1
修
士 学 位 論 文
題
名 シ マ オ オ タ ニ ワ タ リ 類 に
お け る 生 殖 的 隔 離 の 成 立 過 程 の 解 明
指 導 教 授 村 上 哲 明 教 授 平 成 22年 1月 7日 提 出 首都大学東京大学院 理 工 学 研 究 科 生 命 科 学 専 攻 学修番号 09881341 氏 名 山 田 香 菜 子2
目次
ページ Abstract ……….… 2 要旨 ………..……3 はじめに ………...5 材料と方法 ………...9 結果 ………....20 考察 ………23 今後の展望 ………27 謝辞 ………28 引用文献 ………29 Table 1 Figure 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 123
Evolution of reproductive isolation in Asplenium nidus
complex
Kanako Yamada
Abstract.
Species is the most basic and real unit in Taxonomy. At present, most commonly accepted definition of species is the biological species concept that defines species as “groups of actually or potentially interbreeding natural populations, which are
reproductively isolated from other such groups” (Mayr 1942). Following biological species concept, reproductive isolation is the most important for defining species. Asplenium nidus L. and its relatives (A. nidus complex) are a group of epiphytic fern species with simple leaves. It was reported that many cryptic species
(morphologically hard to distinguished but reproductively isolated to each other) are contained in the complex. It was also elucidated that the genetic distances based on rbcL sequences and the degree of reproductive isolation among the cryptic species are
positively correlated (Yatabe et al. 2009).
Therefore, I considered that A. nidus complex might be one of the best materials to reveal the evolutionary process of reproductive isolation, which is the main process of speciation. During the period of master course, I have two kinds of research as follows up to the present: searching members of A. nidus complex which are on the process of speciation in Japan and developing codominant nuclear DNA markers, which will enable me to estimate parents of zygotes or small embryos.
I collected plant samples of Japanese species of the complex: Asplenium
antiquum, A. nidus and A. setoi from 15 localities of Japan, also observing their natural
habitats. Then, I determined their rbcL sequences. As the result, even between the samples of A. setoi collected in Ogasawara Islands and those in Ryukyu Islands, up to 6 base pair differences were observed. As Yatabe et al. (2009) reported that reproductive isolation was observed among individual of the complex whose rbcL sequences differed in 5-6 bases, we can expect that at least partial reproductive isolation might exist among individuals of A. setoi in Ogasawara Islands and those in Ryukyu Islands.
I also tried to develop SSR, gapCp and pgiC markers during period of my master course. Then, I succeeded in developing PCR primers to amplify pgiC marker. Obtained nucleotide sequences of pgiC gene using the developed primers, showed enough
amount of variation even among the samples of Asplenium setoi from various localities in Japan.
In near future, I am planning to conduct artificial crossing experiment between
Asplenium setoi from Ogasawara Islands and those from Ryukyu Islands. Moreover, I
will observe detailed process from fertilization to embryogenesis during artificial crossing among various combination of A. nidus complex in Japan and from adjacent foreign countries, using digital microscope and other facilities.
4
学位論文要旨(修士(理学)
)
論文著者名 山田 香菜子 論文題名:シマオオタニワタリ類における生殖的隔離の成立過程の解明 本文 種(species)は、生物の分類上最も基本的な単位である。現在、有性生殖を 行う生物において最も一般的に用いられている種の定義は、互いに交配して正 常な子孫を残すことができる集団を同じ種と見なす生物学的種概念である。こ の定義に基づくと、生殖的隔離が見られるかどうかが種を区別する上で最も重 要な情報となる。シダ植物において、異なる生物学的種に属する個体同士を交 配させた場合、雑種の胞子体が発生しない、もしくは発生した胞子体が正常な 胞子をつけないという結果が観察されている。しかし、胞子体が発生しなかっ た場合に、受精から胞子体の発生に至るどの段階で生殖的隔離が起こるのか観 察された事例はこれまで報告されていない。特に受精後に隔離が働いている場 合、観察した胚が本当に雑種由来であるかを検証する方法も必要となる。これ までシダ植物の人工交配実験で生じた胞子体の両親判定にはアロザイム多型解 析が用いられてきたが、ごく小さな胚はまだ自力で酵素を生産しないため、両 親判定には共優性でPCR 法によってシグナルの増幅が可能な核 DNA マーカー が必須であると考えられる。 本研究の材料であるシマオオタニワタリ類(チャセンシダ科)は、生殖的隔 離の強さと葉緑体遺伝子の1 つであるrbcL の塩基配列の異なる程度に正の相関 があることが分かっているシダ植物である。そのため、系統関係が遠くなるご とにどのように生殖的隔離が強まっていくのかを解析するには最適の材料と考 えられる。本研究では、シマオオタニワタリ類を材料として、シダ植物の生殖 的隔離がどのような段階を経て進化していくのか解明することを目的としてい る。そのために、今年度までに「rbcL 遺伝子の塩基配列解析による種分化の途 上にある可能性のあるシマオオタニワタリ類の日本国内における探索」と「核 DNA マーカーの開発」の2つを行った。 まず、日本国内各地でシマオオタニワタリ類のサンプリングを行った。特に 先行研究において種内のrbcL の塩基配列の大きなばらつきがある可能性が指摘 されていたオオタニワタリとヤエヤマオオタニワタリを重点的に採集した。採 集したサンプルからDNA を抽出してrbcL 遺伝子の塩基配列を解読し、集団間 で比較した。さらに、得られたサンプルのDNA を用いて、マイクロサテライト マーカー、および核 DNA にコードされている遺伝子である gapCp ならびに pgiC のイントロンを利用した共優性マーカーの開発を試みた。 まずrbcL の塩基配列解析の結果、小笠原諸島産のヤエヤマオオタニワタリと 南西諸島産のヤエヤマオオタニワタリとの間には5-6 塩基のrbcL の塩基配列の 違いがあることが明らかになった。Yatabe et al. (2009) による先行研究の結果 を見ると、この程度の違いがあれば何らかの生殖的隔離が存在することが十分5 に考えられる。また、pgiC のイントロンを利用した、シマオオタニワタリ類全 般で利用可能な核DNA マーカーの開発にも成功した。 今後、小笠原諸島産ヤエヤマオオタニワタリと南西諸島産ヤエヤマオオタニ ワタリを含むrbcL 配列の違いの程度が様々なシマオオタニワタリ類の個体間で 人工交配実験を行い、胞子体が発生しない組み合わせについては、受精から胞 子体発生のプロセスを顕微鏡下で観察して、どの段階で生殖的隔離が生じてい るかを明らかにしたいと考えている。そして、rbcL 配列の違いの程度と生殖的 隔離の起こる段階の関係をより詳しく解析することによって、シダ植物におけ る生殖的隔離の進化過程を解明する予定である。
