ヒアル ロン酸の細胞機能修飾作用 に関する研究
A Study oFModulattve Egects of Hyaluponan
on Cell Functions
当 別 当 健 司
1995
目 次
要 旨
不
日
文 … …… … … …… …… …… … い
英文 …… ……… ……… ……… ……… ……… ……… ……… ……… ……… ……… ……… ……… ………… 癬 筆 論 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 … … … … 和 一 一 一 一 一 一 一 一 一 一 一 一 … … オく論 ――――………――………―…… 第1章
ヒ ト滑膜細胞 の ア ラキ ドン酸 遊離 に対 す る ヒアル ロ ン酸 の抑 制作 用 序 論 一 エ ネ ー ー ー ー ー ー ー ー ー ー ー ー ー … … 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 … … … … … 材 料 と ダテ法 一 一 … … … 一 一 一 … … 一 … 一 一 一 一 一 … … … 一 一 “ 一 “ “ 一 一 一 神 一 一 一 一 一 一 一 結果 細 胞 と 培 養 液 中 の ド 4c]ァ ラ キ ド ン 酸 代 謝 物 単 一 一 “ 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 … 11ビ
4c]ァラキドン酸遊離に対するブラジキニンとヒアルロン酸の作用
………―――…中
11 ヒアル ロ ン酸 の濃度 及 び分 子 量依 存 的 な ど4c]ァラ キ ドン酸 遊離抑 制作 用 ――…―工……14ド
4qァラキドン酸遊離に対するヒアルロン酸とメチルセルロースの粘度の影響 …――い
16 ド 4c]ァ ラ キ ド ン 酸 取 り 込 み に 対 す る ヒ ア ル ロ ン 酸 の 影 響 一 一 一 一 一 一 一 一 一 一 一 一 一 一 一 -17 [3珂ブラジキニ ン結合特1生 ――――――――――――――――――――――――――――――――い17 [3刺ブラジキエ ンの結合 に対 す るヒアル ロン酸 の影響 一一一一……… 19 細胞生存率及 び密度 に対 す るヒアル ロン酸 の影響 “一一一………^22
考 察 ―――――――――――――――――――――――――――――――――――――――――――――-22
第2章
ラッ ト好 中球 に よ るウシ鼻 中隔軟骨分解 に対 す るヒアル ロ ン酸 の抑制作 用 贋テ高缶―――――――――――――――――――――――――――――――――――――――――――――い26 材料 とラ'法
――――――…………―――――――――――――――――――――――――――…―――‐26 結果 好 中球 による軟骨分解 ――――――――――――――――――――――――――――――■―――‐30
好 中球 に よる軟骨分角7の 特徴 ――――――――――――――――――――――――――――――-30
FWILP/cyt∝h』asln B刺激 した好 中球 による軟骨分解 に対す るヒアル ロン酸 の作 用'-32
0pso zed zymosan刺 激 した好 中球 に よる軟骨分解 に対す るヒアル ロン酸 の作用 ――…34
TPAま
たはA23187で刺激 した好 中球 による軟骨分解 に対 す るヒアル ロン酸 の作 用 い37
ド
刺
BンLPの 結合に対するヒアルロン酸の影響 ………
38
1 3 5 7 8要 旨
細胞外 マ トリックス成分 のひ とつであ るヒアル ロ ン酸は ∬―アセチルーD―グル コサ ミンとD―グル クロン酸の繰 り返 し単位 よ り成 る直鎖多糖であ る。生体 中には結合組織 を中心 と して広 く分布 して い るが,関
節液 には高濃度で存在 してい る。炎症性 関節症 では関節液 中の ヒアル ロン酸 の濃度低 下 や低分子化 が起 こるこ とが報告 されてい る。そ こで関節炎症 に関連す る生体現象 に着 目 し)炎
症 反 応 にお け るヒアル ロ ン酸 の役割,特
に ヒアル ロン酸の濃度変化や分子量変化が細胞 の機能 に及 ぼす 影響 について ,肋 ソJ肋 モデル系 を用いて検討 した。本研究では,ヒ ト滑膜細胞 のアラキ ドン酸遊離, ラ ッ ト好 中球 によるウシ鼻 中隔軟骨分解,及
び ラ ッ ト好 中球のスーパーオキサイ ド(02)産
生 を炎 症 モデル と して用い た 。 1.ヒ ト滑 膜 細 胞 の ア ラキ ドン酸 遊 離 に対 す る ヒアル ロ ン酸 の抑 制作 用 炎症 とプロスタグラ ンジ ンは 密接 に関 連 してお り,アラキ ドン酸はプロス タグラ ンジン生 合 成前駆体であ る。そ こで滑膜細胞 の [HC3アラキ ドン酸標 識 リン脂質か らの ビ4c]ァラキ ドン酸遊 離 に対 す る ヒアル ロ ン酸の影響 を検討 した 。ヒア ル ロン酸 は ブラジキニ ン刺 激 した標 識滑膜 細 胞 か らの[μqァ
ラキ ドン酸遊離 を,反
応後lh以
降抑制 した。この抑制効果は ヒアル ロン酸濃度 及 び分子量 に依 存 していたが ぅ単 に粘度 のみ に起 因す るものではなか つた 。ヒアル コ ン酸 は滑膜 細胞 と[3H]ブラジキニ ンの結合 に影響せず,ま
た滑 膜細胞 によ る[“Clアラ キ ドン酸取 り込みに も影響 しなか つた .ヒ アル ロン酸 に よる ピC]アラ キ ドン遊離抑制は)滑
膜細胞 をカル シ ウムーイ オ ノフォアA23187で
刺激 した場合 に も認 め られ た 。これ らの こ と よ りぅヒアル ロ ン酸 は細胞 膜 におけ るホスホ リパ ーゼ活性 に影響 を与 える可能性が示唆 された。 2.ラ ッ ト好 中球 に よ る ウシ鼻 中 隔軟 骨分 解 に対 す る ヒアル ロ ン酸 の抑 制作 用 炎症性 関節症 の特徴 であ る軟骨分解 は,部
