ブドウ
ブドウ
ブドウ
ブドウ‘Syrah’における
における
における
における rotundone 生合成
生合成
生合成機構
生合成
機構
機構
機構の
の
の
の解明
解明
解明
解明
Studies on Mechanism of Rotundone Biosynthesis
in ‘Syrah’ Grapes (Vitis vinifera L.)
山梨大学大学院
山梨大学大学院
山梨大学大学院
山梨大学大学院
医学工学総合教育部
医学工学総合教育部
医学工学総合教育部
医学工学総合教育部
博士課程学位論文
博士課程学位論文
博士課程学位論文
博士課程学位論文
2017 年
年
年 3 月
年
月
月
月
高瀬
高瀬
高瀬
高瀬
秀樹
秀樹
秀樹
秀樹
ブドウ
ブドウ
ブドウ
ブドウ‘Syrah’における
における
における
における rotundone 生合成機構
生合成機構
生合成機構
生合成機構の
の
の
の解明
解明
解明
解明
Studies on Mechanism of Rotundone Biosynthesis
in ‘Syrah’ Grapes (Vitis vinifera L.)
2017 年
年
年 3 月
年
月
月
月
高瀬
高瀬
高瀬
高瀬
秀樹
秀樹
秀樹
秀樹
Hideki Takase
目次 目次 目次 目次 凡例 凡例 凡例 凡例………..………...ii 序論 序論 序論 序論……….1 第 第 第 第 1 章章章 章 Rotundone 定量方法定量方法定量方法の定量方法のの開発の開発開発 開発 1-1 緒言………..3 1-2 実験材料と方法………..4 1-3 結果………..7 1-4 考察………10 1-5 要旨………11 第 第 第 第 2 章章 章章 Rotundone 生合成生合成生合成に生合成ににに関与関与関与する関与する酵素するする酵素酵素酵素のののの同定及同定及同定及び同定及びびび機能解析機能解析機能解析 機能解析 2-1 緒言………16 2-2 実験材料と方法………17 2-3 結果………24 2-4 考察………28 2-5 要旨………32 第 第 第 第 3 章章 章章 Rotundone ののの蓄積の蓄積蓄積を蓄積ををを決定決定する決定決定するする遺伝子する遺伝子遺伝子遺伝子ののの検討の検討検討検討 3-1 緒言………47 3-2 実験材料と方法………49 3-3 結果………52 3-4 考察………55 3-5 要旨………57 総合考察 総合考察 総合考察 総合考察………..63 参考文献 参考文献 参考文献 参考文献………..67 公表文献 公表文献 公表文献 公表文献………..73 謝辞 謝辞 謝辞 謝辞………..74 i
凡例
凡例
凡例
凡例
cDNA complementary deoxyribonucleic acid CTAB cetyl trimethyl ammonium bromide
CYP cytochrome P450
DMAPP dimethylallyl diphosphate
DTT dithiothreitol
DXP 1-deoxy-D-xylulose-5-phophate
DXR 1-deoxy-D-xylulose 5-phosphate reductoisomerase DXS 1-deoxy-D-xylulose-5-phosphate synthase
EIC extracted ion chromatogram FID flame ionization detector FPP farnesyl diphosphate
FPPS farnesyl diphosphate synthase GAP glyceraldehyde 3-phosphate
GC-MS gas chromatography mass spectrometry GPP geranyl diphosphate
GS-MS/MS gas chromatography-tandem mass spectrometry
HDR (E)-4-hydroxy-3-methylbut-2-enyl diphosphate reductase HMB-PP (E)-4-hydroxy-3-methyl-but-2-enyl pyrophosphate HMG-CoA 3-hydroxy-3-methylglutaryl-CoA
HMGR 3-hydroxy-3-methylglutaryl-CoA reductase HMGS 3-hydroxy-3-methylglutaryl-CoA synthase HPLC high performance liquid chromatography HRP horseradish peroxidase
IPP isopentenyl diphosphate KPi buffer potassium phosphate buffer LOD limit of detection
LOQ limit of quantification
MEP 2-C-methyl-D-erythritol-4-phosphate mRNA messenger ribonucleic acid
MRM multiple reaction monitoring MSD mass spectrometric detector
MVA mevalonate
NADPH nicotinamide adenine dinucleotide phosphate NMR nuclear magnetic resonace
ORF open reading frame
PCR polymerase chain reaction PDMS polydimethylsiloxane PVDF polyvinylidene difluoride RefSeq reference sequence
RNA ribonucleic acid
rRNA ribosomal ribonucleic acid RSD relative standard deviation
RT-PCR reverse transcription polymerase chaine reaction SBSE stir bar sorptive extraction
SD synthetic dextrose
SDS-PAGE sodium dodecylsulfate-polyacrylamide gel electrophoresis SIM selected ion monitoring
SPE solid phase extraction SPME solid phase microextraction SRS substrate recognition site TA titratable acidity
TDU thermal desorption unit TLC thin layer chromatography
TPS terpene synthase
Tris tris (hydroxymethyl) aminomethane
Vv Vitis vinifera
序論
序論
序論
序論
ブドウ‘Syrah’はフランス、コート・デュ・ローヌ地方北部を原産とする赤ワイン用 ブドウ品種で、ブドウ‘Mondeuse blanche’と‘Dureza’の交雑によって生まれたと考えら れている (Bowers et al. 1998)。また、‘Syrah’のシノニムとして、‘Shiraz’が知られ、主 にオーストラリアでその名称が使われている。近年フランスだけではなく、オーストラリ ア、南アフリカ共和国、アルゼンチン、チリ、アメリカ合衆国、イタリア、ニュージーラ ンド、ギリシャ、スペイン、スイスなど世界各国で急速に栽培が拡大しており、2000 年か ら 2010 年までに世界全体のワイン用ブドウ栽培面積における‘Syrah’栽培面積割合が 2.1 %から 4.0 %にまで急増し、2010 年時点で‘Cabernet sauvignon’、‘Merlot’、‘Airen’、 ‘Templanillo’、‘Chardonnay’に次ぐ、第 6 番目に世界的に広く栽培されている重要品種 である (Anderson 2014)。ブドウ‘Syrah’は、日本においても山梨県、長野県で栽培が始 められており、ブドウ‘Syrah’の特徴を反映したワインが生産され始めている。‘Syrah’ ワインの香りの特徴は栽培地域の気候によって左右されるが、冷涼な地域で生産された ‘Syrah’ワインにおいて特にスパイス、胡椒などの表現がなされ、この特徴に起因する化 合物としてセスキテルペノイドの一種である rotundone が同定された (Wood et al. 2008)。 実際、この化合物は黒胡椒、白胡椒、オレガノ、バジル、タイムなど、様々なハーブやス パイスにも含まれていることが確認された。
Rotundone は、guaiene 骨格を有したセスキテルペンケトン (Fig.) であり、その官能閾 値は水中 8 ng L-1、赤ワイン中 16 ng L-1と極めて低い (Siebert et al. 2008)。ブドウ‘Syrah’
以外の品種では、オーストラリア産の‘Mourvèdre’、‘Durif’ (Herderich et al. 2012)、欧州 産の‘Duras’、‘Schioppettino’、‘Vespolina’、‘Grüner Veltliner’等で rotundone が同定され ている (Caputi et al. 2011; Geffroy et al. 2014; Mattivi et al. 2011)。また、rotundone はブドウ 果実の中では果皮に局在し、véraison 期から収穫に向けて顕著に増加する。しかしながら、 rotundone は疎水性が高いため、ワイン醸造工程において果皮からの抽出効率は低い (Caputi et al. 2011)。そのため、発酵後のワイン中の rotundone 含有量は、醸造に用いたブド ウ果実中の含有量に依存する。近年の研究の中で、rotundone 蓄積に影響する環境ファクタ ーとして、冷涼な気候 (Caputi et al. 2011)、土壌の地質及び地形 (Scarlett et al. 2014)、灌漑 による土壌湿度や除葉による果房への露光 (Geffroy et al. 2014) などが報告されている。し