6
1. はじめに
種(species)は、生物の分類において基本となる単位であり、生物学上重 要な概念でもある。従来、種は主に形態形質の不連続性によって認識、記載さ れてきた。しかし、形態形質の不連続性に基づく種の認識は、見る者の主観に よって左右されることがままあり、さらに生物の形態は環境によって変化する 場合も多いことから、生物学的な実体を客観的にとらえられる保証がない。 そのため、種をより客観的に定義するため、単系統に属し、他の系統と異 なる特徴・進化的傾向を持つ集団を種とする系統学的種概念(Cracraft, 1989) や、ニッチ(生態的地位)が分化しているかどうかで種のまとまりを判断する 生態的種概念(Van Valen, 1976)など、様々な種の定義が提唱されてきた。中 でも現在、種の定義として最も一般的に用いられているのは、Mayr(1942)ら により提唱された、互いに交配して正常な子孫を残すことができる生物の集団 を種とみなす、生物学的種概念と呼ばれるものである。この定義に基づくと、 自然条件下、もしくは潜在的に交配が起きるかどうか、すなわち生殖的隔離が 見られるかどうかが種を区別する上で最も重要な情報となる。したがって、生 物学的種概念は、有性生殖を行わない生物には適用できない。また、既に絶滅 した生物においては生殖的隔離の有無を検証する方法がないため適用できない といった問題点も存在する。しかしながら、現存する有性生殖を行う高等生物 については、生物学的種概念は十分に有用である(村上 2007)。本研究におい ても、生物学的種概念に基づいて種を定義するものとする。 さて、生物学的種概念において重視される生殖的隔離の機構は、交配前隔 離機構、接合前隔離機構、接合後隔離機構の3 種類に整理されている(Grant, 1985)。交配前隔離機構には、生息場所の違いや、植食性昆虫の場合は食草の違 いなどによる生態的隔離、繁殖の季節の違いや開花の時間の違いなどによる時 間的隔離、ポリネーターの違いや交尾行動の違いなどによる行動的(性的)隔 離、交尾器の形態の不一致やポリネーターと花の形態的対応が不完全であるこ となどによる機械的隔離などが含まれる。接合前隔離には、何らかの原因で精 子が卵子にたどり着くことができない(被子植物の場合、花粉管が胚珠に到達7 できない)配偶子隔離が含まれる。接合後隔離は、接合により雑種の胚が形成 されるものの、雑種の生存率が正常な個体より低いか成熟する前に死亡する、 雑種の生殖力が正常な個体より务るか完全に失われている、雑種の後代子孫の 適応度が正常な個体より低下する(雑種崩壊)などが含まれる。 シダ植物においては、異なる種に分類される個体間で交配させた場合、次 世代の胞子体が発生しない、もしくは不稔の胞子をつける胞子体のみが発生す るという事例が報告されている(Masuyama and Watano 1994; Yatabe et al., 2001)。胞子体が発生しない場合、受精に至るまでのどの段階で生殖的隔離が生 じているのかは、これまでほとんど観察された事例がない。 配偶体の上に造卵 器と造精器がむき出しでついており、水を媒介として精子が造卵器まで泳いで いくという単純な受精様式しかもたないシダ植物において、行動的(性的)隔 離や機械的隔離が生殖的隔離の主たる要因として働いているとは考えにくい。 そのため、精子が造卵器に誘引されない、あるいは誘引されたとしても造卵器 の入り口で精子が死んでしまうなどの原因で受精が起こらないといった接合前 隔離、あるいは受精後の胚発生が起こらないか、発生が途中で停止してしまう といった接合後隔離が起こっていると考えられる。幸い、造卵器がむき出しに なっているシダ植物では、こうした受精から次世代の胞子体発生に至る過程を 観察するのは比較的容易である。ただし、接合後隔離と考えられる現象が観察 できた場合、観察した胚が本当に雑種由来なのかを検証する方法が必要となる。 先行研究において、成長した胞子体が雑種由来か否か判定するにはアロザイム 多型解析が用いられてきた(Masuyama and Watano et al., 1994; Yatabe et al., 2001)。しかし、まだ自力で酵素を生産していない 1 細胞の受精卵や数細胞のご く小さな胚ではアロザイム多型解析による両親判定が不可能である。そのため、 PCR 法などを用いてシグナルを増幅させることで 1 細胞からでも両親判定が可 能な核DNA マーカーの開発が必要となる。また、DNA はタンパク質よりも安 定な物質であるため、もし核DNA マーカーの開発に成功すれば、成長した胞子 体の両親判定もより行いやすくなると期待される。 本研究の材料として用いたのは、旧世界の熱帯~亜熱帯に広く分布するシ
8
マオオタニワタリ類(チャセンシダ科)である。シマオオタニワタリ類は、い ずれも単葉で、種の分類に必要な形態形質に乏しく、染色体数も報告されてい る限りすべて2n=144 の 4 倍体である(Bir, 1960; Abraham et al., 1962;
Kawakami, 1970; Koul, 1970; Tsai and Shieh, 1983; Yatabe et al., 2001)ため、
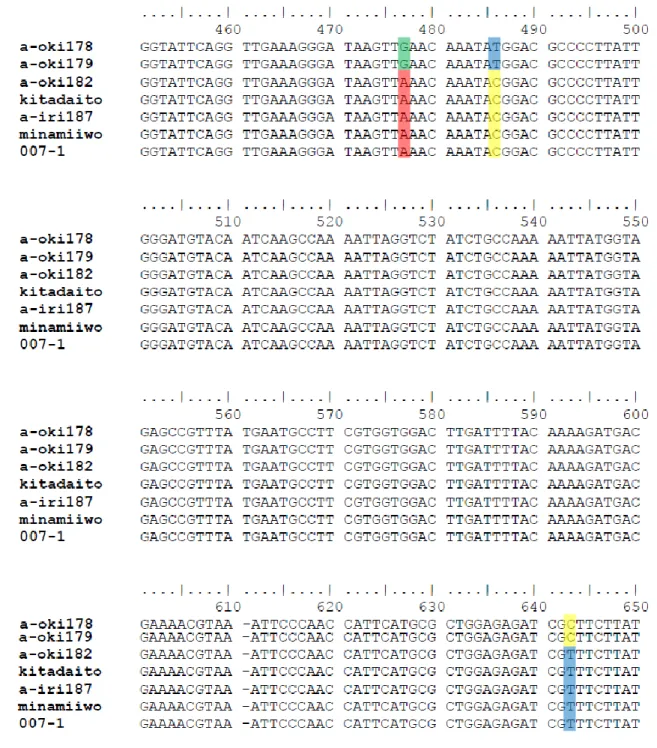
その種の分類は混乱していた。近年になり、葉緑体DNA 上にコードされている 遺伝子の1 つであるrbcL 領域の塩基配列に基づいた分類が行われるようになっ た結果、従来1つの形態的種と認識されてきた集団の中にも、他集団との間に 生殖的隔離が存在し、実質的に異なる生物学的種(隠蔽種)と認められる集団 が複数含まれる場合があることが明らかになった。例えば、Murakami ら(1999a) は、日本に生息するシマオオタニワタリ類のrbcL の塩基配列多型とアロザイム 多型の情報を用いた系統解析を行った結果、先行研究で形態的特徴から Asplenium australasium とされてきた日本産シマオオタニワタリ類の集団は、 オーストラリアで元のタイプ標本が採集されたA. australasium と全く異なる 系統に位置することを解明し、これを新種ヤエヤマオオタニワタリA. setoi と して報告した。またMurakami ら(1999b)は、インドネシア・ハリムン山国 立公園に生育する、従来の分類ですべてシマオオタニワタリAsplenium nidus L. という 1 つの形態種とされていた複数個体のrbcL の塩基配列を解析し、そ れらが3つのrbcL の配列のタイプに分けられ、タイプ間でその生態(生育環境) にも明確な違いが見られることを明らかにした。さらにYatabe ら(2001)は、 Murakami ら(1999b)によってインドネシア・ハリムン山国立公園で認識さ れた、rbcL の塩基配列が異なる 3 タイプのシマオオタニワタリ類の集団間で人 工交配実験を行い、異なるタイプ間では雑種の胞子体が全く得られなかったこ とから、これら3 タイプの間には生殖的隔離があり、互いに隠蔽種の関係にあ ることを明らかにした。さらに、Yatabe ら(2009)は、rbcL 配列が様々な程 度に隠蔽種間で交配実験を行うことで、rbcL 配列の異なっている程度と生殖的 隔離の程度(生じた雑種の胞子体の数ならびにそれらの繁殖能力)には負の相 関があることを明らかにした(Fig. 1-1)。このような研究結果が得られている ことから、シマオオタニワタリ類は、進化の過程でどのような段階を経て生殖
9
的隔離が強まっていき、種分化に至るのかを検証する上で最適な材料だと私は 考えた。
日本国内には、オオタニワタリ(Asplenium antiquum)、シマオオタニワ タリ(Asplenium nidus)、ヤエヤマオオタニワタリ(Asplenium setoi)の3種 のシマオオタニワタリ類が分布しているとされている(Murakami et al., 1999c)。このうち、ヤエヤマオオタニワタリは小笠原と南西諸島の集団との間 に、オオタニワタリは八丈島の集団とその他の地域の集団との間に、かなり大 きなrbcL 配列の隔たりがあり、種分化の途上にあるか、もしくはすでに別々の 種に分化している可能性もあるとMurakami ら(1999a)により発表されてい た。しかし、先行研究で発表されているrbcL の塩基配列は、当時の解析技術の 未発達さから、その精度に不安があり、本当にそれらの集団間に大きなrbcL の 塩基配列の違いがあるのか不確かであった。したがって日本産シマオオタニワ タリ類の系統学的・分類学的実態も未だ不明確なままであった。そのため、本 研究では日本各地でサンプリングしたシマオオタニワタリ類のrbcL の塩基配列 を解析し、日本産シマオオタニワタリ類の実態の解明を目指した。その上で、 もし従来の分類で同種とされていたものの中にもrbcL の塩基配列が異なるもの が見られた場合、Yatabe ら(2009)の結果とも照らし合わせてそれらの集団が 互いに隠蔽種の関係にある可能性があるか検証する。そして、生殖的隔離が成 立していく途中段階である可能性のあるものが見いだされれば、シマオオタニ ワタリ類の種分化の成立の過程を探る研究の材料として用いるのにも最適であ ると考えた。国内で胞子などの材料を容易に得られることになるからである。 本研究では、シマオオタニワタリ類を材料として用い、シダ植物における 生殖的隔離の成立過程を明らかにすることを最終的な研究目的としている。そ の前段階として、今年度までに 1)日本国内における、種分化の途上にある 可能性のあるシマオオタニワタリ類の集団の探索 2)核DNA マーカーの開発 を行った。
