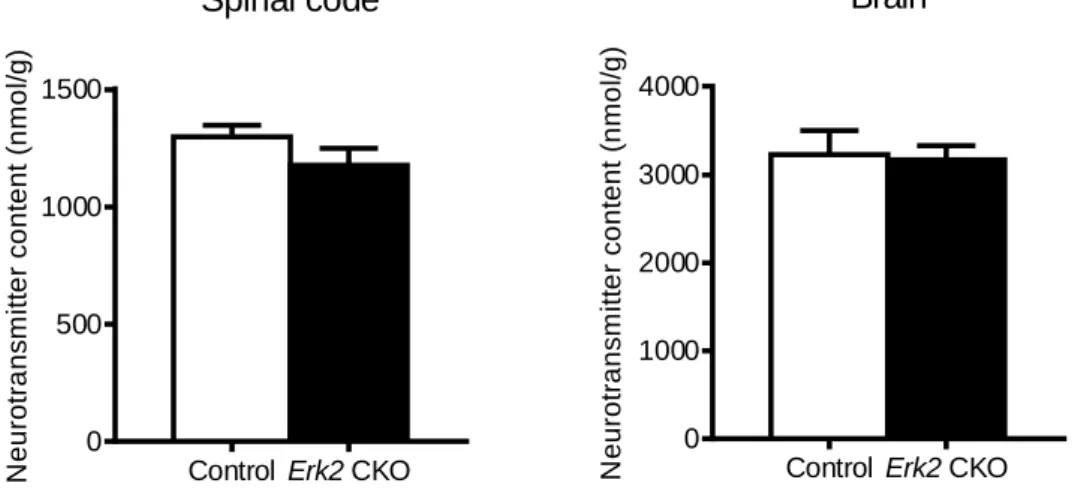
1、Erk2 CKO マウス及びコントロールマウスの脳(Brain)および脊髄(Spinal code)で組織重量当た
りの GABA 濃度に差は認めなかった。(Brain: Erk2 CKO マウス群、3173 ± 158.3 nmol/g、n = 6 対
コントロールマウス群、3230 ± 267.7 nmol/g、n = 9; t = 0.16, P = 0.88)(Spinal code: Erk2 CKO
マウス群、1179 ± 70.6 nmol/g、n = 6 対 コントロールマウス群、1298 ± 50.3 nmol/g、n = 9;
t = 1.42, P = 0.19)(図 8)。
2、Erk2 CKO マウス及びコントロールマウスの脳スライス切片から漏出された単位タンパク当たりの
GABA 濃度に差を認めなかった(Brain Slice: Erk2 CKO マウス群、112.3 ± 50.0 nmol/g、n = 4 対
コントロールマウス群、111.1 ± 23.0 nmol/g、n = 7; t = 0.02, P = 0.98)(図 9)。
第 4 節 小括
Erk2 CKO マウス及びコントロールマウスの脳と脊髄組織において、神経伝達物質 GABA の濃度に差
を認めず、脳スライス切片漏出 GABA 量にも差を認めなかった。プロポフォール投与によりErk2 CKO
マウスで認めた LORR 持続時間の延長は、神経伝達物質 GABA の量によるものでないことが示唆された。
20 第 5 章 考察と結論
MAPK ファミリーに属する ERK は、転写因子を含む標的をリン酸化し、細胞増殖などの現象を制御
する因子であり、MEK から解離して核内移行すると考えられている。阻害剤や中枢神経特異的 ERK2
ノックアウトマウスを用いた実験から、ERK が記憶、学習メカニズムに重要であることが報告されて
いる34, 35)。しかしながら、ERK1/2 と麻酔作用の関係はいまだ報告されておらず、本研究によりその
関係の一端が初めて明らかとなった。
薬剤の腹腔内投与により投与された薬剤は腹膜から速やかに吸収されて全身循環に入る。血漿中濃
度は短時間で最高値を示し、時間経過とともに徐々に低下していく。LORR 到達時間は薬物動態学・
薬力学的に効果部位濃度が入眠に必要な濃度に上昇するまでの時間であり、LORR 持続時間は薬物動
態学・薬力学的に効果部位濃度が入眠に必要な濃度以上に保持される時間である。
本研究では、静脈麻酔薬の LORR 持続時間においてErk1 KO マウスはコントロールマウスと差異を
認めなかったのに対し、Erk2 CKO マウスはコントロールマウスに対して延長を認めた。Erk1 KO マウ
スとErk2 CKO マウスで異なる結果が出た理由として、麻酔作用に ERK1 が関与していない可能性と、
麻酔作用では ERK2 が ERK1 で補償できない生理機能を有する可能性が考えられたが、いずれも麻酔作
用における ERK2 の重要性を示すと考えられる。
また、薬物動態学・薬力学の分野では、臨床現場で頻用されるプロポフォールの研究が盛んに行わ
れており、意識消失と意識回復時の効果部位濃度にはよい相関が存在することが明らかとなっており
36)、このことは一般的には LORR 到達時間と LORR 持続時間にも相関が存在することを示唆している。
しかし、本研究ではErk2 CKO マウスで、コントロールマウスと比較して LORR 到達時間は差を認めな
21
かったが、LORR 持続時間では差を認めるという特異的な結果を得た。Erk2 CKO マウスでは中枢神経
特異的に ERK2 が欠損しており、コントロールマウスと比較して薬物動態的な差異は認めないと考え
られることから、 LORR 到達時間と LORR 持続時間の差の原因としては次のことが考えられる。一つ
は、薬剤投与後の血漿中濃度は短時間で上昇するため、短時間で LORR 到達時間となり差を検出でき
なかったが、代謝・排泄による血漿中濃度の減少は緩徐におこるため、LORR 持続時間の差として認
