C
TIN
O
M
Y
C
E
T
O
L
O
G
IC
201 5 VOL. 29 NO. 2 ACTINOMYCETOLOGI CAVOL. 29
NO. 2
1990年12月18日 第4種郵便物認可ISSN 0914-5818
http://www. actino.jp/
Published by
The Society for Actinomycetes Japan
日本放線菌学会誌 第28 巻
1
号
ACTINOMYCETOLOGICA VOL.28 NO.1, 2014
誌会 学 菌 線 放 本 日 A B C D
(
201
5
公開
SAJ NEWS
Vol. 29, No. 2, 2015
Contents
Outline of SAJ: Activities and Membership
S 2
Congratulatory message to Professor Satoshi Omura, a Nobel prize laureate
in Physiology or Medicine
S 3
Preface of Dr. Satoshi Omura
S 4
Publications contributed by Dr. Satoshi Omura in Actinomycetologica
S 8
Award Lecture
S 10
Publication of Award Lecture
S 18
58th Regular Colloquim
S 19
The 2015 Annual Meeting of the Society for Actinomycetes Japan (Program)
S 20
The 2016 Annual Meeting of the Society for Actinomycetes Japan
S 28
Outline of SAJ: Activities and Membership
The Society for Actinomycetes Japan (SAJ)
was established in 1955 and authorized as a scientific organization by Science Council of Japan in 1985. The Society for Applied Genetics of Actinomycetes, which was established in 1972, merged in SAJ in 1990. SAJ aims at promoting actinomycete researches as well as social and scientific exchanges between members domestically and internationally. The Activities of
SAJ have included annual and regular scientific
meetings, workshops and publications of The Journal of Antibiotics (the official journal, joint publication with Japan Antibiotics Research Association), Actinomycetologica (Newsletter) and laboratory manuals. Contributions to International Streptomyces Project (ISP) and International Symposium on Biology of Actinomycetes (ISBA) have also been SAJ's activities. In addition, SAJ have occasional special projects such as the publication of books related to actinomycetes: “Atlas of Actinomycetes, 1997”, “Identification Manual of Actinomycetes, 2001” and “Digital Atlas of Actinomycetes, 2002” (http://www.actino.jp/DigitalAtlas/). These activities have been planned and organized by the board of directors with association of executive committees consisting of active members who belong to academic and nonacademic organizations.
The SAJ Memberships comprise active
members, student members, supporting members and honorary members. Currently (as
of Mar. 31, 2012), SAJ has about 413 active members including student members, 22 oversea members, 11 honorary members, 5 oversea honorary members, 1 special member and 12 supporting members. The SAJ members are allowed to join the scientific and social meetings or projects (regular and specific) of SAJ on a membership basis and to browse The Journal of Antibiotics from a link on the SAJ website and
will receive each issue of Actinomycetologica, currently published in June and December. Actinomycete researchers in foreign countries are welcome to join SAJ. For application of SAJ membership, please contact the SAJ secretariat (see below). Annual membership fees are currently 5,000 yen for active members, 3,000 yen for student members and 20,000 yen or more for supporting members (mainly companies), provided that the fees may be changed without advance announcement.
The current members (April 2014 - March 2016) of the Board of Directors are: Hiroyuki Osada (Chairperson; RIKEN), Haruo Ikeda (Vice Chairperson; Kitasato Univ.), Tomohiko Tamura (Secretary General; NITE), Takayuki Kajiura (Ajinomoto Co., Inc.), Kenji Ueda (Nihon Univ.), Yojiro Anzai (Toho Univ.), Koji Ichinose (Musashino Univ.), Jun Ishikawa (NIID), Takashi Sakai (Eisai Co., Ltd.), Kenji Arakawa (Hiroshima Univ.), Tohru Dairi (Hokkaido Univ.), Hiroyasu Onaka (Tokyo Univ.), Masahiro Sota (Nagase & Co., Ltd.), and Takuji Kudo (RIKEN).
The members of the Advisory Board are: Yoko Takahashi, Kunimoto Hotta, Kozo Ochi, Yuzuru Mikami, and Akira Yokota.
Copyright: The copyright of the articles published in Actinomycetologica is transferred from the authors to the publisher, The Society for Actinomycetes Japan, upon acceptance of the manuscript.
The SAJ Secretariat
c/o Culture Collection Division, Biological Resource Center,
National Institute of Technology and Evaluation (NBRC) 2-5-8, Kazusakamatari, Kisarazu, Chiba 292-0818, Japan Phone: +81-438-20-5763 Fax: +81-438-52-2329 E-mail: [email protected]
Congratulatory message to Professor Satoshi Omura, a Nobel prize
laureate in Physiology or Medicine
Hiroyuki Osada (Chairperson of the Society for Actinomycete Japan)
Dr. Satoshi Omura, an honorary member of the Society for Actinomycetes Japan (SAJ) and an emeritus
professor of Kitasato University, was nominated as a recipient of the Nobel Prize in Physiology or
Medicine 2015. On behalf of all the SAJ members, I extend my healthful congratulations on his winning
Nobel Prize.
Dr. Omura and his collaborators discovered new compounds more than 480, since he was appointed as
a head of Antibiotics Laboratory of Kitasato Institute, 1973.
Among these compounds, 26 compounds have been put on market as pharmaceutical drugs, pesticides or
reagents. In particular, anti-parasitic drugs, avermectin and ivermectin, were developed by the collaboration
with Merck & Co. Ivermectin and showed the excellent efficacy against onchocerciasis and lymphatic
system filariasis. Ivermectin has been distributed to the people living in the tropical and sub-tropical
countries of Africa and South America by the aid of the World Health Organization (WHO). It is expected
that these infectious diseases will be eradicated by the drug in near future.
Dr. Omura has long been aiming at the research which gave benefit to the people, and he always says to
students and young researchers that the research should be "useful for society" and "not mimicry of other
people". I admire his clear vision and a strong leadership. Moreover, I appreciate the great achievement of
Dr. Omura and his co-workers.
I wish Dr. Omura's good health in spite of his busy schedule. I hope that Dr. Omura is continuously
Preface of Dr. Satoshi Omura
Satoshi Omura was born in Yamanashi Prefecture, Japan, in 1935. He received an M.S. degree in 1963 from Tokyo University of Science, followed by a Ph.D. in Pharmaceutical Sciences from the University of
Tokyo in 1968, and the second Ph.D. in Chemistry from the Tokyo University of Science in 1970. He
started his research career as a Research Associate at Yamanashi University from 1963. After 2 two years,
he joined the Kitasato institute in 1965, initially as a researcher, over the years occupying various posts,
and culminating on his appointment in 1984 as Vice-President and then in 1990 as President. During the
period, he was as Visiting-Professor of Wesleyan University (USA) from 1971 to 1975 and as Professor of
School of Pharmaceutical Sciences, Kitasato University from 1975 to 1984. He served as President
Emeritus of the Kitasato Institute from 2008 to 2012. He is currently a Distinguished Emeritus Professor.
He was also appointed as inaugural Max Tishler Professor of Chemistry at Wesleyan University (USA) in
2005.
For 50 years, the drug discovery group at the Kitasato Institute led by him has continued advanced and
pioneering research based on the profound belief that organic compounds produced by microorganisms
have immeasurable promise for use in improving the health of mankind. He discovered more than 480
novel organic compounds possessing interesting chemical structures and/or biological activities, including
many widely-used antibiotics. Among them, 26 compounds and/or their derivatives have entered into
widespread common usage as medicines, agents to improve animal health and husbandry, as agrochemicals
and as reagents for biochemical research. The following are some representative compounds to illustrate
the breadth and scope of his discoveries. Cerulenin, produced by a fungus, Acremonium caerulensm, with
the isolation, structure elucidation and research on the mode of action being carried out by him and his
colleague, was the first inhibitor of fatty acid biosynthesis. It became the lead compound for development
of the medically important statins, inhibitors of cholesterol biosynthesis. The compound remains a crucial
reagent for research in the biosynthesis of polyketide compounds. Because cerulenin is also specific
inhibitor of polyketide synthesis on both type-I and type-II polyketide synthases. Furthermore, he applied
to construct the hybrid macrolide, “Chimeramycins”, by ‘hybrid biosynthesis’ using cerulenin.
