コンカナバリン A 誘導性急性肝炎マウスにおける VDR の肝免疫調節作用
日本大学大学院医学研究科博士課程 病理系病態代謝学専攻
梅田 直
修了年 2018 年
指導教員 槇島 誠
目 次
第1章 概 要・・・・・・・・・・・・・・・・・・・・・・・・・ 1
第2章 緒 言・・・・・・・・・・・・・・・・・・・・・・・・・ 2
第3章 実験方法・・・・・・・・・・・・・・・・・・・・・・・・ 7
第4章 実験結果・・・・・・・・・・・・・・・・・・・・・・・・15
第5章 考 察・・・・・・・・・・・・・・・・・・・・・・・・・23
第6章 まとめ・・・・・・・・・・・・・・・・・・・・・・・・・27
謝 辞・・・・・・・・・・・・・・・・・・・・・・・・・・・・・28
表・図・・・・・・・・・・・・・・・・・・・・・・・・・・・・・29
引用文献・・・・・・・・・・・・・・・・・・・・・・・・・・・・52
研究業績・・・・・・・・・・・・・・・・・・・・・・・・・・・・57
略語表
1,25(OH)2D3 1α,25-dihydroxyvitamin D3
α-GalCer α-galactosylceramide AST aspartate aminotransferase ALT alanine aminotransferase
BMDM bone marrow derived macrophages CCL C-C chemokine ligand
cDNA complementary DNA
Con-A concanavalin A
Cy cyanine
CYP2R1 vitamin D 25-hydroxylase CYP24A1 vitamin D 24-hydroxylase CYP27A1 sterol 25-hydroxylase
CYP27B1 25-hydroxyvitamin D 1α-hydroxylase D-Bil direct bilirubin
DNA deoxyribonucleic acid
ELISA enzyme-linked immunosorbent assay FBS fetal bovine serum
FITC fluorescein isothiocyanate FXR farnesoid X receptor
HBSS Hanks' balanced salt solution
IFN interferon
Ig immunoglobulin
IL interleukin
iNOS inducible nitric oxide synthase
iNKT invariant NKT
MNC mononuclear cells
NAFLD non-alcoholic fatty liver disease NF-κB nuclear factor-κB
NK natural killer
NADPH nicotinamide adenine dinucleotide phosphate PBS phosphate buffered saline
PCR polymerase chain reaction
PE phycoerythrin
PXR pregnane X receptor RNA ribonucleic acid
ROS reactive oxygen species
RPMI Roswell Park Memorial Institute medium RXR retinoid X receptor
T-Bil total bilirubin TCR T cell receptor
TGF transforming growth factor TLR toll-like receptor
TNF tumor necrosis factor VDR vitamin D receptor
WT wild-type
第1章 概 要
ビタミンDは脂溶性ビタミンであり、コレステロール合成の中間代謝産物で ある7-デヒドロコレステロールから日光照射による光開裂反応を経て生合成さ れるホルモンである。活性型ビタミンDである 1α,25-ジヒドロキシビタミンD3
は、核内受容体であるビタミンD受容体 (vitamin D receptor, VDR) に作用する ことにより、骨・カルシウム代謝調節や細胞増殖・分化、心血管機能、毛周期、
抗菌・抗炎症作用など多岐にわたる機能を有している。VDRはリガンド依存性 転写因子として腸管や腎臓などの標的臓器において様々な遺伝子の発現制御を 行うが、肝臓における VDR の発現は非常に低いことから、その機能には不明 な点が多く残されていた。近年、VDRは肝細胞ではなく肝常在マクロファージ
である Kupffer 細胞や間葉系星細胞などの非実質細胞で発現することや、ビタ
ミンD投与が胆管結紮による肝障害を抑制することが明らかとなった。しかし、
肝臓における自然免疫応答に対する VDR シグナルの関与についての報告は少 ない。そこで、本研究では野生型または VDR 欠損マウスを用いて急性肝炎を 比較した。まず、野生型マウスの肝非実質細胞において VDR が発現及び機能 していることを確認した。次に、野生型または VDR 欠損マウスにコンカナバ リンAを投与し、急性肝炎を惹起させ経過を観察した。その結果、野生型では 投与後の肝組織において多数の死細胞が観察されたが、VDR欠損での変化は軽 微であった。また、野生型と比較し VDR 欠損マウスの血中トランスアミナー ゼ値増加は軽度であった。一方、血中サイトカイン値を定量したところ、増加 の程度に変化を認めなかった。また、肝単核球を単離し、遺伝子発現解析を行 った結果、血中と同様に両群において顕著なサイトカイン発現を認めた。一方、
肝常在 Kupffer 細胞が産生する活性酸素種は VDR 欠損において有意に減少し
た。以上の結果から、VDR欠損マウスにおいて、サイトカイン産生は正常であ るのに対し肝常在 Kupffer 細胞による活性酸素種産生が抑制されることにより 肝炎が軽減することが明らかとなった。本研究により肝常在 Kupffer 細胞での VDRの機能及び肝炎の病態への関与が示された。
第2章 緒 言
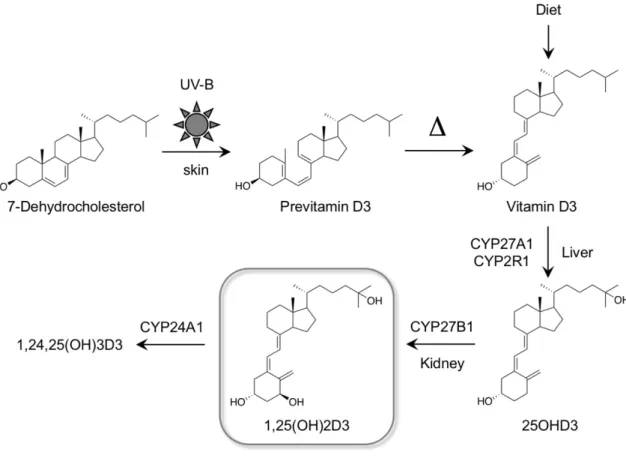
2-1. ビタミンDの生合成と代謝
ビタミン D は脂溶性ビタミンであり、動物由来のビタミン D3と植物由来の ビタミンD2からなる。ビタミンD3は主に魚肉や肝油に、ビタミンD2の前駆体 であるエルゴステロールはシイタケやキクラゲなどのきのこ類に多く含まれて いる。また、ビタミンDは多くの他のビタミンとは異なり体内で合成可能であ
る[1] (図1)。コレステロール生合成経路の中間代謝産物である7-デヒドロコレ