めた可能性である。別の可能性としては、LORR 到達時間以後にErk2 CKO マウスで薬剤感受性が亢進
ししたため、LORR 到達時間では差を認めず、LORR 持続時間では差を認めた可能性が考えられた。Erk2
CKO マウスで認めた静脈麻酔薬による LORR 持続時間の延長は、Erk2 CKO マウスでは中枢神経特異的
に ERK2 が欠損していることから、麻酔薬の神経系への直接的な作用による可能性が高く、中枢神経
の ERK2 が静脈麻酔薬の麻酔作用に重要な役割を果たしていると考えられる。
GABAA受容体作動薬であるプロポフォールなどの投与後の LORR 持続時間は、コントロール群と比較
してErk2 CKO マウス群で延長した。GABAA受容体と ERK との関係については本章の後半で考察する。
NMDA 受容体拮抗薬であるケタミンについても、投与後の LORR 持続時間はコントロール群と比較し
てErk2 CKO マウス群で延長した。記憶関連では NMDA 受容体と ERK 経路との直接のかかわりが報告さ
れているが37)、麻酔作用との関連での報告はない。ケタミンの作用機序としては、NMDA 受容体拮抗
作用の他、過分極活性化型陽イオン電流(Hyperpolarization-activated cationic currents: HCN
電流)に対する作用38, 39)、シナプス領域外 GABA 受容体への作用 40)等が考えられているが明らかで
ない点が多い。ケタミンの麻酔作用に対する ERK2 の関与については今回の結果で示唆されたが、ま
だ明らかでない点が多く今後の課題である。
一方、吸入麻酔薬であるセボフルランではErk1 KO マウスとErk2 CKO マウスともに LORR に至る
22
濃度と MAC で差を認めなかった。LORR 濃度は脳における麻酔作用の指標であり、セボフルランの脳
に対する作用において ERK1 と ERK2 の関与は少ないと考えられた。また、MAC は侵害刺激に対する
体動抑制に必要な脊髄における麻酔薬の濃度であり、Erk2 CKO マウスは脊髄や後根神経節において
も ERK2 の欠損が確認されている41)ことから、セボフルランの脊髄に対する作用においても ERK1 と
ERK2 の関与は少ないと考えられた。
GABAA受容体では抑制性の神経伝達物質である GABA が結合すると、受容体の構造が変化し陰イオン
である Cl-を細胞内に流入させるチャネルが開く。Cl-の流入により細胞内の陰イオン濃度が上昇する
と、細胞膜にマイナス電位が生じ、ニューロンは興奮性の活動電位を発生できなくなる。麻酔薬は
GABAA受容体の特定アミノ酸残基に結合してイオンチャネルの開口時間を引き延ばし、GABA が受容体
に結合した際の神経抑制効果を強めると考えられている42)。GABAA受容体を介した抑制性作用は 2 種
類存在し、Phasic inhibition はシナプス小胞に存在する GABA がシナプス間隙に放出され、シナプ
ス後膜に存在する受容体に結合することによって発生する一過性の Cl- 電流であり、シナプスに存在
する受容体は比較的高濃度の GABA により活性化されるため、Phasic inhibition が発生するときに
は 0.3 〜l mM 程度の高濃度の GABA が放出されると考えられている29)。一方、Tonic inhibition は
シナプス外に漏出してきた数 μM 程度の低濃度の GABA が、シナプス外に存在する GABAA受容体に高
い親和性をもって結合して発生する持続的な Cl- 電流である。Tonic inhibition は、Phasic
inhibition と比べて、麻酔薬への感受性が非常に高く、海馬においてプロポフォールは Phasic
inhibition と Tonic inhibition をともに増強するが、その比率は Tonic inhibition のほうが約 7
倍高いとされ 43)、麻酔薬の作用メカニズムに重要な役割を果たしていると考えられている。Phasic
inhibition と Tonic inhibition と ERK の関係は今後の研究課題である。
23