分 的 には浸潤 した好 中球 に起 因す る。そ こで硫酸 化 グ リコサ ミノグ リカ ン遊離 を指標 とした好 中球 による軟 骨分解 に対す るヒアル ロン酸 の影響 を18hま
で 検 討 した 。好 中球 を cytochalasin B存 在下 洋fofmyl―L―methionyl―L―leucyl―L―phenyl―alanine(FMLD,opsO Zed zymosan,12-0-tetradecanoylphorbol-13-acetate(TPA)ま た は カ ル シ ウ
ムーイオノフォア
A23187で
刺激 す る と,軟
骨分解 は顕著 に促進 され た。活性化 され た好 中球 による軟骨分解は,ヒアル ロ ン酸に よ り濃度 及 び分子量依存 的に抑制 された 。Opsonized zymosall
を用いた場合
,低
分子 量 ヒアル ロン酸(猛28× 105)は 軟骨 分解 を抑制せ ず,高
分 子量 ヒアル ロあ る と考 え られ る 。ヒアル ロン酸の 軟骨 分解 抑制 パ タ ー ンは,opso
zed zymosan,TPA及
びA23187を
刺激剤 と して用 いた 場合,相互 に近似 していた。曲 【LP/cytochalasin Bを用 いた場合, ヒアル ロン酸 は軟骨分解 を少な くとも18h抑
制 した。ヒアル ロ ン酸 は好 中球へ の[3H]FMLPの 結合 に影響 せ ず,ま
た予 めF ILPで 刺 激 した好 中球に対 して も軟骨分 解抑制作 用 を示 した 。こ れ らの こ とよ リヒアル ロン酸の抑制作用 には,粘度 に起 因す るものに加 え,粘度以外 の細胞 に対 す る直接 的な作用 もあ るもの と推察 され る. 3.ラ ッ ト好 中球 の ス ー パ ー オ キサ イ ド産 生 に対 す る接 着 表 面 に コ ー トされ た 免 疫 グ ロプ リンGD関
節 液 成 分 及 び ヒアル ロ ン酸 の 影 響 炎症性 関節症 において関節腔 内へ浸潤 した好 中球の産生す る活性酸素 は,関
節組織 に対 し種 々 の傷害 を与 え る。関節軟骨表面 には関節液 中の免疫グ ロブ リン(Ig)沈着が認 め られ,接
着 した 好 中球 の活性化 リガ ン ドとなつてい る.そ こでIgG,関
節液及 び ヒアル ロン酸 に よる好 中球 の接 着表面処理が,接着好 中球 の02産
生 に及ぼす影 響 について検討 した。関節液 中には表面にコー トされ たIgGと 共働 して,好
中球 の02産
生 を促進 す る物質が存在 す ることを見 いだ した。この 02 産 生 促 進 物 質 は 高 分 子 で あ り,酵
素 処 理 に 対 す る感 受 性 よ リム チ ン型 糖 鎖 を持 つ glycocottugatesで あ る と推定 された。IgG及
び 関節液で 表面コー トす る際 に ヒアル ロン酸 を共 存 させ る と,接
着 好 中球 の02産
生が抑 制 された 。表面 を ヒアル ロン酸前処理後,IgG及
び関節 液 で コー トした場合,及
びIgGコ
ー ト表面 をヒアル ロ ン酸で後処 理 した場合 には,高
分子量 ヒ アル ロン酸(猛 2.0× 106)の みが 02 産 生 を抑制 した。いずれの場合で もぅ接着表面 をヒアル ロン 酸(猛 20× 106)で 処理す るこ とによる好 中球の接着抑制は認め られなかった 。これ らの ことよ り,ヒ アル ロン酸 はIgG及
び関節液 中のIsG共
働性02 産生促進物質の表面沈着 を抑制す るか) あ るいは表 面コー トされ た ヒアル ロン酸が直接 的に好 中球の02産
生 を抑制す る可能性が示唆 さ れた 。 以上 の結果 よ りぅ炎症 の進展 に ともな う組織傷 害 に対 して,高
分子量 ヒアル ロ ン酸は組織保護 的 な作用 を持つ もの と考 え られ る。高分子量 ヒアル ロン酸 を生体 由来抗炎症物質 と して位 置づけるこ とが可能であ る。炎症 に ともな う発熱や関節液 中へ遊離 され る炎症性サイ トカイ ンはぅ滑膜細胞 の ヒアル ロン酸合成 を促進す るこ とが知 られてい る。これ らの現象は炎症の修復過程 のひ とつであ り, 生体 の恒常性維持作用の一環 であ ると推察 され る。S剪
“
陶αリ
Hyaluronan(HAl,a component of extracellular matrices,is a linear polysaccharide composed of
alternating u ts of N―acetyl…D-81ucosamne and}81ucuro c acid HA is secrettd from solne
organisms and is found in he highest concentrations in soft connectlve tissues,especially in joint synovial auids ln inflarllmatory arthritls,he synovial HA is reduced in concentratton and lnolecular
welght This smdy was undertaken to elucldate he role of HA in he syno al auid ofinllamedjoints
Effects of HA with various lnolecular weights on some of he cell functlons responsible for