ステロールは皮膚における日光照射によりステロイド骨格のB環が開裂し、プ レビタミン D3となる。さらに体温による熱反応によって速やかにビタミン D3
(コレカルシフェロール) が産生される。産生されたビタミン D3は食事由来の ものも含めて肝臓に運ばれ、シトクローム P450 酵素である CYP27A1 または
CYP2R1によって25位が水酸化され 25-ヒドロキシビタミンD3 (カルシフェジ
オール) となる。さらに腎臓において CYP27B1 により 1α 位が水酸化され、
1α,25-ジヒドロキシビタミン D3 (1,25(OH)2D3, カルシトリオール) が産生され る。1,25(OH)2D3は活性型ビタミンD3として後述のVDRに作用し、多岐にわた る機能を発揮する。また、1,25(OH)2D3は代謝酵素であるCYP24A1による 24位 水酸化を経て速やかに代謝され、最終的にカルシトロン酸となり、尿中へ排泄 される。
2-2. VDRの発現と機能
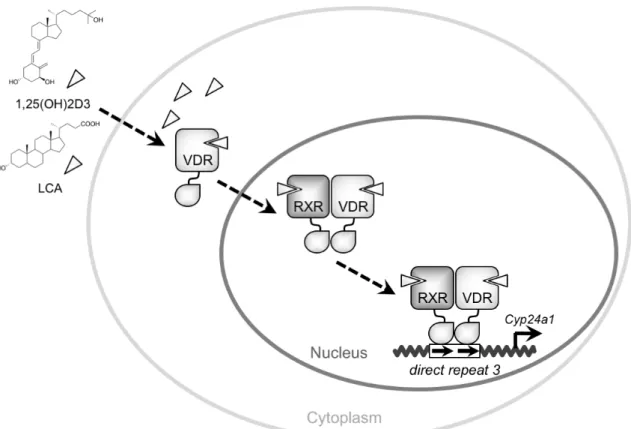
VDRは1,25(OH)2D3の受容体としてクローニングされた核内受容体である[2]。
核内受容体は DNA 結合ドメインとリガンド結合ドメインを有する転写因子で あり、ヒトでは48種類存在することが確認されている[3]。VDRはオキシステ ロール受容体である liver X receptor、胆汁酸受容体である farnesoid X receptor (FXR) および薬物代謝受容体であるpregnane X receptor (PXR) などと同じ核内 受容体サブファミリー1に属し、これらの受容体と同様にレチノイドX受容体
重要なリガンドは1,25(OH)2D3であるが、PXRやFXRなどの類似の受容体の機 能解析が行われた過程において、二次胆汁酸であるリトコール酸 (lithocolic acid,
LCA) が同じく VDR の生体内リガンドとして作用することが明らかとなった
[5]。1,25(OH)2D3や LCA などの VDR リガンドは容易に細胞膜の脂質二重膜を 通過し VDRタンパク質のリガンド結合ドメインに結合する。VDR はコンホメ ーションを変化させ核内へと移行し、RXRとヘテロ二量体を形成する。転写共 役因子群との転写複合体を形成後、標的遺伝子の DNA 上の規則的な応答配列 (ダイレクトリピート3型) に結合し標的遺伝子群の発現を制御する (図2)。 ビタミン Dの最も重要な生理作用はホルモンとしてVDR に作用し、骨・カ ルシウム代謝を制御することである。日照不足などによるビタミンD欠乏は小 児ではくる病を、成人では骨軟化症を惹起することが知られている。1997年に
VDR 欠損 (VDR-KO) マウスが作製され、低カルシウム血症、骨形成障害、成
長障害、脱毛などが観察された[6]。これらの表現型はヒト遺伝性くる病の症状 と類似しており、VDRによるカルシウム代謝制御の重要性を示している。VDR は骨・カルシウム代謝疾患以外にも、細胞増殖・分化、心血管作用、毛周期の 調節、抗菌、抗炎症作用など多岐にわたる機能を有しており、くる病、骨軟化 症だけでなく、骨粗鬆症、悪性腫瘍、動脈硬化、乾癬、自己免疫疾患など様々 な疾患の分子標的とされており、多くのビタミンD製剤が開発されている。
2-3. 肝臓における免疫細胞の機能
肝臓は食事由来の栄養成分や薬物など、さまざまな分子の代謝臓器として機 能するが、同時に門脈を介して腸管から多くの異物を受け付けることから免疫 細胞が豊富に存在するため主要な免疫臓器でもある。肝臓を構成する細胞は約 60〜70 %は肝細胞 (実質細胞) であるが、残りの約 30 %は非実質細胞である。
非実質細胞にはマクロファージである肝常在 Kupffer 細胞やナチュラルキラー (natural killer, NK) 細胞やNKT 細胞、T細胞、B細胞などのリンパ球や間葉系 星細胞、類洞内皮細胞などが含まれる。これらの細胞群は、肝細胞と協調しな
がら外来異物に対する防御、炎症、抗腫瘍活性など、多岐にわたる機能を担っ ており、肝障害の病態に重要な役割を果たしている。
近年、組織特異的マクロファージの fate mapping 解析が行われ、グリア細胞 は胎生期に卵黄嚢で産生された後に運ばれ自己増殖を繰り返す、骨髄造血には 全く依存しない細胞であることが示された[7]。肝臓マクロファージについても 解 析 が 行 わ れ 、 肝 臓 に は 2 種 類 の マ ク ロ フ ァ ー ジ が 存 在 し 、 そ れ ぞ れ F4/80+CD68+と F4/80+CD11b+の細胞表面抗原パターンを示すこと、前者は卵黄 嚢または胎生期肝臓にて産生される肝常在 Kupffer 細胞で放射線照射に対して 耐性であるのに対し、後者は放射線感受性骨髄由来マクロファージ (bone marrow derived macrophages, BMDM) であることが明らかとなった[8, 9]。CD32 が新たな肝臓マクロファージの新しいマーカーであるという報告がされたが [10]、その後の検討により CD32 陽性細胞については類洞内皮細胞が混在する 可能性が示されたため、現在は上述の F4/80+CD68+とF4/80+CD11b+の2種類の マーカーを用いて肝常在Kupffer細胞と BMDMを定義付けている。また、これ ら 2 種類の肝臓マクロファージの機能が比較され、肝常在 Kupffer 細胞は貪食 能 や 活 性 酸 素 種 (reactive oxygen species, ROS) 産 生 に 特 化 す る の に 対 し 、 BMDMは高い炎症性サイトカイン産生能を有することが明らかとなった[8, 10]。 マウスを用いた肝炎モデルの解析は様々な刺激分子の投与により検討が行わ れている。四塩化炭素投与モデルは、肝臓における間葉系星細胞の活性化を惹 起し、肝線維化を誘導する[11]。また、コリン欠乏アミノ酸置換食付加による肝 炎モデルは肝細胞への中性脂肪蓄積を誘発する非アルコール性肝炎モデルであ る[12]。コンカナバリン A (concanavalin A, Con-A) はナタマメ由来の糖脂質で あり、マウスへの尾静脈投与により自己免疫性急性肝炎を発症することが報告 された[13]。ヒトの自己免疫性肝炎と異なり、投与後数時間で劇症肝炎を発症 し、24時間後には収束する急性肝炎のモデルであるため、ヒトの病態を再現す ることは困難であるが、肝臓免疫細胞の機能解析のためにマウスを用いた検討 で頻繁に用いられる肝炎モデルである。Con-A 肝炎の発症には様々な肝免疫細