最近では、遺伝学的手法を用いてアミノ酸を点変異させた GABAA受容体を用いることにより、麻酔
薬のより詳細な作用部位と GABAA受容体の関係が検討されるようになった。GABAA受容体は4回膜貫
通型蛋白質サブユニットの複合体で、通常はヘテロ 5 量体で構成される。現在までにα1-6,β1-3,
γ1-3,δ,ε1-3,θ,πのサブユニットが同定されている44, 45)。サブユニットの発現は、脳の部
位によって異なり、受容体を構成するサブユニットの組み合わせによって薬物感受性が異なることも
わかっている46)。脳神経細胞では 2 つのαサブユニットと 2 つのβサブユニットと 1 つγサブユニ
ットで構成されるものが一般的で47)、特にα1、β2、γ2 が 2:2:1 の 5 量体で構成されているサブタ
イプ構成が GABA 受容体の 60%を占めていると考えられている48, 49)。
プロポフォールはβ1 サブユニットの膜貫通部位(Transmembrane: TM)のなかで TM2 および TM3 に
作用するものの、α2 サブユニットの変異には影響されない50)。また、ラット GABAA受容体β3 サブ
ユニットの TM2 および TM3 領域の変異は、プロポフォールによる GABA 誘発電流の増強を抑制し51)、
TM2 のみの変異でもプロポフォールによる LORR が強く抑制されることが報告されている52)。
ERK は核内転写因子のリン酸化による遺伝子の発現作用や細胞質タンパク等への作用を有してい
る。遺伝子の発現には時間を必要とするのに対して、電位依存性カリウムチャネルの制御、局所的な
タンパク質の合成やグルタミン酸受容体局所制御などが迅速な作用として報告されており 53, 54)、今
回 MEK 阻害薬を用いた実験で認められたプロポフォール投与後の LORR 持続時間の延長も、時間的観
点から遺伝子発現に依存しない ERK による直接的な作用機序の存在が示唆された。これまでに、ERK
の GABAA受容体に対する直接的な作用として、GABAA受容体サブユニットに存在する ERK リン酸化部
位への作用による GABAA受容体の抑制が報告 7)されており、麻酔作用との関連は今後の課題である。
また、神経系の分化に重要な働きをするとされているグリコーゲン合成酵素キナーゼ 3β(GSK3β)
24
と ERK が協調的にゲフィリンのシナプス後膜でのゲフェリンのクラスタリングを制御しているとも
報告されている55)。ゲフィリンは、GABA 受容体のクラスター形成の足場タンパク質として機能して
おり、ERK はゲフェリンを介して GABAA受容体のシナプス後膜における局在を制御している可能性が
考えられ、その機能的役割の検討は今後の検討課題である。
25 結論
1、 中枢神経における静脈麻酔薬の作用機序及びその制御において、細胞内情報伝達機構が重要な役
割を果たしている。
2、 ERK は静脈麻酔薬の作用を抑制する方向に働き、ERK の活性化を抑制すると麻酔薬の作用は増強
する。
3、 上記メカニズムは ERK2 が主導的な役割を果たしているのに対し、ERK1 の関与は少ない。
26 謝辞
本研究を実施にするにあたり、細部にまでご指導いただきました
防衛医科大学校 麻酔学講座教授 風間富栄先生、
防衛医科大学校 麻酔学講座講師 佐藤泰司先生 に、
感謝申し上げます。ありがとうございました。
また、研究にご協力いただきました、
国立大学法人 浜松医科大学 神経生理学講座教授 福田敦夫先生
地方独立行政法人 東京都健康長寿医療センター研究所 老化制御研究チーム 遠藤昌吾先生
防衛医科大学校 麻酔学講座 スタッフ、研究科、専修医の先生方
防衛医科大学校 麻酔学講座 実験助手 高宮希代子さん
ありがとうございました。
27 引用文献
1)Yamakura T, Bertaccini E, Trudell JR, Harris RA. Anesthetics and Ion Channels: Molecular Models
and Sites of Action 1. Annual review of pharmacology and toxicology. 2001;41(1):23-51.