Staurosporine, identified through a search for alkaloids of microbial origin, was found to inhibit protein
kinase C, this observation occurring over 10 years after he discovered the compound. During that time, the
discovery of staurosporine stimulated development of novel anti-cancer substances, all based on the
structure and bioactivity of staurosporine. Staurosporine is said to be the most widely-used biochemical
reagent originating from a naturally-occurring microorganism, with the number of scientific papers based
Ivermectin, a 22,23-dihydro-derivative of avermectin, producing by Streptomyces avermitilis, was
discovered and developed by collaborative research with the Merck Shape & Dohme Corp. (USA) as an
anthelmintic macrocyclic-lactone. It was made available on the animal health market in 1981. Two years
later, it became the biggest selling drug in animal health and the position was maintained for over 20 years.
This compound was later found to be highly safe and effective anti-nematode agent for treating several
human diseases. The eradication programs for two devastating tropical diseases, Onchocerciasis and
Lymphatic filariasis, which afflict around 1 in 7 of the entire world population, being orchestrated by the
World Health Organization (WHO), are based primarily on the use of Ivermectin, which is being donated
by the Merck Sharp & Dohme Corp. and the Kitasato Institute. The drug is being administered to almost
300 million people annually in some of the world’s poorest and remotest of communities. It is envisaged
that Onchcerciasis will be eliminated globally by 2025 and Lymphatic filariasis by 2020. With respect to
Onchocerciasis, disease elimination has virtually been completed in Latin America and is well on the way
to success in Africa. S. avermitilis, discovered by him and his co-workers, is the only avermectin-producing
organism ever found and that a single organism has been the sole source of industrial production ever since.
It is recognized that the discovery and use of ivermectin represents the greatest public health intervention of
the last quarter of the 20th century and can be regarded as on a par with the discovery and use of penicillin.
He has also built up an excellent body of research findings on the relationships between chemical
structure, biological activity and mode of action of many macrolide antibiotics, such as leucomycin, tylosin,
spiramycin and erythromycin. Among of them, rokitamycin, a derivative of leucomycin coupled with
tilmicosin, a derivative of tylosin, have been used with great effect as human medicins and in animal health,
respectively. Overall, the natural products from microbial origin which have appeared as a result of his
initiatives and creative endeavors have been used worldwide as medicines, agrochemicals, and reagents for
biochemical research, and have greatly contributed to progress in the health of mankind, as well as being
essential elements in making major steps forward in biomedical knowledge and understanding possible.
Alongside his discovery work, he has studied in depth the mechanisms used by microorganisms to produce
various kinds of substances, in particular carrying out extensive research at the genetic and molecular
biology level. His elucidation and application of the genetic control of chemical compound formation has
made it possible to manipulate microorganisms to produce novel compounds, as well as creating the
potential and ability to produce completely synthetic compounds. He and his colleagues obtained various
mutants of Streptomyces avermitilis in which part of the biosynthetic pathway of avermectin was blocked.
They determined the production pathway by isolating various precursors accumulated in each mutant. Each
mutated point on the chromosome was identified, eventually allowing him to clone all eighteen genes
providing a complete understanding of the biosynthetic mechanism for avermectin. The ground-breaking
genome analysis of S. avermitilis was the world’s first for a commercially-important actinomycete, and it
also demonstrated that the S. avermitilis has gene clusters involving biosynthesis of 37 kinds of secondary
metabolites, beside the immeasurably important avermectin.
Following his lead, creation of novel compounds using genetic engineering is now routinely carried out.
His original collaboration and continued pioneering in this area resulted in the appearance of hybrid
antibiotic ‘Mederrhodin’, created by him together with Prof. Sir David A. Hopwood in U.K. This
remarkable compounds was the first novel synthetic antibiotic created using genetic engineering and is
worthy of recognition as being the pioneer compounds in a completely new research field with almost
limitless potential.
He is being continually recognized internationally in the natural-products chemistry field, and for the
application of his discoveries, as evidenced by his numerous awards and honors. Among these are the
Hoeschst-Roussel Award from American Society for Microbiology (1985), the Pharmaceutical Society of
Japan Award (1986), Uehara Prize from the Uehara Memorial Foundation Japan (1989), the Japan
Academy Prize (1990), Charles Thom Award from Society for Industrial Microbiology, USA (1991),
Fujiwara Award from the Fujiwara Foundation of Sciences Japan (1995), Robert Koch Gold Medal from
Koch Institute, Germany (1997), Prince Mahidol Award from Thailand (1998), Nakanishi Prize of Japan
Chemical Society and the American Chemical Society (2000), Ernest Gunther Award of the American
Chemical Society (2005), Hamao Umezawa Memorial Award of the International Society of Chemotherapy
(2007), Tetrahedron Prize for Creativity in Organic Chemistry from Elsevier (2010), Arima Award of the
International Union of Microbiological Society (2011), Research Achievement Award of American Society
of Pharmacognosy (2013), Canada Garidner Global Health Award (2014), the Asahi Prize from the Asahi
Shimbun Company Japan (2015), and Nobel Prize in Physiology or Medicine (2015). He has been
decorated with Medal with Purple Ribbon from Japan (1992), Medal with Dark Blue Ribbon (1999), with
French L’Ordre National de la Legio d’honneur Chevalier (2007), and with the Order of the Sacred
Treasure, Gold and Silver Star from Government of Japan (2011). He has also been designated by Japan in
2012 as a highly-prestigious Person of Cultural Merit. He is a member of the Japan Academy, the German
Academy of Sciences Leopoldina, and the European Academy of Sciences, and is a Foreign Associate of
the United States National Academy of Sciences, the French Academy of Sciences, and the Chinese
Academy of Engineering. His honorary memberships include those of the American Society of
Biochemistry and Molecular Biology, the Royal Society of Chemistry, the Chemical Society of Japan,
Society for Actinomycetes Japan, Japan Society for Bioscience, Biotechnology and Agrochemistry, as well
long-standing member of the Editorial Board of the Journal of Antibiotics, serving as an Editor-in-Chief
from 2004 to 2013. He has published well over 1,000 scientific articles, continuing to publish extensively.
He still works as a Special Coordinator of the Research Project for Drug Discovery from Natural Products
in the Kitasato Institute for Life Sciences, Kitasato University for the discovery of microbial products
possessing biologically important activities.
Publications contributed by Dr. Satoshi Omura in Actinomycetologica
(※ The following electronic journals are available via J-STAGE)
Vertical distribution of microorganisms in soils (in Japanese with English abstract)
Yoko Takahashi, Yoshiko Seki, Yoshitake Tanaka, Ruiko Oiwa, Yuzuru Iwai, Satoshi Omura
Vol. 4 (1990) No. 1, p.1-6
Metabolism and Products of Actinomycetes - An Introduction
Yoshitake Tanaka, Satoshi Omura
Vol. 4 (1990) No. 1, p.13-14
Strategic Strain Improvement of Antibiotic Producer
Haruo Ikeda, Satoshi Omura
Vol. 5 (1991) No. 2, p.86-99
Quantitative Analysis for Madurose and Other Sugars in A Small Amount of Actinomycete Whole
Cells by Gas-Liquid Chromatography
Yoko Takahashi, Hisako Egusa, Baodi Deng, Hiroaki Kiyohara, Haruki Yamada, Yuzuru Iwai,
Satoshi Omura
Vol. 6 (1992) No. 2, p.69-78
Genetic Aspects of the Selective Production of Useful Components in the Avermectin Producer
Streptomyces avermitilis
Haruo Ikeda, Satoshi Omura
Vol. 7 (1993) No. 2, p.133-144
Plasmid-Mediated Gene Disruption in Streptomyces griseus
Nobuaki Kudo, Kenji Ueda, Haruo Ikeda, Satoshi Omura, Teruhiko Beppu, Sueharu Horinouchi
Vol. 8 (1994) No. 1, p.17-20
Cosmid Vector for Cloning and Analysis of Streptomyces DNA
Chang-Hong Pang, Masako Shiiyama, Haruo Ikeda, Haruo Tanaka, Satoshi Omura
Vol. 8 (1994) No. 1, p.21-25
Transfer of Staurosporine-Producing Strain Streptomyces staurosporeus AM-2282 to the Genus
Saccharothrix as Saccharothrix aerocolonigenes (Labeda 1986) subsp. staurosporeus subsp. nov.
