トピ
ックス
171 1. 遺伝子発現量の細胞間多様性 細胞はゲノム DNA から遺伝情報を読み取り,必要 に応じて,必要な場所で,適切な遺伝子を適量発現 させ,遺伝子発現産物である RNA およびタンパク質 を産生する.遺伝子発現産物は細胞の様々な機能を 担っているので,その発現量によって細胞の性質が決 まる. 細菌などの単細胞生物は,細胞間で同じゲノム配列 を持ち,ほぼ同じ環境下でも,細胞間で遺伝子発現産 物量に著しい差があり,異なる環境応答(表現型多様 性)を示すことが知られている.多くの単細胞生物 は,細胞分裂によってほぼ同じゲノム配列を持つ娘細 胞をつくるが,すべての細胞がほとんど同じ表現型を 示す場合,急速な環境変化に耐えられず,死滅する可 能性がある.しかし,表現型に大きな多様性がある場 合,急速な環境変化に対して生き残る細胞が増加する 可能性があることから,単細胞生物の生存戦略として 遺伝子発現量の細胞間多様性が重要と考えられる1). 単細胞生物に比べ,多細胞生物では細胞間に様々な 細胞間相互作用があり,急激かつ極端な遺伝子発現量 の多様性が現れにくいと考えられる.しかし,細胞分 化という不可逆的な表現型の多様性が生み出される過 程で,一過的な遺伝子発現量の多様性が誘引され,そ れによって細胞運命が決まることが知られている2). 遺伝子発現量の多様性(表現型の多様性)が生じる のは,細胞の遺伝子発現過程(転写因子の確率的活性 化,転写因子の結合乖離,RNA の転写,RNA の分解, RNA の輸送,タンパク質の翻訳,分解など)や微小 環境での化学反応の確率性に起因する.さらに,遺伝 子発現制御ネットワーク(転写因子間での発現制御 ネットワーク),ヒストン修飾等のエピジェネティッ クな制御等によって細胞ごとの多様性が固定化され る.多細胞生物における細胞分化はその最たる例であ る.遺伝子発現過程において,特に転写バーストと呼 ばれる現象が遺伝子発現量の細胞間多様性の誘引に寄 与している可能性が示唆されている. 2. 転写バースト 遺伝子発現はDNA(遺伝子)からRNA,タンパク質 へのセントラルドグマを介して起こる(図 1).転写は 細胞内に 1 または 2 コピーしかない遺伝子上で行われ る.また多くの遺伝子では,連続的に転写される ON 状態と,ほとんど転写が行われない OFF 状態が確率 的に切り替わる.この現象を転写バーストと呼ぶ3). 転写バーストの動態はプロモーターの ON 状態の 割合を表すバースト頻度と,1 回の ON 状態での平均 mRNA 発現量を示すバーストサイズで記述できる4). さらに,RNA 分解速度も明らかになれば,細胞内の 平均 RNA 分子数と RNA 発現量の細胞間多様性の大 きさ(後述)を見積もることができる.RNA 分解速 度と平均 mRNA 分子数が同じであれば,バースト頻 度が低くバーストサイズが大きい遺伝子は,その逆の 遺伝子よりも遺伝子発現量の細胞間多様性が大きい (図 2A).すなわち,転写バーストは遺伝子発現量の 細胞間多様性を生み出す要因となりうる.また転写 生物物理 61(3),171-173(2021) DOI: 10.2142/biophys.61.171 受理日:2020 年 11 月 11 日哺乳類細胞における転写バーストの網羅解析:
遺伝子発現量の細胞間多様性の理解に向けて
落合 博
1,2 1広島大学大学院統合生命科学研究科 2広島大学ゲノム編集イノベーションセンターComprehensive Analysis of Transcriptional Bursting in Mammalian Cells
Hiroshi OCHIAI1,2
1Graduate School of Integrated Sciences for Life, Hiroshima University 2Genome Editing Innovation Center, Hiroshima University
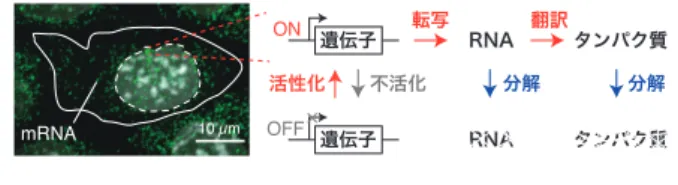
図 1 転写バースト.左パネルは,マウス ES 細胞の特定の RNA 分子を 1 分子 RNA-FISH(smFISH)で緑色に,細胞核を白色で可視化した ものである.ON 状態の遺伝子領域では強い輝点が認められる. 白い実線は細胞膜を示し,白い点線で囲まれた領域は細胞核であ る.右パネルは遺伝子発現の模式図を示す.