2)Krasowski M, Harrison N. General anaesthetic actions on ligand-gated ion channels. Cellular and
Molecular Life Sciences CMLS. 1999;55(10):1278-303.
3)Rudolph U, Antkowiak B. Molecular and neuronal substrates for general anaesthetics. Nat Rev
Neurosci. 2004 Sep;5(9):709-20.
4)Hemmings Jr HC, Akabas MH, Goldstein PA, Trudell JR, Orser BA, Harrison NL. Emerging
molecular mechanisms of general anesthetic action. Trends in pharmacological sciences.
2005;26(10):503-10.
5)Franks NP. General anaesthesia: from molecular targets to neuronal pathways of sleep and arousal.
Nature Reviews Neuroscience. 2008;9(5):370-86.
6)Bell-Horner CL, Dohi A, Nguyen Q, Dillon GH, Singh M. ERK/MAPK pathway regulates GABAA
receptors. Journal of neurobiology. 2006 Nov;66(13):1467-74.
7)Bell‐Horner CL, Dohi A, Nguyen Q, Dillon GH, Singh M. ERK/MAPK pathway regulates GABAA
receptors. Journal of neurobiology. 2006;66(13):1467-74.
8)Nishida E, Gotoh Y. The MAP kinase cascade is essential for diverse signal transduction pathways.
Trends in biochemical sciences. 1993;18(4):128-31.
9)Sweatt JD. Mitogen-activated protein kinases in synaptic plasticity and memory. Curr Opin
28 Neurobiol. 2004 Jun;14(3):311-7.
10)English JD, Sweatt JD. Activation of p42 mitogen-activated protein kinase in hippocampal long
term potentiation. The Journal of biological chemistry. 1996 Oct 4;271(40):24329-32.
11)Pagès G, Guérin S, Grall D, Bonino F, Smith A, Anjuere F, et al. Defective thymocyte maturation in
p44 MAP kinase (Erk 1) knockout mice. Science. 1999;286(5443):1374-7.
12)Selcher JC, Nekrasova T, Paylor R, Landreth GE, Sweatt JD. Mice lacking the ERK1 isoform of
MAP kinase are unimpaired in emotional learning. Learning & Memory. 2001;8(1):11-9.
13)Mazzucchelli C, Vantaggiato C, Ciamei A, Fasano S, Pakhotin P, Krezel W, et al. Knockout of
ERK1 MAP kinase enhances synaptic plasticity in the striatum and facilitates striatal-mediated
learning and memory. Neuron. 2002;34(5):807-20.
14)Satoh Y, Endo S, Ikeda T, Yamada K, Ito M, Kuroki M, et al. Extracellular signal-regulated kinase
2 (ERK2) knockdown mice show deficits in long-term memory; ERK2 has a specific function in
learning and memory. The Journal of Neuroscience. 2007;27(40):10765-76.
15)Imamura O, Satoh Y, Endo S, Takishima K. Analysis of Extracellular Signal‐Regulated Kinase 2
Function in Neural Stem/Progenitor Cells via Nervous System‐Specific Gene Disruption. Stem Cells.