10
2. 材料と方法
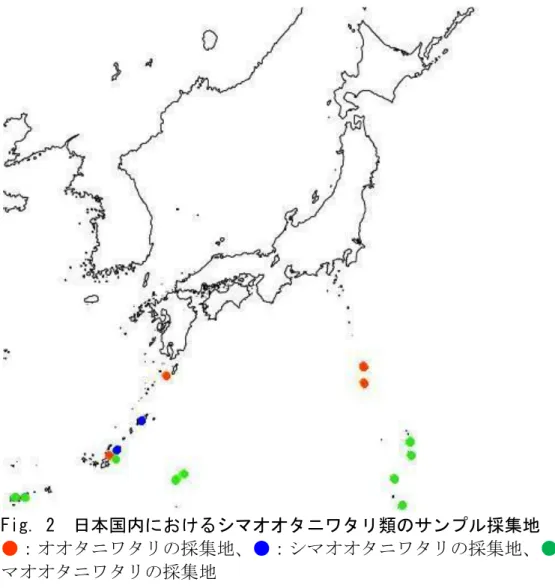
【材料】 日本国内において、オオタニワタリは紀伊半島・伊豆諸島・九州・南西諸 島、シマオオタニワタリは南西諸島、ヤエヤマオオタニワタリは南西諸島と小 笠原諸島にそれぞれ分布しているとされている(倉田・中池 1981; Murakami et al., 1999a)。既に野生集団が絶滅したと思われる場所を除く日本産シマオオ タニワタリ類の分布域をほぼ網羅する形で、国内計15 カ所から、オオタニワタ リ11 個体、シマオオタニワタリ 5 個体、ヤエヤマオオタニワタリ 23 個体を採 集し、材料として用いた(サンプルコード、採集地、採集年月日、採集者の情 報はTable. 1 にまとめた。また採集地の情報は Fig. 2-1 にも示した。)。 これらの各個体から、証拠標本のための胞子嚢群をつけた葉、胞子、シリ カゲル乾燥サンプルを採集した。また、生育していた環境を写真やメモにより 記録した。 【方法】 1)rbcL 遺伝子の塩基配列解析による種分化の途上にある可能性のあるシ マオオタニワタリ類の日本国内における探索日本産シマオオタニワタリ類の植物サンプルよりCTAB 抽出法(Doyle and Doyle, 1987)により全 DNA を抽出した。まず、シリカゲルで乾燥させた葉片 約5mm 四方を直径 5mm のセラミックボール(ニッカトー製)を入れた 2ml のプラスチックチューブに入れ、ミキサーミル(Retsch 製 MM300)で粉砕 した。粉砕したサンプルに抽出用バッファー(2% cetyltrimechylammonium bromide, 0.1M Tris-HCl (pH 8.0), 20mM EDTA (pH 8.0), 1.4M NaCl, 0.1% (v/v) β-メルカプトエタノール)500μl を加え、ヒートブロック(IWAKI 製 LO2-628)を用いて 55℃で 20 分間インキュベートした。そしてクロロホルム・ イソアミルアルコール混合液(24:1 v/v)を 500μl 加え、test tube mixer(SIBATA 製 TTM-1)で撹拌した。その後、微量高速冷却遠心機(HITACHI 製 himac CF 15R)を用いて遠心分離(15000rpm×4 分, 20℃)を行い、上清 350μl を 1.5ml
11 チューブに回収した。上清にイソプロパノール300μl を加え、軽く撹拌して DNA を沈殿させ、再び遠心分離(14000rpm×10 分, 4℃)を行った。遠心後、上清 を捨て、70%エタノール 500μl を加え、再度遠心分離(10000rpm×3 分, 4℃) を行い、上清を捨てた。吸引乾燥機(東京理科機械製 A-3S)で約 10 分間減圧 乾燥を行った後、チューブに100μl の TE バッファー(10mM Tris-HCl, 1mM EDTA (pH8.0))を加え、DNA の沈殿を溶解した。これを PCR 増幅のためのテ ンプレートDNA とした。 その後、rbcL 遺伝子のコード領域 1194bp を PCR 法により増幅した。プ ライマーはHasebe et al. (1994) に記載されたもの(aF, cR)を用いた。PCR 反応液(2×Ampdirect Plus 10μl, 0.5U NovaTaq, 50μM forward primer 0.2 μl, 50μM reverse primer 0.2μl, SDW 9μl)19.5μl にテンプレート DNA 0.5μl を混合し、サーマルサイクラー (Applied Biosystems 製 GeneAmp PCR System 9700)を用いてPCR を行った。PCR 条件は、まず 95℃で DNA の変性 とTaq ポリメラーゼの活性化を 10 分間行った後、94℃30 秒、53℃30 秒、72℃ 3 分のサイクルを 45 回繰り返し、72℃でさらに 7 分間伸長反応を行った。得ら れた産物を、1%アガロースゲルを用いて電気泳動した。100bp ladder もサイ ズマーカーとして同時に流した。 一部で非特異的増幅が見られたため、1000bp 付近に相当する部分をメス で切り出し、QIAquick Gel Extraction Kit (QIAGEN 製) を用いゲルから DNA を精製した。まず、切り出したゲルに対して3 倍量の Buffer QG を加え、50℃ で約10 分間(ゲルが完全に溶解するまで)インキュベートした。そしてゲル と同量のイソプロパノールを混ぜ、これをQIAquick スピンカラムに入れた。 13000rpm×1 分, 20℃で遠心した後、フロースルー液を捨て、500μl の Buffer QG をカラムに添加した。13000rpm×1 分, 20℃で遠心した後、フロースルー 液を捨て、700μl の Buffer PE をカラムに添加し、再び 13000rpm×1 分, 20℃ で遠心した。フロースルー液を捨て、13000rpm×1 分, 20℃で遠心した。10μ l の溶出バッファーをカラムに加え、1 分間放置した後 13000rpm×1 分, 20℃ で遠心した。
12
その後、BigDye Terminator v3.1 Cycle Sequencing Kit (Applied
Biosystem) を用いてシークエンス反応を行った。シークエンス反応に用いた プライマーは、Hasebe et al. (1994) に記載されたもの(aF, aR, cF, cR)およ び、Murakami et al. (1999b) に記載されたシマオオタニワタリ類専用のプラ イマー(OT-NP1, OT-2PR)である。PCR 産物溶液 0.5μl に、反応液(Ready Reaction mix 0.5μl, 1×Bigdye sequencing Buffer 1.75μl, 50μM primer 1μl, SDW 5.75μl)9.5μl を加えた。反応条件は、最初に 96℃で 1 分間変性させた 後、96℃30 秒、50℃5 秒、60℃2 分のサイクルを 25 回繰り返した。シークエ ンス反応産物に70%エタノール/0.5mM MgCl₂ 37μl を加え、スイング型プレ ート遠心機(TOMY 製 Multipurpose Refrigerated Centrifuge LX-120) で 遠心(3500rpm×30 分, 20℃)した後、5 分間減圧乾燥させ、沈殿を 15μl の Hi-Di Formamide (Applied Biosystems 製) で溶かし、95℃2 分でヒートショックを 行った。その後、氷上で約3 分冷却してから DNA シークエンサーABI PRISM 3100 Genetic analyzer(Applied Biosystems 製)により塩基配列を決定した。 得られた配列データをコンピューターソフトウエアChromasPro version 1.34(Technelysium 製)で編集した後、MEGA4 (Tamura 2007) を用いてア ライメントし、分子系統樹を作成した。系統解析には外群としてホウビシダの 配列データを用いて、近隣結合法(Saitou and Nei, 1987)と最節約法(Eck and Daynoff, 1966)の2つの方法で行った。いずれの系統解析法においても、各 枝の統計学的信頼度を知るためにブートストラップ値を計算した。 2)核DNA マーカーの開発 2-1 マイクロサテライトマーカー マイクロサテライトマーカーの開発には、dual-suppression PCR 法(Lian et al., 2006)を用いた。まず、CTAB 法により抽出した母島産ヤエヤマオオタ ニワタリの全DNA 1μl を、制限酵素 EcoRⅤ (15U/μl) と SspⅠ(8U/μl) (TaKaRa 製)で切断した。具体的には、DNA 1~2.5μg を含む DNA 溶液に、 EcoRⅤ 0.5μl もしくは SspⅠ 0.75μl、バッファー(EcoRⅤは 10×H Buffer、 SspⅠは 10×SspⅠ Buffer 各 10μl を使用)を加え、ミリ Q を加えて全体の量
13 を100μl にし、37℃で 12 時間制限酵素反応を行った。その後、クロロホルム・ イソアミルアルコール混合液500μl を加えて酵素を失活させ、エタノール沈殿 処理を行った。沈殿を13μl の TE バッファーに溶解させた。次に 100μM Upper adapter (5’-GTAATACGACTCACTATAGGGCACGCGTGGTCGACGGCCCGGGCTG GT-3’) と 100μM Lower adapter (5’-ACCAGCCC-NH₂-3’) を 0.2ml チューブに 5μl ずつ加えて、サーマルサイクラーで 70℃10 分、37℃10 分処理し、不等長ア ダプターを作成した。別のチューブにLigation kit ver. 1(TaKaRa 製)の A 液 を40μl、B 液を 10μl 加え、次に処理済みの不等長アダプター溶液を 1μl 加えた。 最後に、制限処理後のDNA 溶液を 0.5μg 分だけ加え、サーマルサイクラーを用 いて16℃、12 時間インキュベートした。 アダプターのライゲーション反応の後、70℃で 10 分処理することで酵素を失 活させた。エタノール沈殿処理後、25μl の TE バッファーに溶解し、DNA 量を 測定して超純水を加えて10ng/μl の濃度に調整した。その後、ddGTP 反応溶液 (1×PCR buffer with 1.5mM MgCl₂ 3μl, 100mM ddGTP 1μl, 1.25U
AmpliTaqGold (Applied Biosystems) , SDW 18.75μl)25μl を、ライゲーショ ン処理後のDNA 溶液 25μl に加え、サーマルサイクラーを用いて 94℃9 分、50℃ 10 分、72℃5 分処理を行った。次に、TE バッファー250μl とクロロホルム・イ ソアミルアルコール混合液300μl を加えて酵素を失活させ、エタノール沈殿処 理を行った。その後、50μl の TE バッファーに溶解した。