inflaFXllnatory arthritis were exanuned in this study. The attention was focused on the,刀 ッケ施 effects of HA on he release of arachidonic acid froHl synovial fibroblasts, neutrophil― mediated cartilage degradation,and the superoxide release ffom meutrophils
′.れ肪♭,ゎっ 匂他 じ佑 げrfA οЮ肋じrじJglvdじげ α肥じ肪】οれた αじ
"″
ο躍 乃"刑αれりれοガα′デ♭Ю♭ル∫ぬProstaglttdins ha1/e been implicated in many features of infla―atlon A substrate fof
prosぬglandin biosynhesis is frec arachidonic acid(AAJ from phOSpholipids in plasma membranes
Therefore,the effects of HA on the release of AA from phosphohpids, which was induced by the
actlon of bradykiェun to the synovial fibroblasts of osteoarth dc patients,were exanuned.I‐ IA inhibited
he[14c]AA release from he prelれeled synovial cells sttmulated wih or wihout bradykinin l h after
incubation and hereafter The inhibitory effects of HA on he[14c]AA felease were dependent on he
concentration and molecular weight of HA However,he inhibition of the[14c]AA release by HA
was not merely due to he scosity ofI― IA I―IA did not affect he specittc binding ofド H]bradyki n to
the cells The[14c]AA relette induced by calcium― ionophore A23187 was also inhibitt by HA wih
a high molecular weight ln add on,HA did not affect[14c]AA uptake by he cells.These results
suggest hat HA with a high lnolecular weight suppresses he acttvitles of lnettbrane― associated
phospholipases
2.れ肪 ♭テル リ 妍 杉Cぬ げ IA ο泥 れじ′放 η 肪 ′´刑♂J触″′ 臼 的 肋
=9砲 盟 肱所 οη
lnaa_atory joint diseases are characterized by he degradatlon of articular canllages,which is partly mediattd by neutrophils accumulatlng at inflamed sites The effects of HA on neutrophil― mediated degradattom of the cartilage were exaHllned for up to 18 h in a coculmre systern of rat
peritoneal neutrophils with he cartilage of bovine nasal septa The release of sulfated gycosalmmo― glycans from he cartilage、 vas markedly enhanced by the neutrophils、 vhich、vere acttvated、 vith elher
Ⅳ―formyl―L―methionyl―L―leuCyl―L―phenylalanine(FMLP)cO bined wih cytochalasin B,opso zed
zylnosan,12-O tetradecanoylphorbol-13-ace協 俺(TPAl or calCium―
ionophore A23187 However,HA
inhibited he cartilage degradatlon by the actlvated neutrophils The inhibitory effects were dependent
on the con∝ntratton and molecular weight of I― IA When he neu,ophils were incubated wlth
opsonized zymosan,HA with a higher molecular weight(′ yr 2 0X 106)inhiblted he cartilage
degradation for an incubatlon peiod of up to 2 h,but HA wlth a lower∬ Юlecular weight(上 2,8×
1。5)did nOt ehcit significant inhibition on the cartllage degradation for up to 18 h HA did not