かしながら、これらの環境ファクターが直接的あるいは間接的に影響を及ぼす、ブドウ果 実における rotundone 生合成機構はこれまでに明らかにされていない。 そこで本研究では、日本で栽培されたブドウ‘Syrah’を用い、rotundone 生合成メカ ニズムを解明することを目的とした。第 1 章では、複雑な前処理が必要だった従来法と比 較して、より簡便で且つ高感度な新規 rotundone 定量法の開発を目指した。さらに日本で 栽培したブドウ‘Syrah’と他のブドウ品種の rotundone 濃度の定量及び、フランス産及び 日本産‘Syrah’ワインの rotundone 濃度の定量を行い、其々の比較を行った。第 2 章では、 ブドウにおける rotundone 生合成機構上の未同定酵素の単離を目的として、‘Syrah’果実か らその候補遺伝子を単離するとともに、単離遺伝子がコードする酵素の機能性解析を試み た。第 3 章では、rotundone 生合成の遺伝的制御に関する知見を得るために、α-guaiene 及 び rotundone の異なる蓄積パターンを示す、2 つの異なる畑で栽培された日本産‘Syrah’ 果実を用いて rotundone 生合成に関連する遺伝子のブドウ成熟期間中の発現パターンを比 較し、ブドウ果実中の rotundone 蓄積を決定する遺伝子の検討を行った。
第
第
第
第 1 章
章
章
章
Rotundone 定量方法
定量方法
定量方法
定量方法の
の
の
の開発
開発
開発
開発
1-1
諸言
諸言
諸言
諸言
これまで rotundone の定量は、ブドウ果実またはワインに極微量にしか含まれていな いために固相抽出 (SPE) カラムを用いた前処理後に SPME (solid phase micro extraction) 法 による抽出を行うなど、複雑な操作が必要とされてきた (Caputi et al. 2011; Geffroy et al. 2014; Mattivi et al. 2011; Siebert et al. 2008)。また、この SPE カラムを用いた前処理には、例 えば、ワイン分析のためには約 100 mL 程度の比較的大容量のサンプル量が必要で、さら にブドウ果実抽出物といった濁度の高いサンプルを分析する場合、SPE カラムが頻繁に詰 まるため、多数のサンプルを効率的に分析するのに不都合があった。Stir bar sorptive extraction (SBSE) 法は、溶液中に含まれる化合物の抽出、濃縮を目的に用いられる手法で (Baltussen et al. 1999)、近年ブドウ果実やワイン中に極微量に含まれる様々な香気成分分析 に採用されている。SBSE 法は、SPE カラムを用いた前処理と比べて、化合物の高い濃縮 効率のほか、幾つかの利点を有する。例えば、SPE カラムはその使用直前に数種の有機溶 媒を用いたコンディショニングが必要となるが、SBSE 法に用いるスターバーのコンディ ショングは予め事前に実施でき、且つ容易である。また、SBSE 法は SPE カラムのように 濁度の高いサンプルを用いたとしても詰まることはない。 そこで本章では、第一に、高感度でより簡便な方法として SBSE 法と 2 次元ガスクマ トグラフ質量分析計 (2 次元 GC-MS) を組み合わせた定量法を開発した。
1-2
実験材料
実験材料
実験材料
実験材料および
および
および
および方法
方法
方法
方法
1-2-1
試薬
試薬
試薬
試薬
Rotundone は Mattivi らの方法 (Mattivi et al. 2011)、2H5-rotundone は Siebert らの方法
(Siebert et al. 2008) により合成した。Rotundone の1H nuclear magnetic resonance (NMR, 400 MHz)及び13C nulear magnetic resonance (NMR, 100MHz) スペクトルは、Bruker AVANCE 400 spectrometer によって取得し、既報と同一であることを確認した。NMR の測定は常温、重 水素化 chloroform 中で行われた。化学シフトは ppm 単位で記録された。2
H5-rotundone は
GC-MS 分析 (スキャンモード) で得られたマススペクトルを用いて同定した。
合成で用いたすべての試薬、NMR で使用した重水素化 chloroform、その他分析で使用 した HPLC-grade ethanol、HPLC-grade methanol、dichloromethane、acetone、n-hexane、n-pentane、 ethyl acetate、酒石酸はシグマアルドリッチジャパンから購入した。Mili-Q 水は Mili-Q purification system (Merk Millipore) で調整した。
1-2-2
ブドウ
ブドウ
ブドウ
ブドウ及
及
及
及びワインサンプル
びワインサンプル
びワインサンプル
びワインサンプル
山梨県韮崎市 (lat. 35o
41’6’’N; long. 138o22’9’’E; 標高 約 540 m) で栽培したブドウ‘甲 州 (Koshu) [Vitis vinifera] ’、笛吹市 (lat. 35o
38’51’’N; long. 138o41’54’’E; 標高 約 350m)で栽 培されたブドウ‘Muscat Bailey A [V. labrusca (Bailey) × V. vinifera (Muscat Hamburg)]’、甲 州市 (城の平ヴィンヤード、lat. 35º38'59"N; long. 138º44'43"E; 標高 589 m) で栽培されたブ ドウ‘Syrah [V. vinifera]’及び‘Cabernet Sauvignon [V. vinifera]’、及び長野県上田市 (マリ コヴィンヤード、lat. 36º20'35"N; long. 138º18'5"E; 標高 640 m) で栽培されたブドウ‘Syrah’、 ‘Merlot [V. vinifera]’、‘Sauvignon Blanc [V. vinifera]’のブドウ果実を rotundone の定量に用 いた。ブドウのサンプリングは 2012 年に行われた。収穫されたブドウは、直ちに除梗され た後、分析に使用されるまで-30 o C で保存された。 分析に供試した‘Syrah’ワインは、市販品を購入し、分析に使用されるまで 20 o C で 保存された。
1-2-3
サンプルの
サンプルの
サンプルの
サンプルの調整
調整
調整
調整
ブドウ果実に含まれる rotundone を定量するために、冷凍ブドウ果実 (約 200 g) は、 種子の破砕を最低限に抑えながら、ミル (IFM-62DG、岩谷産業) を用いて 30 秒間破砕・ 均一化された。均質化された果実の破砕物 10 g を計量し、内部標準として2 H5-rotundone (100 µL、100 µg L-1) を添加した後、n-pentane:ethyl acetate (9:1、v/v)を 20 mL 加え、シェイカー を用いて 1 時間撹拌・抽出した。得られた抽出物は、3000 rpm、10 分間遠心分離した。分 離した有機溶媒画分を試験管に移し替え、窒素ガスパージにより有機溶媒を注意深く取り 除いた。残った残差を ethanol 500 mL を用いて溶解し、その後酒石酸バッファー (pH3.2) 4.5 mL を加えた。以上のように調整された試料を、SBSE 法による抽出、2 次元 GC-MS 分析 に供試した。 ワインに含まれる rotundone の定量には、ワイン 5 mL に対し、内部標準として 2 H5-rotundone (100 µL、100 µg L -1 ) を添加したものを、SBSE 法による抽出、2 次元 GC-MS 分析に供した。 果実成分分析のために用いた果汁は、ミル (IFM-62DG) で破砕・均一化されたブドウ 破砕物を搾汁率 60 % (重量) で搾汁することで調整された。全可溶性固形物は屈折計 (Pocket PAL-1、株式会社アタゴ) を用いて計測し、oBrix として現した。滴定酸度 (TA、 titratable acidity) は 0.1 N 水酸化ナトリウムを用いた滴定で計測した。TA は酒石酸換算 (g L-1) で現した。1-2-4
Stir bar sorptive extraction (SBSE) 法
法
法
法
Polydimethylsiloxane (PDMS、24 µL) がコーティングされたスターバー (Twister、 GESTEL GmbH) は、窒素ガス雰囲気下で、300 oC、60 分間加熱し、調整された。Rotundone 定量用に調整されたブドウ果実或いはワインサンプルを、10 mL 容量のヘッドスペースバ イアルに移し、スターバーを加えた後、スクリューキャップで密閉した。SBSE は、1500 rpm、 60 分間、室温で撹拌することで行われた。撹拌後、スターバーを取り出し、Mili-Q 水で洗 浄、キムワイプ (日本製紙クレシア) にて水分を取り除いた後、抽出用ガラスライナーに 入れ、熱脱着ユニット (TDU、GESTEL GmbH) にセットし、2 次元 GC-MS 分析に供試し
た。使用後のスターバーは、methanol/dichloromethane (1:1) 中に 24 時間浸漬後、窒素雰囲 気下で 300 o
C、60 分間加熱による再調整後、次回の使用まで常温で保存された。
1-2-5
2 次元
次元
次元 GC-MS 分析条件
次元
分析条件
分析条件
分析条件
2 次元 GC-MS 分析は、7890A GC システム (Agilent Technologies) を基本として、1 次 元目の検出向けに Flame ionization detector (FID)、2 次元目の検出向けに質量分析計 (5975C inert XL MSD、Agilent Technologies)、加熱脱着ユニット (TDU、GESTEL GmbH) 、クール ドインジェクションシステム (CIS4、GESTEL GmbH)、Agilent Low Thermal Mass II モジュ ール (LTMII、Agilent Technologies) を接続し、行った。