PCR 反応溶液(1×QIAGEN Multiplex PCR Master Mix 10μl, 20μM SSR compound primer ((AC)₆(AG)₅または(TC)₆(AC)₅) 0.2μl, 20μM AP2 プライマ ー(5’-CTATAGGGCACGCGTGGT-3’) 0.2μl, ミリ Q 9.1μl)19.5μl に ddGTP 処理したDNA 溶液を加えて PCR 反応を行った。反応条件は最初に 95℃15 分 でDNA の変性と Taq ポリメラーゼの活性化を行った後、94℃30 秒の変性、57℃ 1 分 30 秒のアニーリング反応、72℃1 分の伸長反応のサイクルを 40 回繰り返 し、最後に60℃30 分の伸長反応を行った。
14
精製した。まずPCR 反応産物をすべて 1.5ml チューブに移し、超純水を加えて 100μl にした。その後 500μl の Binding Buffer を加えて混ぜ、DNA 溶液を Filter Tube に入れた。14000rpm×1 分, 20℃で遠心後、フロースルー液を捨て、 500μl の Wash Buffer を Filter tube に加え、14000rpm, 1 分, 20℃で遠心した。 フロースルー液を捨て、200μl の Wash Buffer を Filter Tube に加え、14000rpm, 1 分, 20℃で遠心した。フロースルー液を捨て、50μl の Elution Buffer を加え、 1 分間放置した後 14000rpm×1 分, 20℃で遠心した。溶出液にエタノール沈殿 処理を行い、10μl の TE バッファーに溶解した。これを 1.5%アガロースゲル にロードして100V で約 15 分間電気泳動を行った。ゲルの両端には 100bp ladder をサイズマーカーとして入れた。泳動後、トランスイルミネーター上で 400bp~800bp の DNA が存在すると思われる箇所をメスで切り出し、QIAquick Gel Extraction Kit を用いて精製した。
クローニング用のLBプレートの作成は、まず Bacto tryptone 6g, Bacto yeast extract 3g, NaCl 6g を蒸留水に溶かし、1N NaOH 0.6ml を加えた。次に寒天 9g を加え、蒸留水で 600ml までメスアップした後、三角フラスコに移し、オー トクレーブ(TOMY 製 BS-245)を用いて 121℃15 分で加熱し、滅菌と寒天の 溶解を行った。そしてオートクレーブ後の三角フラスコを55℃のウォーターバ スに浸し、55℃まで冷ました。これに 20mg/ml アンピシリンナトリウム水溶液 3ml, 20mg/ml X-gal 溶液 1.2ml、23.8mg/mlIPTG 溶液 3ml を加え、混合した。 滅菌シャーレ(9cm×2cm)に分注し、30 分放置した後、シャーレを上下逆さ まにして4℃で保存した。
クローニングはQIAGEN PCR Cloningplus Kit (QIAGEN) を用いた。まず
PCR 産物を、分光光度計(フナコシ製、UV1101 Biotech Photometer)を用い て定量し、65ng 分だけ 0.2ml チューブに移し、超純水を加えて 4μl にした。 pDrive Cloning Vector を 1μl、2×Ligation Master Mix を 5μl それぞれ加え て混ぜ、16℃で 2 時間インキュベートした。QIAGEN EZ Competent Cell の入 ったチューブ1 本あたり 2μl のライゲーション反応液を加え、静かに混ぜた。 氷上で5 分間放置した後、42℃のウォーターバスで 30 秒間加熱した。再び氷上
15 で2 分間放置した後、キット付属の SOC 培養液を 250μl ずつ各チューブに加 えた。クリーンベンチで120μl、100μl、残り(およそ 80μl)をそれぞれ違う LB プレートにプレーティングした。パラフィルムを巻き、上下逆さまにして LB プレートをインキュベーター(ヤマト科学製 IC43)の中で、37℃で約 12 時間インキュベートした。その後、4℃で約 4 時間インキュベートした。プレー トに出来た白色コロニーを10μl 用チップでつつき、50μl の超純水中で撹拌し た。この溶液を95℃、5 分でヒートショックを行った後、氷上で 5 分間冷却し た。 コロニーが溶けた溶液0.5μl に PCR 反応用混合液(10×PCR Buffer 1μl, dNTP 1μl, 50μM M13-forward primer (5’-CGCCAGGGTTTTCCCAGTCACGAC-3’) 0.1μl, 50μM M13-reverse primer (5’-CGGATAACAATTTCACACAGGAAAC-3’) 0.1μl, 0.25U AmpliTaq Gold (Applied Biosystems 製) , SDW 7.25μl)10μl を加えて、インサートの PCR 増幅を行った。PCR の反応条件は、最初に 94℃15 分で変性させた後に、 94℃30 秒で変性、54℃30 秒でアニーリング、72℃1 分 15 秒の伸長反応を 40 回繰り返し、最後に72℃4 分の伸長反応を行った。PCR 産物に ExoSAP-IT(usb 製)0.15μl、滅菌水 2.85μl を加え、37℃30 分、80℃20 分で精製を行った。 そして、精製済みのPCR 産物に対してシークエンス反応と精製を行い、イン サートの塩基配列の決定を行った。
得られた塩基配列データはコンピューターソフトウエアのABI PRISM DNA SEQUENCING ANALYSIS 5.1.1 (Applied Biosystems 製) で編集を行った。 MEGA 4 でアライメントを行い、同じ配列があれば片方を除いた。そして OLIGO ver.6 (Molecular Biology Insights, Inc) を用いて compound primer と
対になるPCR 用プライマー(領域特異的プライマー)を設計した。プライマー
は、PCR 産物の長さが 50-300bp であること、Tm 値が 55℃-65℃になることに 留意して設計を行った。設計したプライマーはシグマ アルドリッチ ジャパ ン株式会社に注文して作成した。
16
んだ16 個体の全 DNA を用いてスクリーニングを行った。全 DNA 溶液 0.5μl に反応液(1×QIAGEN Multiplex PCR Master Mix 3μl, 20μM compound primer 0.06μl, 20μM 領域特異的プライマー0.06μl)の入ったマイクロチュ ーブに分注し、PCR 増幅を行った。compound primer の(AC)6(AG)5は6-FAM、
(TC)6(AC)5はVIC の蛍光を添加した。反応条件は、95℃15 分で変性させた後、
94℃30 秒の変性、52℃1 分 30 秒のアニーリング反応、72℃45 秒の伸長反応を 24~26 サイクル行い、最後に 60℃30 分の伸長反応を行った。PCR 産物を 1μl ずつシーケンス用プレートに入れ、Hi-Di Formamide 12μl、GeneScan-500LIZ (Applied Biosystems 製) 0.2μl をそれぞれ加え、95℃2 分でヒートショック処 理を行ってから、ABI PRISM 3100 Genetic Analyzer にセットし、フラグメン ト解析を行った。
得られた波形データはGENEMAPPER ver. 4.0 (Applied Biosystems 製) で
解析を行い、マーカーの有効性を調べた。マーカーを選択する基準として、PCR
増幅していること、変異(多型)が見られること、遺伝子型を決定できること の3 点に基づいてスクリーニングを行った。
2-2 gapCp を用いた共優性マーカーの開発
まず、CTAB 法により抽出した母島産ヤエヤマオオタニワタリの全 DNA 0.5 μl に PCR 反応液(2×Ampdirect Plus 10μl, 0.5U NovaTaq, 50μM
ESGAPCP8F1(5’-ATYCCACGYTCAACTGGTGCTGC-3’) 0.2μl, 50μM ESGAPCP11R1(5’-GTATCCCCAYTCRTTGTCRTACC) 0.2μl, SDW 9μl) 19.5μl を加え、PCR 反応を行った。反応条件は1)で述べたものと同様である。
得られたPCR 反応産物を、1%アガロースゲルを用いて電気泳動し、バンドが
見られた部位をメスで切り出し、QIAquick Gel Extraction Kit を用いて精製し た。そして、精製済みのPCR 産物に対してシークエンス反応と精製を行った後、 塩基配列の決定を行った。
得られた配列データ(901bp)を NCBI に登録された植物のgapCp の塩基配 列のデータと比較し、シダ植物のgapCp の塩基配列であることを確認した。そ
17
の後、得られた配列を元にOLIGO ver.6 (Molecular Biology Insights 製) を用 いて2 対のプライマー(101F, 101R, 103F, 103R)を設計した。プライマーは、 forward, reverse 共に Tm 値が 55℃-65℃になることに留意して設計を行った。 設計したプライマーはシグマ アルドリッチ ジャパン株式会社に発注して作 成した。