actlvation of neutrophils by opsoni‐Led zymosan 「Fhe inhibitory pattem of HA on he cartilage degradatton stimulated witt opsonized zylnosan was essentially siIIlllτ r to hose widi TPA and
A23187,as a stimulating agent of neutrophils,When FTILP/cytochalasin B was used to simulate he
neutrophils,HA samples(haVing different HЮ lecular weights)inhibited he neutrophil― mediated
cartllage degrad ion for notless than 18 h of incubatlon HA did not affect he binding ofド H]Fヽ僻
to the cells, In addition, HA inhibited he cartilage degradatton by he neutrophils preincubated、 vith
FMLP/9朽
chalasin B These results suggest hat the inhibitory eff∝ 偽 of HA on he cartllagedegradation by the neuttophils are not only due to an increased viscoslty Of he lnedium,but also due to a direct rnodulatton of cell functions,
J.聯
じじ倉 げ ∫"機rじ じο″'ng∫ 崩 加 加 刑朋 οgん♭′Jtt C,dノηοッ麓 ′′″ 側ゴrrA οη加♂∫叩 θЙοχ″じ
P/οど′じチわれ妙 2沈じχtt ηじ脱opttJd
ln inaarmatory arthritls,oxygen radical de vattves,、 vhich、vere produced by activated neutrophils in inflamed joint, play a maJor role in he generatlon of joint tissue inJury. IIrllnunoglobuhns in
syno Л nuid are ttapped in he supe ciЛ la_yers of attcular cattlage in inaamedjoint,The trapped
iIImunoglobuhns on he cartilage surface actlvate he neutrophils attached to he surface The effects
of surface treatment witt lgG,syno al auid and HA on he production of supero de a on((乳‐)
were exanuned with he adherent neutrophils lgG and synovial fluids ffonl Pattents with
inaa_atofy joint disease synergisuc』 ly enhanced he鏡‐producion by he adherent rat pe toneal
neutrophils A gel filtratlon sttdy indicated hat he neutrophil― sumulating substance in the synovial auid、vtt a large mol∝ular substance and l1/as inactivated by digestion with boh Proteinases and
O―glycanase,indicating a glycocottugate wih a muciniype sugar chain
When the surfaces were treated witt lgG and syovial fluid in the presence of HA,HA decreased
働‐rdease levels from he adherent neurophils.The surface tteatment wih HA having a high
molecular weight(γ r2 0× 106)p Or tO treating wih lgG and synovial auid resulted in an inhibidon
Of he覺
prOduction by he adherent cells on he surface,The覺 ‐prOductlon in resPonse tO thesurface―coated lgG was also inhibited by subsequent surface― coatlng with HA of妊 2.OX 106 The
nu ber of surface―adherent cells was unchanged after he surface treatlnent wih HA of γ「
20X106
in all cases These results suggestthat(1)HA inhibits he surface― deposidons oflgG and he syno alauid compOnent,resuling in m inhibidon of he働 production and,(2)血 e surface_coated HA with a