Yoko Takahashi, Mayumi Shinose, Akio Seino, Yuzuru Iwai, Satoshi Omura
Vol. 9 (1995) No. 1, p.19-26
Physiological Regulation of Sporulation of Kitasatosporia setae in Submerged Culture
Yoko Takahashi, Yuzuru Iwai, Satoshi Omura
Rare Actinomycetes Isolated from Desert Soils
Yoko Takahashi, Atsuko Matsumoto, Akio Seino, Yuzuru Iwai, Satoshi Omura
Vol. 10 (1996) No. 2, p.91-97
Characterization of Actinomycetes Isolated from Fallen Leaves
Atsuko Matsumoto, Yoko Takahashi, Maki Mochizuki, Akio Seino, Yuzuru Iwai, Satoshi Omura
Vol. 12 (1998) No. 1, p.46-48
Application of PCR for Selection of Gram-Positive Bacteria with High DNA G+C Content among
New Isolates
Liyan Yu, Yoko Takahashi, Atsuko Matsumoto, Akio Seino, Yuzuru Iwai, Satoshi Omura
Vol. 16 (2002) No. 1, p.1-5
Microbacterium flavum sp. nov. and Microbacterium lacus sp. nov., isolated from marine
environments.
Akiko Kageyama, Yoko Takahashi, Yoshihide Matsuo, Kyoko Adachi, Hiroaki Kasai, Yoshikazu
Shizuri, Satoshi Omura
Vol. 21 (2007) No. 2, p.53-58
Microbacterium awajiense sp. nov., Microbacterium fluvii sp. nov. and Microbacterium pygmaeum sp.
nov.
Akiko Kageyama, Yoshihide Matsuo, Hiroaki Kasai, Yoshikazu Shizuri, Satoshi Omura, Yoko
Takahashi
Hamada Award 2011
Deciphering regulatory mechanisms for antibiotic production
by Streptomyces hormones
Shigeru Kitani
International Center for Biotechnology, Osaka University, 2-1 Yamadaoka, Suita, Osaka 565-0871, Japan.
INTRODUCTION
Members of the Gram-positive, soil-dwelling filamentous bacterial genus Streptomyces have been extensively studied due to their complex life cycle of morphological development and their ability to synthesize a wide variety of secondary metabolites, including antibiotics with important applications in human/veterinary medicine and agriculture. The regulation of secondary metabolism is a complicated process controlled by multiple-types of regulators that respond to various physiological and environmental factors. One of the well-known regulatory systems for Streptomyces secondary metabolism is a signaling system that uses the γ-butyrolactone autoregulator as a small diffusible signaling molecule (Bibb, 2005; Takano, 2006). γ-Butyrolactone autoregulators are active at nanomolar concentrations and elicit the onset of biosynthesis of secondary metabolites by modulating the DNA-binding activity of cognate receptor proteins; hence, they have been referred to as bacterial hormones, and sometimes as Streptomyces hormones.
The most characterized γ-butyrolactone
autoregulator is A-factor (Fig. 1A), which was discovered in the early 1960s as the first γ-butyrolactone autoregulator, which triggers both streptomycin production and morphological development in Streptomyces griseus (Khokhlov et
al., 1967, 1973). In the 1980s, our laboratory
demonstrated the structural diversity of γ-butyrolactone autoregulators by the identification of virginiae butanolides (VBs), which induce the production of the antibiotic virginiamycin in
Streptomyces virginiae, and IM-2, which switches on
the production of the blue pigment indigoidine in
Streptomyces lavendulae FRI-5 (Fig. 1A) (Yamada et al., 1987; Sato et al., 1989). In 1995, a receptor
protein for γ-butyrolactone autoregulators was first characterized by our identification of BarA as a
VB-specific receptor (Okamoto et al., 1995). The autoregulator signaling cascades in many
Streptomyces species and non-Streptomyces
actinomycetes continue to be gradually unraveled (Table 1).
In general, the autoregulator receptor binds to a specific DNA sequence called an autoregulatory element in the promoter region of its target genes, repressing their transcription (Folcher et al., 2001). Once production of the autoregulator is initiated and the concentration reaches a threshold concentration, the autoregulator interacts with its cognate receptor to lose the DNA-binding activity and induces the transcription of the target regulatory genes, and in turn activates the coordinated expression of regulatory and biosynthetic genes involved in secondary metabolism. Although the detailed regulatory machinery after the perception of autoregulators with the receptor appears to vary
among the strains, the pair set of γ-butyrolactone
autoregulators and their cognate receptors is a conserved and common system in the autoregulator
Fig. 1. Chemical structures of Streptomyces hormones. (A) Representative γ-butyrolactone autoregulators. The structures of A-factor from S. griseus, IM-2 from S. lavendulae FRI-5, VB-A from S. virginiae, and SCBs from S. coelicolor A3(2) are shown. (B) A representative γ-butenolide autoregulator. The structure of avenolide from S. avermitilis is shown.
signaling cascade. Thus, the autoregulators and the receptors have been a pivotal factor for deciphering the regulatory mechanism of secondary metabolism in actinomycetes.
In this review, I describe our recent studies on the regulatory mechanism for virginiamycin production controlled by VB, the identification of a new type of
Streptomyces hormone, and the application of our
knowledge of the autoregulator signaling cascades in the discovery of novel natural products.
Virginiamycin production controlled by VB
In Streptomyces species, multiple regulatory genes that directly or indirectly control the biosynthesis of a particular secondary metabolite(s) are frequently localized in the vicinity or in the middle of the biosynthetic gene cluster, and they occasionally form a regulatory island (Aigle et al., 2005; Pulsawat et al., 2007b). In particular, an autoregulator receptor gene is often accompanied by pathway-specific regulatory genes, autoregulator biosynthetic genes, and genes encoding a homologue of the autoregulator receptor (Fig. 2). These genes operate at several regulation layers for the biosynthesis of the cognate secondary metabolite through transcriptional regulation.
VB A, B, C, D, and E (VB-A-E), are isolated as γ-butyrolactone autoregulators containing a
1’-α-hydroxy group from the culture
broth of S. virginiae (Fig. 1A). They induce the coordinated production of two
different antibiotics, virginiamycin M1
(VM) and virginiamycin S (VS), a polyunsaturated macrolactone and a cyclic hexadepsipeptide, respectively, through the binding of VBs to BarA, a VB-specific receptor and DNA-binding transcriptional repressor (Fig. 3) (Kinoshita et al., 1997). The flaking
regions of barA contain several
important genes that are involved in the production of VM and VS (barB and
vmsR), the VB biosynthesis (barX, barS1,
and barS2), and the mechanisms of resistance to VS (varR and varS) (Fig. 2). These genes are clustered in a 10 kb region, and thus are considered to form a virginiamycin regulatory island. One interesting gene (varM) is found to be present at the barS1 upstream region in the regulatory island. Because the varM gene encodes a homologue of the type II ATP-binding cassette (ABC) transporter, we presume that VarM is involved in self-resistance to avoid suicide of the producer strain. To characterize the function of varM, we performed transcriptional analysis and antibiotic-susceptibility testing by heterologous expression of varM in Streptomyces lividans, and the results suggested that the expression of varM was intensely and tightly controlled by an intracellular concentration of VM, not by VS, and that proper
accessory proteins for providing the full function of VarM might be required for the transportation of VM (Kitani et al., 2010b).