分析条件
カラム:1 次元 ; BP20 カラム (50 m x 0.22 mm x 0.25 µm、Trajan Scientific and Medical) 2 次元 ; DB-1 カラム (10 m x 0.18 mm x 0.40 µm、Agilent Technologies) TDU 条件:40 o
C (0.5 min) - 200 oC min-1 - 300 oC (3 min) CIS4 条件:10 o
C (0 min) - 720 oC min-1 - 250 oC (1 min) 脱着流量:50 mL min-1
(Nitrogen)
注入モード:スプリットレスモード (スプリットレス時間:3 min) オーブン:BP20 カラム;60 o
C (3 min) - 5 oC min-1 - 240 oC (17 min) DB-1 カラム;35 o
C (40.8 min) - 20 oC min-1 - 250 oC (5 min) ハートカッティング時間:37 min - 40 min
FID 温度:250 oC
インターフェース温度:250 oC
イオン化モード:electron impact ionization (70 eV)
測定モード:スキャンモード (定性) 、スキャンレンジ;m/z 40 - 300
Selected ion monitoring (SIM)モード (定量) 、選択イオン;m/z 147、161、 203、208、218、223
ターゲットイオン:rotundone ; m/z 218、2
1-3
結果
結果
結果
結果
1-3-1
SBSE 法及
法及
法及び
法及
び
び GC-MS 分析条件
び
分析条件の
分析条件
分析条件
の
の
の最適化
最適化
最適化
最適化
SBSE 法は SPME 法と同じ原理に基づいているが、SPME 法と比較してより大きい PDMS 相を有しているため、より高い感度での分析が可能となる。SBSE 法における化合 物の回収率は PDMS 相と液相間の分配係数により制御される (Prieto et al. 2010)。分配係数 は octanol/水分配係数 (Ko/w) と相関しているため、抽出する対象の化合物の Ko/wをもとめ
ることで、SBSE 法による理論的な回収率を求めることができる。既報によると、rotundone の log Ko/wは 4.98 と予測されており (Mattivi et al. 2011)、それをもとに今回採用した SBSE
法の条件 (PDMS 24 mL、5 mL サンプル溶液中) での rotundone の回収率を計算した結果、 99.8 %と高い数値であった。このことから、rotundone を抽出する方法として、SBSE 法は 優れていることが示唆された。 2 次元 GC-MS 分析条件は、rotundone 及び2H5-rotundone に対するターゲットイオンを より効率的に同定するために最適化された。Rotundone 定量に向けたハートカッティング 時間は、スキャンモードでの GC-MS 分析において rotundone をリテンションタイム 49.39 min に検出したことから (Fig.)、その時間を含む 37 min - 40 min に決定した。
1-3-2
バリデーション
バリデーション
バリデーション
バリデーション
Rotundone の定量法のバリデーションには、rotundone が検出限界以下であったブドウ ‘Cabernet Sauvignon’のブドウ果実及びワインに、rotundone 及び、内部標準として 2 H5-rotundone がそれぞれ添加されたものを用いた。校正曲線の直線性は、ブドウ果実では 10 から 3000 ng kg-1の範囲 (r2 = 0.99967)、ワインでは 10 から 1200 ng L-1の範囲 (r2 = 0.99956) で確認された (Table 1-1)。検出限界 (LOD) 及び定量限界 (LOQ) は、それぞれ signal-to-noise ratios (S/N) で 3:1、 10:1 を示す rotundone 濃度として決定した。その結果、ブドウ果実を分析する際の LOD 及 び LOQ はそれぞれ 2.1、6.5 ng kg-1、ワインを分析する際の LOD 及び LOQ はそれぞれ 2.4、
Mattivi et al. 2011)。次に、ブドウ果実及びワインからの rotundone の回収率はそれぞれ 106 % (添加濃度 120 ng kg-1)、96 % (添加濃度 120 ng L-1) となり、相対標準偏差 (RSD) は 1.7 %、 2.3 %であった (n = 3)。
1-3-3
日本産
日本産
日本産
日本産ブドウに
ブドウに
ブドウに
ブドウに含
含まれる
含
含
まれる
まれる rotundone の
まれる
の
の定量
の
定量
定量
定量
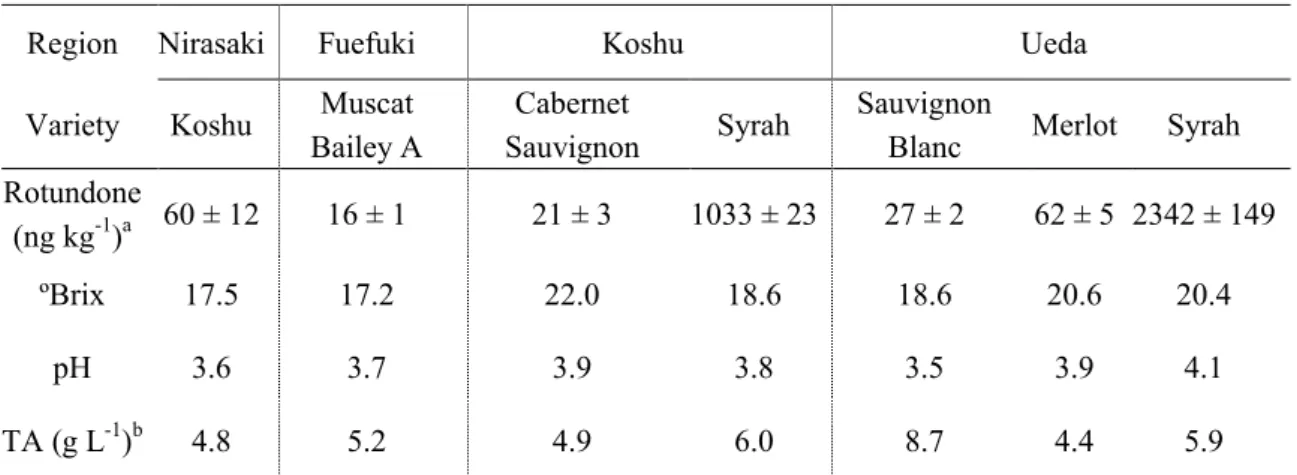
開発した分析法で日本の異なる産地で栽培される、‘Syrah’を含む 6 種類のブドウ品 種に含まれる rotundone 濃度を定量した。長野県上田市 (マリコヴィンヤード) および山梨 県甲州市 (城の平ヴィンヤード) で栽培された‘Syrah’果実の rotundone 含有量は、それ ぞれ 2342、1033 ng kg-1と、国内で栽培される他のブドウ品種と比較して顕著に高いことが認められた (Table 1-2)。これらの結果は、rotundone が‘Syrah’果実及びワインの特徴香 として貢献していることを支持した (Wood et al. 2008)。既報において、冷涼な地域で栽培 されたブドウ果実中に高い水準で rotundone が蓄積することが報告されている。また、上 に示したようにマリコヴィンヤードで栽培された‘Syrah’果実の rotundone 含有量は、城 の平ヴィンヤードで栽培されたものと比較して顕著に高かった。マリコヴィンヤードは、 緯度の違いや地理的要因のために城の平ヴィンヤードと比べて冷涼な気候を有する。この 気候の違いによって両産地間の‘Syrah’果実における rotundone 含有量に差が生まれたと 考えられた。さらに、日本で栽培された‘Syrah’果実、特にマリコヴィンヤードで栽培さ れた‘Syrah’果実は、オーストラリアで栽培された‘Syrah’果実で報告された rotundone の最大濃度 (1082 ng kg-1
) よりも高い水準の rotundone を含有していた (Scarlett et al. 2014)。
1-3-4
Syrah ワインに
ワインに
ワインに含
ワインに
含
含まれる
含
まれる rotundone の
まれる
まれる
の
の
の定量
定量
定量
定量
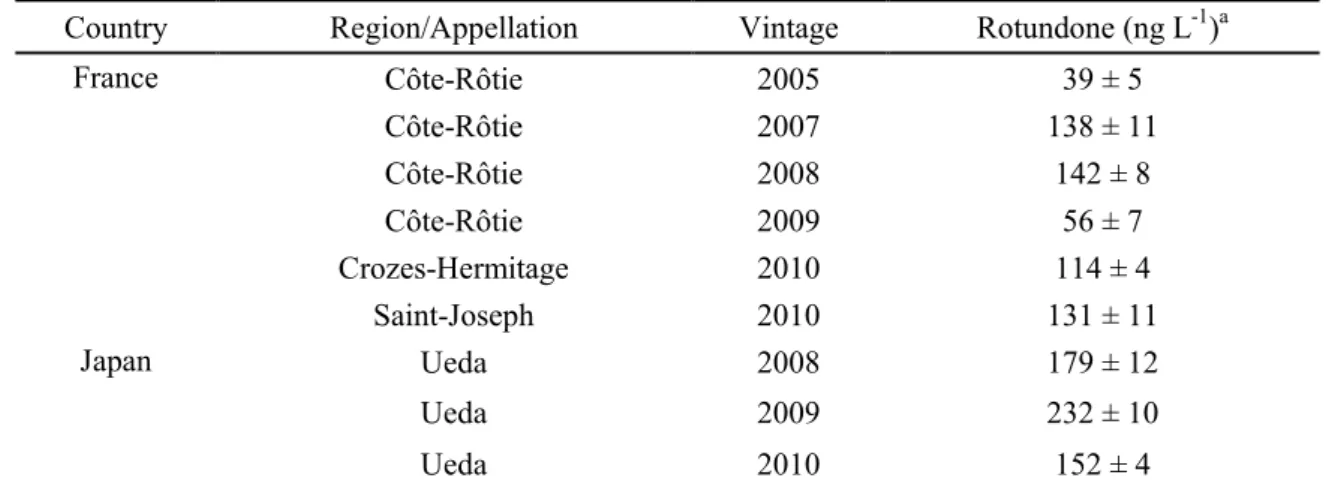
開発した分析法で市販の‘Syrah’ワイン中の rotundone の定量を行った (Table 1-3)。 市販の‘Syrah’ワインは、マリコヴィンヤードで栽培された‘Syrah’果実を用いて醸造 された‘Syrah’ワインに加えて、フランスの Côte-Rôtie、Crozes-Hermitage、Saint-Joseph といったアペラシオンで生産されたワインが選ばれた。生産年 (2008 年、2009 年、2010 年) が異なるマリコヴィンヤードで栽培された‘Syrah’果実を用いて醸造されたワインを 分析した結果、179±12、232±10、152 ± 4 ng L-1の rotundone を検出した。これらの濃度
は、これまで海外で報告されている‘Syrah’ワイン中の含有量と比較しても高い数値 (オ ーストラリア、グランピアンズ産‘Shiraz’ワイン:最大値 161 ng L-1
) であった (Herderich et al. 2012)。
1-4
考察
考察
考察
考察
本章では、SBSE 法及び 2 次元 GC-MS を用いて、高感度でより簡便な、ブドウ果実及 びワイン中の rotundone 定量方法の開発を行った。開発した分析法により、従来法で行わ れていた SPE を用いた前処理工程を省略でき、更に少量のサンプル (例、ワイン 5 mL) を 用いた定量が可能となった。日本で栽培された様々なブドウ品種を分析した結果、‘Syrah’ 果実が他の品種と比較して顕著に rotundone を高含有しており、この化合物がブドウ‘Syrah’ に特徴的に含まれる成分であること改めて確認した。また、日本、特に長野県上田市のマ リコヴィンヤードで栽培された‘Syrah’果実及びそれを用いて醸造されたワインを分析し た結果、フランス或いはオーストラリア産の‘Shiraz’ワインと比較しても、高い水準で rotundone を含有した。これらの結果は、日本は rotundone を高濃度で含み、胡椒様アロマ の豊かな‘Syrah’ワインの醸造に適しており、日本に存在する何らかの環境ファクターが そこに育つ‘Syrah’果実中の rotundone 蓄積を促進していることを示唆した。高い品質の ‘Syrah’果実を継続的に得るために、地域的な気候特徴が‘Syrah’果実中の rotundone 蓄 積に及ぼす影響を調査する必要性が考えられた。1-5
要旨
要旨
要旨
要旨