設計したプライマーのスクリーニングのために、日本産シマオオタニワタリ 類の各種・各地域から選んだ16 個体から、QIAGEN DNeasy Plant Mini Kit (QIAGEN 製)を用いて、再度、全 DNA を抽出した。これはより純度の高い DNA を得るためである。まず、約 5mm 角のシリカゲル乾燥サンプルを CTAB 抽出のときと同様にミキサーミルで破砕し、400μl の Buffer AP1 と 4μl の RNase A ストック溶液を添加し、激しく混合した。混合液を 65℃のヒートブロ ックで10 分間インキュベートした。インキュベート中に 2,3 回チューブを転 倒混和した。130μl の Buffer AP2 を混合液に添加後、軽く混和し、5 分間氷上 でインキュベートした後、14000rpm×5 分、20℃で遠心した。上清を 2ml コレ クションチューブ中にセットしたQIAshredder Mini スピンカラムにアプライ し、14000rpm, 2 分, 20℃で遠心した。濾液を 1.5ml チューブに移し、1.5 倍量 のBuffer AP3/E を添加し、ピペッティングで混合した。混合液 650μl を 2ml コレクションチューブ中のDNeasy Mini スピンカラムに添加し、8000rpm×1 分, 20℃で遠心し、濾液を捨てた。カラムに残りの混合液を加え、同様に遠心し た。カラムを新しい2ml コレクションチューブに移し、Buffer AW 500μl を添 加し、8000rpm×1 分, 20℃で遠心した。濾液を捨て、Buffer AW 500μl をカラ ムに添加し、14000rpm×2 分, 20℃で遠心し、メンブレンを乾燥させた。カラ ムを新しい1.5ml チューブに移し、100μl の Buffer AE をメンブレン上に添加 し、室温で5 分間放置した後、8000rpm×1 分, 20℃で遠心し、メンブレンに吸 着していた全DNA を溶出した。
このようにして得られた全DNA 溶液 1μl に反応液(2×Ampdirect plus 10 μl, 0.5U Z-Taq, 10μM forward primer 0.5μl, 10μM reverse primer 0.5μl) 19.5μl を加え、PCR 反応を行った。反応条件は、94℃で 5 分 DNA を変性させ
18
た後、94℃5 秒、60℃10 秒、72℃20 秒のサイクルを 50 回繰り返し、その後 72℃ で7 分間変性させた。PCR 反応産物を 2%アガロースゲルで電気泳動し、増幅 をチェックした後、増幅が見られたPCR 産物各 4μl に ExoSAP-IT0.8μl と滅 菌水4.2μl を加えて 37℃30 分、80℃20 分で精製した。そして、BigDye Terminator v3.1 Cycle Sequencing Kit を用いて、精製済みの PCR 産物に対し てシークエンス反応を行った。PCR 産物溶液 4.5μl に、反応液(Ready Reaction mix 1μl, 1×BigDye Sequencing Buffer 2μl, 1μM primer 1.6μl, SDW 0.9μ l)9.5μl を加えた。反応条件は、最初に 96℃で 1 分変性させた後、96℃10 秒、 50℃15 秒、60℃4 分のサイクルを 25 回繰り返した。その後、精製を行い、シ ーケンサーを用いて塩基配列の決定を行った。
得られた配列は、NCBI BLAST を利用して、本当に植物のgapCp の配列で あるか確認した。
2-3 pgiC を用いた共優性マーカーの開発
まず、QIAGEN DNeasy Plant Mini Kit を用いて抽出したシマオオタニワタ リ類のDNA16 個体分の全 DNA をそれぞれテンプレートとし、Ishikawa ら (2002)の開発したプライマー14F と 16R(Fig. 2-2)を用いた増幅を試みた。 全DNA 溶液 1μl に、反応液(PrimeSTAR Max 5μl, 10μM 14F 0.2μl, 10μ M 16R 0.2μl, SDW 3.6μl)9.5μl を加え、PCR 反応を行った。反応条件は、 まず98℃で 10 秒 DNA を変性させた後、98℃10 秒、50℃15 秒、72℃5 秒のサ イクルを35 回繰り返し、72℃で 7 分伸長反応を行った。そして、反応液に ExoSAP-IT 0.15μl を加えて 35℃30 分、80℃20 分処理した後、精製済みの PCR 産物に対して1)と同様の方法でシークエンス反応を行い、ABI の 3100 型 DNA シークエンサーを用いて塩基配列の決定を行った。 このようにして得られた配列を元に、Primer3 (version 0.4.0) (http://frodo.wi.mit.edu/primer3/)を用いて新規のプライマー184F, 536R, 97F, 489R を開発した。設計したプライマーは、株式会社ベックスに発注して作 成した。しかし、これらのプライマーを用いたPCR では十分な増幅が見られな
19
かった。
そこで、シマオオタニワタリ類と同じくチャセンシダ科に属するチャセンシ ダ(Asplenium trichomanes)の pgiC 配列(James et al.,2008)と、Ishikawa ら(2002)のプライマー開発に用いられたハチジョウベニシダ(Dryopteris caudipinna)の配列をアライメントし、Ishikawa ら(2002)が開発したプライ マー15F, 15R と相同なチャセンシダのpgiC の塩基配列を元に新規のプライマ ーAsp15F, Asp15R(Fig. 2-3)を設計した。設計したプライマーを Ishikawa ら (2002)が開発したプライマーと組み合わせ、プライマーの種類が異なる 2 種 類の反応液(2×Ampdirect plus 10μl, Z-Taq 0.2μl, 10μM 14F 0.5μl, 10μM Asp15R 0.5μl, SDW 7.8μl)と(2×Ampdirect plus 10μl, Z-Taq 0.2μl, 10 μl Asp15F 0.5μl, 10μl 16R 0.5μl, SDW 7.8μl)にテンプレート DNA1μl を加え、PCR 反応を行った。2%アガロースゲルで PCR 産物を泳動し、増幅を 確認したところ、Asp15F と 16R の組み合わせで PCR 反応を行った場合の一部 のシマオオタニワタリ類のサンプルについて増幅が見られた。そこで、増幅が 見られたPCR 産物 4μl に ExoSAP-IT0.8μl と滅菌水を加え、37℃15 分、80℃ 15 分で反応させ、精製した。精製済みの PCR 産物に対して(2-2)と同様 の方法でシークエンス反応を行い、塩基配列の決定を行った。決定できた配列 は、シダ植物のpgiC の一部であることをその相同性から確認し、16R の内側の 位置にプライマー471R, 481R (Fig. 2-2) を設計した。プライマーの設計には Primer3 (version 0.4.0) を用いた。プライマーの設計の際には、なるべく長い 配列を増幅できること、シークエンスできたサンプル同士で高い保存性が見ら れることに留意した。設計したプライマーは株式会社ベックスに発注して作成 した。 設計したプライマーを用いて変異のある配列を得られるか、QIAGEN DNeasy Plant Mini Kit を用いて抽出したシマオオタニワタリとヤエヤマオオ タニワタリの全DNA 16 個体分を用いて PCR 反応を行い確かめた。反応液(2 ×Ampdirect plus 10μl, Z-Taq 0.2μl, 10μM Asp15F 0.5μl, 10μM 471R 0.5 μl, SDW 7.8μl)と(2×Ampdirect plus 10μl, Z-Taq 0.2μl, 10μl Asp15F 0.5
20 μl, 10μl 481R 0.5μl, SDW 7.8μl)各 19.5μl にテンプレート DNA1μl を加 え、PCR 反応を行った。2%アガロースゲルで PCR 産物を泳動し、増幅を確認 したところ、いずれのプライマーの組み合わせでもほとんどのサンプルで増幅 が見られた。増幅が見られたPCR 産物を ExoSAP-IT を用いて精製した。精製 済みのPCR 産物に対して2-2と同様にシークエンス反応を行い、その後精製 と塩基配列の決定を行った。得られた配列データをChromasPro version 1.34 で編集した後、MEGA4 を用いてアライメントし、変異が見られるかチェック した。 その後、シマオオタニワタリ類の中では、ヤエヤマオオタニワタリやシマオ オタニワタリとやや遠縁のオオタニワタリや、オニヒノキシダ、オトメシダ、 ヌリトラノオ、ナンゴクホウビシダなどのシマオオタニワタリ類に比較的近縁 なチャセンシダ科のシダ植物の種の全DNA サンプルをテンプレートに用いた ときにPCR 増幅が可能かも確かめた。
21
3. 結果
1)日本国内における、種分化の途上にある可能性のあるシマオオタニワ タリ類の探索 本研究では、オオタニワタリ4 産地 11 個体、シマオオタニワタリ 1 産地 3 個 体、ヤエヤマオオタニワタリ9 産地 23 個体について、そのrbcL の塩基配列を 決定した(Fig. 3-1)。得られた配列の間の関係が分かるように分子系統樹の形 で示したのがFig. 3-2 である。また、Yatabe ら(2009)によって報告されてい た外国産のシマオオタニワタリ類のrbcL の配列に今回得られた日本産シマオオ タニワタリ類の配列を加え、系統解析した結果を示したのがFig. 3-3 である。 父島・母島産ヤエヤマオオタニワタリのrbcL 配列はすべて同一であったが、 南西諸島産ヤエヤマオオタニワタリは北大東島産の個体とそれ以外の個体の間 で1 塩基の配列の違いが見られた。父島・母島産の個体と北硫黄島産の個体の rbcL の塩基配列の違いは 6 塩基、父島・母島産の個体と北硫黄島産以外の南西 諸島産の個体のrbcL の塩基配列の違いは 5 塩基であった。また、父島・母島産 ヤエヤマオオタニワタリと北硫黄島・南硫黄島(火山列島)産ヤエヤマオオタ ニワタリの間には、rbcL の塩基配列に 2 塩基の違いがあることが明らかになっ た。火山列島産ヤエヤマオオタニワタリの方が系統樹のより根本に位置してお り、南西諸島産ヤエヤマオオタニワタリとのrbcL の塩基配列の差は 3~4 塩基 であった。 また、サンプル採集の際に私自身が行ったフィールドでの生育環境の観察の 結果、南西諸島産のヤエヤマオオタニワタリが主に日当たりの悪い場所の樹木 や岩に着生していたのに対し、小笠原諸島・母島のヤエヤマオオタニワタリは 地面から直接生えていたり、開けた草原にも見られたりする場合があるなど、 生態的にも差異が見られた(Fig. 3-4, 3-5)。 一方、Murakami ら(1999c)による先行研究で、八丈島産と紀伊半島、九 州、沖縄本島産の個体間で大きなrbcL の塩基配列の違いがあると報告されてい たオオタニワタリは、沖縄本島で採集したものが他の集団との間で1 塩基の rbcL の塩基配列の違いが見られた他は、目立った違いは見られなかった。今回、22 確実に野生のものである八丈島産のオオタニワタリのサンプルは得られなかっ たが、八丈島に地理的に比較的近い青ヶ島のオオタニワタリについては、屋久 島のものと全く同一のrbcL 配列が得られた。 2)核DNA マーカーの開発 2-1 マイクロサテライトマーカー プラスミドの挿入が確認された大腸菌のコロニー176 個をピックアップして シークエンスし、34 のマイクロサテライト領域を増幅できるプライマーを設計 した。これらのプライマーをスクリーニングしたところ、いずれもPCR 増幅が 見られない、あるいはアレルの判読が不可能である(Fig. 3-6)などの理由で、 残念ながら遺伝マーカーとして使用できるものは得られなかった。 2-2 gapCp を用いた共優性マーカーの開発 新規に開発したプライマー101F, 101R, 103F, 103R を用いた PCR により、3 サンプルで200bp 前後の長さの塩基配列が得られた。しかし、NCBI BLAST search を用いて類似の登録済み塩基配列を検索した結果、得られた配列は植物 よりも菌類のgapCp の配列に近いという結果となり、作成したプライマーでは シダ植物のgapCp の配列を特異的に PCR 増幅させることはできなかった。 2-3 pgiC を用いた共優性マーカーの開発