2008;26(12):3247-56.
16)Satoh Y, Endo S, Nakata T, Kobayashi Y, Yamada K, Ikeda T, et al. ERK2 contributes to the control
of social behaviors in mice. The Journal of Neuroscience. 2011;31(33):11953-67.
17)Samuels IS, Karlo JC, Faruzzi AN, Pickering K, Herrup K, Sweatt JD, et al. Deletion of ERK2
mitogen-activated protein kinase identifies its key roles in cortical neurogenesis and cognitive
29
function. The Journal of Neuroscience. 2008;28(27):6983-95.
18)Pages G, Guerin S, Grall D, Bonino F, Smith A, Anjuere F, et al. Defective thymocyte maturation
in p44 MAP kinase (Erk 1) knockout mice. Science. 1999 Nov 12;286(5443):1374-7.
19)Satoh Y, Endo S, Nakata T, Kobayashi Y, Yamada K, Ikeda T, et al. ERK2 contributes to the control
of social behaviors in mice. The Journal of neuroscience : the official journal of the Society for
Neuroscience. 31. United States2011. p. 11953-67.
20)Heurteaux C, Guy N, Laigle C, Blondeau N, Duprat F, Mazzuca M, et al. TREK-1, a K+
channel involved in neuroprotection and general anesthesia. The EMBO journal.
2004;23(13):2684-95.
21)Eger EI, Saidman LJ, Brandstater B. Minimum alveolar anesthetic concentration: a standard of
anesthetic potency. Anesthesiology. 1965;26(6):756-63.
22)Sato Y, Kobayashi E, Hakamata Y, Kobahashi M, Wainai T, Murayama T, et al.
Chronopharmacological studies of ketamine in normal and NMDA ϵ1 receptor knockout mice†.
British journal of anaesthesia. 2004;92(6):859-64.
23)Durieux ME. Editorial I: powerful tools require careful handling--the case of the circadian
ketamine effect. Br J Anaesth. 92. England2004. p. 783-92.
24)Franks NP. Molecular targets underlying general anaesthesia. British journal of pharmacology.
2006;147(S1):S72-S81.
25)Petrenko AB, Yamakura T, Kohno T, Sakimura K, Baba H. Reduced immobilizing properties of
isoflurane and nitrous oxide in mutant mice lacking the N-methyl-D-aspartate receptor
30
GluR(epsilon)1 subunit are caused by the secondary effects of gene knockout. Anesthesia and
analgesia. 2010 Feb 1;110(2):461-5.
26)Jin H, Wu H, Osterhaus G, Wei J, Davis K, Sha D, et al. Demonstration of functional coupling
between γ-aminobutyric acid (GABA) synthesis and vesicular GABA transport into synaptic vesicles.
Proceedings of the National Academy of Sciences. 2003;100(7):4293-8.
27)Somogyi P, Takagi H, Richards JG, Mohler H. Subcellular localization of benzodiazepine/GABAA
receptors in the cerebellum of rat, cat, and monkey using monoclonal antibodies. The Journal of
neuroscience : the official journal of the Society for Neuroscience. 1989 Jun;9:2197-209.
28)Farrant M, Nusser Z. Variations on an inhibitory theme: phasic and tonic activation of GABAA
receptors. Nature Reviews Neuroscience. 2005;6(3):215-29.
29)Yeh J-H, Jeng C-J, Chen Y-W, Lin H-M, Wu Y-S, Tang C-Y. Selective enhancement of tonic
inhibition by increasing ambient GABA is insufficient to suppress excitotoxicity in hippocampal
neurons. Biochemical and biophysical research communications. 2005;338(3):1417-25.
30)Belelli D, Harrison NL, Maguire J, Macdonald RL, Walker MC, Cope DW. Extrasynaptic GABAA
receptors: form, pharmacology, and function. The Journal of Neuroscience. 2009;29(41):12757-63.
31)Antognini JF, Schwartz K. Exaggerated anesthetic requirements in the preferentially
anesthetized brain. Anesthesiology. 1993;79(6):1244-9.
32)Rampil IJ, Mason P, Singh H. Anesthetic potency (MAC) is independent of forebrain structures in
the rat. Anesthesiology. 1993;78(4):707-12.