Table 1. Functional properties of Streptomyces hormones and their autoregulator receptors for secondary metabolite production.
Fig. 2. Organization and comparison of the regulatory gene clusters for antibiotic production. Genes are color-coded by the predicted function of their products: autoregulator receptors (black), transcriptional regulators (dark gray), and autoregulator biosynthetic enzymes (light gray).
Next, to clarify the regulatory mechanism of virginiamycin production, we tried to find genes related to the virginiamycin biosynthesis by investigating the regions adjacent to the virginiamycin regulatory island. By screening of a cosmid library and gene disruption analysis, we identified a virginiamycin biosynthetic gene cluster composed of 27 genes located in the 87-kb region downstream of the barZ gene (Pulsawat et al., 2007b). The polyketide synthase (PKS) modules for the VM biosynthesis contain no acyltransferase (AT) domain,
although such a domain is found in the canonical modular type I PKSs, and have AT-docking sites immediately downstream of the ketosynthase (KS) domains in each PKS module (Fig. 4). In addition, within the gene cluster, virI encodes a discrete AT that provides the missing AT activity by acting in
trans with every PKS module of the VM-PKS. These
findings demonstrate that the VM biosynthetic gene cluster is a new member of the “AT-less” PKS cluster in streptomycetes. To achieve increased VM production selectively, we introduced an additional copy of virI or bkdA [encoding the branched-chain α-keto acid dehydrogenase (BCDH) E1 α and β fusion subunit] driven by the strong and constitutive promoter ermEp* into the wild-type strain (Pulsawat
et al., 2007a, 2007b). These engineered strains
showed a 1.5-fold increase of VM production, suggesting that the supply of malonyl-CoA and isobutyryl-CoA is one of the rate-limiting factors in the VM biosynthesis.
The further left-hand region of the VM biosynthetic gene cluster contains visG, which is necessary for the VS biosynthesis as a provider of the nonproteinogenic amino acid phenylglycine, together with the visE and visF genes, which encode plausible nonribosomal peptide synthetases for the VS framework (Ningsih et al., 2011). Taken together Fig. 3. Regulatory mechanism for virginiamycin production governed by
VB. Lines ending with a perpendicular line and arrows show repression and activation of the regulation, respectively.
Fig. 4. Proposed pathway for the biosynthesis of virginiamycin M1 in S. virginiae. The circles and squares represent enzymatic domains in the PKS and
NRPS, respectively. A, adenylation domain; ACP, acyl carrier protein; AT, acyltransferase; C, condensation domain; Cy, condensation/cyclization domain; DH, dehydratase; KR, ketoreductase; KS, ketosynthase; MT, methyltransferase; TE, thioesterase. The gray ovals symbolize the AT-docking sites and the gray circle in module 6 represents a region of unknown function. The presumed inactive DH and KS domains of module 1 and module 9 are shaded in black.
with the finding that four VS biosynthetic genes (visA,
visB, visC, and visD) are localized in the right-hand
region of varR (Namwat et al., 2002), these results indicate that the VS biosynthetic gene cluster is separated by both the virginiamycin regulatory island and the VM biosynthetic gene cluster.
The region between visE and the VM biosynthetic genes contains two additional regulatory genes, vmsS and vmsT, which encode a homologue of
Streptomyces antibiotic regulatory protein (SARP)
family regulators and response regulators of a bacterial two-component signal transduction system, respectively. Gene disruption analysis demonstrated that VmsS positively controls the production of both VM and VS, whereas VmsT regulates VM biosynthesis only (Pulsawat et al., 2009). To determine how these pathway-specific regulators are involved in virginiamycin biosynthesis, we performed extensive transcriptional analysis in the gene disruptants. The gene expression profiles indicated that coordinated virginiamycin biosynthesis is controlled by VmsS and VmsT with another pathway-specific regulator, VmsR, and these transcriptional regulators hierarchically control the expression of the biosynthetic gene cluster.
So far, we have clarified the regulatory mechanism of virginiamycin production, which is composed of VB biosynthetic enzymes (Lee et al., 2010) and several kinds of transcriptional regulators, and biosynthetic mechanisms of virginiamycin (VM and VS) at the molecular level (Fig. 3, Fig. 4). Thus, based on this information of virginiamycin biosynthesis, integral genetic manipulation should allow the construction of a high-level producer of virginiamycin as well as a single-producer for either VM or VS alone.
Avenolide, a new type of Streptomyces hormone
At least 60% of Streptomyces species appear to synthesize γ-butyrolactone autoregulators (Yamada, 1995), whereas the signaling molecules produced by many other species in order to elicit secondary metabolism still remain to be unraveled, largely
because Streptomyces hormones such as
γ-butyrolactone autoregulators are produced at extremely low concentrations in the culture medium and are difficult to isolate. In 2001, we isolated 320 µg of SCB1 from 280 L of the culture supernatant from Streptomyces coelicolor A3(2) (Fig. 1A), and characterized SCB1 as a γ-butyrolactone autoregulator that induces production of the pigmented antibiotics actinorhodin and undecylprodigiosin (Takano et al., 2000). In 2009, we reported that two further minor peaks possessing the antibiotic stimulatory activity were identified as SCB2 and SCB3 by using mass spectrometry (MS), tandem MS, and chemically synthesized
γ-butyrolactone analogues (Fig. 1A) (Hsiao et al., 2009). As of 2009, 14 γ-butyrolactone autoregulators had been identified in several Streptomyces species.
Through the functional analysis of autoregulator receptors in the regulation of secondary metabolism, we have learned that the genes involved in autoregulator biosynthesis and which encode the autoregulator receptor are frequently clustered at the same locus in various Streptomyces species (Fig. 2). AfsA is the key enzyme for A-factor biosynthesis to produce an A-factor intermediate (Kato et al., 2007), and thus homologues of AfsA, which probably produce β-keto ester precursors, are regarded as important enzymes for the biosynthesis of γ-butyrolactone autoregulators. Currently, our studies and the NCBI database show that many afsA-family genes are either adjacent to or closely localized with γ-butyrolactone receptor gene homologues with either the same or the opposite transcription orientation (Nishida et al., 2007; Kitani et al., 2010a; Lee et al., 2010; Aroonsri et al., 2012a). Genes encoding tailoring enzymes for producing the mature γ-butyrolactone autoregulators, such as BarS1 and BarS2 in the VB biosynthetic pathway (Shikura et al., 2002; Lee et al., 2008), are also sometimes found to be present at the flanking region of the pair set of an
afsA-family gene and a γ-butyrolactone receptor gene
homologue. However, in Streptomyces avermitilis, which produces the important anthelmintic agent known as avermectin, no afsA-family genes are localized in the avaR locus (Kitani et al., 2011; Miyamoto et al., 2011). The avaR locus includes
avaR1, avaR2, and avaR3, which encode homologues
of autoregulator receptors. Gene disruption analysis revealed that AvaR3 acts as a global regulator that controls the production of avermectin and other antibiotics, and the cell morphology. These findings suggested that the avaR locus is involved in the regulation of avermectin production via the perception of signaling molecules, and strongly encouraged us to investigate the function of two genes, aco and cyp17, that encode an acyl-CoA oxidase and a cytochrome P450 hydroxylase, respectively (Fig. 2), although aco and cyp17 have no assigned functions in the context of avermectin biosynthesis itself.