high mol∝ular weight directly affects on he adherent neutrophils,resultlng in an inhibidon of he呪 ‐
pЮducion by the cells
Results obtalned in this study conclude血 証 HA wih a high molecular welght eliclts a tissue
protectlve function against he tlssue damage induced by inaa―atlon,HA、vith a large rnolecule rnay
be considered as an endogeneous antl―inaa_atory substance lt has been reported hatI‐IA synhesis
in human synovial cells is stlmulated、 vith infla―atory cytoklnes in synovial fluids and hyperthemia
in the inflaIImatory reactton These phenomena seem to be a natural recovery process ffom
論
緒
生体反応 の根 源は個 々の細胞 の生理機能 にあ る。細胞 自身は生体 中でコ ラーゲ ン)エ
ラステ ン, プロテオグ リカ ン及 び構造糖 タンパ ク質 を主要な もの とす る細胞外マ トリックスに接触,支
持 され てお り)細
胞 と細胞外 マ トリックスの相互作用 に基 づ く生体反応 について幅広 い研究が行 なわれて い る。細胞 と細胞外 マ トリックスの相互作用には,細胞外 マ トリックス生合成 に影響 す るこ とに よ る細胞か ら細胞外マ トリックスヘの作用 と,細胞骨格 と会合 してい る細胞膜上 の レセ プター を介 す る細胞外マ トリックスか ら細胞への作用が考 え られてお り,細
胞外 マ トリックスの挙動は細胞機能 と密接 に関連 す るもの とされてい る[13]. 細胞外 マ トリックス成 分のひ とつ であ るヒアル ロン酸 は/―アセチル ーD―グル コサ ミンとD―グル クロン酸の繰 り返 し単位 よ り成 る直鎖多糖であ る。生体 中には結合組織 を中心 と して広 く分布 して お り,特
に膳帯,関
節液,軟
骨,硝
子体及 び皮膚 には高濃度で存在 してい る 141.高分子量の ヒアル ロン酸は水溶液 中でグル状 とな り,高
い粘性 を与える[5]。 この性質は組織 に流体 力学的な強度 を付 与 した り,水
分保持や浸透圧 の調整な ど生体機能の 維持に重要 とされてい る 辟,6,7]。 また溶液 中で ヒアル ロン酸分子鎖 の形成す る3次
元 ネ ッ トワー クは フィル ター機能 を有 し,細
胞 や 巨大分子 の移 動 を制限す るこ とが示 されて い る14,8,9].従来,本多糖 の機能 は この よ うにその物理 的性 質 と関連 づけ られて きたがぅ細胞 に対する直接 的な生理機能 についてはほ とん ど関心が払 われて こなか った。 しか しなが ら近 年,細
胞表 面上 に ヒア ル ロ ン酸 と結合 す る タ ンパ ク質 (レセ プ ター)が発見 され [10-14],ヒアル ロン酸 の細胞機能 に対 す る研究は急速に進展 して きた。 骨 と骨 を連結 す る部位 が関節 であ る。骨 の末端 は軟骨で被覆 され,関
節全体 は 関節包で包 まれ て い る。軟骨 と軟骨の間には関節液が満 たされてお り,ヒアル ロン酸はその主要な構成成分のひ とつ である。関節包の最 も内側 は滑膜細胞で覆われて お り,関
節液 の ヒアル ロン酸 は この滑膜細胞 で 生 合成 されてい る[15].ヒアル ロ ン酸合成 酵素は細胞膜 に存在 し,細
胞膜 で合成 され た ヒアル ロ ン酸 は細胞外へ と運 ばれ る[16].慢性 関節 リューマチ は滑膜組織 に生 じる炎症 を特徴 とす る関節疾 患で あ り[17],変形性 関 節症 で も二 次 的に滑 膜 炎症が発 生 す る[18]。 この ような関節 では関節液 中の ヒ アル ロン酸は濃度低下や低分子化 を起 こ し,こ れに付随 して関節液 の粘度が低下 す ることが報告 さ れてい る[19]。 また炎症性 関節症 の関節液 中には種 々の炎症 メデ ィエ ーターが検 出され るが,イ ン ター ロイキ ンー1は
そ の 中で も重要な もののひ とつであ る[20,21].イ ンター ロイキ ンー1は
滑膜 細胞 や軟骨細胞 のマ トリックス分解酵素産性 を促進 し関節 の破壊 に深 く関与 す るが pO,21],そ の一方で 滑膜細胞 の ヒアル ロ ン酸合成 を促進 す る こ とが報告 されてい る[22つ旬。また 炎症 は通常 発熱 を伴うが
,滑
膜細胞 の ヒアル ロン酸合成 は温度 の上昇 に よって も促 進 され るこ とが報 告 されてい るp5]. この ような現象は生体 の恒常性維持 に関連 し,炎
症 の修復過程 のひ とつではないか と考 えることが で きる(下図参照).し
か しなが ら関節炎症 とヒアル ロン酸の関わ りについて,細
胞 レベルでの理解 は進んでいない 。そ こで 関節炎症 に随伴 す る生体現 象 に着 目 し,ヒ アル ロ ン酸の役割,特
に ヒアル ロン酸の濃 度変化 や分 子量変化が 細胞の機能 に及 ぼす影 響 について,Jη ソj施 モデル系 を用いて検 討 した 。関節変 と
rrtte∬evttЛ…
TPと‐
り
Prostagiandin E2産 生 炎症 の進 展 発 熱 関節液中への 好中球遊走十
ヒアル ロン酸の分解 ヒアル ロン酸合成 可>(??)