33)Wakai A, Kohno T, Yamakura T, Okamoto M, Ataka T, Baba H. Action of isoflurane on the
31
substantia gelatinosa neurons of the adult rat spinal cord. Anesthesiology. 2005 Feb;102(2):379-86.
34)Kelleher RJ, 3rd, Govindarajan A, Jung HY, Kang H, Tonegawa S. Translational control by MAPK
signaling in long-term synaptic plasticity and memory. Cell. 2004 Feb 6;116(3):467-79.
35)Thomas GM, Huganir RL. MAPK cascade signalling and synaptic plasticity. Nature reviews
Neuroscience. 2004 Mar;5(3):173-83.
36)Iwakiri H, Nishihara N, Nagata O, Matsukawa T, Ozaki M, Sessler DI. Individual effect-site
concentrations of propofol are similar at loss of consciousness and at awakening. Anesthesia and
analgesia. 2005 Jan;100(1):107-10.
37)Krapivinsky G, Krapivinsky L, Manasian Y, Ivanov A, Tyzio R, Pellegrino C, et al. The NMDA
receptor is coupled to the ERK pathway by a direct interaction between NR2B and RasGRF1. Neuron.
2003;40(4):775-84.
38)Chen X, Shu S, Bayliss DA. HCN1 channel subunits are a molecular substrate for hypnotic actions
of ketamine. The Journal of Neuroscience. 2009;29(3):600-9.
39)Zhou C, Douglas JE, Kumar NN, Shu S, Bayliss DA, Chen X. Forebrain HCN1 channels contribute
to hypnotic actions of ketamine. Anesthesiology. 2013 Apr;118(4):785-95.
40)Hevers W, Hadley SH, Lüddens H, Amin J. Ketamine, but not phencyclidine, selectively modulates
cerebellar GABAA receptors containing α6 and δ subunits. The Journal of Neuroscience.
2008;28(20):5383-93.
41)Otsubo Y, Satoh Y, Kodama M, Araki Y, Satomoto M, Sakamoto E, et al. Mechanical allodynia but
not thermal hyperalgesia is impaired in mice deficient for ERK2 in the central nervous system. Pain.
32 2012 Nov;153(11):2241-52.
42)Steinbach JH, Akk G. Modulation of GABAA receptor channel gating by pentobarbital. The
Journal of physiology. 2001;537(3):715-33.
43)Bai D, Zhu G, Pennefather P, Jackson MF, MacDonald JF, Orser BA. Distinct functional and
pharmacological properties of tonic and quantal inhibitory postsynaptic currents mediated by
γ-aminobutyric acidA receptors in hippocampal neurons. Molecular pharmacology. 2001;59(4):814-24.
44)Jacob TC, Moss SJ, Jurd R. GABAA receptor trafficking and its role in the dynamic modulation of
neuronal inhibition. Nature Reviews Neuroscience. 2008;9(5):331-43.
45)大熊 誠, 芝崎 真, 黒川 和. キーワード解説 GABAA receptors GABAA受容体. 日本薬理学雑誌.
2008 2008.05;131(5):388-90.
46)Barnard E, Skolnick P, Olsen R, Mohler H, Sieghart W, Biggio G, et al. International Union of
Pharmacology. XV. Subtypes of γ-aminobutyric acidA receptors: classification on the basis of subunit
structure and receptor function. Pharmacological reviews. 1998;50(2):291-314.
47)Chang Y, Wang R, Barot S, Weiss DS. Stoichiometry of a recombinant GABAA receptor. The
Journal of neuroscience. 1996;16(17):5415-24.
48)Möhler H. GABAA receptor diversity and pharmacology. Cell and tissue research.
2006;326(2):505-16.
49)田中 佐, 吉田 武, 木内 祐. 臨床睡眠薬理の豆知識 GABAA受容体の構造と機能. 睡眠医療. 2008
2008.06;2(3):364-6.
50)Krasowski MD, Koltchine VV, Rick CE, Ye Q, Finn SE, Harrison NL. Propofol and other
33
intravenous anesthetics have sites of action on the γ-aminobutyric acid type A receptor distinct from
that for isoflurane. Molecular pharmacology. 1998;53(3):530-8.