The level of avermectin production in the aco disruptant was remarkably reduced to 6% of the wild-type level, and this defect in the avermectin production was restored by the addition of an ethyl acetate extract of culture broth from the wild-type strain (Kitani et al., 2011). Interestingly, an ethyl
acetate extract of the aco disruptant and
representative γ-butyrolactone autoregulators were unable to restore the impaired avermectin production, implying that a signaling molecule distinct from known γ-butyrolactone autoregulators is involved in
the regulation of avermectin production. To isolate the signaling molecule from the 2,000 L of wild-type culture, we purified the AvaR1-interactive ligand that is biosynthesized by the wild-type strain, not by the
aco disruptant. Based on the structural elucidation
and the chemical synthesis of the AvaR1-interactive ligand (Uchida et al., 2011), we successfully identified “avenolide” (for S. avermitilis butenolide) as a signaling molecule involved in the induction of avermectin production (Fig. 1B). By adding synthetic avenolide into the aco disruptant, we found that avenolide triggered avermectin production at a minimum effective concentration of 4 nM. This result indicated that avenolide is new type of Streptomyces hormone involved in regulating secondary metabolism.
The focus on the difference between gene arrangements around the locus of the autoregulator receptor led us to the identification of avenolide as a new Streptomyces hormone. Gene arrangements similar to the aco/avaR1/cyp17 cluster are found in the genomes of several Streptomyces strains (Kitani
et al., 2011), suggesting that other streptomycetes
may produce avenolide or avenolide-like molecules and that they function as a general class of signaling molecules in regulating Streptomyces secondary metabolism. It will be interesting to investigate the distribution of producers for butenolide-type autoregulators.
Discovery of new compounds by the gene disruption of autoregulator receptors
Many genes encoding autoregulator receptors and their homologues have been reported to control secondary metabolism, and are classified into the following three groups based on their gene loci and properties in the regulatory system: (i) autoregulator receptor genes that are found within a biosynthetic gene cluster of a particular secondary metabolite(s), and operate at several regulation layers of biosynthesis of the cognate secondary metabolite (Kinoshita et al., 1997; Nakano et al., 2000; Matsuno
et al., 2004; Pulsawat et al., 2007b, 2009); (ii)
autoregulator receptor genes in the biosynthetic gene cluster that simultaneously control production of both the corresponding secondary metabolite and another secondary metabolite whose biosynthetic genes are distal to the locus of the receptor gene (Takano et al., 2005; Gottelt et al., 2010); (iii) autoregulator receptor genes that have no adjacent biosynthetic gene clusters but regulate the expression of biosynthetic genes that are located apart from the locus of the receptor gene (Horinouchi, 2007). These findings prompted us to investigate the possibility that an autoregulator receptor gene with no adjacent biosynthetic gene clusters controls the production of not only interesting secondary metabolites but also other
multiple secondary metabolites that have not yet been identified.
Kitasatospora setae NBRC 14216T produces
bafilomycin A1 and B1, specific inhibitors of vacuolar ATPase. The complete genome sequence revealed that K. setae has at least 24 genes or gene clusters for the biosynthesis of secondary metabolites, including bafilomycin (Ichikawa et al., 2010). However, a vast majority of these genes and clusters are of unknown function in the biosynthetic process and are presumably cryptic biosynthetic pathways. The genome of K. setae harbors three homologues (ksbA, ksbB, and ksbC) of autoregulator receptor genes together with afsA-family genes in the proximal region, suggesting that three kinds of autoregulator signaling cascades control the production of secondary metabolites in K. setae. To investigate the in vivo function of autoregulator receptor genes, we performed gene disruption analysis, and the results showed that KsbA functions as a negative regulator of bafilomycin production but has no influence on morphological development, whereas KsbC positively controls bafilomycin production and aerial mycelium formation (Table 1). The ksbA locus and the ksbC locus, neither of which have any biosynthetic gene cluster for secondary metabolites in the flanking regions, are 1.8 and 3.4 Mb, respectively, away from the bafilomycin biosynthetic genes. To explore the possibility that KsbC controls the production of other secondary metabolites, we carefully compared HPLC profiles of the methanol extract from the wild-type strain and the
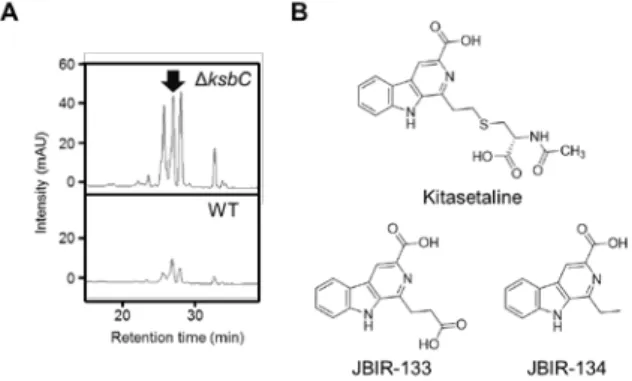
ksbC disruptant, and found that several peaks from
the ksbC disruptant were larger than those from the wild-type strain (Fig. 5A). The structure of the major peak was elucidated on the basis of physicochemical evidence, and thus the compound named kitasetaline was identified as a novel β-carboline alkaloid (Fig. 5B) (Aroonsri et al., 2012b). Subsequently, we identified putative kitasetaline biosynthetic genes by
Fig. 5. Overproduction of kitasetaline by the gene disruption of ksbC. (A) Kitasetaline production in the ksbC disruptant (ΔksbC) and the wild-type strain (WT). The peak of kitasetaline is indicated as a black arrow. (B) Chemical structures of kitasetaline, JBIR-133, and JBIR-134.
using the heterologous expression of a genomic BAC library of K. setae with S. avermitilis SUKA22, a model host for heterologous expression of secondary metabolism (Komatsu et al., 2010; Aroonsri et al., 2012a). Heterologous expression of the putative kitasetaline biosynthetic genes led to the production of not only kitasetaline but also JBIR-133, the production of which is also detected in the ksbC
disruptant, and JBIR-134 as novel β-carboline
alkaloids (Fig. 5B). These results indicated that disruption of the ksbC gene led to the discovery of new natural compounds such as kitasetaline and KBIR-133, together with the finding of a novel
biosynthetic route for microbial β-carboline
alkaloids.
The actinomycetes genome has numerous cryptic biosynthetic pathways in which genes appear to be expressed poorly, if at all, under the given cultivation condition. In fact, the K. setae wild-type strain produces a negligible amount of kitasetaline, while the ksbC disruptant shows precocious and abundant production of kitasetaline, enabling identification of the structure. As in the case of KsbC, genetic manipulation of an autoregulator receptor (or pseudoreceptor regulator), even one which has no proximal gene cluster for secondary metabolites, will be a promising strategy in the discovery of novel natural products and new biosynthetic pathways.
CONCLUSION
Streptomyces hormones, exemplified by
γ-butyrolactone-type autoregulators and
butenolide-type autoregulators, and their cognate receptors could provide a key to decipher the complicated regulatory mechanism of secondary metabolism, including antibiotic production, in actinomycetes. Our previous and current studies have contributed to an increase in antibiotic production (Table 1), to the deduction of common features and differences of regulatory pathways for the biosynthesis of secondary metabolites, and to the awakening of silent gene clusters for the discovery of novel compounds. Further understanding and application of the regulatory mechanism governed by
Streptomyces hormones will shed new light on the
remarkable ability encoded in the actinomycetes genome to produce clinically and agrochemically useful natural compounds.
ACKNOWLEDGEMENTS
It was my great honor to receive the Hamada Award of the Society for Actinomycetes, Japan (SAJ) in 2011. I would like to express my appreciation to
all the members of the Laboratory of Molecular Microbiology, International Center for Biotechnology, Osaka University for their support during this work. I am most grateful to Prof. Takuya Nihira for inspiring me throughout my research work with insightful guidance and compassionate advice. I am deeply indebted to Prof. Haruo Ikeda for his constant help and suggestions. I would also like to express my thanks to Prof. Yasuhiro Yamada, Prof. Mervyn Bibb, and Prof. Eriko Takano. Finally, I would like to thank the members of the SAJ for their continuing interest and support.
REFERENCES
Aigle, B., et al. (2005). Involvement of AlpV, a new
member of the Streptomyces antibiotic regulatory protein family, in regulation of the duplicated type II polyketide synthase alp gene cluster in
Streptomyces ambofaciens. J. Bacteriol. 187,
2491-2500.