Matrix meta‖o‐
Proteinases産 生
(欝!稿
掲評
)│
軟骨れ壊
―
齢
:▼ide(。2‐ )) 滑膜細胞 軟 骨細胞百田
第
1章
ヒ ト滑膜細胞のアラキ ドン酸遊離 に対するヒアル ロン酸の抑制作用
序 論
疼痛 は変形性膝 関節症 の主要 な症状 のひ とつであ る。ステ ロイ ド性及び非ステ ロイ ド性抗炎症薬 を用 い る変形性膝関節症 の化学療法は,二
次的に生 じた滑膜 炎 に起 因す る疼痛 の緩和 を 目的 と して い る p6,27].炎 症 に よ り組織が傷害 され ると組 織 よ り種 々 の化学物質 が遊離 され る。この中で)ブ ラジキニ ンは極 めて強力な発痛作用 を有す る[28,29].ブ ラジキニ ンは9個
のア ミノ酸 よ り成 るペ プ チ ドで,細
胞表面上 の プラジキニ ンレセ プタ ーに結合 し,ホス ホ リパーゼ ーA2及
び ―Cを
同時 に活 性化 す るpO,31]。 これ らの ホスホ リパーゼは細胞膜 リン脂質 を加水分解 し)直
接 的あ るいは間接 的 にア ラキ ドン酸 を遊離 させ るB2]。 また,カル シウムーイオ ノ フォアA23187は
細胞 内カル シウム濃 度 を直接上昇 させ るこ とに よ リホスホ リパーゼA2を活性化 す るこ とが知 られて い るp3].ブラジキ ニ ンに誘発 され る疼痛 は,末
梢組織痛覚 レセ プターの 感 受性 を上 げるプロスタグラ ンジンE2に よ り増強 され るB41。 シクロオキシゲナーゼ産物の生成 には予めア ラキ ドン酸が遊 離 され ることが 必 要であ るので B5,36],細 胞 膜 リン脂 質か らのアラキ ドン酸遊離 と,炎
症反 応 の主徴候 のひ とつであ る疼痛は密接 に関連 してい るもの と考 え られ る。近年,変
形性膝 関節症 患者の関節腔 内ヘ ヒアル ロ ン酸 を投与す る臨床試験 において,ヒアル ロン酸の鎮痛効果が示されてい るp7,38]。 しか しなが ら その作用機構 は明 らかではない。そ こで ヒ ト滑膜 細胞 を用いて,エ
イコサ ノイ ド生合成前駆体 で あ るアラキ ドン酸の遊離 に対す るヒアル ロン酸の作用 を検討 した。滑膜細胞のアラキ ドン酸遊離 を促 進 させ る手段 と して)細
胞膜 レセ プター を介 して 作用 す るブラジキニ ンと)レセ プターの介在 しな いA23187を
用 いた 。この結果,いずれ の場合 もヒアル ロン酸は濃度及び分子量 に依存 して滑膜細 胞 のアラキ ドン酸遊離 を抑制 す るこ とが示 され た 。しか しなが らヒアル ロン酸 に よるアラキ ドン酸 遊離抑制 は,単
に ヒアル ロン酸の粘度のみに起 因す るものではなかった。さ らに,ヒ アル ロン酸 は 滑膜細胞 に よるアラキ ドン酸取 り込 み には影響 を与 えずぅまた ブラジキニ ンとその レセ プター の結 合 に も影響 しなか った 。これ らの結果 よ り,ヒ
アル ロ ン酸は細胞膜 におけるホスホ リパーゼ活性 発 現 に影響 す る可能性 が示唆 され た 。材 料 と 方 法
ヒアル ロ ン酸 エ ン ドトキシンを含 まない4種
類の分子量の異 なるヒアル ロン酸 鮎 度平均分子量 狂2.0× 106, 14×106,98×105及び2.8x105)は 働η ヵCοCC′∫つ0?'冴ιれた,∫ の培養 上清 よ り精製 されp9],資 生堂医薬品研究所 よ り提供 された。ブタ皮膚 由来 のヒアルロン酸(狂40∼
6.0× 104)及 び ヒ ト勝帯 由来のヒアル ロン酸(猛08∼
12× 106)は 生化学工業 よ り購入 した。 滑膜 細 胞 滑膜 は関節症治療 のための手術時 に変形性 関節症患者 の膝 よ り採取 し,周
辺組織 を除いた後 さ ら に1∼2 mm2の 断片 と した 。滑膜 断片は培養 デ イ ッシュに付 着 させ,約 5分
間放 置後,20%非
働化 ウシ胎児血清 (FBS,Flow iaboratories,U.SA),100U/mlペ ニシ リン(明治製菓)及び100 μg/mlス ト レプ トマイシン(明治製菓)を含 むグルベ ッコ変法 イー グル培地(DMEM;Π
ow laboratories,U.S.A) を添加 し,5%C02,37°
Cで
培養 した 。滑膜 断片 よ リデ ィ ッシュに付着 して周 囲に広 が つた細胞 は, 形態 的に線維芽細胞様 であ った 。この細 胞 は025%ト
リプシ ン(Πow labOratories,US.A.)で 剥離 し た後,6穴