を産生できないヌードマウスにおいては肝炎耐性であることから[14, 15]、 T細 胞の関与が示唆された。その後、NKT細胞が機能しないCD1d欠損マウスにお いて肝炎が起こらないこと[16]、抗 NK1.1 抗体投与で肝障害は軽減するが抗ア シアロ GM1 抗体では変化しないことから[17]、NKT 細胞が肝炎発症に必須で あることが明らかとなった。また、様々な中和抗体の投与や欠損マウスの解析 から、インターフェロン (interferon, IFN) –γなどの炎症性サイトカイン産生や inducible nitric oxide synthase (iNOS) の発現誘導が必須であることも報告された
[18-20]。さらに、エダラボンなどのフリーラジカルスカベンジャーの投与によ
り肝障害が抑制されることから ROS 産生も肝炎発症に必須であること、また
Con-A誘導性ROS産生は肝常在Kupffer細胞によって行われることが明らかと
なった[21]。以上の報告から、Con-A肝炎の発症には少なくとも (1) NKT細胞 の活性化、(2) IFN-γなどの炎症性サイトカイン産生、(3) 肝常在Kupffer細胞に よるROS産生が必要であることがわかっている。
ビタミン D と肝障害については、ビタミン D欠乏が C 型肝炎に対するイン ターフェロン療法の感受性低下に関与すること、非アルコール性脂肪性肝疾患 (non-alcoholic fatty liver disease, NAFLD) の病態を増悪させること、一方でビタ
ミンDの補充がNAFLDや肝線維化を抑制するなど多数の疫学的な報告がなさ
れており、ビタミンDの肝機能に対する重要性は様々な検証によって示されて いる[22]。これらの作用は受容体であるVDRを介していると考えられるが、同 じNR1サブファミリーに属するFXRやPXRなどの核内受容体が肝臓に高発現 し脂質代謝や薬物代謝制御因子として働くのに対し、VDRは肝臓での発現が非 常に低いことから[23]、肝臓におけるVDRの機能はまだ十分に解明されていな い。近年、ラットを用いた検討により、肝細胞においてはVDRの発現は非常に 低いものの、肝常在 Kupffer 細胞、類洞内皮細胞、間葉系星細胞などの非実質 細胞に発現を認めることが明らかとなった[24]。また、マウスにおける胆管結 紮による胆汁鬱滞の病態において1α-ヒドロキシビタミンD3投与が炎症性サイ トカイン産生を抑制すること[25]、間葉系星細胞において VDR は transforming
growth factor (TGF)-β/Smad経路をエピジェネティックに抑制するメカニズムで
肝線維化を抑制すること[11]などが報告されたが、肝臓自然免疫制御における
Kupffer細胞などの免疫細胞に関する検討は未だ報告されていない。そこで本研
究では、肝臓免疫細胞における VDR の選択機能を見出すため、野生型 (wild-
type, WT) および VDR-KO マウスにおける免疫細胞の分布と機能解析および
Con-A投与による急性肝炎の病態の比較を行った。
第3章 実験方法
3-1. マウスの飼育及び繁殖
WT (C57BL/6J) マウスは日本クレア株式会社 (Tokyo, Japan) より購入した。
VDR-KO (Vdr-/-) マウスは加藤茂明博士の承諾を得て中外製薬株式会社より供
与して頂いた[6]。VDR-KOマウスはC57BL/6Jマウスと8回以上バッククロス させたのちに繁殖に用いた。自然交配による繁殖が困難であるため、雄及び雌 Vdr+/-マウスを自然交配させることによって得た。生まれたマウスは遺伝子型を 判別する必要があるため、ケージ内での個体識別のため耳にパンチで穴を開け、
尻尾を 1 cm 程度カットし、DNA 抽出を行った後、PCR 反応を行い遺伝子型
(+/-または-/-) を判別した。遺伝子型判定は Vdr または Neo カセット遺伝子を
検出することにより決定した。ジェノタイピング用PCR反応に用いたプライマ ーは表1に示した。マウスは室温23±1 °C 及び湿度45–65 %の制御下で飼育し、
水及び飼料 (CE-2; 日本クレア株式会社, 2 %水酸化カルシウム、1.5 %リン、20 % ラクトースを付加) は自由摂取させた[26, 27]。実験には8〜12週齢の雄マウス を用い、炭酸ガスで安楽死させた後、各組織を摘出した。全てのマウス実験は 文部科学省もしくは日本大学医学部の定めた動物実験指針に従い行った。
3-2. 使用試薬
1,25(OH)2D3 は和光純薬株式会社 (Osaka, Japan) から購入した。Con-A は Vector Laboratories, Inc. (Burlingame, CA) から、α-ガラクトシルセラミド (α- galactosylceramide, α-GalCer) はフナコシ株式会社 (Tokyo, Japan) から購入した。
3-3. マウス肝臓由来単核球 (mononuclear cells, MNC) の単離
マウス肝常在 Kupffer 細胞を含む MNC は以下に示す方法で単離した。マウ スを炭酸ガス吸引により安楽死させ、後大静脈より全採血を行った。肝臓を摘 出後、胆嚢や結合組織を除去し、1×phosphate bufferd saline (PBS) にて洗浄し た。肝臓はハサミを用いて細切し、50 mL チューブに移し、1 %ウシ胎児血清
(fetal bovine serum, FBS) を含むRPMI培地 (和光純薬株式会社) (以下1 % FBS- RPMI培地) 15 mLに懸濁した。1,500 rpmで5分間遠心後、上清を吸引し肝臓1 gあたり20 mLのHBSS緩衝液 (Thermo Fisher Scientific, Waltham, MA) に懸濁 した。1 M塩化カルシウム溶液、1 M塩化マグネシウム溶液、1 M硫酸マグネ シウム溶液をHBSS 溶液20 mLに対し25 µL、さらにPBSを用いて25 mg/mL に調製したコラゲナーゼ溶液 (和光純薬株式会社) をHBSS溶液 20 mLに対し
200 µL加えた。37 ℃に設定した恒温槽にて22分間インキュベートさせ、肝組
織を分散させた。コラゲナーゼ反応後、1 % FBS-RPMI培地を HBSS 溶液と等 量加え、1,500 rpmにて5分間遠心した。培地を吸引後、再度1 % FBS-RPMI培
地を10 mL加え、1,500 rpmにて5分間遠心し培地を吸引した。ペレットは1 %
FBS-RPMI培地50 mL を加えて懸濁し、10 cm ディッシュ上に載せた金属メッ
シュを用いて裏ごしした。再び 50 mLチューブに移し、1,500 rpm にて 5分間 遠心した。培地を吸引後、以下に示す組成で調製した33 % Percoll溶液を25 mL 加え懸濁した。Percoll溶液の調製は、まず、Percoll® (pH8.5-9.5, 20 ℃) (Sigma- Aldrich, St. Louis, MO) 92.5 mLに対し10×PBS 7.2 mL、10 %炭酸水素ナトリウム 溶液2.4 mL加えたものを100 % Percoll溶液とし、ストック溶液とした。次に 100 % Percoll溶液33 mLにRPMI培地67 mLおよびノボ・ヘパリン注 (持田製 薬株式会社, Tokyo, Japan) 500 µLを加え33 % Percoll溶液とし、実験に用いた。