Aroonsri, A., et al. (2012a). Pleiotropic control of
secondary metabolism and morphological development by KsbC, a butyrolactone autoregulator receptor homologue in
Kitasatospora setae. Appl. Environ. Microbiol. 78,
8015-8024.
Aroonsri, A., et al. (2012b). Kitasetaline, a novel
beta-carboline alkaloid from Kitasatospora setae NBRC 14216T. J. Biosci. Bioeng. 114, 56-58.
Bibb, M.J. (2005). Regulation of secondary
metabolism in streptomycetes. Curr. Opin.
Microbiol. 8, 208-215.
Choi, S.U., et al. (2004). Cloning and functional
analysis by gene disruption of a gene encoding a γ-butyrolactone autoregulator receptor from
Kitasatospora setae. J. Bacteriol. 186, 3423-3430.
Folcher, M., et al. (2001). Pleiotropic functions of a
Streptomyces pristinaespiralis autoregulator
receptor in development, antibiotic biosynthesis, and expression of a superoxide dismutase. J. Biol.
Chem. 276, 44297-44306.
Gottelt, M., et al. (2010). Deletion of a regulatory
gene within the cpk gene cluster reveals novel antibacterial activity in Streptomyces coelicolor A3(2). Microbiology. 156, 2343-2353.
Horinouchi, S. (2007). Mining and polishing of the
treasure trove in the bacterial genus Streptomyces.
Biosci. Biotechnol. Biochem. 71, 283-299.
Hsiao, N.H., et al. (2009). Analysis of two additional
signaling molecules in Streptomyces coelicolor and the development of a butyrolactone-specific reporter system. Chem. Biol. 16, 951-960.
Ichikawa, N., et al. (2010). Genome sequence of
Kitasatospora setae NBRC 14216T: an
Streptomycetaceae. DNA. Res. 17, 393-406.
Kato, J.Y., et al. (2007). Biosynthesis of
γ-butyrolactone autoregulators that switch on secondary metabolism and morphological development in Streptomyces. Proc. Natl. Acad.
Sci. U. S. A. 104, 2378-2383.
Khokhlov, A.S., et al. (1967). The A-factor,
responsible for streptomycin biosynthesis by mutant strains of Actinomyces streptomycini. Dokl.
Akad. Nauk. SSSR. 177, 232-235.
Khokhlov, A.S., et al. (1973). Effect of A-factor on
the growth of asporogenous mutants of
Streptomyces griseus, not producing this factor. Z. Allg. Mikrobiol. 13, 647-655.
Kim, H.S., et al. (2004). Cloning and characterization of a gene encoding the γ-butyrolactone autoregulator receptor from
Streptomyces clavuligerus. Arch. Microbiol. 182,
44-50.
Kinoshita, H., et al. (1997). Butyrolactone
autoregulator receptor protein (BarA) as a transcriptional regulator in Streptomyces virginiae.
J. Bacteriol. 179, 6986-6993.
Kitani, S., et al. (2001). Gene replacement analysis
of the butyrolactone autoregulator receptor (FarA) reveals that FarA acts as a novel regulator in secondary metabolism of Streptomyces lavendulae FRI-5. J. Bacteriol. 183, 4357-4363.
Kitani, S., et al. (2008). Disruption of sscR encoding
a γ-butyrolactone autoregulator receptor in
Streptomyces scabies NBRC 12914 affects
production of secondary metabolites. Folia.
Microbiol. (Praha). 53, 115-124.
Kitani, S., et al. (2010a). Control of secondary
metabolism by farX, which is involved in the γ-butyrolactone biosynthesis of Streptomyces
lavendulae FRI-5. Arch. Microbiol. 192, 211-220.
Kitani, S., et al. (2010b). Characterization of varM
encoding type II ABC transporter in Streptomyces
virginiae, a virginiamycin M1 producer.
Actinomycetologica. 24, 51-57.
Kitani, S., et al. (2011). Avenolide, a Streptomyces
hormone controlling antibiotic production in
Streptomyces avermitilis. Proc. Natl. Acad. Sci. U. S. A. 108, 16410-16415.
Komatsu, M., et al. (2010). Genome-minimized
Streptomyces host for the heterologous expression
of secondary metabolism. Proc. Natl. Acad. Sci. U.
S. A. 107, 2646-2651.
Lee, K.M., et al. (2005). Cloning and in vivo
functional analysis by disruption of a gene encoding the γ-butyrolactone autoregulator receptor from Streptomyces natalensis. Arch.
Microbiol. 184, 249-257.
Lee, Y.J., et al. (2008). Identification by gene
deletion analysis of barS2, a gene involved in the biosynthesis of γ-butyrolactone autoregulator in
Streptomyces virginiae. Arch. Microbiol. 189,
367-374.
Lee, Y.J., et al. (2010). Null mutation analysis of an
afsA-family gene, barX, that is involved in
biosynthesis of the γ-butyrolactone autoregulator in Streptomyces virginiae. Microbiology. 156, 206-210.
Matsuno, K., et al. (2004). Identification by gene
deletion analysis of barB as a negative regulator controlling an early process of virginiamycin biosynthesis in Streptomyces virginiae. Arch.
Microbiol. 181, 52-59.
Miyamoto, K.T., et al. (2011). The autoregulator-receptor homologue AvaR3 plays a regulatory role in antibiotic production, mycelial aggregation and colony development of
Streptomyces avermitilis. Microbiology. 157,
2266-2275.
Nakano, H., et al. (1998). Gene replacement analysis
of the Streptomyces virginiae barA gene encoding the butyrolactone autoregulator receptor reveals that BarA acts as a repressor in virginiamycin biosynthesis. J. Bacteriol. 180, 3317-3322.
Nakano, H., et al. (2000). A null mutant of the
Streptomyces virginiae barA gene encoding a
butyrolactone autoregulator receptor and its phenotypic and transcriptional analysis. J. Biosci.
Bioeng. 90, 204-207.
Namwat, W., et al. (2002). Characterization of
virginiamycin S biosynthetic genes from
Streptomyces virginiae. Gene. 286, 283-290.
Ningsih, F., et al. (2011). VisG is essential for
biosynthesis of virginiamycin S, a streptogramin type B antibiotic, as a provider of the nonproteinogenic amino acid phenylglycine.
Microbiology. 157, 3213-3220.
Nishida, H., et al. (2007). Evolution of γ-butyrolactone synthases and receptors in
Streptomyces. Environ. Microbiol. 9, 1986-1994.
Okamoto, S., et al. (1995). Virginiae butanolide
binding protein from Streptomyces virginiae. Evidence that VbrA is not the virginiae butanolide binding protein and reidentification of the true binding protein. J. Biol. Chem. 270, 12319-12326.
Onaka, H., et al. (1995). Cloning and characterization of the A-factor receptor gene from
Streptomyces griseus. J. Bacteriol. 177,
6083-6092.
Pulsawat, N., et al. (2007a). Identification of the
bkdAB gene cluster, a plausible source of the
starter-unit for virginiamycin M production in
Streptomyces virginiae. Arch. Microbiol. 187,
459-466.
Pulsawat, N., et al. (2007b). Characterization of
biosynthetic gene cluster for the production of virginiamycin M, a streptogramin type A antibiotic, in Streptomyces virginiae. Gene. 393, 31-42.
Pulsawat, N., et al. (2009). Hierarchical control of
virginiamycin production in Streptomyces
virginiae by three pathway-specific regulators:
VmsS, VmsT and VmsR. Microbiology. 155, 1250-1259.
Santamarta, I., et al. (2005). Different proteins bind
to the butyrolactone receptor protein ARE sequence located upstream of the regulatory ccaR gene of Streptomyces clavuligerus. Mol. Microbiol.
56, 824-835.
Sato, K., et al. (1989). Isolation and structure of a
new butyrolactone autoregulator from
Streptomyces sp. FRI-5. J. Ferment. Bioeng. 68,
170–173..