Rotundone は、ワイン中の胡椒様香気に貢献する酸化型セスキテルペノイドである。 これまでの rotundone の定量方法は、ブドウ果実及びワイン中に極微量にしか含まれてい ないために SPE を用いた前処理後に SPME 法による抽出を行うなど、複雑な操作が必要 とされてきた。本研究において、我々はブドウ果実及びワイン中に微量に含まれる rotundone を高感度で簡便な定量法を開発した。この定量法には SBSE 法を用いた 2 次元 GC-MS 分析を採用し、従来法で行われていた SPE を用いた前処理工程を省略した。ブド ウ果実を分析する際の LOD 及び LOQ はそれぞれ 2.1、6.5 ng kg-1、ワインを分析する際の LOD 及び LOQ はそれぞれ 2.4、7.2 ng L-1であった。次に、ブドウ果実及びワインからの rotundone の回収率はそれぞれ 106 % (添加濃度 120 ng kg-1)、96 % (添加濃度 120 ng L-1) と なり、相対標準偏差 (RSD) は 1.7 %、2.3 %であった (n = 3)。開発した定量法を用いた分 析の結果、マリコヴィンヤードで栽培された‘Syrah’果実及びそれを用いて醸造されたワ イン中の rotundone 濃度は、フランス産‘Syrah’ワインと比較しても高い水準であり、さ らにオーストラリア産‘Shiraz’果実及びワインで報告された最大 rotundone 濃度より高か った。これらの結果は、日本に存在する環境因子がそこに育つ‘Syrah’果実中の rotundone 蓄積を促進していることを示唆するとともに、高い品質の‘Syrah’果実を継続的に得るた めに、地域的な気候特徴が‘Syrah’果実中の rotundone 蓄積に及ぼす影響を調査する必要 性を示した。Fig. Two-dimensional chromatogram of rotundone. Six hundred ng L-1 of rotundone was spiked in Cabernet Sauvignon wine. (A) SIM mode (m/z 218). (B) The mass spectrum of rotundone obtained in scan mode at the retention time of 49.39 min.
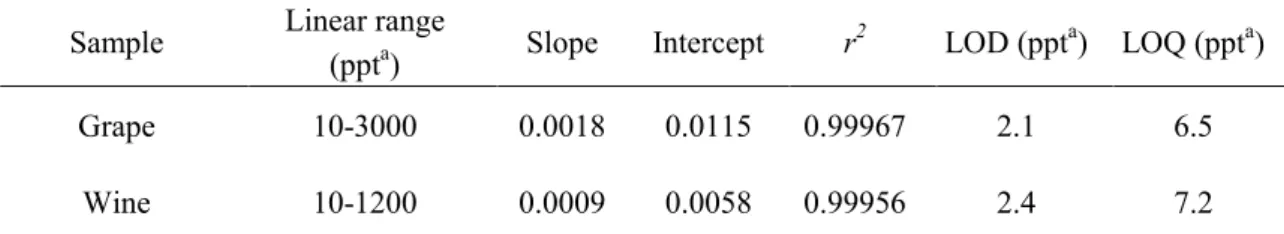
Table 1-1 Linearity, LOD, and LOQ of rotundone in Cabernet Sauvignon grape and wine
Sample Linear range
(ppta) Slope Intercept r 2 LOD (ppta) LOQ (ppta) Grape 10-3000 0.0018 0.0115 0.99967 2.1 6.5 Wine 10-1200 0.0009 0.0058 0.99956 2.4 7.2 a
ppt (parts per trillion), rotundone concentration expressed as ng kg-1 of grape or ng L-1 of wine; LOD, limit of detection; LOQ, limit of quantification
Table 1-2 Berry parameters and rotundone concentration in various grapes cultivated in Japan
Region Nirasaki Fuefuki Koshu Ueda
Variety Koshu Muscat Bailey A
Cabernet
Sauvignon Syrah
Sauvignon
Blanc Merlot Syrah Rotundone (ng kg-1)a 60 ± 12 16 ± 1 21 ± 3 1033 ± 23 27 ± 2 62 ± 5 2342 ± 149 ºBrix 17.5 17.2 22.0 18.6 18.6 20.6 20.4 pH 3.6 3.7 3.9 3.8 3.5 3.9 4.1 TA (g L-1)b 4.8 5.2 4.9 6.0 8.7 4.4 5.9 a
Rotundone concentration are shown as means ± standard deviation of triplicate experiments.
b
Table 1-3 Rotundone concentrations in Syrah wines from different regions
Country Region/Appellation Vintage Rotundone (ng L-1)a
France Côte-Rôtie 2005 39 ± 5 Côte-Rôtie 2007 138 ± 11 Côte-Rôtie 2008 142 ± 8 Côte-Rôtie 2009 56 ± 7 Crozes-Hermitage 2010 114 ± 4 Saint-Joseph 2010 131 ± 11 Japan Ueda 2008 179 ± 12 Ueda 2009 232 ± 10 Ueda 2010 152 ± 4 a
第
第
第
第 2 章
章
章
章
Rotundone 生合成
生合成
生合成
生合成に
に関与
に
に
関与
関与する
関与
する
する
する
酵素
酵素
酵素
酵素の
の
の同定及
の
同定及
同定及び
同定及
び
び
び機能解析
機能解析
機能解析
機能解析
2-1
諸言
諸言
諸言
諸言
α-Guaiene は、セスキテルペン炭化水素類に属し、様々な植物由来の精油やブドウ果実 中に含有されることが報告されている (Coelho et al. 2006; Kapadia et al. 1967; Pino et al. 2001; Rakotonirainy et al. 1997; Schreier et al. 1976)。この化合物は rotundone と共通し、特有の 構造である五員環と七員環を有していることから、rotundone の前駆物質と考えられている。 つまり、rotundone は、α-guaiene 上の 2 位の炭素がケトン基へと酸化された結果、生成され る。実際、α-guaiene が空気酸化された結果、(2R)-rotundol や (2S)-rotundol といった中間体 を介し、rotundone が生成することが提案された (Huang et al. 2014; Huang et al. 2015a; Huang et al. 2015b)。しかしながら、ブドウ由来の酵素によるα-guaiene から rotundone を生合成する メカニズムについてはこれまで報告されてこなかった。セスキテルペン炭化水素類の酸化 を触媒可能な酵素としては、Cytochrome P450 (CYP) に属する酵素が挙げられ、具体的には premnaspirodiene oxygenase CYP71D55 (Takahashi et al. 2007)、α-humulene 10-hydroxylase CYP71BA1 (Yu et al. 2011)、(+)-valencene oxidase CYP71AV8 (Cankar et al. 2011) 等が挙げられ る。そこで本章では、α-guaiene から rotundone への変換と同様な酵素反応を触媒する premnaspirodiene oxygenase の遺伝子配列を基に、12-fold coverage genome sequence assembly (Pinot Noir PN40024) (Jaillon et al. 2007) から候補遺伝子を選出し、ブドウ‘Syrah’から候補 遺伝子を単離するとともに、単離遺伝子がコードする酵素の機能性解析を試みた。
2-2
実験材料
実験材料
実験材料
実験材料および
および
および
および方法
方法
方法
方法
2-2-1
植物材料及
植物材料及
植物材料及
植物材料及び
び
び
び試薬
試薬
試薬
試薬
本章の実験で用いたブドウ‘Syrah’及び‘メルロー’は、長野県上田市に位置するマ リコヴィンヤード (lat. 36o 20’35’’N; long. 138o18’5’’E; 標高 約 640 m) で栽培され、2012 年 及び 2013 年に収穫されたブドウ果実を試験に供した。 Rotundone 及び2H5-rotundone は、1-2-1 で示した方法で調整した。α-Guaiene は、guaiol
に thionyl chloride を加え、脱水することで合成された。 合成したα-guaiene の精製は、Merck silica gel 64F-254 (0.25 mm、precoated) を用いた薄層クロマトグラフィにより行い、NMR により同定した。GC-MS により純度を求めた結果、90 %であった。β-Nootkatol は Takahashi らの方法 (Takahashi et al. 2007) に従い、nootkatone の還元によって合成され、NMR により 同定した。
合成で用いたすべての試薬、NMR で使用した重水素化 chloroform、その他分析で使用 し た HPLC-grade ethanol 、 HPLC-grade methanol 、 n-hexane 、 n-pentane 、 ethyl acetate 、 (-)-α-cedrene、(+)-valencene はシグマアルドリッチジャパンから購入した。Mili-Q 水は Mili-Q purification system (Merk Millipore) で調整した。
2-2-2
Syrah 果実
果実
果実からの
果実
からの
からの候補遺伝子
からの
候補遺伝子の
候補遺伝子
候補遺伝子
の
の
の単離
単離
単離
単離
長野県上田市で栽培された‘Syrah’果実の外果皮から CTAB 法により、total RNA を抽 出した (Chang et al. 1993)。抽出された total RNA は、RNase-Free DNase set (Qiagen) を用い て処理した後、RNeasy Plant Mini kit (Qiagen) を用いて精製した。cDNA は SuperScript III reverse transcriptase (Life Technologies Inc.) を用いて合成した。候補遺伝子の full-length cDNA を取得するために、polymerase chain reaction (PCR) は Phusion High-Fidelity DNA Polymerase (New England BioLabs Inc.) を用いて実施した。PCR のテンプレートには精製し た cDNA を用いた。各候補遺伝子に対する特異的プライマーは、12-fold coverage genome sequence assembly (Pinot Noir PN40024) (Jaillon et al. 2007) の情報を基に設計した。VvSTO2 向けのプライマー1 及び 2 は XM_010646246、VvSTO4 向けのプライマー3 及び 4 は