肝組織は33 % Percoll溶液に懸濁後、2,500 rpmにて30 分間遠心し、肝細胞と MNC を含む非実質細胞とを分離した。遠心後、上清を全て吸引し、Red blood cell lysis solution (Sigma-Aldrich) を2 mL加え、再び1 % FBS-RPMI培地50 mL で懸濁した。1,500 rpm で 5 分間遠心後、培地を吸引し赤血球を完全に除去し た。最後に1 % FBS-RPMI培地10 mLを加え懸濁後、1,500 rpmで5分間遠心 後、培地を吸引し1×PBSで懸濁した。得られた細胞はTC20™全自動セルカウ ンター (Bio-Rad Laboratories, Hercules, CA) を用いてカウントし、各実験に用い た。
3-4. 細胞及びマウス組織からの総RNA抽出、逆転写反応によるcDNA合成 マウス各組織または肝MNC からの総RNA 抽出は以下に示すようにRNAiso plus (タカラバイオ株式会社, Shiga, Japan) を用いて行った。凍結したマウス各
臓器は約5 mm片にRNAiso plus溶液1.4 mLを加え、ホモジェナイザーを用い
て氷冷下で破砕した。破砕液1.26 mL を 2 mL チューブに移し、クロロホルム 0.52 mLを加え混合した。10分間氷上で静置後、15,000 rpm、4 ℃で15分間遠 心した。水層0.6 mLを1.5 mLチューブに移し等量のイソプロパノールを加え 氷上で15分間静置した。15,000 rpm、4 ℃で10分間遠心し、上清を除去後75 % エタノールを0.75 mL加え再び15,000 rpm、4 ℃で10分間遠心した。上清を完 全に除去し15分間風乾後、超純水を50 µL加えRNAを溶解した。肝由来MNC
由来総 RNAは、1,500 gで5分間遠心して得られた細胞のペレットから組織と
同様の方法でRNAiso plusを用いて抽出した。得られた総RNA液は、NanoDrop (Thermo Fisher Scientific) を用いて測定し、濃度を算出した。総RNA抽出液は 超純水を用いて250 ng/µLに希釈した。
cDNA の合成は ImProm-II Reverse Transcription system (Promega Corporation, Madison, WI) を用いて行った。500 ngの総RNA (250 ng/µLを2 µL) に10 µM oligo dT プライマー 0.5 µL と超純水を 2.5 µL 加え、5 µL の反応液とし、
GeneAmp® PCR System 9700 (Thermo Fisher Scientific) を用いて70 ℃で5分間 加温し、4 ℃で5分間冷却した。この反応液にImProm-II 5×Reaction buffer 2 µL、 25 mM塩化マグネシウム1.2 µL、10 mM deoxynucleotide triphosphates mixture 0.5 µL、ImProm-II Transcriptase 0.5 µL及び超純水0.8 µLの混合液を加え10 µLの反 応液を調製した。逆転写反応はGeneAmp® PCR System 9700 を用いて25 ℃で 15分間加温後、42 ℃で60分間伸長反応を行った。その後70 ℃で15分間加温 することで酵素を失活させ、4 ℃に冷却した。得られたcDNA溶液10 µL にTris-
EDTA緩衝液を20 µL加えて3倍希釈し、-20 ℃で保存した。
3-5. リアルタイムPCR
マウス肝MNCまたは組織から抽出した総RNAを用いて合成したcDNA 2 µL にPower SYBR Green PCR Master Mix (Thermo Fisher Scientific) 7.5 µL、forward primer (10 µM) 0.3 µL、reverse primer (10 µM) 0.3 µL及び超純水を加えて反応溶 液を調製した。反応に用いたマウス各プライマーの配列はイントロンを挟む配 列で設計し、表2に示した。リアルタイムPCR反応はStepOnePlusTM Real-time PCR System (Thermo Fisher Scientific) を用いて次のように行った。まず、95 ℃ で 10 分間反応させた。次に 94 ℃で 15 秒間熱変性させ、アニール及び伸長反
応は60 ℃、1分間で行い、2ステップを1サイクルとし40サイクル行った。発
現量の計算は逆転写させた総 RNA 1 ng あたりのコピー数を算出することによ って行った。内部標準として Cyclophilin B (Gene symbol: Ppi6) の値を用いて mRNA発現量を補正した。
3-6. フローサイトメトリー
各肝臓免疫細胞の分布については以下の方法を用いて実施した。免疫細胞を同 定するために用いた抗体の詳細は表3に示した。
3-6-1. 肝常在 Kupffer細胞およびBMDMの染色
マウス肝臓からコラゲナーゼ処理を行って単離したMNC を1.5mLチューブ に移し3,000 rpmで2分間遠心し、上清を除去後、1×stein buffer (2 % FBSおよ び0.1 %アジ化ナトリウムを含む1×PBS) 1 mL を加え懸濁し、再び3,000 rpm で2分間遠心した。再度1×stein buffer 1 mLで細胞を洗浄後、抗マウスCD16/32 Fc blocker (Thermo Fisher Scientific) を 10 µL 加え氷上で 15 分間静置した。
fluorescein isothiocyanate (FITC)標 識 抗 マ ウ ス F4/80 抗 体 (Thermo Fisher Scientific) 、phycoerythrin (PE)標 識 抗 マ ウ ス CD11b 抗 体 (Thermo Fisher Scientific) および biotin標識抗マウスCD68抗体 (Bio-Rad Laboratories) 10 µLを 加え再び氷上で15分間静置した。1×stein buffer 1 mLで細胞を2回洗浄後、PE 標識ストレプトアビジン (Thermo Fisher Scientific) を10 µL加え氷上で15分間
Coulter, Indianapolis, IN) を用いて細胞分布を測定した。