哺乳類細胞における転写バーストの網羅解析 172 多様性を示す.NANOG は多能性獲得に重要な因子で あり,NANOG 発現量の高い内部細胞塊細胞は将来エ ピブラストへと分化する.内部細胞塊細胞やマウス ES 細胞における遺伝子発現量の細胞間多様性の誘引 には転写因子間の負のフィードバックや細胞間相互作 用等が関係すると考えられているが,これら制御機構 が作用する以前から発現量多様性が誘引されているこ となどから,詳細は明らかになっていない. 興味深い報告として,Nanog 遺伝子はマウスの初期 発生過程およびマウス ES 細胞で,しばしば 1 つの対 立遺伝子のみから転写されることが示されている7). このことから,Nanog 遺伝子は顕著な転写バーストを 示し,それが NANOG 発現量の細胞間多様性を誘引 する一因である可能性が示唆されていた. 著者のグループは,転写ライブイメージングおよび smFISH 解析によって,マウス ES 細胞において転写 バーストに起因する intrinsic noise が Nanog 遺伝子の発
現量多様性に有意に寄与することを明らかにした8). 一方,マウス ES 細胞で認められる他の多くの遺伝子 の発現量多様性に転写バーストがどの程度寄与してい るのか,また転写バーストがどのように制御されてい るのかは明らかにされていなかった. 4. 1 細胞 RNA-seq による転写バースト動態の解析 上述したように,各対立遺伝子の発現分布がわかれ ば,intrinsic noise を推定でき,さらに mRNA 分解速度 が既知であれば,バーストサイズおよびバースト頻度 も推定できる.そこで著者は,ゲノムワイドに mRNA 分解速度が既知であるマウス ES 細胞に着目した9). さらに,ハイブリッド ES 細胞と呼ばれる,異なるマ ウス系統を交配して得られた個体由来の ES 細胞を用 いて 1 細胞 RNA-seq を実施することで,各遺伝子に ついて対立遺伝子間の発現分布が得られると考えた (図 3A).ハイブリッド ES 細胞では,対立遺伝子間 にゲノム配列多型があるため,その配列の違いによっ て 1 細胞 RNA-seq 解析データから,由来対立遺伝子 を区別して発現量を定量できる.著者のグループは, 約 450 細胞の 1 細胞 RNA-seq 解析を行い,各遺伝子 について,対立遺伝子間の発現量分布を決定すること で,網羅的に intrinsic noise を推定した.また,既知の RNA 分解速度を利用して,バーストサイズ,バース ト頻度も網羅的に推定した.その結果,遺伝子によっ て intrinsic noise,バーストサイズ,バースト頻度が大 きく異なることがわかった(図 3B). 次に,intrinsic noise,バーストサイズ,バースト頻 度の制御機構を調べるために,種々の転写制御関連因 バーストは,同一細胞内にある対立遺伝子間での遺伝 子発現量多様性さえ誘引する(図 2B).このことか ら,転写バーストによって生じる遺伝子発現量多様性 の出現要因(noise)は細胞内部で生じる要因である intrinsic noise に分類される.それ以外の要因(別の遺 伝子の発現量の細胞間多様性によって誘引される遺 伝子発現量の多様性や細胞間相互作用等に起因する 要 因) は extrinsic noise と 呼 ば れ る.intrinsic noise と extrinsic noise は,対立遺伝子間の発現量の分布から見 積もることができる5)(図 2C).遺伝子によってバー ストサイズ,バースト頻度,intrinsic noise は大きく異 なっていることが知られていたが,これらがどのよう な分子機構で制御されているかは明らかにされていな かった. 3. マウス初期発生過程における細胞運命決定 マウスの初期発生過程で,内部細胞塊細胞がエピ ブラストまたは原始内胚葉細胞に分化する.この運 命決定が起こる前に,転写因子 NANOG を含む複数 の因子の発現量は細胞間で大きな多様性を示すこと から,確率的な遺伝子発現が細胞運命を決定し得るこ とが示唆されている6).マウス胚性幹(ES)細胞は内 部細胞塊と似た性質の細胞で,血清存在下で転写因子 NANOG を含む複数の遺伝子が大きな遺伝子発現量の 図 2 転写バーストによる遺伝子発現量の細胞間多様性の誘引.(A) RNA 分解速度を一定にした場合の,バーストサイズ,バースト頻 度,細胞内平均 RNA 分子数,遺伝子発現量の細胞間多様性の関 係.点線は,細胞内の平均 RNA 分子数が同じであることを,点線 の色は細胞内の平均 RNA 分子数の違いを意味する.(B)転写バー ストは,対立遺伝子間での遺伝子発現量の多様性も誘引する. (C)1 細胞レベルでの対立遺伝子ごとの発現量分布から intrinsic noise を見積もることができる.1 つの点は 1 つの細胞における各 対立遺伝子由来の mRNA 数を示す.