XM_010654905、VvSTO6 向けのプライマー5 及び 6 は XM_010657579 及び NC_012016 の情 報を基にそれぞれ設計された (Table 2-1)。約 1.5 kb の得られた PCR 産物は、DNA ligation kit (Takara Bio Inc.) を用いた TA クローニングを介し、pT7 Blue vector にサブクローニングさ れた後、シークエンスにより遺伝子配列を確認した。
2-2-3
形質転換酵母
形質転換酵母
形質転換酵母
形質転換酵母の
の
の作成
の
作成
作成
作成
C 末端 V5 epitope 融合タンパク質の生成するために、プライマー7 及び 8(VvSTO2)、 9 及び 10 (VvSTO4)、11 及び 12 (VvSTO6) を用いた PCR により、ストップコドンを取り除 いた候補遺伝子 cDNA フラグメントをそれぞれ作成した (Table 2-1)。作成した cDNA フラ グメントは、pYES2.1/V5-His-TOPO® vector (Invitrogen) を用い、Gietz 及び Schiestl の方 法 (Gietz and Schiestl 2007) に従い、BJ2168 yeast strain (MATa, prc1-407, prb1-1122, pep4-3, leu2, trp1, ura3-52, gal2; Nippon Gene) に導入した。
2-2-4
レコンビナ
レコンビナ
レコンビナ
レコンビナン
ン
ン
ントタンパク
トタンパク質
トタンパク
トタンパク
質
質
質の
の
の
の調整
調整
調整
調整
前培養として、50 mL の uracil を含まない SD 培地 ( 2% glucose) 中に、30 o C、200 rpm で一晩培養した。およそ定常期まで生育した前培養液の OD600を計測した後、本培養に使 用する 250 mL の uracil を含まない SD 培地 (2% galactose) を OD600 = 0.4 に調整するための 必要量を算出した。上記必要量分の前培養液を 3000 x g、4 o C で 5 分間遠心分離し、集菌 された酵母細胞を本培養用の培地に加えた後、30 o C、200 rpm にておよそ 10-16 時間、 OD600 = 1.2 前後になるまで培養した。 酵母からのタンパク質抽出は、スフェロプラスト法を用いた。本培養後の菌液を 3000 x g、4 oC 条件で 5 分間の遠心分離による集菌を行い、ペレットとなった菌体に湿潤体重量 1 g に対して 10 mL の 100 mM Tris-HCL pH 9.2 (10 mM DTT) を添加した。30 oC、90 rpm で 30 分間処理し、3000 x g、4 oC 条件で 5 分間の遠心分離により、ペレットを回収した後、 10 mL sorbitol/KPi buffer (1.2 M sorbitol、20 mM potassium phosphate buffer pH 7.4) で洗浄し た。再度遠心により、ペレットを回収した後、菌体に 10 mL の sorbitol/KPi buffer を加え、 Zymolyase 20-T 5 mg を添加、30 oC、90 rpm、60 分間処理した。細胞壁消化程度の確認は、蒸留水中でスフェロプラストが破裂して処理後の OD600値が処理前の 10 % 程度にまで減 少することを利用して行った。酵母スフェロプラストを 3000 x g、4 o C 条件で 5 分間の遠 心分離により回収し、上澄みを除いた後、氷冷しておいた 10 mL の sorbitol/KPi buffer に て洗浄し、再度 3000 g、4 o C にて 5 分間遠心してペレットにした。得られたペレットに氷 冷しておいた 7 mL breaking buffer (0.6 M sorbitol、20 mM Tris-HCL pH 7.4) を加え、15 mL 容ダウンス型破砕機 (WHEATON) に移し、泡立てないように 20 回上下させて細胞を破砕 した。細胞破砕液を、氷冷しておいた 50 mL 容ファルコンチューブに移し、3000 x g、4 o C 条件で 5 分間の遠心分離により、未破砕細胞や核等を沈殿させた。レコンビナントタンパ ク質を含むミクロソーム画分は 100,000 x g、4 o C にて 60 分間超遠心機 (Ultracentrifuge TL、 BECKMAN) により遠心することで回収した。ミクロソーム画分の沈殿を、500 µL の 50 mM Tris-HCl pH 7.5 (20 % glycerol) に溶かし込み、粗酵素液として用いた。タンパク質濃度の 定量は、Bradford 法により行った。粗酵素液はこの状態で-80 o C で凍結し、実験に使用す る前に流水で融解して使用した。 レコンビナントタンパク質の発現有無を確認するために、調整したタンパク質 (10 µg/lane) は、10 % (w/v) slab gels (e-PAGEL、ATTO Co.) を用いた SDS-PAGE により分離さ れ後、PVDF メンブラン (Amersham) に転写された。 レコンビナントタンパク質の検出は、 mouse monoclonal anti-V5-HRP antibody (Invitrogen) と ECL PrimeWestern Blotting Detection Reagent (Amersham) を用いたウェスタンブロッティングにより行った。
2-2-5
In vitro 酵素
酵素
酵素アッセイ
酵素
アッセイ
アッセイ
アッセイ
前述の方法により酵母から調整した粗酵素液を酵素アッセイに用いた。1.5 mL 用のマ イクロチューブを容器として用い、全容量 200 µL として、50 mM Tris-HCl pH 7.5、1 mM NADPH、100 µM 基質、200 µg のミクロソームタンパク質を含むように調整した。反応は、 30 oC、2 時間行われた。反応生成物は、200 µL の ethyl acetate を加え、抽出した。有機溶 媒層をガラスチューブに移し、20 µL 前後になるまで窒素パージによる濃縮を行った後、 後述する GC-MS 分析に供試した。 各酵素の酵素化学的特性は、それぞれの基質濃度範囲 (α-guaiene ; 2.6-130 µM、 (+)-valencene ; 2-100 µM) で、30 oC、30 分間反応させ、決定した (n = 3)。また、両基質、α-guaiene 及び (+)-valencene が相互に酵素反応を競合阻害するかどうかを調べるために、 両基質をそれぞれ同濃度、80 mM 同時に加え、30 o C、30 分間反応させた (n = 3)。200 ml の methanol を添加することで反応を停止し、10 ng µl-1 になるように(-)-α-cedrene を内部標 準として加えた後、Mili-Q 水を加えて 2 mL まで希釈したものを後述する SBSE 法を用い た GC-MS 分析に供試した。酵素化学的特性は、Hanes-Woolf plot から算出した。
2-2-6
GC-MS 分析条件
分析条件
分析条件
分析条件
酵素反応生成物を直接酢酸エチルで抽出したサンプルは、Agilent 5973 insert mass selective detector (MSD) が連結された Agilent 6890 Series GC system を用いて分析された。
分析条件
カラム:BP20 カラム (50 m x 0.22 mm x 0.25 µm、Trajan Scientific and Medical) カラム流量:コンスタントフロー 1.1 mL min-1
(Helium) オーブン:40 o
C (2 min) - 15 oC min-1 - 130 oC (0 min) - 10 oC min-1 - 260 oC (5 min) インターフェース温度:260 oC
イオン化モード:electron impact ionization (70 eV)
測定モード:スキャンモード (定性) 、スキャンレンジ;m/z 40 - 300
2-2-7
Stir bar sorptive extraction (SBSE) 法
法
法
法
SBSE 法は、1-2-4 に記述した方法で実施した。
2-2-8
SBSE 法
法
法を
法
を
を用
を
用いた
用
用
いた
いた GC-MS 分析条件
いた
分析条件
分析条件
分析条件
酵素反応生成物を SBSE 法で抽出したサンプルは、Triple-Axis Detector を備えた Agilent 5975C inert XL MSD、加熱脱着ユニット (TDU、GESTEL GmbH)、クールドインジ ェクションシステム (CIS4、GESTEL GmbH) が連結された Agilent 7890A Series GC system を用いて分析された。
分析条件
カラム:BP20 カラム (50 m x 0.22 mm x 0.25 µm、Trajan Scientific and Medical) カラム流量:コンスタントフロー 1.1 mL min-1
(Helium) TDU 条件:40 o
C (0.5 min) - 200 oC min-1 - 300 oC (3 min) CIS4 条件:10 o
C (0 min) - 720 oC min-1 - 250 oC (1 min) 脱着流量:50 mL min-1
(Nitrogen)
注入モード:スプリットレスモード (スプリットレス時間:2 min) オーブン:40 o
C (2 min) - 15 oC min-1 - 130 oC (0 min) - 10 oC min-1 - 260 oC (5 min) インターフェース温度:260 oC