または24穴
培養 プ レー トに1.4∼20× 104 cellS/cm2の細 胞密度 で播種 し,10%劇
BSを含 むDMEM中
で コ ンフルエ ン トにな るまで培養 した。実験 には4∼ 7回
継代 した細胞 を使用 した 。 滑膜 細 胞 か らの ア ラ キ ドン酸 遊 離24穴
プ レー トで 培養 した滑膜細 胞 は)10%劇
BSを含 むDMEM中
[1-HC]アラキ ドン酸(10∼2.2kBq/0.5m1/well,19∼ 2.O GBq/mmole,New EngiaIIld Nuclear,US.A.)で 37°
C,24h標
識 し た 。標 識滑膜 細胞 は コBSを含 ま な い
DMEM l.O mlで
3回
洗 浄後,分
子 量 の異 な る ヒアル ロ ン酸(025,0.5,10及
び15 mg/ml)存
在 下 ま た は非 存 在 下 で っ ブ ラ ジ キニ ン(10″M,Bachem,U.SA)ま
た はA23187(1″M,Sigma Chemic江,U`S.A.)を 添 加 した
10%劇
BSを含 むDM酌
1(05d/we■
)中 で3h培
養 した。培養液 の粘度が及ぼす影響を検討 するため
,他
の培養系 において メナルセル ロース(0.4,15及
び 4.O mg/ml,東 京化成)を添加 した。培養終 了後,培
養液 を採取 し)細
胞はCa21ΔIg2+_不含phosphよe―buffered sdine(PBS)0.5 mlで
2回
洗浄 した。洗浄液は培養液 と混合 し)混
入の可能性 のある剥離細胞 または細胞断片を除去するためぅ室温で 2000×g,15 min遠心 した。この上清を取 り,
放射活性を液体 シンテ レーションカウンター(アロカ LSC-700)で 測定 した。
離に対する影響を次の方法で検討 した。先ず滑膜細胞のウェルを
3群
に分けた。第1群
はヒアル ロ ン酸を添加 した10%劇
BSを含むDMEM(0 5 mVwell)中
で48h前
培養 した。後半24hに
は[“Clア ラキ ドン酸(1 6 kBq)を加え細胞 を標識 した。第2群
はヒアル ロン酸及び[HC]アラキ ドン酸を含む 同一の培養液で24h前
培養 した。第3群
は ヒアル ロン酸無添加 としっ[HClアラキ ドン酸のみ加 えて24h培
養 した。これ らのヒアル ロン酸処理及 び無処理の標識滑膜細胞はDMEM l.O mlで 3回
洗浄 後,ブ
ラジキニン(10/M)を
添加 した10%FBSを
含むDMEM(0.5m1/well)中
で45 min培養 した。 ブラジキニンを添加 しないコン トロールは各群 ごとに設定 した。培養終了後,培
養液 中へ遊離 した 放射活性を測定 した。 滑膜 細 胞 へ の ア ラ キ ドン酸 の 取 り込み24穴
プ レー トでコ ンフルエ ン トとな った滑膜細胞 を,ヒ アル ロン酸(15 mg/ml,猛 2.0×106)存 在下 または非 存在下,10%同
BSを含 むDMEM中
で[Hqァ
ラキ ドン酸 (1.2 kBq/05m1/well)と とも に,時
間 を変 えなが ら5%C02,37°
Cで
培養 した 。培養終 了後,培
養 液 中に残存 す る放射活性 を測 定 した。 細胞及 び培養液 中の脂 質分析 総脂質は酌lchらの方法140]に よ り塩酸酸性下で抽出 した。6穴
プレー トで [HC]ア ラキ ドン酸に よる標識を行 なった滑膜 細胞はセル スクレーパ ーにより剥離 し,少
量の希塩酸 を含むメタノール/ クロロホルム混液(1/2,v/v)に より脂質を抽出 した。培養液は l M HClに よ りpH 3と し,約8倍
量 のメタノール/ク ロロホルム混液(1/2,v/v)イこよ り脂質を抽出 した。得 られた有機溶媒相は窒素気流 中で乾固 させ,ク
ロロホル ムに溶解 した後,薄
層クロマ トグラ フィー(シ リカゲル 60,Merck, Germally)に よ り分析 した。展開溶媒 として,細
胞抽出物 中の リン脂質,アラキ ドン酸,ジアシル グ リセロール及び トリアシル グ リセロール を分 離するた め,石
油エー>ル
/ジエナルエー>ル
/酢酸 (50/50/1,v/1r)を 用いた。培養液抽出物中のアラキ ドン酸の分析にはクロロホルム/メ タノール(93/7, v/v)を用いた。各試料 とも脂質標準品を担体 として展開 した。ヨウ素蒸 気で着色 した脂質スポ ッ ト をかき取 り,シンチ レーションカクテル(ク リアゾール,ナカライ>ス
ク)と混合後)放