3-6-2. NK細胞またはNKT細胞の染色
コラゲナーゼ反応を行わずに単離した肝MNCを1×stein buffer 1 mL で2回 洗浄後、抗マウス CD16/32 Fc blocker を 10 µL 加え氷上で 15 分間静置した。
FITC標識抗マウスαβ-T cell receptor (TCR) (Thermo Fisher Scientific)、PE標識抗 マウス NK1.1 (Thermo Fisher Scientific) および biotin 標識抗マウス CD69 抗体 (Thermo Fisher Scientific) 10 µLを加え再び氷上で15分間静置した。1×stein buffer 1 mL で細胞を 2 回洗浄後、PE-Cy5 標識ストレプトアビジン (Thermo Fisher Scientific) を10 µL加え氷上で15分間静置した。1×stein buffer 1 mLで細胞を 2回洗浄後、Cytomics FC500を用いて細胞分布を測定した。
3-6-3. インバリアントNKT (invariant NKT, iNKT) 細胞の染色
染色を行う前日に、α-GalCer (0.2 mg/mL) 3 µLとDimerX I: Recombinant Soluble Dimeric Mouse CD1d:Ig Fusion Protein (BD Biosciences, San Jose, CA) 8 µLを混合 し37 ℃で18時間反応させた。18時間後、CD1d-α-GalCer complexにPE標識抗 ラットIgG1 (BD Biosciences) 11 µL を加え室温遮光下で60分間反応させた。コ ラゲナーゼ反応を行わずに単離した肝MNCを1×stein buffer 1 mL で2回洗浄 後、抗マウス CD16/32 Fc blockerを10 µL加え氷上で15分間静置した。PE標 識CD1d-α-GalCer-IgG1抗体複合体を1.5 µLおよびPE-Cy5標識抗マウスαβTCR、
FITC標識抗マウスNK1.1抗体 (Thermo Fisher Scientific) を各10 µLを加え再び 氷上で 15 分間静置した。1×stein buffer 1 mL で細胞を 2 回洗浄後、Cytomics
FC500を用いて細胞分布を測定した。
3-6-4. B細胞の染色
コラゲナーゼ反応を行わずに単離した肝MNCを1×stein buffer 1 mL で2回 洗浄後、抗マウス CD16/32 Fc blocker を 10 µL 加え氷上で 15 分間静置した。
FITC 標識抗マウスIgM (Thermo Fisher Scientific) および PE-Cy5標識抗マウス
B220 抗体 (Thermo Fisher Scientific) を各10 µL加え、再び氷上で 15分間静置 した。1×stein buffer 1 mLで細胞を2回洗浄後、Cytomics FC500を用いて細胞 分布を測定した。
3-7. マウスへの薬剤投与
1,25(OH)2D3 (0.1 mM、エタノール溶液) はエタノールで2倍に希釈後、50 µL
に対し 1×PBS を 1 mL 加えて溶解した。コントロール (1×PBS) または
1,25(OH)2D3をマウス体重20 gあたり200 µL腹腔内投与し、10 nmol/kg体重と した。約14時間後、もう一度同量腹腔内投与し2時間後に炭酸ガスを用いて安 楽死させ、腎臓および肝臓を摘出した。
Con-A は 1×PBSを用いて 2.5 mg/mLに溶解し、ストック溶液とした。マウ
ス投与前に1×PBSを用いて1.25 mg/mLに調製し、投与に用いた。マウス体重
20 gあたり200 µLを尾静脈投与し、最終投与濃度を12.5 mg/kg体重とした。血
中トランスアミナーゼ値およびサイトカイン値測定のため、投与前 (0 時間)、
Con-A投与1、3、6、12、24、48、72時間後に経時採血を行った。投与72時間
後、炭酸ガス吸引によってマウスを安楽死させた。肝組織観察およびROS測定 のため、マウスにコントロール (1×PBS) または Con-A 投与 24 時間後、炭酸 ガス吸引によってマウスを安楽死させ、後大静脈より全採血を行い肝臓を摘出 した。遺伝子発現解析を行うため、マウスにコントロール (1×PBS) または
Con-A 投与6時間後、炭酸ガス吸引によってマウスを安楽死させ、後大静脈か
らの全採血および肝臓を摘出した。
α-GalCer (1 mg/mL、ジメチルスルホキシド溶液) は65 ℃に加温した1×PBS を用いて 0.01 mg/mL に溶解し、マウス体重 20 g あたり 200 µL を尾静脈投与 し、最終投与濃度を0.1 mg/kg体重とした。血中サイトカイン値測定のため、投 与前 (0時間)、α-GalCer投与1、3、6、12、24、48、72時間後に採血を行った。
投与72時間後、炭酸ガス吸引によってマウスを安楽死させた。
採血した血液は 3,000 g にて 10 分間遠心分離し、血漿を得て実験に用いた。
3-8. 肝MNCにおけるin vivo貪食能評価
FITC標識マイクロスフェアビーズ (Fluoresbrite® YG Microspheres, Calibration Grade 3.00 µm) (Polysciences, Inc., Warrington, PA) を1×PBSで10倍希釈し、マ
ウス体重20 gあたり200 µL尾静脈投与した。10分後、マウスを炭酸ガス吸引
により安楽死させ、肝 MNC をコラゼナーゼ処理を用いる方法で単離した。単 離したMNCを1×stein buffer 1 mLで2回洗浄後、抗マウスCD16/32 Fc blocker (Thermo Fisher Scientific) を10 µL加え氷上で15分間静置した。PE標識抗マウ ス CD11b抗体 (Thermo Fisher Scientific) および PE-Cy5標識抗マウスF4/80抗 体 (Thermo Fisher Scientific) 10 µLを加え再び氷上で15分間静置した。1×stein buffer 1 mLで細胞を2回洗浄後、Cytomics FC500 (Beckman Coulter, Indianapolis, IN) を用いて貪食細胞分布を測定した。
3-9. 血中成分分析
マウス血中アスパラギン酸アミノトランスフェラーゼ (aspartate aminotrans- ferase, AST) ま た は ア ラ ニ ンアミノトランスフェラーゼ (alanine aminotrans-
ferase, ALT) 値はトランスアミナーゼ CII-テストワコー (和光純薬株式会社)