Shikura, N., et al. (2002). barS1 a gene for
biosynthesis of a γ-butyrolactone autoregulator, a microbial signaling molecule eliciting antibiotic production in Streptomyces species. J. Bacteriol.
184, 5151-5157.
Takano, E., et al. (2000). Purification and structural
determination of SCB1, a γ-butyrolactone that elicits antibiotic production in Streptomyces
coelicolor A3(2). J. Biol. Chem. 275,
11010-11016.
Takano, E., et al. (2001). A complex role for the
γ-butyrolactone SCB1 in regulating antibiotic production in Streptomyces coelicolor A3(2). Mol.
Microbiol. 41, 1015-1028.
Takano, E., et al. (2005). A bacterial hormone (the
SCB1) directly controls the expression of a pathway-specific regulatory gene in the cryptic type I polyketide biosynthetic gene cluster of
Streptomyces coelicolor. Mol. Microbiol. 56,
465-479.
Takano, E. (2006). γ-Butyrolactones: Streptomyces
signalling molecules regulating antibiotic production and differentiation. Curr. Opin.
Microbiol. 9, 287-294.
Uchida, M., et al. (2011). Total synthesis and
absolute configuration of avenolide, extracellular factor in Streptomyces avermitilis. J. Antibiot.
(Tokyo). 64, 781-787.
Yamada, Y., et al. (1987). The structure of inducing
factors for virginiamycin production in
Streptomyces virginiae. J. Antibiot. (Tokyo). 40,
496-504.
Yamada, Y. (1995). Butyrolactone autoregulators,
inducers of secondary metabolites, in
Publication of Award Lecture
The Society for Actinomycetes Japan Hamada Award 2011,
Dr. Shigeru Kitani
“Deciphering regulatory mechanisms for antibiotic production
by Streptomyces hormones”
Actimomycetologica (2015) 29 [2], S10-S17.
International Center for Biotechnology, Osaka University, 2-1 Yamadaoka, Suita, Osaka 565-0871, Japan
58 th Regular Colloquim
Date: Oct. 30 (Fri), 2015
Place: The National Institute of Infectious
Diseases
Program:
1. “RNA-guided DNA cleavage mechanism by CRISPR-Cas9”
Hiroshi NISHIMASU (The University of Tokyo, JST, PRESTO)
2. “Development of cosmetic ingredients using actinomycetes”
Yukio NAKAMURA (KOBAYASHI
Pharmaceutical Co., Ltd. Skincare Division)
3. “Intestinal microbiota-derived metabolites shape intestinal immune system through epigenetic modifications”
Koji HASE (Faculty of Pharmacy, Keio University)
4. “Splendid Gifts from Microorganisms -A charm of actinomycetes is endless”
Yoko TAKAHASHI (Laboratory of
Microbiology for Drug Discovery, Kitasato Institute for Life Sciences, Kitasato University)
The 2015 Annual Meeting of the Society for Actinomycetes Japan
Dates Sept 7 (Mon) –Sept 8 (Tue), 2015
Venue: Toyama International Conference Center
Oote-machi 1-2, Toyama 930-0084, Japan TEL: +81-76-424-5931 Fax: +81-76-493-7170
September 7 (Mon)
9:20 Opening Remarks
9:30 Oral Presentation –Session 1
O-1 Search for new actinomycete secondary metabolites based on molecular networking
○Nobuhiro Koyama1, 3, Dimitrios J. Floros2, Paul R. Jensen4, Pieter C. Dorrestein3
(1Graduate School of Pharmaceutical Sciences, Kitasato University, 2Chemistry and
Biochemistry, University of California San Diego, 3Skaggs School of Pharmacy &
Pharmaceutical Sciences, University of California San Diego, 4Scripps Institution of
Oceanography, University of California San Diego)
O-2 New compound produced by an actinomycete strain, Streptomyces griseus OS-3601
○Rei Miyano 1, Hirotaka Matsuo 2, Toru Kimura 1, Yukihiro Asami 2, Masato Iwatsuki 1,2,
Noriko Sato 3, Kazuro Shiomi 1,2, Yōko Takahashi 2, Satoshi Omura2, Takuji Nakashima 2
(1Graduate School of Infection Control Sciences, 2Kitasato Institute for Life Science, Kitasato
Univ., 3Department of Pharmacy, Faculty of Pharmaceutical Science, Kitasato Univ.)
O-3 Genomic exploration for unexploited biosynthetic machinery mediated by amino group
carrier protein in Actinomycetes
○Kenichi Matsuda1, Fumihito Hasebe1, Takeo Tomita1, Yuh Shiwa2, Yu Kanesaki2,
Hirofumi Yoshikawa2, Kazuo Shin-ya3, Tomohisa Kuzuyama1, Makoto Nishiyama1
(1Univ. Tokyo, 2Tokyo Univ. of Agric., 3AIST)
O-4 Mycolic acid-containing bacteria induce the production of novel secondary metabolites in
Streptomyces strains
○Shotaro Hoshino1, Lihan Zhang1, Takayoshi Awakawa1, Toshiyuki Wakimoto2,
Hiroyasu Onaka3 and Ikuro Abe1
(1Graduate School of Pharmaceutical Sciences, The University of Tokyo, 2Graduate School of
Pharmaceutical Sciences, Hokkaido University, 3Graduate School of Agricultural and Life
Sciences, The University of Tokyo)
O-5 Viability of mycolic acid-containing bacteria is crucial for Streptomyces antibiotic
production in combined-culture
○Shumpei Asamizu 1, Taro Ozaki 1, Kanae Teramoto 2, Katsuya Satoh 3, Hiroyasu Onaka 1
(1Graduate School of Agricultural and Life Sciences, The University of Tokyo, 2Advanced
Technology Department, JEOL Ltd., 3Takasaki Advanced Radiation Research Institute, Japan
Atomic Energy Agency)
10:45 Break
11:00 Oral Presentation –Session 2
O-6 Identification of telomestatin biosynthetic gene cluster by a series of gene inactivation
experiment
○Keita Amagai1,2, Fumitaka Kudo3, Tadashi Eguchi, Hiroyuki Osada2, Haruo Ikeda4,
(1Technology Research Association for Next generation natural products chemistry, 2RIKEN
CSRS, Chemical Biology, 3Tokyo Institute of Technology, 4Kitasato University, 5AIST)
O-7 Functional analysis of the antibiotic BD-12 biosynthetic genes
○Haruka Niikura 1, Chitose Maruyama 1, Miho Izumikawa 2, Jun Ishikawa 3, Haruo Ikeda 4,
Kazuo Shin-ya 5, Yoshimitsu Hamano 1
(1 Fukui Pref. Univ., 2 JBIC, 3 NIID, 4 Kitasato Univ., 5 AIST)
O-8 Biosynthesis of azoxyalkene compound produced by a mutant of Streptomyces rochei
Hirofumi Kunitake, Haruyasu Kinashi, ○Kenji Arakawa (Graduate School of AdSM, Hiroshima University)
O-9 Formation of the unique diazepanone-ring in caprazamycin biosynthesis
○Taro Shiraishi,1 Masayuki Igarashi,2 Makoto Nishiyama,1 Tomohisa Kuzuyama1
(1BRC, UTokyo, 2BIKAKEN)
12:00 Lunch
13:30 The SAJ Plenary Meeting
14:00 Awarding Ceremony
13:20 Award Lectures
SAJ Award
Mechanism of terpenoid biosynthesis in actinomycetes
Dr. Tomohisa Kuzuyama
(Biotechnology Research Center, The University of Tokyo)
SAJ Merit Award
Research on the diverse capability of Actinomycetes and contribution to SAJ activities
Dr. Fumio Kato
(School of Pharmaceutical Sciences, Toho University)
Hamada Award
Development of the selective isolation method for non-filamentous actinobacteria and taxonomic studies
Dr. Moriyuki Hamada
(Biological Resource Center, National Institute of Technology and Evaluation)
Discovery of type III polyketide synthases from Streptomyces, and their physiological implications
Dr. Nobutaka Funa
(Graduate Division of Nutritional and Environmental Sciences, University of Shizuoka)
16:20 Invited Lectures
Made in Japan, Made in Toyama - Drug discovery from the medicine city -
Dr. Nobuhiko Nomura
(Research Laboratories, Toyama Chemical Co.,Ltd)
Actinomycetes-derived carbohydrate-binding agents (CBA): mechanistically novel agents with antiviral and antiparasitic activity
Dr. Jan Balzarini
(Rega Institute for Medical Research, KU Leuven)
18:00 Break
18:30 Reception
September 8 (Tue)
9:15 Poster Short Presentation (odd numbers) 9:45 Poster Session (odd numbers)
11:00 Oral Presentation –Session 3
O-10 Analysis of the molecular mechanism by which lincomycin at subinhibitory
concentrations alters gene expression and potentiates secondary metabolism of
Streptomyces spp.