イオン化モード:electron impact ionization (70 eV)
測定モード:スキャンモード (定性) 、スキャンレンジ;m/z 40 - 300
2-2-9
SBSE 法
法
法を
法
を
を用
を
用いた
用
用
いた
いた GC-MS/MS 分析条件
いた
分析条件
分析条件
分析条件
ブドウ組織中のα-guaiene 及び rotundone の定量は SBSE を用い、Agilent 7000 GC/MS Triple Quad system、TDU、CIS4 が連結された Agilent 7890A Series GC system によって行 った。
分析条件
カラム:BP20 カラム (50 m x 0.22 mm x 0.25 µm、Trajan Scientific and Medical) カラム流量:コンスタントフロー 1.1 mL min-1
(Helium) TDU 条件:40 o
C (0.5 min) - 200 oC min-1 - 300 oC (3 min) CIS4 条件:10 o
C (0 min) - 720 oC min-1 - 250 oC (1 min) 脱着流量:50 mL min-1
(Nitrogen)
注入モード:スプリットレスモード (スプリットレス時間:2 min) オーブン:40 o
C (2 min) - 15 oC min-1 - 130 oC (0 min) - 10 oC min-1 - 260 oC (5 min) インターフェース温度:260 oC
イオン化モード:electron impact ionization (70 eV) コリジョンセル内流量:1.5 mL min-1
クエンチ流量:2.25 mL min-1
(Helium)
測定モード:Multiple reaction monitoring (MRM) mode トランジション:α-guaiene; m/z 204 → 147
rotundone; m/z 218 → 163
2
H5-rotundone; m/z 223 → 166 (内部標準)
コリジョンエナジー:α-guaiene; 3 eV、rotundone; 8 eV、2H5-rotundone; 7 eV
2-2-10
定量的
定量的
定量的
定量的リアルタイム
リアルタイム
リアルタイム
リアルタイム RT-PCR 分析
分析
分析
分析
長野県上田市で栽培された‘Syrah’及び‘Merlot’果実の外果皮或いは中果皮から CTAB 法により、total RNA を抽出した (Chang et al. 1993)。抽出された total RNA は、RNase-Free DNase set (Qiagen) を用いて処理した後、RNeasy Plant Mini kit (Qiagen) を用いて精製し、 定量的リアルタイム RT-PCR のテンプレートとして用いた。定量的リアルタイム RT-PCR 分析は、One-Step SYBR PrimeScript PLUS RT-PCR kit (Takara-Bio Inc.) を使い、ABI Prism 7300 realtime PCR system (Life Technologies Inc.) で行った。特異的プライマーの設計には、 Primer Express 1.0 software (Life Technologies Inc.) を用いた。VvSTO2 向けにプライマー13 及び 14、18S rRNA (Gen Bank accession no. AF207053) 向けにプライマー15 及び 16 をそれ ぞれ設計した (Table 2-1)。定量的リアルタイム RT-PCR は 3 反復実施され、全てのサンプ ルは 18S rRNA のデータを内部標準として標準化された。
2-2-11
ブドウ
ブドウ
ブドウ
ブドウ組織
組織
組織
組織からの
からの
からのα
からの
α-guaiene 及
α
α
及
及
及び
び
び
び rotundone 抽出
抽出
抽出
抽出
ブドウ組織からのα-guaiene 及び rotundone 抽出は、1-2-3 に記載した方法をわずかに 修正加え、実施した。果実組織は、液体窒素を用いて冷凍された後、ミル (MM400、Retsch) を用いて粉砕した。1 g (生重量) の粉末を計量し、50 µL の2 H5-rotundone (50 µg L -1 ) を内 部標準として添加した。次に 5 mL n-pentane:ethyl acetate (9:1、v/v)を加え、室温で 1 時間振 盪機にかけて抽出した。有機溶媒相を 3000 rpm、10 分間遠心し、ガラスチューブに移した 後、窒素ガスパージにより有機溶媒を除去した。ガラスチューブ底に残った残留物に 500µL の ethanol を添加し、溶かし込んだ後、4.5 mL の酒石酸 buffer (pH 3.2) を加え、SBSE 法 を用いた GC-MS/MS の分析サンプルとした。
2-2-12
分子
分子
分子
分子系統解析
系統解析
系統解析
系統解析
Vitis vinifera CYP71BE ファミリーに属するタンパク質及び Gen Bank データベースに 登録されたテルペノイドを修飾する CYP のアミノ酸シークエンスをもとに、系統解析を行 った。ClustalW program (Thompson et al. 1994) によるアライメントを行った後、MEGA 6.06 software により、neighbor-joining 法に基づいて系統樹を作成した (Tamura et al. 2013)。ブー ストラップ値は繰り返し数 1000 で計算した。
2-3
結果
結果
結果
結果
2-3-1
‘
‘
‘
‘Syrah’
’
’
’果実
果実
果実
果実からのセスキテルペンオキシダーゼ
からのセスキテルペンオキシダーゼ遺伝子
からのセスキテルペンオキシダーゼ
からのセスキテルペンオキシダーゼ
遺伝子
遺伝子の
遺伝子
の
の単離
の
単離
単離
単離
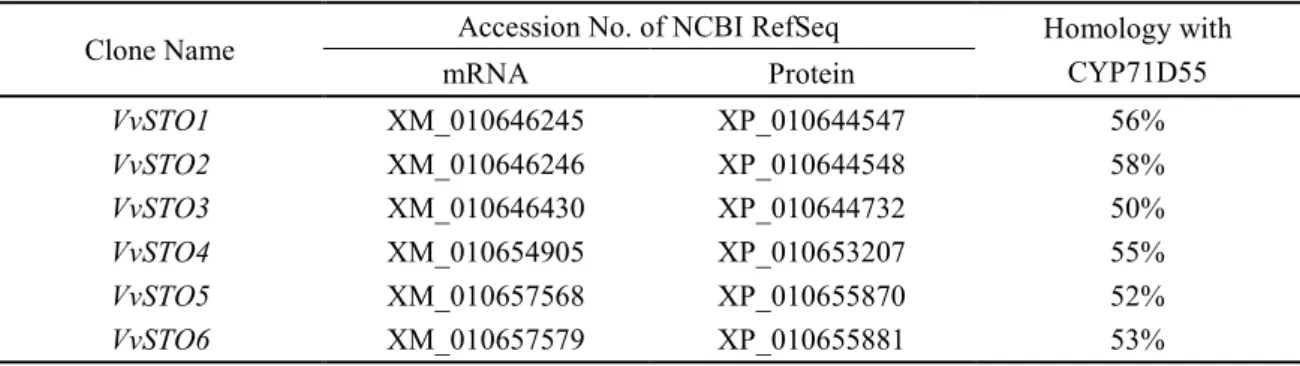
CYP に属し、さらに CYP71D サブファミリーに属する premnaspirodiene oxygenase CYP71D55 は、premnaspirodiene (2 位のカーボン部位) から solavetivone までの連続的は酸 化反応を触媒することができる。また、(+)-valancene (2 位のカーボン部位) からβ-nootkatol への酸化反応も触媒することができる (Takahashi et al. 2007)。特に、CYP71D55 による premnaspirodiene の連続的な酸化反応は、我々のターゲットとするα-guaiene から rotundone への酸化反応と近似している。そこで、α-guaiene の酸化を触媒しうる、ブドウ果実由来の セスキテルペンオキダーゼ遺伝子を単離するために、CYP71D55 の遺伝子配列を基に、 12-fold coverage genome sequence assembly (Pinot Noir PN40024) (Jaillon et al. 2007) から候補 遺伝子を絞り込んだ。その結果、CYP71D55 の遺伝子配列と 50 %以上の相同性を有する 6 つの候補遺伝子が選抜された。Cytochrome P450 Homepage (Nelson 2009) に掲載されている
V.vinifera cytochrome P450 リストと比較した結果、6 つの候補遺伝子は、CYP71BE ファミ
リーに属していた。このファミリーは CYP71D ファミリーと重複して存在している (Cytochrome P450 nomeclature committee か ら の 情 報 ) 。 こ れ ら の 遺 伝 子 を V.vinifera sesquiterpen oxidase、VvSTO1 から VvSTO6 と名付け、其々と合致する 12-fold coverage genome sequence assembly (Pinot Noir PN40024) (Jaillon et al. 2007) 上のレファレンスシークエンス (RefSeq) を一覧にまとめた (Table 2-2)。VvSTO1 から VvSTO6 のうち、3 遺伝子 (VvSTO2、
VvSTO4、VvSTO6) を‘Syrah’外果皮から単離することに成功した。単離された VvSTO2、 VvSTO4、VvSTO6 の cDNA は、それぞれ 1527、1515、1518 bp のオープンリーディングフ
レーム (ORF) を有し、それぞれ 508、504、505 のアミノ酸を有するタンパク質であるこ とが推定された。これら 3 つの遺伝子がコードするタンパク質と CYP71D55 のアミノ酸シ ークエンスのアライメントの結果、上記タンパク質は、真核生物の CYP に共通する保存モ チーフ FxxGxRxCxG (Chapple 1998; Ralston et al. 2001) 及び、推定基質認識部位 (SRS) (Gotoh 1992) を有することを確認した (Fig. 2-1)。VvSTO2、VvSTO4、VvSTO6 は、アミノ 酸シークエンスレベルで CYP71D55 とそれぞれ 53-58 %の相同性を示した。これらは、 P450 nomenclature committee (Nelson 2009) により それぞれ CYP71BE5、 CYP71BE1 、
CYP71BE10 と命名され、DDBJ accession number はそれぞれ LC055499、LC055500、LC055501 が与えられた。
2-3-2
In vitro 酵素
酵素
酵素
酵素アッセイによる
アッセイによる
アッセイによる V.vinifera cytochrome P450 機能解析
アッセイによる
機能解析
機能解析
機能解析