51)Siegwart R, Jurd R, Rudolph U. Molecular determinants for the action of general anesthetics at
recombinant alpha(2)beta(3)gamma(2)gamma-aminobutyric acid(A) receptors. Journal of
neurochemistry. 2002 Jan;80:140-8.
52)Jurd R, Arras M, Lambert S, Drexler B, Siegwart R, Crestani F, et al. General anesthetic actions
in vivo strongly attenuated by a point mutation in the GABA(A) receptor beta3 subunit. FASEB J.
17. United States2003. p. 250-2.
53)Dong P, Zhou J-Y, Wu GS. Post-translational regulation of mitogen-activated protein kinase
phosphatase-2 (MKP-2) by ERK. Cell Cycle. 2010;9(23):4650-5.
54)Im BH, Rhim H. GABA(B) receptor-mediated ERK(1/2) phosphorylation via a direct interaction
with Ca(V)1.3 channels. Neurosci Lett. 2012 Mar 28;513(1):89-94.
55)Tyagarajan SK, Ghosh H, Yévenes GE, Imanishi SY, Zeilhofer HU, Gerrits B, et al. Extracellular
Signal-regulated Kinase and Glycogen Synthase Kinase 3β Regulate Gephyrin Postsynaptic
Aggregation and GABAergic Synaptic Function in a Calpain-dependent Mechanism. Journal of
Biological Chemistry. 2013;288(14):9634-47.
34 図表
図 1 ERK 経路
ERK経路は細胞外からの刺激を受けてRaf-MEK-ERKの順で活性化される。核内に移行したERKは細胞
増殖や細胞分化などに関する遺伝子の発現を制御する転写因子をリン酸化する。また、細胞質では
様々なタンパク質を基質としている。
転写因子
遺伝子発現 核
Raf
MEK1/2
ERK1/2
細胞質タンパク質
イオンチャネル
受容体
35 A
B
36
37
図2 Cre-loxPシステムを用いたErk2 CKOマウスのスキーム
Erk2のエクソン2(Ex2)、エクソン3(Ex3)をloxP塩基配列で挟み、Creと呼ばれる酵素を導入して遺
伝子を欠損させる方法である(A)。Erk2をloxP配列で挟んだErk2 floxマウスと、中枢神経特異的に
Creを発現するトランスジェニックマウスを掛け合わせることでErk2 conditional knockout mouse
(Erk2 CKOマウス、またはErk2 (flox/flox), Cre+、またはErk2(ΔCNS/ΔCNS))を作製した(B
1-3)。このマウスは、Creが発現している中枢神経ではErk2が欠損しており、Creが発現していな
い細胞は正常である。Erk2 (flox/+), Cre(-)、Erk2 (+/+), Cre(-)、Erk2 (flox/flox), Cre(-)、
Erk2 (+/+), Cre+をコントロールマウスとして用いた。
38 A
B
図3 Erk1 KOマウスに静脈麻酔薬投与後のLORR到達時間(Latency to LORR)と持続時間(Duration of
LORR)
(A,B)Erk1 KOマウス群とコントロールマウス群に、ペントバルビタール、プロポフォール、ケタ
ミン投与後のLORR到達時間と持続時間に差はなかった。(ペントバルビタール:Erk1 KOマウス群 n = 15,
コントロールマウス群 n = 16)(プロポフォール:Erk1 KOマウス群 n = 15, コントロールマウス群
n = 15)(ケタミン:Erk1 KOマウス群 n = 9, コントロールマウス群 n = 10)
Propofol (100 mg/kg)
Control Erk1KO 0
100 200 300
LatencytoLORR(s)
Pentobarbital (50 mg/kg)
Control Erk1KO 0
100 200 300
LatencytoLORR(s)
Control Erk1KO 0
50 100 150
Ketamine (200 mg/kg)
LatencytoLORR(s)
Propofol (100 mg/kg)
Control Erk1KO 0
10 20 30
DurationofLORR(min)
Pentobarbital (50 mg/kg)
Control Erk1KO 0
20 40 60 80
DurationofLORR(min)
Control Erk1KO 0
20 40 60 80 100
Ketamine (200 mg/kg)
DurationofLORR(min)