○Kazuma Shimono1, Tomoko Maruyama1, Yu Imai1, Yukinori Tanaka2, Seizo Sato3,
Kozo Ochi,2 Takeshi Hosaka1
(1Sch. Agric. Shinshu Univ, 2Fac. Life. Sci. Hiroshima Inst. Technol, 3Nippon Suisan Kaisha. Ltd.
Tokyo Innovation Center)
O-11 Production Enhancement of sinefungin by multiple mutations introduced in rpoB gene at the rif-1 cluster of Streptomyces incarnatus NRRL8089
○Takashi Tamura1, Hitomi Shimizu1, Michiko Nemoto1, Kenji Inagaki1
(1Graduate School of Life & Environmental Science, Okayama University)
O-12 Genome analysis and modification of Rhodococcus erythropolis strains; toward creating improved host microorganisms
○Wataru Kitagawa1, 2, Miyako Hata1
(1Bioproduction research institute, AIST, 2Grad. Sch. Agric., Hokkaido Univ.)
O-13 Structural analyses of cell-wall peptidoglycans of the family Micromonosporacae strains
○Akira Také1, Takuji Nakashima2, Yuki Inahashi2, Kazuro Shiomi1,2, Yōko Takahashi2,
Satoshi Ōmura2, Atsuko Matsumoto1,2
(1Grad. Sch. Infection Control Sci.; 2Kitasato Inst. Life Sci., Kitasato Univ.)
12:00 Lunch
13:30 Poster Short Presentation (even numbers) 14:00 Poster Session (even numbers)
15:00 Break
15:15 Oral Presentation –Session 4
O-14 Structure and biological activity of a new compound sagamilactam produced by an actinomycete strain, Actinomadura sp. K13-0306
○Toru Kimura1, Yukihiro Asami1,2, Masato Iwatsuki1,2, Atsuko Matsumoto1,2, Noriko Sato3,
Kazuro Shiomi1,2, Yoko Takahashi2, Satoshi Ōmura2, Takuji Nakashima1,2
(1Grad. Sch. Infection Control Sci., Infect. Cont. Sci., 2Kitasato Inst. Life Sci., Kitasato Univ., 3Kitasato pham., Kitasato Univ.)
O-15 Isolation and structure determination of new siderophore albachelin from Amycolatopsis
alba
○Shinya Kodani1,Hisayuki Komaki2,Masahiro Suzuki1,Hikaru Hemmi3,
Mayumi Ohnishi-Kameyama3
(1Graduate School of Agriculture, Shizuoka University, 2Biological Resource Center, National
Institute of Technology and Evaluation, 3National Food Research Institute, National Agriculture
and Food Research Organization)
O-16 Structure-based engineering of a crotonyl-CoA reductase/carboxylase AntE for biogenesis of unnatural polyketides
○Lihan Zhang1, Takahiro Mori1, Takayoshi Awakawa1, Wen Liu2, Ikuro Abe1
(1Graduate School of Pharmaceutical Sciences, The University of Tokyo, 2 Shanghai Institute of
Organic Chemistry, Chinese Academy of Sciences)
O-17 Bacterial indole prenyltransferases for chemoenzymatic synthesis of prenylated compounds
○Intan Timur Maisyarah,1 Kazuo Shin-ya,2 Makoto Nishiyama,1 Tomohisa Kuzuyama1
O-18 An improved design for acyltransferase domain replacements in modular polyketide synthases
○Satoshi Yuzawa1, Kai Deng2,3, Trent Northen2,4, Paul Adams2,4,5, Leonard Katz1,6,
Jay Keasling1,2,4,5,6,7
(1QB3 Inst., UC Berkeley, 2JBEI, 3SNL, 4Phys. Biosci. Div., LBNL, 5Dept. Bioeng.,
UC Berkeley, 6SynBERC, 7Dept. Chem. Biomol. Eng., UC Berkeley)
16:30 Closing Remarks, Poster Award Ceremony
Poster Session
P-1 Construction of gene expression database by combination of next generation sequencing
and HiCEP method
Misugi Uraji1, Harunobu Yunokawa2, Kun Wan1, ○Tadashi Hatanaka1
(1Research Institute for Biological Sciences, Okayama, 2Messenger Scape Co. Ltd.)
P-2 Image analysis of mycelial morphology and application to Tacrolimus production
○Ayana Ide, Fujio Goto, Shiho Shimizu (Astellas Pharma Tech Co., Ltd.)
P-3 Production Inhibition for FK506 (Tacrolimus) Caused by Culture Medium (defatted wheat
germ) of Different Suppliers and Its Prevention
○Ayako Futase1, Kuniharu Watanabe1, Ayana ide1, Shiho Shimizu
(Astellas Pharma Tech Co., Ltd)
P-4 Discrimination of deep-sea Micromonospora species by Matrix-Assisted Laser Desorption
Ionization-Time of Flight Mass Spectrometry (MALDI-TOF MS)
○Masaki Hatano1, Yuko Takahashi1, Ryuichi Sawa1, Masayuki Igarashi1, Chiaki Kato2,
Masakatsu Shibasaki1
(1Institute of Microbial Chemistry (BIKAKEN), 2Japan Agency for Marine-Earth Science and
Technology)
P-5 Discovery of a new function of second metabolite produced by Streptomyces as
organocatalysis
○Tatsuya Nishiyama1, Yoshiteru Hashimoto1, Hitoshi Kusakabe2, Takuto Kumano1,
Michihiko Kobayashi1
(1 Institute of Applied Biochemistry, and Graduate School of Life and Environmental Sciences,
University of Tsukuba, 2 Life Science Research Center, College of Bioresource Science,
Nihon University, 3 Enzyme-Sensor Co., Ltd.)
P-6 Structure determination of a siderophore peucechelin from Streptomyces peucetius
○Masahiro Suzuki1,Shinya Kodani1,Hisayuki Komaki2,Fumiya Kobayakawa1, Hikaru Hemmi3
(1Graduate School of Agriculture, Shizuoka University, 2Biological Resource Center, National
Institute of Technology and Evaluation, 3National Food Research Institute, National Agriculture
and Food Research Organization)
P-7 Antibiotics as the words of microorganisms
○Ryota Kuramoto1 , Yukinori Tanaka1 , Takeshi Hosaka2 , Kozo Ochi1
(1Hiroshima Institute of Technology, 2Shinshu University)
P-8 Complete genome sequence of Actinomyces sp. Chiba101 isolated from a pig
Tomoko Kobayashi1, Yuriko Sekigawa1, Torii Yasushi1, Eiji Yokoyama2, Ishige Taichiro3,
Kanesaki Yu3,, Satoshi Murakami1
(1Laboratory of Animal Hygiene, Department of Animal Science, Tokyo University of Agriculture, 2Division of Bacteriology, Chiba Prefectural Institute of Public Health, 3Genome Research Center,
Tokyo University of Agriculture)
P-9 The study on discovery of new compounds from actinomycete strains by physicochemical
screening
Hirotaka Matsuo1, Panitch Boonsnongcheep 2, Toru Kimura3, Masato Iwatusuki1, Michiko Sato4,