本研究で新規に開発したForward 側の PCR プライマーAsp15F と Reverse 側の471R および 481R の組み合わせにより、500bp 前後の配列を得ることがで きた。NCBI BLAST search により、この配列がシダ植物のpgiC の配列にほぼ 間違いないことが確認できた。そこで、得られたpgiC の配列をアライメントし て系統解析を行い、最節約系統樹の形で示したのがFig. 3-4 である。確実に配 列の違いが見られた箇所を数えたところ、シマオオタニワタリ(沖縄本島産) と南西諸島産ヤエヤマオオタニワタリの間の配列の違いが13bp 以上、南西諸島 産ヤエヤマオオタニワタリと母島産ヤエヤマオオタニワタリの間の配列の違い
23 が22bp 以上、母島産ヤエヤマオオタニワタリと火山列島産ヤエヤマオオタニワ タリの間の配列の違いが15bp 以上見られた(Fig. 3-7, Fig. 3-8)。ちなみに火山 列島産ヤエヤマオオタニワタリと南西諸島産ヤエヤマオオタニワタリの間の配 列の違いは7bp で、南西諸島産ヤエヤマオオタニワタリと母島産ヤエヤマオオ タニワタリの間の配列の違いより小さかった。これはrbcL の配列による系統解 析とも一致した結果であった。 また、シマオオタニワタリの配列に基づいて設計したプライマーAsp15F と 471R および 481R の組み合わせで、シマオオタニワタリ類の中ではシマオオタ ニワタリ、ヤエヤマオオタニワタリ、オオタニワタリ、さらにオニヒノキシダ、 オトメシダ、ナンカイシダなどのシマオオタニワタリ類には含まれないものの、 比較的近縁なチャセンシダ属の一部の種でもPCR 増幅が可能であることが確認 できた。
24
4. 考察
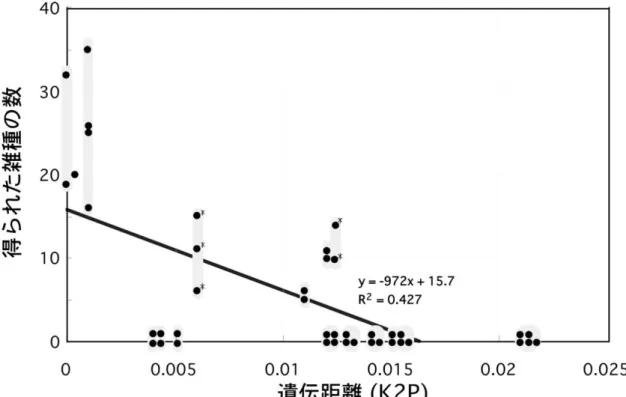
1)日本国内に種分化の途上にあると考えられるシマオオタニワタリ類の 集団は存在するか? 今回、小笠原諸島(父島・母島・北硫黄島・南硫黄島)産ヤエヤマオオタニ ワタリと、南西諸島(沖縄本島・石垣島・西表島・北大東島・南大東島)産ヤ エヤマオオタニワタリの間には、そのrbcL 配列に 5~6 塩基の違いがあること が明らかとなった。 Yatabe ら(2009)の報告により、rbcL の塩基配列の違いが 5~6 塩基見られ る(K2P により求めた遺伝的距離が 0.005 程度)集団間では、人工交配実験を 行っても雑種の胞子体が発生しない、もしくは発生してもそれらが不稔である (稔性のある胞子を形成できない)場合があることが報告されている(Fig. 1-1)。 そのため、今回5~6 塩基のrbcL の塩基配列の違いが見られた南西諸島産ヤエ ヤマオオタニワタリと小笠原諸島(特に父島・母島)産ヤエヤマオオタニワタ リの間にも、部分的にでも生殖的隔離が成立している可能性がある。 また、従来から認識されているシマオオタニワタリ類3 種の間には、生殖的 隔離が存在する可能性は十分にあるが、これら3 種間でどの程度の段階の生殖 的隔離が起きているのか検証された事例はない。そこで、DNA 解析用サンプル と同時に採集した胞子を用いてさまざまな日本産シマオオタニワタリ類同士の 組み合わせでの人工交配実験を現在行っているところである。異なる集団間の かけ合わせで胞子体の発生が見られたら、新規に開発したpgiC マーカーを用い て、得られた胞子体の両親判定を行う予定である。 2)小笠原諸島産シマオオタニワタリ類の起源 今回、rbcL を用いた系統解析の結果、火山列島の南硫黄島・北硫黄島産ヤエ ヤマオオタニワタリと父島列島・母島列島の父島・母島産ヤエヤマオオタニワ タリの間にはrbcL の塩基配列に 2 塩基程度の差があること、分子系統樹(Fig. 3-1)では父島・母島集団の方が派生的な位置にあり、火山列島集団の方がより 根本に近い位置にあることが明らかとなった。pgiC の塩基配列の比較でも同様25 の結果が得られた。これらの結果から、父島・母島集団と火山列島集団は互い に近縁ではあるものの、異なる起源を持っている可能性も考えられる。これら 小笠原諸島産のヤエヤマオオタニワタリの由来として、以下のような可能性が 考えられる。1)父島・母島に南西諸島からヤエヤマオオタニワタリが進入し た後、より島としての起源が新しい火山列島に独立に南西諸島からヤエヤマオ オタニワタリが侵入した。 2)南西諸島から父島・母島に進入したヤエヤマ オオタニワタリの一部が火山列島に侵入し、その後両集団の遺伝的な交流が途 絶え、rbcL の塩基配列に違いが生じた。 3)南西諸島以外の地域に生息する ヤエヤマオオタニワタリが小笠原諸島の父島・母島と火山列島に別々に侵入し た。 などが考えられる。父島・母島列島の起源は数千~数百万年前、火山列 島の起源は十数万年前と言われており(海野ほか、2009)、100 万~500 万年に 1 塩基程度といわれているrbcL の進化速度(Yatabe et al., 1999)と火山列島 の成立の新しさを考慮すると、上記の3つの仮説のうち2)もしくは3)の可 能性が高いと考えられる。 また、Fig. 3-3 を見ると、南西諸島産ヤエヤマオオタニワタリとほぼ同じrbcL の塩基配列をもつ個体がインドネシアのジャワ島で採集されている。したがっ てrbcL の塩基配列に基づいた系統樹上でヤエヤマオオタニワタリと同じクレー ドに属するシマオオタニワタリ類の集団がかなり広範囲にわたって分布してい る可能性がある、すなわち小笠原諸島産ヤエヤマオオタニワタリの起源が海外 の集団にある可能性も十分にあることになる。小笠原諸島産ヤエヤマオオタニ ワタリと同一のrbcL の塩基配列を持つ海外産シマオオタニワタリ類はまだ見つ かっていない。しかしながら、海外産のヤエヤマオオタニワタリを含むシマオ オタニワタリ類については、まだ調査が十分でない地域の方が多く、そういっ た地域に南西諸島集団よりさらに小笠原諸島の集団に近縁なヤエヤマオオタニ ワタリが見いだされる可能性も考えられる。今後、小笠原諸島に見られるヤエ ヤマオオタニワタリの起源地である可能性のある海外の地域(台湾、中国南部、 フィリピン、北マリアナ諸島、インドネシア、マレーシア、ハワイ、フィジー など)からもシマオオタニワタリ類のサンプルを集め、それらのrbcL の塩基配
26 列の解析などを行うことも考えている。 また、サンプル採集の際にフィールドで行った観察の結果、小笠原諸島産ヤ エヤマオオタニワタリが、南西諸島においてヤエヤマオオタニワタリの生育が 見られないような乾燥した環境を含む様々な環境で生育していることが明らか になった。これは、小笠原という比較的乾燥した海洋島における適応進化の結 果とも考えられるが、小笠原諸島のヤエヤマオオタニワタリが南西諸島の集団 とは異なる起源をもつことを反映している可能性もある。 3)pgiC マーカーの有用性 今回開発したpgiC 領域の DNA マーカーは、rbcL の塩基配列の違いがごく 小さなシマオオタニワタリ類の集団間でも変異を検出することが可能であり、 またシマオオタニワタリ類以外のチャセンシダ科の近縁種群で同じプライマー を用いたPCR 増幅が可能であった。したがって、シマオオタニワタリ類全般に おいて核マーカーとしての使用が可能であると考えられる。現在、人工交配実 験で用いる予定の株のうち、異なるpgiC の配列をヘテロでもつと思われる個体 の各アリルの配列を解読する作業を行っている。親個体のアリルがすべて特定 できたら、1 粒の花粉から複数の遺伝マーカー(マイクロサテライトマーカー) をPCR 増幅する技術(Isagi and Suyama ed. 2011)などを参考にして、シダ 植物のごく小さな胚からDNA マーカーを PCR 増幅する技術を確立し、人工交 配させた後の受精卵や胚の親子判定に用いる予定である。 また、本研究で開発したプライマーにより増幅できるpgiC の塩基配列は、核 DNA でありながら比較的塩基の挿入・欠失が尐ないことから、核遺伝子を用い た系統解析にも利用できると考えている。日本産のみならず海外産のシマオオ タニワタリ類のサンプルも含めた核DNA にもとづく分子系統解析を行い、これ までに報告されている葉緑体DNA にコードされているrbcL 遺伝子の情報に基 づく分子系統樹と比較することも考えている。それは前述した小笠原諸島のヤ エヤマオオタニワタリの起源を解明する研究にもつながる可能性がある。なぜ なら、核DNA にコードされているpgiC の方が葉緑体 DNA よりも変異が多く