を用いて測定した。血中総ビリルビン (total bilirubin, T-Bil) 値または直接 (抱 合型) ビリルビン (direct bilirubin, D-Bil) 値はビリルビンBIIテストワコー (和 光純薬株式会社) を用いて測定した。
血 中 イ ン タ ー ロ イ キ ン (interleukin, IL)-4 ま た は IFN-γ 値 は そ れ ぞ れ Quantikine® ELISA Mouse IL-4 Immunoassay (R&D Systems, Inc., Minneapolis, MN)、
Quantikine® ELISA Mouse IFN-γ Immunoassay (R&D Systems, Inc.) を用いて測定 した。
3-10. 肝組織のヘマトキシリン及びエオシン染色
マウスより摘出した肝組織は10 %中性緩衝ホルマリン液 (武藤化学株式会社, Tokyo, Japan) を用いて 24時間固定した。固定組織サンプルは 1×PBS で 2回 洗浄後、50 %エタノール溶液中にて保存した。パラフィン包埋は自動固定包埋
装置を用いて行い、50 %エタノール (4時間)、60 %エタノール (2時間)、70 % エタノール (2時間)、90 %エタノール (2時間)、95 %エタノール (2時間)、100 % エタノール (2 時間)、キシレン (2 時間)、キシレン (1 時間)、キシレン (1 時 間)、パラフィン (1時間)、パラフィン (2時間)、パラフィン (2時間) のプログ ラムで行った。包埋した肝組織を用いてパラフィンブロックを作製し、切片は ミクロトームを用いて 5 µm の厚さで切り出した。切り出した切片はキシレン 15分間を3回、100 %エタノール (10分)、70 %エタノール (5分)、50 %エタノ ール (5 分)、30 %エタノール (5 分)、超純水 (5 分) の順で脱パラフィン処理 し、ヘマトキシリンおよびエオシンを用いて染色した。染色後、ヒストファイ ン (株式会社ニチレイバイオサイエンス, Tokyo, Japan) を用いて封入し、光学 顕微鏡にて観察した。
3-11. 血中および肝臓 ROS量の測定
マウスより摘出し凍結した肝臓は40 mg/mLとなるように1×PBSに懸濁し、
ホモジェナイザーを用いて氷冷下で 30 秒間破砕し、2 回繰り返した。全量を
1.5mL チューブに移し、10,000 gで 5 分間遠心し、得られた上清を実験に用い
た。血漿または肝臓破砕液中の総ROS量はOxiSelectTM In Vitro ROS/RNS Assay Kit (Green Fluorescence) (Cell Biolabs, Inc., San Diego, CA) を用いて行った。肝臓 破砕液については内部標準として総タンパク質量をPierce™ BCA Protein Assay Kit (Thermo Fisher Scientific) によって測定し、得られたROS量を補正した。
3-12. 統計処理
全ての実験結果は平均値±標準誤差として示した。有意差検定は Prism 6.0h (GraphPad Software, Inc., La Jolla, CA) を用いて行った。二群間の比較にはTwo- tailed Student’s-t検定を用い、多群間比較にはTwo-way ANOVAまたはOne-way ANOVA followed by Tukey’s multiple comparison法を用いて算出した。
第4章 実験結果
4-1. 肝MNCにおけるVDRの発現と機能
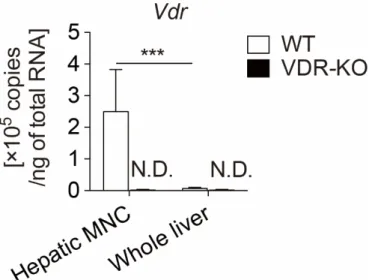
まず始めに肝 MNC における VDR の発現を検討した。WT または VDR-KO マウスより肝臓を摘出し、一部を残し肝 MNC を単離した。本研究で用いた単 離方法により、肝MNC中には少なくとも肝常在Kupffer細胞、BMDM、NK細 胞、NKT細胞、T細胞およびB細胞が含まれることを確認している。総肝臓ま たは肝MNCよりRNAを抽出し、リアルタイムPCRを用いてVdrのmRNA量
(総RNA 1 ngあたりのコピー数) を定量した。その結果、肝MNCにおけるVdr
のmRNA発現量は2.5×105コピーであるのに対し、肝細胞が60-70%を占める
総肝臓は6.8×103コピーであり、肝MNCで約36倍発現量が高いことが示され
た (図3)。同時にVDR-KOマウス由来の検体ではVdrの発現は検出されないこ
とを確認した。
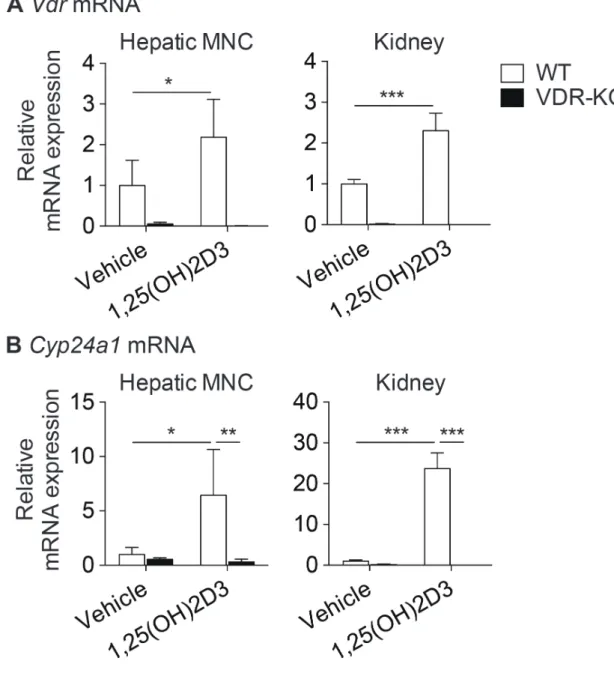
次に、肝MNCにおけるVDR標的遺伝子の発現を調べるために、WTまたは
VDR-KOマウスにVDRの天然リガンドである1,25(OH)2D3を腹腔内投与した。
14 時間後、再度投与し2時間後にマウスを安楽死させ、肝臓およびVDRの代 表的な標的臓器である腎臓をコントロールと用いるために摘出した。肝臓より 単離した肝 MNC または腎臓における Vdr および VDR の標的遺伝子である
Cyp24a1発現をリアルタイムPCR法によって解析した。その結果、肝MNCお
よび腎臓において1,25(OH)2D3投与によってVdrの発現が増加すること、VDR- KO マウスにおける発現は非常に低いバックグラウンドレベルであることを確 認した (図4A)。また、WT マウスの肝MNCにおいて1,25(OH)2D3投与により Cyp24a1発現が約 6.4倍誘導された (図4B)。腎臓においては約23 倍の発現誘 導を認めたことから、肝MNCは腎臓よりも低いものの、同様にVDR標的遺伝 子発現を誘導することを確認した。VDR-KOマウスでは発現誘導されないこと からVDR依存的な現象であることが示された。
以上の結果より、マウス肝MNCにおいてVDRは十分に発現しており、転写 因子としての機能を保持することが示された (表4)。
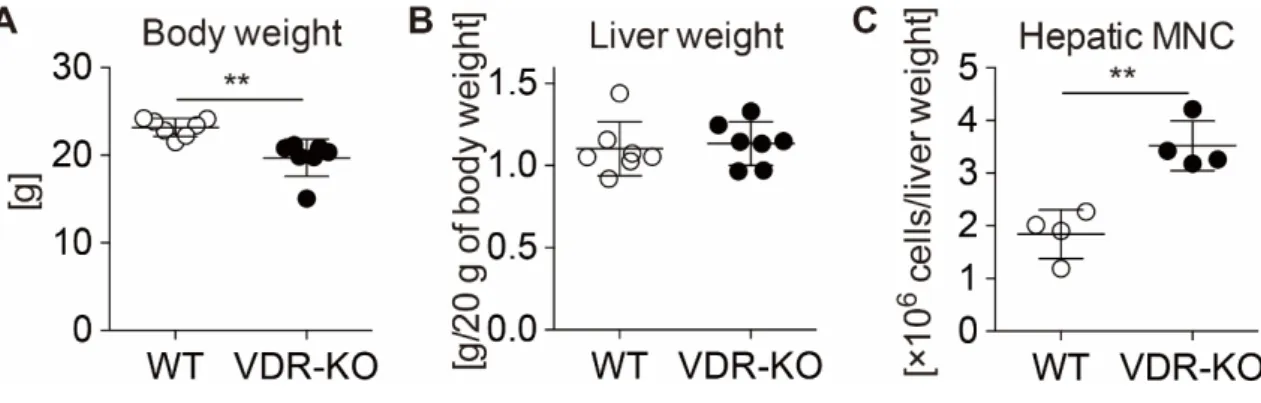
4-2. VDR-KOマウスの表現型
WT または VDR-KO マウスの表現型を比較した。過去の報告と一致し[6]、
VDR-KO マウスは加齢とともに脱毛し同週齢のマウスの体重は減少すること
を確認した (図 5A)。一方、体重 20 g あたりの肝重量は差を認めなかった (図
5B)。次に、WTまたはVDR-KOマウスから摘出した肝臓より肝MNCを単離し
た。肝重量あたりの肝MNC数の比較を行ったところ、WTマウス (1.8 ± 0.5× 106個) に対しVDR-KOマウス (3.5 ± 0.5×106個) において1.9倍増加していた (図5C)。
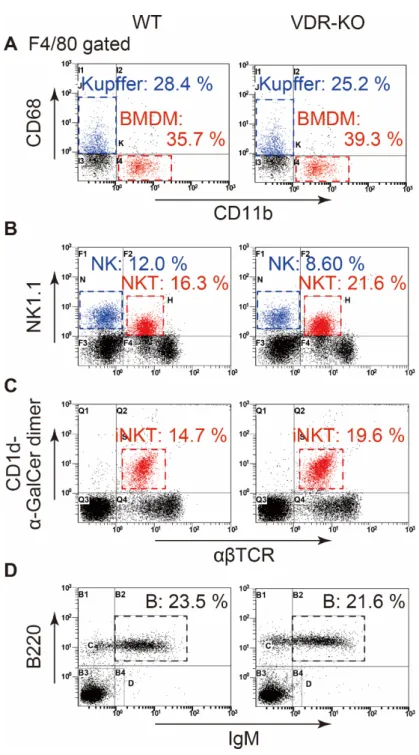