単離した遺伝子の酵素的機能を明らかにするために、それぞれの ORF を含む発現プラ スミドを用いて酵母 BJ2168 株を形質転換した。VvSTO2、VvSTO4、VvSTO6 の発現有無 は、anti-V5-HRP antibody を用いたウェスタンブロッティングにて確認した (Fig. 2-2)。 VvSTO2、VvSTO4、VvSTO6 の C 末端 V5 epitope 融合タンパク質の推定分子量は、それぞ れ 61.4、60.9、61.1 kDa であった。ウェスタンブロッティングの結果、それぞれの融合タ ンパク質の推定分子量と一致した位置に明確なバンドを検出した。 V. vinifera cytochrome P450 を発現させた酵母細胞から調整したミクロソーム画分を用 いて、α-guaiene を基質として用いた in vitro 酵素アッセイを行った。酵素反応生成物は GC-MS 分析された。空の pYES vector を組み込んだ酵母細胞から調整したミクロソーム画 分 (vector control) を用いた酵素アッセイでは、酵素反応生成物は検出されなかった (Fig. 2-3A)。一方、発現したすべての酵素がα-guaiene を基質として用いることができたが、 VvSTO2 が唯一 rotundone を生成した。Rotundone の分子量を示す m/z 218 の extracted-ion chromatogram (EIC) 上においても、VvSTO2 の反応生成物として rotundone の大きなピー ク (peak1) を示し、 そのマススペクトルは有機合成で得た rotundone のものと一致した (Fig. 2-3B-D)。さらに m/z 163 の EIC 上には、rotundone 以外の反応生成物として 2 つの微 小なピークを検出し、そのマススペクトルから (2R)-rotundol 及び (2S)-rotundol と推定した (Huang et al. 2015a) (Fig. 2-3C-E)。また、VvSTO4、VvSTO6 はα-guaiene を基質として複数 の反応生成物を生成した (Fig. 2-4)。これらの生成物は、親イオンとして m/z 220、222 が検 出されたことから、セスキテルペン水酸化物と推定されるが、本研究では同定できなかっ た。In vitro 酵素アッセイにおいてα-guaiene から rotundone への自発的な酸化反応が起こら ないことは、空の pYES vector、VvSTO4 及び VvSTO6 を導入した酵母細胞から調整したミ クロソーム画分を用いた多数の反復試験 (酵素反応時間 2-48 時間) によって確認した。
2-3-3
VvSTO2 の
の
の
の基質特異性及
基質特異性及
基質特異性及
基質特異性及び
び
び
び酵素化学的特性
酵素化学的特性
酵素化学的特性
酵素化学的特性
α-Guaiene から rotundone の合成を触媒した VvSTO2 に関して、その基質特異性及び酵 素化学的特性をさらに調査した。VvSTO2 の酵素活性評価は、潜在的に基質になりえる他 のセスキテルペノイド、モノテルペノイド等を用いた (Fig. 2-5)。その結果、VvSTO2 は α-guaiene だけではなく(+)-valencene も基質としたが、その他セスキテルペノイド、モノテ ルペノイド、C13 ノルイソプレノイドは基質としなかった。(+)-Valencene は、報告されて いた CYP71D55 の機能と同様に、 β-nootkatol に変換された (Takahashi et al. 2007) (Fig. 2-6)。 しかしながら、α-nootkatol や nootkatone などのその他反応生成物は検出されなかった。
α-Guaiene から rotundone、(+)-valencene からβ-nootkatol への変換に対する Km 値は、 Hanes-Woolf plot によって計算され、それぞれ 30 µM、35 µM であった (Fig. 2-7)。これら の数値は(+)-valencene からβ-nootkatol への変換を触媒可能な他のセスキテルペンオキシダ ーゼの数値 (7.4±1.2 µM-52±2 µM) と同等のレベルであった (Gavira et al. 2013)。 α-Guaiene と(+)-valencene を同時に加えた酵素アッセイの結果、それぞれの基質を独立して 加えた酵素アッセイと比較して、rotundone とβ-nootkatol の生成率はそれぞれ 82.7 %、 73.3 %であった。これらの結果は、α-guaiene と(+)-valencene はその程度は比較的小さいが、 お互いの酵素反応に干渉することを示唆した。 α-Guaiene から rotundone への変換について、その最適温度は 30-40 o C、最適 pH は 7.0-8.0 であった (Fig. 2-8)。
2-3-4
果実中
果実中
果実中
果実中の
の
の
のα
α
α-guaiene、
α
、rotundone 蓄積及
、
、
蓄積及
蓄積及
蓄積及び
び
び
び VvSTO mRNA 発現
発現
発現
発現パターン
パターン
パターン
パターン
ブドウ果実中のα-guaiene、rotundone 濃度は、SBSE 法を用いた GC-MS/MS 分析により 定量した。また、VvSTO2 転写レベルはリアルタイム RT-PCR により定量した。過去の研究 では、rotundone はブドウ外果皮に局在することが報告されていた (Caputi et al. 2011)。し かしながら、α-guaiene の局在は明らかではなかった。α-Guaiene、rotundone 濃度、VvSTO2 転写レベルのブドウ果実における組織特異性は、2012 年に収穫した‘Syrah’果実を用い て調査した。‘Syrah’外果皮中のα-guaiene、rotundone 濃度は、内果皮中よりも高かった (Fig. 2-9A、B)。これに一致し、外果皮中の VvSTO2 転写レベルも、内果皮中のレベルよりも顕
著に高かった (Fig. 2-9C)。これらの知見は、セスキテルペン生合成及びその蓄積はブドウ 果実中の外果皮に制限されるという過去の報告を支持した (May et al. 2013)。ブドウ成熟期 間 (開花後 8 から 18 週) における、ブドウ外果皮中のα-guaiene、rotundone 蓄積及び VvSTO2 転写レベルが、rotundone の高い品種である‘Syrah’と低い品種である‘Merlot’の比較を しながら、調査された。同時に、全可溶性固形物、滴定酸度といった一般果実成分分析も 行われた (Fig. 2-10)。‘Syrah’外果皮中のα-guaiene の蓄積は、開花 12 週で最大となり、そ の後減少した (Fig. 2-9D)。興味深いことに、rotundone の蓄積は、α-guaiene の蓄積より 2 週 遅い開花 14 週で最大となり、その後減少した (Fig. 2-9E)。対照的に‘Merlot’外果皮中の α-guaiene 及び rotundone 濃度は成熟期間を通じて常に‘Syrah’外果皮よりも低かった (Fig. 2-9F)。さらに、成熟期間における両品種の外果皮中の VvSTO2 転写レベルは完全に異なり、 rotundone 蓄積パターンと一致した。これらの結果は、VvSTO2 がブドウ果実における rotundone 生合成に関与していることを示唆した。
2-4
考察
考察
考察
考察
Rotundone は、オーストラリア産‘Shiraz’ワインから 2008 年に同定されて以来 (Wood et al. 2008)、ワインが持つスパイシーな特徴に貢献する、最も重要な化合物の 1 つとして 注目されている。Rotundone 蓄積に影響をあたえる環境ファクターやα-guaiene の自発的酸 化による化学的な rotundone 形成は報告されてきたが、植物中の rotundone 生合成機構はこ れまで明らかではなかった。
本章では、rotundone 生合成に関与するキー酵素を同定するために、我々は既に機能が 報告されている sesquiterpene oxidase gene、CYP71D55 のシークエンスをもとに 12-fold coverage genome sequence assembly (Pinot Noir PN40024) (Jaillon et al. 2007) から候補遺伝子 を絞り込んだ。その結果、α-guaiene の 2 位のカーボンを酸化し、rotundone 生合成を触媒 する新規の CYP、α-guaiene 2-oxidase VvSTO2 を同定することに成功した。VvSTO2 は、 CYP71BE5 と命名され、CYP71 族、CYP71 ファミリーの中の CYP71BE サブファミリーに 属した。また、CYP71BE サブファミリーは、CYP71D サブファミリーと重複して存在する。 CYP71 族は、全ての植物 CYP のうち 50 %以上を占めている。これらの CYP は、植物のテ ルペノイド代謝機構の中でモノテルペノイドやセスキテルペノイドの酸化を含む、多種多 様な機能を有することが報告されているが、α-guaiene から rotundone の生成を触媒可能な 植物 CYP はこれまで報告されてこなかった。分子系統解析は、VvSTO2 が他のテルペンノ イドを修飾可能な CYP と密接に関連していることを示した (Fig. 2-11)。
これまで植物の中でセスキテルペンノイドを多段階で酸化可能な sesquiterpene oxidase が複数報告されている。例えば、Artemisia annua 由来の amorphadiene oxidase CYP71AV1 は amorpha-4,11-diene から artemisinic acid の 3 段階での酸化を触媒する (Ro et al. 2006)。
Nicotiana tabacum 由 来 の 5-epi-aristolochene-1,3-dihydroxylase CYP71D20 は 5-epi-aristolochene から capsidiol の 2 段階での酸化を触媒する (Takahashi et al. 2005)。