27 見られ、情報量がより多いため、より解像度の高いデータを得られると考えら れるからである。 3)gapCp マーカーの開発 今回、gapCp の領域を用いた核マーカー開発に失敗したのは、Schuettpelz ら(2008)の論文に、チャセンシダ科に属するホウビシダのgapCp がシングルコ ピーであったと述べられていたため、シマオオタニワタリ類のgapCp もシング ルコピーであると私が誤解していたためである可能性がある(実際はlong と short の 2 つのコピーが存在する)。そのため、縮重プライマーを用いてアニー リング温度を下げて増幅したgapCp のアガロースゲル中のバンドがスメア状に なっているのを1 本のバンドが見えていると勘違いしてバンドの切り出しをし てしまったこと、またプライマーを設計する際に保存性の高い領域を探さずに 適当な位置に設計してしまったことなどが失敗の原因である可能性が考えられ る。縮重プライマーでも、非特異的な増幅を起こさない適切なアニーリング温 度でPCR 増幅し、クローニングして long と short それぞれの塩基配列を特定 し、保存性の高い部位にlong と short それぞれに特異的な増幅が可能な PCR プライマーを設計すれば、gapCp も核マーカーとして、あるいは系統解析に用 いる核DNA の塩基配列情報として有効に用いることができる可能性が十分に ある。今後、gapCp マーカーの開発にも再チャレンジしたいと考えている。
28
5.今後の展望
今年度までの研究で、日本産シマオオタニワタリ類の中に新たな隠蔽種、あ るいはまだ生殖的隔離が完全に成立しておらず、種分化の途上にある可能性の ある集団も発見し、そして今後行う生殖的隔離の成立の過程を調べる研究にお いて必須となる核DNA マーカーの開発も行うことができた。特に核 DNA マー カーの開発は、今後行う予定の人工交配実験の結果、雑種の胞子体の発生が見 られない場合、受精から次世代胞子体の発生に至る過程のどこで生殖的隔離が 働いているかを実体あるいは顕微鏡下での観察により明らかにする研究を行う 上で重要なポイントとなると考えている。また、日本産シマオオタニワタリ類 の中で新たな種に分化しつつある途上にあると思われる集団が発見されたこと で、今後の実験の材料を国内で入手できる可能性が示された。とはいえ、日本 産シマオオタニワタリ類の起源を探る上でも、海外、その中でも特に日本列島 をとりまく亜熱帯~熱帯地域の材料を採集し、解析することも必要である。そ うすることで、さらに種分化途上の異なる段階にある、ヤエヤマオオタニワタ リを含むシマオオタニワタリ類が得られる可能性もあるからである。このよう な材料も利用して、シダ植物の種分化(生殖的隔離の進化)過程を解明してい ければと考えている。29
6.謝辞
本研究を行うに当たり、忍耐強く助言とサポートをして下さった植物系統分 類学研究室の村上哲明教授に厚く御礼申し上げます。 東京大学理学研究科附属植物園の角川(谷田辺)洋子博士には、シダ植物を 用いた各種の実験の手法について色々とご教授いただきました。 また、植物系統分類学研究室の加藤英寿博士には、フィールドワークの方法 の指導ならびにサンプル採集のお手伝いをしていただきました。加藤朗子氏に は、サンプルの採集や研究室生活における指導など様々な面で大変お世話にな りました。マイクロサテライトマーカーの開発においては、森啓悟氏に手法を 教えていただきました。さらに、圃場の杉山栄徹氏、安木孝子氏には、材料植 物の栽培において多大なご協力をいただきました。 また、沖縄本島での調査の際には地元の高良拓夫氏に大変お世話になりまし た。さらに、小笠原諸島での調査の際には、小笠原総合事務所国有林課の方々、 環境省自然保護官並びに東京都自然職員の方々に大変お世話になりました。 そのほか、植物系統分類学研究室の皆様には、サンプルの採集、実験の助言 など、研究のあらゆる面においてお世話になりました。 皆様にこの場を借りて深く御礼申し上げます。30
引用文献
Abraham, A., Ninan, C. A. and Mathew, P. M. 1962. Studies on the cytology and phylogeny of the pteridophytes. Journal of the Indian Botanical Society 41: 339-421
Bir, S. S., 1960. Cytological observations on the East Himalayan members of Asplenium Linn. Current Science 29: 445-447
Cracraft, J. 1989. Species as Entities of Biological Theory. What the philosophy of biology is 31-52
Doyle, J. J. and Doyle, J. L. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin 19, 11-15
Eck, R. V. and Daynoff, M. O. 1966. In: Daynoff, M. O. (ed. ), Atlas and Protein Sequence and Structure. National Biomedical Research Foundation, Silver Springs Md.
Grant, V. 1985. The Evolutionary Process. Columbia Univ. Press, N. Y. Hasebe, M., Omori, T., Nakazawa, M., Sano, T., Kato, M. and Iwatsuki, K.
1994. rbcL gene sequences provide evidence for the evolutionary lineages of leptosporangiate ferns. Evolution 91: 5730-5734 Isagi, Y. and Suyama, Y. 2011. Single-Pollen Genotyping Springer
Ishikawa, H., Watano, Y., Kano, K., Ito, M. and Kurita, S. 2002. Development of primer sets for PCR amplification of the PgiC gene in ferns. Journal of Plant Research 115: 65-70
James, K. E., Schneider, H., Ansel, S. W., Evers, M., Robba L., Uszynski, G., Pedersen, N., Newton, A. E., Russell, S. J., Vogel, J. C. and Kilian, A. 2008. Diversity Arrays Technology (DArT) for Pan-Genomic
Evolutionary Studies of Non-Model Organisms. PLoS ONE 3: e1682. doi: 10.1371/journal. pone. 0001682
Kawakami, S., 1970. Karyological studies on Aspleniaceae. Ⅱ.
Chromosomes of seven species in Aspleniaceae. Botanical Magazine Tokyo 83: 74-81
31
spider plant. Genetica 41: 305-310
Lian, C., Wadud, M. A., Geng,Q., Shimatani, K. and Hogetsu, T. 2006. An improved technique for isolating codominant compound microsatellite markers. Journal of Plant Research 119: 415-517
Masuyama, S. and Watano, Y. 1994. Hybrid sterility between two isozymic types of the fern Ceratopteris thalictroides in Japan. Journal of Plant Research 107: 269-274
Mayr, E. 1942. Systematics and the Origin of Species. Columbia University Press, New York
Murakami, N., Watanabe, M., Yokoyama, J., Yatabe, Y., Iwasaki, H. and Serizawa, S. 1999a. Molecular Taxonomic Study and Revision of the Three Japanese Species of Asplenium sect. Thamnopteris. Journal of Plant Research 112: 15-25
Murakami, N., Yatabe, Y., Iwasaki, H., Darnaedi, D. and Iwatsuki, K. 1999b. Molecular α-Taxononomy of a Morphologically Simple Fern
Asplenium nidus Complex from Mt. Halimun National Park, Indonesia. In: Kato M, (ed. ) The biology of biodiversity Berlin: Springer, 53-66
Saitou, N and Nei, M. 1987. The neighbor-joining method; A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution 4: 406-425
Schuettpeltz, E., Grusz, A. L., Windham, M. D. and Pryer, K. M. 2008. The Utility of Nuclear gapCp in Resolving Polyploid Fern Origin.