4-3. WTまたはVDR-KOマウスにおける肝免疫細胞分布
図 5C において、WT と比較しVDR-KOマウス由来総肝 MNC 数が増加して いたため、フローサイトメトリーを用いて肝 MNC に含まれる代表的な免疫細 胞の分布を評価した。まず、肝常在 Kupffer 細胞または BMDM を評価するた
め、WT または VDR-KO マウスより肝臓を摘出し、コラゲナーゼを用いて肝
MNCを単離した。肝常在Kupffer細胞を単離するためにはコラゲナーゼ反応を 用 い た 肝 臓 組 織 の 分 散 化 が 必 要 で あ る 。 肝 常 在 Kupffer 細 胞 は F4/80+CD68+CD11b-、BMDM は F4/80+CD11b+CD68-の分画で検出可能であるた め、F4/80、CD68およびCD11bの抗体を用いて染色した。その結果、両群にお いて同程度の分布を認め、差を認めなかった (図6A)。次にNK細胞またはNKT 細胞の比較を行った。NKT 細胞については VDR-KO マウスの肝臓や脾臓にお いて細胞分布や機能が低下することが報告されている[28]。WTまたはVDR-KO マウスより肝 MNC をコラゲナーゼ反応を行わずに単離し、αβTCR、NK1.1、
CD69の抗体を用いて染色した。その結果、NK細胞 (αβTCR-NK1.1+細胞) また は NKT 細胞 (αβTCR+NK1.1+細胞) の分布についてもマクロファージと同様に 両群で同程度の細胞分布を認め、NKT細胞については過去の報告と異なる結果 を得た (図6B)。リンパ球の活性化マーカーであるCD69発現についてもNK細 胞、NKT細胞ともに差を認めなかった。さらに Type I NKT細胞であるiNKT細
WT、VDR-KOともに同程度の細胞分布を認めた (図6C)。最後にB220および IgM抗体を用いてB細胞を染色したが、NK細胞や NKT細胞と同様に変化を認 めなかった (図 6D)。以上の結果から、VDR-KO マウスにおいて総 MNC 数は 増加していたが、ある種の免疫細胞が特異的に増加している現象は見出されな かった。また、無刺激の状態では肝組織は正常であったことから (図 10)、肝 MNCの分布はWTと同様に正常であると判断した。
4-4. WTおよびVDR-KOマウスにおけるin vivo貪食能評価
VDR-KO マウスにおける肝常在 Kupffer細胞の機能を調べるため、代表的な
機能である貪食能についてin vivoで検討した。WTまたはVDR-KOマウスに蛍 光ビーズを投与し10分後に肝MNCを単離後、フローサイトメトリーを用いて ビーズを貪食した免疫細胞数を評価した。その結果、まずWTマウスにおいて 肝常在 Kupffer細胞の 43 %が蛍光ビーズを貪食した (図7A)。VDR-KOマウス において貪食能はわずかではあるが有意に減少した (図7A、C)。一方、過去の 報告と一致し[8]、WTのBMDMにおけるビーズの貪食能は肝常在Kupffer細胞 と比較し約1.5 %と非常に低く、2種類の肝マクロファージにおける貪食能は肝 常在Kupffer細胞が優位であることが示された (図7B、C)。また、VDR-KOマ ウスのBMDMにおける貪食能は WTと変化を認めなかった。さらに、VDR-KO マウスの好中球 (F4/80-CD11b+細胞) においても貪食能は有意に減少した (図
7C)。以上の結果より、VDR-KOマウスにおいて肝常在Kupffer細胞または好中
球などの食細胞の機能低下が起こっていることが示された。
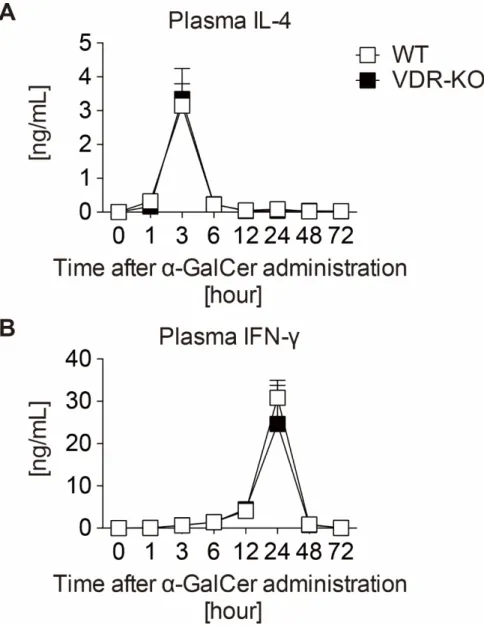
4-5. WT または VDR-KO マウスにおける NKT 細胞特異的リガンド投与の効
果
次に、NKT 細胞の機能を直接評価するために、iNKT 細胞特異的投与の効果 を比較した。α-GalCerは海綿Agelas mauritianusから単離された糖脂質であり、
抗原提示細胞の CD1d 分子に結合し iNKT 細胞が持つ特定の TCR (ヒトでは
Vα24 Vβ11、マウスではVα鎖はVα14、Vβ鎖はVβ8.2、Vβ7またはVβ2) に提 示されることで iNKT を特異的にかつ強力に活性化することが知られている
[29]。WTまたはVDR-KOマウスにα-GalCerを尾静脈投与し、各マウスより経
時採血を行い、得られた血漿を用いてIL-4またはIFN-γ濃度をそれぞれELISA 法を用いて測定した。WT および VDR-KOマウスにおいて血中 IL-4 濃度は α-
GalCer 投与後 3 時間に一過性に増加し、一方で IFN-γ 濃度は 24 時間後に増加
した (図8)。WT、VDR-KO間で差を認めなかった。
以上の結果および細胞分布の検討から (図6B、C)、VDR-KOマウスの肝臓に おいて NKT 細胞は WT と同程度に存在し、正常に機能していることが示され た。
以上、WTとVDR-KOマウスにおける肝 MNCの分布および機能を比較した結
果、VDR-KOマウスにおいて、総肝MNCが増加していたこと、免疫細胞分布
に変化を認めないこと、肝常在 Kupffer 細胞における貪食能が減弱していたこ と、NKT細胞リガンド投与によるサイトカイン産生能に変化を認めないことが 明らかとなった (表5)。
4-6. WTまたはVDR-KOマウスにおけるCon-A誘導性肝障害の比較
肝障害に対する VDR の影響を調べるため、WT または VDR-KOマウスにお
けるCon-A誘導性肝炎の病態を比較した。Con-Aはマウスへの尾静脈投与によ
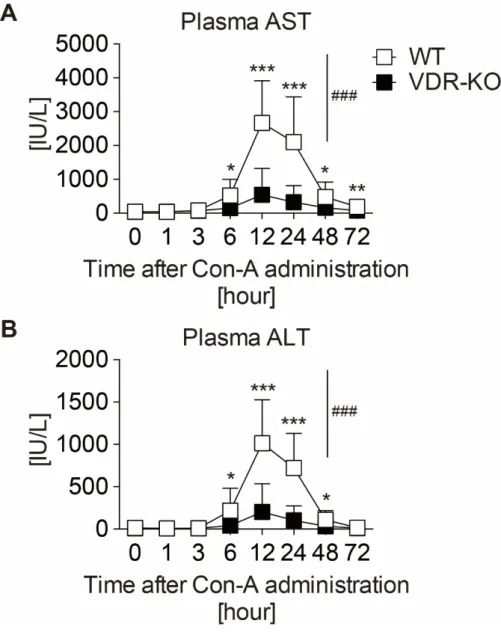
り急性肝炎を発症し、約 12〜24 時間後肝障害は最大となり、48 時間後には収 束することから[21]、WTまたは VDR-KOマウスにCon-Aを投与後、各マウス より経時採血を行い、得られた血漿を用いて肝逸脱酵素であるASTまたはALT 値をそれぞれ測定した。その結果、WTではCon-A投与6時間後から一過性に ASTおよびALT値の顕著な増加を認め、12時間後に最大となり、48時間後に 正常程度まで減少した (図9A、B)。一方、VDR-KOマウスにおいても一過性ト ランスアミナーゼ値増加を認めたが、予想に反しその増加はWTと比較し有意 に減少した。
次に、WT または VDR-KO マウスにおける Con-A 投与後の肝組織を比較す
るためにCon-A 投与 24時間後に肝臓を摘出し、ホルマリンで固定後、パラフ
ィン切片を作製しヘマトキシリンまたはエオシンを用いて染色した。その結果、
WT の肝臓において投与 24 時間後に多数のネクローシス誘導肝細胞が観察さ れた (図 10)。一方、VDR-KO マウスの肝臓において、肝細胞死は WT と比較 し軽微であり、血中ASTおよびALT値と同様の傾向が観察された。