Hyoscyamus muticus 由来の premnaspirodiene oxygenase CYP71D55 は premnaspirodiene から
solavetivone の 2 段階での酸化を触媒する (Takahashi et al. 2007)。また、Cupressus nootkatensis
由来の valencene oxidase CYP706M1 は(+)-valencene から(+)-nootkatone の 2 段階での酸化を 触媒する (Cankar et al. 2014)。これらの反応には、同時にセスキテルペノイドの水酸化物な ど、中間体が存在することが報告されてきた。過去の報告と同様に、in vitro 酵素アッセイ
の中で VvSTO2 も rotundone を主に生成したが、小さなのピークとして(2R)-rotundol、 (2S)-rotundol と推定される中間体を検出した。これらの結果は、VvSTO2 はα-guaiene から rotundone への酵素的酸化に関与しており、α-guaiene から rotundone への 1 段階での酸化、 或いは (2R)-rotundol、(2S)-rotundol から rotundone への 2 段階目のすばやい酸化反応を介し た 2 段階での酸化を触媒することを示唆した (Fig. 2-12)。さらに本章で立てた rotundone 生合成仮説を証明するためには、(2R)-rotundol、(2S)-rotundol の標準試料を用いた検証が必 要となる。
α-Guaiene 2-oxidase VvSTO2 は、α-guaiene だけではなく(+)-valencene の 2 位のカーボ ンを酸化し、 β-nootkatol を生成することができる。一方で、VvSTO2 は他のセスキテルペ ノイド、モノテルペノイド、C13 ノルイソプレノイドを基質として利用できなかった。こ のことは、α-guaiene 2-oxidase が比較的狭い基質特異性を持つことを示唆した。VvSTO2 は α-guaiene、(+)-valencene に対して低い Km 値を持ち、高い親和性を示した。両基質に対す る Km 値は同水準にあるため、VvSTO2 はブドウの中でα-guaiene 2-oxidase、(+)-valencene oxidase の両方或いはいずれか一方として機能しているかもしれない。しかしながら、本研 究においては‘Syrah’及び‘Merlot’外果皮中に (+)-valencene 及びβ-nootkatol を検出する ことはできなかった。このことは、ブドウ外果皮中に (+)-valencene 及びβ-nootkatol は極微 量のみ含まれている、或いは含まれていないことを示唆した。実際、我々の知る限りでは 両化合物がブドウ外果皮から検出された報告はない。これらのことから、VvSTO2 は、ブ ドウ外果皮においては、(+)-valencene oxidase というよりはむしろ α-guaiene 2-oxidase とし て機能していることが示唆された。
VvSTO4、VvSTO6 もまたα-guaiene を酸化し、未知の化合物を生成した。それら未知 の化合物のうち、2 つは両酵素で共通して生成された (peak 4、5) (Fig. 2-4)。しかしながら、 VvSTO2 とは異なり、rotundone や (2R)-rotundol、(2S)-rotundol を生成しなかった。このよ うな両酵素の共通する酵素的特性は、両酵素間アミノ酸シークエンスの高い相同性 (71 %) に起因するかもしれない。また、これらのことは、VvSTO2 が他の V.vinifera CYP71BE と 比べてユニークな部位特異性を持つことを示唆した。VvSTO4、VvSTO6 と VvSTO2 は、 アミノ酸シークエンスにおいて 61-62 %の相同性を有する。それ故、α-guaiene から rotundone へ修飾は、相同性を示さない、残りのアミノ酸残基の部分に起因していることが 示唆された。過去の報告では、植物 CYP を部位特異的突然変異させた研究において、SRS 領域内のアミノ酸残基が基質選択性、反応生成物の特異性、酵素化学的特性、部位特異性
に影響を与えることを示唆された (Kahn et al. 2001; Komori et al. 2013; Takahashi et al. 2007; Takahashi et al. 2005)。VvSTO2 の基質特異性及び部位特異性を更に理解するためには、部 位特異的突然変異による機能且つ構造的な研究が必要とされる。
定量的リアルタイム RT-PCR 分析の結果、CYP71BE5 の遺伝子発現は、ブドウ果実に おける rotundone の局在パターンと一致して、‘Syrah’外果皮でより高かった (Fig. 2-9A-C)。 また、‘Syrah’外果皮にはα-guaiene も顕著に高い濃度で存在した。これらの結果は、 rotundone の蓄積はα-guaiene の生合成と供に、CYP71BE5 の発現によって制御されている ことを示唆した。また、CYP71BE5 遺伝子は、α-guaiene 及び rotundone の蓄積パターンと 一致して、ブドウ成熟期間中を通して‘Merlot’外果皮より‘Syrah’外果皮で顕著に高く 発現した (Fig. 2-9D-F)。このことから、α-Guaiene 生合成や VvSTO2 発現性の違いによって、 両品種間のブドウ果実中の rotundone 濃度に差が生まれているかもしれない。以上の結果 に基づき、CYP71BE5 は、α-guaiene 2-oxidase としての活性を有する、ブドウ中の rotundone 生合成における鍵酵素であると結論付けた。
α-Guaiene から rotundone への化学的酸化による非酵素的変換が起こることが報告され ているが (Huang et al. 2014)、本研究で行った in vitro 酵素アッセイでは観察されなかった。 これらの事実は、α-guaiene の酵素的変換は、非酵素的変換よりも溶液中ではより速く反応 が進むことを示唆した。それ故、ブドウ果実におけるα-guaiene からの rotundone 生合成は 主に酵素的に起こっている可能性が高い。さらにブドウにおける in vivo での VvSTO2 の機 能を決定するためには、rotundone を含まない、或いは無視できる水準で微量に含むブドウ 組織中で VvSTO2 を過剰発現させる試験 (必要に応じてα-guaiene を基質としてフィード)、 または rotundone を高く含有するブドウ‘Syrah’を用いた VvSTO2 ノックアウト試験によ る検証が必要とされる。
以上、本章では rotundone 生合成経路を介し、α-guaiene から rotundone を生成する α-guaiene 2-oxidase の同定に成功した。VvSTO2 発現解析結果は、VvSTO2 がブドウ果実中 の rotundone 生合成において鍵酵素としての役割を持つことを支持するとともに、rotundone の蓄積が VvSTO2 発現及びその前駆物質であるα-guaiene 生合成によって制御されることを 示唆した。近年、rotundone 蓄積に影響を与える環境ファクター、地域的な特徴に関する研 究が多数進行している。しかしながら、rotundone 生合成機構を制御するマーカー遺伝子に 関する情報が無い状態で、各環境ファクターが及ぼす効果を検証、評価することは困難で あった。今回の研究で発見された遺伝子の情報が活用され、ブドウにおける rotundone 生
2-5
要旨
要旨
要旨
要旨
Rotundone は強力な香気を有する化合物で、ブドウを含む様々な植物中に存在し、特徴 的な胡椒様のアロマを与えている。本研究において、我々はブドウ‘Syrah’からα-guaiene を rotundone に変換するα-guaiene 2-oxidase VvSTO2 を同定した。この酵素は CYP に属し、 さらに CYP71D サブファミリーと重複して存在する CYP71BE サブファミリーに属した。
VvSTO2 はブドウ果実における rotundone の局在に一致し、‘Syrah’外果皮でより高いレベル
で発現した。α-Guaiene もまたブドウ外果皮中に非常に高い濃度で検出した。これらの結果 は、rotundone の蓄積は前駆物質であるα-guaiene の利用可能性及び VvSTO2 の発現によって 制御されていることを示唆した。果実成熟期間における VvSTO2 の発現は、‘Merlot’外果 皮よりも‘Syrah’外果皮でより高く、α-guaiene 及び rotundone の蓄積と一致した。これら の結果に基づき、VvSTO2 はブドウ中の rotundone 生合成機構における鍵酵素と考えられた。
Table 2-1. The specific primers used in this work.
Primer No. Orientation Sequence (5' to 3')
1 F CCTCTCTACTGGTCATCTCTTTTCTCC 2 R GCATTGCATTTCAGATAAATAGCATCG 3 F CAAAGATGGCGCTTGTGCTGAG 4 R CATTACATCACAGCGGCTTCTG 5 F GGTATCTCTTCTAAGAAACCTCTAACA 6 R GCATTGATCCAAATACAAAACACCCA 7 F ATGGAGCTCCAATTCTCCTTCTTCC 8 R TTCAACAGGCAGAGGACGATAAGTAATG 9 F ATGGAGTTTCCCTCTTCTTTCCTC 10 R CCCAACAGTCAATGGACTATAAGG 11 F ATGGAGTTCTTCTCCTCTTCTC 12 R ATCAGCATGCGATGGATTATAAG 13 F CTACTGGTCATCTCTTTTCTCCTC 14 R ACTTCGGAGCAGGATGGTTG 15 F CGAAAGCATTTGCCAAGGAT 16 R CCTGGTCGGCATCGTTTATG