Systematic Botany 33: 621-629
Tamura, T., Dudley, D., Nei, M. and Kumar, S. 2007. MEGA4: Molecular Evolutinonary Genetics Analysis (MEGA) Software Version 4.0. 2007. Molecular Biology and Evolution 24: 1596-1599
Tsai, J. L. and Shieh, W. C. 1983. A cytotaxonomic survey of the
pteridophytes in Taiwan. Journal of Scientific Computing 20: 137-158 Van Valen, L. 1976. Ecological Species, Multispecies, and Oaks. Taxon Vol.
32
25, No. 2/3, 233-239
Yatabe, Y., Masuyama, S., Darnaedy, D. and Murakami, N., 2001. Molecular systematics of the Asplenium nidus complex from Mt. Halimun
national park, Indonesia: Evidence for reproductive isolation among three sympatric rbcL sequence types. American Journal of Botany 88: 1517-1522
Yatabe, Y., Nishida, H. and Murakami, N. 1999. Phylogeny of osmundaceae inferred from rbcL nucleotide sequences and comparison to the fossil evidences. Journal of Plant Research 112: 397-404
Yatabe, Y., Shinohara, W., Matsumoto, S. and Murakami, N. 2009. Patterns of hybrid formation among cryptic species of bird-nest fern, Asplenium nidus complex (Aspleniaceae), in West Malesia. Botanical Journal of Linnean Society 160: 42-63 海野進・中野俊・石塚治・駒澤正夫.2009. 20 万分の 1 地質図幅「小笠原諸島」 独立行政法人産業技術総合研究所地質調査総合センター,つくば 岩槻邦夫・加藤雅啓.2000. 多様性の植物学Ⅰ 植物の世界 東京大学出版会, 東京 岩槻邦夫・馬渡峻輔.1996. 生物の種多様性 裳華房,東京 高山浩司・加藤英寿.2010. 遙かな波路を越え、たどり着いた植物たち.i-Bo 20: 6-7 清水健太郎・長谷部光泰.2007. 植物の進化 基本概念からモデル生物学を活 用した比較・進化ゲノム学まで 秀潤社,東京 倉田悟・中池敏之編.1981. 日本のシダ図鑑 2 東京大学出版会,東京
33 Table 1 研究材料の種名、サンプルコード、採集地、採集年月日ならびに採集 者 サンプルコー ド 種名 採集地 採集年月 日 採集者 chichi001 ヤエヤマオオタニワタリ 東京都小笠原村父島東 平 2008/12/1 山田香菜子 higashidaira ヤエヤマオオタニワタリ 東京都小笠原村父島東平 2008/7/5 加藤英寿 otaki ヤエヤマオオタニワタリ 東京都小笠原村父島大滝 2008/7/5 加藤英寿 haha007-1 ヤエヤマオオタニワタリ 東京都小笠原村母島乳房山 2008/12/3 山田香菜子 haha037 ヤエヤマオオタニワタリ 東京都小笠原村母島石門 2008/12/4 山田香菜子 haha038 ヤエヤマオオタニワタリ 東京都小笠原村母島石 門 2008/12/4 山田香菜子 haha084 ヤエヤマオオタニワタリ 東京都小笠原村母島乳房山 2008/12/6 山田香菜子 haha088-1 ヤエヤマオオタニワタリ 東京都小笠原村母島乳房山 2008/12/6 山田香菜子 haha1 ヤエヤマオオタニワタリ 東京都小笠原村母島石門 2008/5/16 加藤朗子 haha2 ヤエヤマオオタニワタリ 東京都小笠原村母島石門 2008/5/16 加藤朗子 haha3 ヤエヤマオオタニワタリ 東京都小笠原村母島石 門 2008/5/16 加藤朗子 haha4 ヤエヤマオオタニワタリ 東京都小笠原村母島石門 2008/5/16 加藤朗子 haha5 ヤエヤマオオタニワタリ 東京都小笠原村母島石門 2008/5/16 加藤朗子 kitaiwo014 ヤエヤマオオタニワタリ 東京都小笠原村北硫黄 島 2009/6/17 加藤英寿 kitaiwo061 ヤエヤマオオタニワタリ 東京都小笠原村北硫黄島 2009/6/19 加藤英寿 minamiiwo ヤエヤマオオタニワタリ 東京都小笠原村南硫黄 島 2007/6/25 加藤英寿 daitom507 ヤエヤマオオタニワタリ 沖縄県島尻郡北大東村 2008/6/27 加藤英寿 daito509 ヤエヤマオオタニワタリ 沖縄県島尻郡南大東村 2008/6/28 加藤英寿 daito514 ヤエヤマオオタニワタリ 沖縄県島尻郡南大東村 2008/6/28 加藤英寿 oki177 ヤエヤマオオタニワタリ 沖縄県国頭村与那覇岳 2009/7/7 山田香菜子 ishi186 ヤエヤマオオタニワタリ 沖縄県石垣市バンナ岳 2009/7/12 山田香菜子 iri187 ヤエヤマオオタニワタリ 沖縄県竹富町西表島 2009/7/17 山田香菜子 iri188 ヤエヤマオオタニワタリ 沖縄県竹富町西表島 2009/7/17 山田香菜子 oki182 シマオオタニワタリ 沖縄県国頭村辺戸岬 2009/7/8 山田香菜子 oki184 シマオオタニワタリ 沖縄県国頭村辺戸岬 2009/7/8 山田香菜子 oki185 シマオオタニワタリ 沖縄県国頭村与那覇岳 2009/7/8 山田香菜子
34 amami006 シマオオタニワタリ 鹿児島県住用町川内 2010/3/4 中路正嘉 amami007 シマオオタニワタリ 鹿児島県住用町川内 2010/3/4 中路正嘉 oki178 オオタニワタリ 沖縄県国頭村与那覇岳 2009/7/7 山田香菜子 oki179 オオタニワタリ 沖縄県国頭村与那覇岳 2009/7/7 山田香菜子 oki181 オオタニワタリ 沖縄県国頭村与那覇岳 2009/7/7 山田香菜子 ao002 オオタニワタリ 東京都青ヶ島村池の沢 2009/8/19 山田香菜子 ao003 オオタニワタリ 東京都青ヶ島村池の沢 2009/8/19 山田香菜子 ao004 オオタニワタリ 東京都青ヶ島村池の沢 2009/8/19 山田香菜子 ao005 オオタニワタリ 東京都青ヶ島村池の沢 2009/8/19 山田香菜子 ao006 オオタニワタリ 東京都青ヶ島村池の沢 2009/8/19 山田香菜子 yaku009 オオタニワタリ 鹿児島県熊毛郡屋久島町鈴之江 2009/8/24 山田香菜子 yaku010 オオタニワタリ 鹿児島県熊毛郡屋久島町鈴之江 2009/8/24 山田香菜子 hachijo オオタニワタリ 東京都八丈町 2009/5/29 山本薫 hachihoubi ホウビシダ(外群) 東京都八丈町三原山 2009/6/25 村上哲明
35
Fig. 1 rbcL の塩基配列が異なるシマオオタニワタリ類の個体間の交配実験で得 られた雑種の数
縦軸:それぞれの組み合わせで父親と母親を入れ替えて20 回ずつ、合計 40 回 交雑させた際に得られた雑種個体の数。横軸:Kimura’s two-parameter method (K2P) によりrbcL 領域の配列を元に求めた遺伝的距離。*がついたものは、両 親種のどちらかが母親であった場合のみ雑種の胞子体の発生が見られた組み合 わせ。Yatabe et al. (2009)より引用・一部改変。
36
Fig. 2 日本国内におけるシマオオタニワタリ類のサンプル採集地
●:オオタニワタリの採集地、●:シマオオタニワタリの採集地、●:ヤエヤ マオオタニワタリの採集地
37
Fig. 3 Ishikawa ら(2002)の開発したシダ植物のpgiC 領域の PCR 増幅用プラ イマーの位置
38
Fig. 4 Ishikawa ら(2002)が開発したpgiC 領域の PCR 増幅用プライマーと今回 新たに開発したプライマーの位置
配列は上からDryopteris caudipinna (Ishikawa et al., 2002), Asplenium trichomanes (James et al., 2008), A. nidus のもの。
15F, R
Asp15F, R
16R
471R 481R
39 Fig. 5 日本産シマオオタニワタリ類から見いだされた 7 タイプのrbcL の塩基 配列 上から順に沖縄本島産オオタニワタリ(タイプ1)、沖縄本島産オオタニワタリ (タイプ2)、沖縄本島産シマオオタニワタリ(タイプ 3)、北大東島産ヤエヤマ オオタニワタリ(タイプ4)、西表島産ヤエヤマオオタニワタリ(タイプ 5)、南 硫黄島産ヤエヤマオオタニワタリ(タイプ6)、母島産ヤエヤマオオタニワタリ (タイプ7)の塩基配列。色つきの四角で囲んだ位置に塩基の挿入・欠失・置換 が見られる。
40
41