4-7. WTまたはVDR-KOマウスにおけるCon-A刺激後のサイトカイン産生量
の比較
VDR-KO マウスにおける Con-A 誘導性肝障害軽減メカニズムを明らかにす
るため、まず始めに WT または VDR-KOマウスにおける Con-A 誘導性血中サ イトカイン濃度を測定した。Con-A 投与により NKT 細胞や T 細胞が活性化さ れ、IL -4やIFN-γなどのサイトカインが産生される[17, 21]。そこで、WTまた
は VDR-KOマウスにCon-A を尾静脈投与後、各マウスより経時採血を行い、
得 ら れ た 血 漿 を 用 い て IL-4 ま た は IFN-γ 濃 度 を そ れ ぞ れ enzyme-linked immunosorbent assay (ELISA) 法を用いて測定した。その結果、血中IL-4濃度は
WT、VDR-KOマウスともに Con-A 投与 1 時間後に顕著な増加を認め、6 時間
後には投与前と同程度まで減少した (図11A)。Con-A投与3時間後のIL-4濃度
はVDR-KOにおいて有意に減少したが、1時間後の最大濃度についてはWT、
VDR-KO間で差を認めなかった。IFN-γ濃度は WT、VDR-KOともにCon-A 投
与1時間後から増加を認め、12時間後に最大に達し、24時間後には正常程度に 減少した (図 11B)。投与 1、3、24時間後のIFN-γ濃度はVDR-KOマウスにお いて有意に減少したが、IL-4 と同様に 12 時間後の最大濃度については差を認 めなかった。
次に、WT またはVDR-KOマウスにコントロール (PBS) または Con-A投与 し、6 時間後の総肝臓または肝 MNC におけるサイトカインなどの炎症メディ エーター遺伝子の発現をリアルタイム PCRを用いて評価した。WTマウスを用 いて予備検討に行い、Con-A投与6時間後がtumour necrosis factor (TNF)-α (Gene
symbol: Tnf)、IFN-γ (Gene symbol: Ifng) などの炎症性サイトカインおよびiNOS
(Gene symbol: Nos2)の顕著な発現増加の検出に最適な時間であることを確認し
た。WTとVDR-KOマウスにおけるこれらの発現を比較したところ、VDR-KO
マウスにおいて肝障害は軽減していたにも関わらず、Con-A 誘導性Tnf、Ifngお
よび Nos2の発現はVDR-KOマウスでむしろ増加した (図 12A)。次に、Vdrま
たは 1,25(OH)2D3合成に必須なビタミン D-1α-水酸化酵素である CYP27B1 (1α- hydroxylase) (Gene symbol: Cyp27b1) の発現を評価した。単球、マクロファージ において、Vdrおよび Cyp27b1は結核菌感染などによるToll様受容体 (toll-like
receptor, TLR) シグナルの活性化によって発現増加し、カテリシジンなどの抗
菌ペプチドを誘導することが報告されている[30]。同様にWTマウスの肝MNC において、Con-A刺激によりVdrの発現は約11倍の、Cyp27b1についても約19 倍の顕著な発現増加を認めた (図12B)。VDR-KOマウスにおいても、WTと比 較して低いものの Con-A 投与によりVDR 非依存的に9.3倍の Cyp27b1発現増 加を認めた。以上の結果から、VDR-KOマウスの肝MNCにおいてもCon-A投 与によるTLRシグナルは活性化され、WTと同程度もしくはそれ以上のサイト カイン産生能を有することが示された。
また、肝臓においてTLR刺激によりケモカインであるC-C chemokine ligand (CCL)2 (Mcp-1, Gene symbol: Ccl2) の発現が増加し、骨髄から肝臓への単球、マ クロファージの動員を誘導することにより肝障害が増悪することが知られてい る[31]。我々の検討により、肝障害時のマウス肝臓におけるCcl2発現は肝MNC ではなく肝細胞であり、総肝臓において誘導が認められる知見を得ている。そ こで、肝細胞におけるCcl2発現を解析したいと考えたが、肝 MNCを単離する 際に肝細胞を同時に分離することは不可能である。全肝臓は MNC を含有して いるものの70 %が肝細胞であることから、肝細胞での発現を想定し、Con-A投 与 6 時間後の総肝臓におけるサイトカインまたは Ccl2 遺伝子の発現について もリアルタイムPCRで評価した。その結果、まずTnfまたはIfng発現は肝MNC における発現と同様に、WTと同程度または発現増加を認めた (図13)。Ccl2に
ついても同様に、VDR-KO マウスにおいて Con-A 投与により顕著に増加し、
WTとVDR-KO間での差は認めなかった。
以上の結果から、VDR-KO マウスは Con-A 刺激に対し肝障害を軽減させる が、サイトカイン産生能は有していることが示された。NKT細胞がCon-A刺激 初期のサイトカイン産生を担当することから、NKT細胞はWTと同様に機能し ていることが示唆された。
4-8. WT および VDR-KO マウスにおける Con-A 刺激後の血中または肝臓中
ROS産生の比較
これまでの検討より、VDR-KO マウスにおいて NKT 細胞は正常に機能して おり、サイトカイン産生がCon-A肝炎軽減の原因にはならないことが示された。
そこで、次にCon-A誘導性のROS産生について検討した。WTマウスを用いた 予備検討において、Con-A 投与24時間後が血中または肝臓中ROS 産生能を評 価するのに最適な時間であることを見出した。WT または VDR-KO マウスに
Con-Aを尾静脈投与し、24時間後に各マウスを安楽死させ、全採血後、肝臓を
摘出した。まず、血中ASTおよびALT値を評価し、これまでの結果 (図9) と
同様に VDR-KO において肝障害マーカーが顕著に減少していることを確認し
た (図14A)。また、血中AST、ALT値と同様に、Con-A 投与によって増加する 総ビリルビンおよび直接ビリルビン値についても VDR-KO マウスにおいて減 少していることを確認した (図14B)。以上、肝障害が軽減した検体を用いて血 中または肝臓破砕液中の総 ROS量を測定した。その結果、WTと比較しVDR- KO マウスの血中または肝臓において Con-A 誘導性 ROS 産生が有意に減少し た (図15A、B)。Con-A刺激によるROS産生は肝常在 Kupffer細胞が担うこと から[21]、VDR-KOマウスにおいて肝常在Kupffer細胞によるROS 産生が減少 することにより肝障害が軽減すると考えられた。