哺乳類細胞における転写バーストの網羅解析 173 落合 博(おちあい ひろし) 広島大学大学院統合生命科学研究科准教授 広島大学大学院理学研究科修了.同原爆放射線医 科学研究所,JST さきがけ専任研究員を経て現職. 研究内容:転写バーストの制御機構の解明,転写 バーストが細胞運命決定に与える影響の解明 連絡先:〒 739-0046 広島県東広島市鏡山 3-10-23 イノベーションプラザ 1A01 E-mail: [email protected] URL: https://home.hiroshima-u.ac.jp/ochiai/ 落合 博 5. 今後の展望 網羅的解析から,転写バースト制御機構は遺伝子 ごとに異なることが明らかになった.今後遺伝子ご とに転写バースト制御機構の詳細な分子機構を解明 していく必要がある.今回の解析から,特に遺伝子内 領域での転写伸長因子の局在やプロモーター領域への H3K27me3 およびその調節因子の局在が転写バースト を制御している可能性が示された.これらを含む特定 因子の遺伝子領域への結合と転写との関係を経時的に 明らかにするためにはライブイメージングが有効であ る.著者のグループは,特定の内在遺伝子の転写活性 と,特定転写制御因子のライブイメージング解析を実 施し,特定転写制御因子が転写活性状態の遺伝子近傍 でクラスターを形成していることを見出した10).今 後,この解析を転写伸長因子や H3K27me3 などへ適 用することで,特定遺伝子の転写バースト制御機構の 解明が期待される. 文 献
1) Fraser, D., Kaern, M. (2009) Mol. Microbiol. 71, 1333-1340. DOI: 10.1111/j.1365-2958.2009.06605.x.
2) Chalancon, G. et al. (2012) Trends Genet. 28, 221-232. DOI: 10.1016/j.tig.2012.01.006.
3) Rodriguez, J., Larson, D. R. (2020) Annu. Rev. Biochem. 89, 189-212. DOI: 10.1146/annurev-biochem-011520-105250.
4) So, L.-H. et al. (2011) Nat. Genet. 43, 554-560. DOI: 10.1038/ ng.821.
5) Elowitz, M. B. et al. (2002) Science 297, 1183-1186. DOI: 10.1126/science.1070919.
6) Fiorentino, J. et al. (2020) Annu. Rev. Genet. 54, annurev-genet- 021920-110200. DOI: 10.1146/annurev-genet-021920-110200. 7) Miyanari, Y. et al. (2012) Nature 483, 470-473. DOI: 10.1038/
nature10807.
8) Ochiai, H. et al. (2014) Sci. Rep. 4, 7125. DOI: 10.1038/ srep07125.
9) Ochiai, H. et al. (2020) Sci. Adv. 6, eaaz6699. DOI: 10.1126/sciadv. aaz6699.
10) Li, J. et al. (2019) Cell 178, 491-506.e28. DOI: 10.1016/ j.cell.2019.05.029. 子のプロモーターや遺伝子内領域(gene body)への局 在の程度との相関解析を実施した.その結果,転写伸 長因子が遺伝子内領域に多く局在する遺伝子ほど転写 頻度が高く intrinsic noise が小さいこと,またプロモー ターにヒストン H3 の 27 番目のリジンがトリメチル 化されている状態(H3K27me3)や,それを制御する ポリコーム抑制複合体 2 関連因子が多いほど,バース ト頻度が低く intrinsic noise が大きくなる傾向があるこ とがわかった.しかしさらなる詳細な解析から,転写 伸長因子や H3K27me3 のみで転写バーストの動態が 決まっている訳ではなく,これらとその他の因子の組 み合わせによって,遺伝子ごとに転写バーストの動態 が決定されていることが示唆された(図 3C). 図 3 転写バーストの網羅解析.(A)ハイブリッドマウス ES 細胞の 1 細胞 RNA-seq 解析の模式図.(B)マウス ES 細胞における遺伝子 ごとのバーストサイズ,バースト頻度,intrinsic noise の関係.各 点は特定遺伝子のバーストサイズ,バースト頻度の大きさを示し ている.(C)網羅解析によって明らかになった転写バースト制御 機構.図は広島大学プレスリリースより引用.