射活性 を測 定 した。 滑膜 細胞へ の プラ ジキニ ンの結合(Sigma chemical,U,S,A.),0.01 mM Phosphoramidon(和 光純薬),1 0 mM dithiothfeitol lfrI光 純薬汲
び
0179脂
肪酸不含 ウシ血清アルブミン(コスモバイオ)を添加 したCa21ふlg2+_含有 PBS(pH 7.4)を用いた 酔1-43].6穴 プレー トで培養 した滑膜細胞は1.O mlの この緩衝液で
3回
洗浄後,直
ちにブラジキニ ン結合実験に供 した。滑膜細胞に,ヒ アルロン酸 (1.5 mg/ml,猛 2.Ox 106)及 び非標識 プラ ジキニン(5μ
M,Pe
nsula Laboratoies,USA)存在下 または非存在下,[2,3-prolyl-3,牛 aH(N)ト ブラジキニン(0.7∼35 nM,3774 GBq/mmole,New Englalld Nuclear,U,S,A.)を 含む結合実験緩衝液0.5
mlを
添加 し,37°Cで
1∼180 minイ ンキュベー トした。イ ンキュベー ト後,細胞 を結合実験緩衝液 10 mlで3回
洗浄 し,少
量の2 M NaOHで
可溶化及 び2 M HClに よる中和後ぅ放射活性 を測定 し た。一部の滑膜細胞 につ いては ヒアルロ ン酸前処理 を行なつた。先ず細胞 を,ヒ
アル ロ ン酸(15
mg/ml,″ :20× 106)を 添加 した10%劇
BSを 含 むDMEM(0.5m1/we11)中
で37°C,30∼
180 min培 養 した 。培養終 了後,細
胞 は結合実験緩衝液 で洗浄 し,ブ
ラジキニ ン結合実験 に供 した。 [3H]ブラジキニ ンの特異的結合量は,全
結合量か ら非特異 的結合 量(5〃M非
標 識 ブラジキニ ン存 在下 で測定 した もの)を差 し引いて算 出 した 。 一部の実験では滑膜細胞の ブラ ジキニ ンレセ プターサ ブクラスを明 らかにす るため,[3H]ブ ラ ジキ ニ ン の 結 合 に 対 す る [Lysyl]―,[Thi5ぉ,}Phe71_,[》Afg°,Hyp3,Thf,B,}Phe7]_,及 び [des―Arg9]―ブ ラ
ジキニ ン(いずれ もPe nsula Laborato es,U.S,A.)の 影響 を検討 した.
粘度測定 ヒアル ロ ン酸 (1.5 mg/ml)と メナルセル ロース (0.4,1.5及 び 4.O mg/ml)は
10%劇
BSを
含むDMEMに
溶解 した 。これ らの多糖溶液の粘度(mPA・Sで表示)は)校
正 した回転型粘度計 (東京計 器 た 用い37°Cで
測定 した。分子量 2.O x 106,1.4x106,98× 105及 び2.8x10Sの微生物 由来のヒア ル ロン酸 (1.5 mg/ml)の 粘度はそれぞれ 14.5,8.1,3.8及 び1 8 mPAoSであった。ブタ皮膚 由来及 び ヒ ト膳帯由来の ヒアルロン酸 (15 mg/ml)1よ それぞれ11及
び5.l mPA・ Sとなった。メナルセル ロースの粘度は,0.4,15及
び40 mg/mlで それぞれ 3.1,7.6及 び 12.9 mPAasで ぁった。培養液の 粘度は0,6 mPA・Sであった。 細胞 生 存率及 び密 度 の測定6穴
プレー トで培養 した滑膜細胞の培養液 を,分
子量の異なるヒアル ロン酸(15 mg/ml)を添加 した10%日
BSを含 むDMEM(2 0 mVwell)と
交 換 した 。滑 膜 細 胞 は 5%C02,37°Cで
さ ら に48h培
養 した。培養終了後025%ト
リプシンを加 え,37°C,10 min処理することによ り細胞 を剥離 した。細胞懸濁 液 は
10%FBSを
含 むDMEMと
混合 し,100×g,10 min遠
心 した 。沈澱 した細胞 は10%
配Sを 含むDM酌
1025
に再懸濁 し,血
球計算盤 によ り細胞数 を計測 した。細胞生存率は トリパ ンブルー排除法 によ り評価 した。 統計解析 結果は平均 ±標準偏 差で示 した。同一の ヒアル ロン酸濃度におけるヒアル ロン酸分子量差に対 する統計学的有意差は,対
応のないStude∬sサ ー検定により評価 した。 結 果 細胞 と培 養 液 中 の [HC]ア ラ キ ドン酸 代 謝 物 滑膜細胞 を[HC]アラ キ ドン酸(6.3×104 dPm)で24h標
識 す る とっ4,7× 104 dPmの 標 識アラキ ド ン酸が細胞 に取 り込 まれ た。細 胞 に取 り込 まれた[HClアラキ ドン酸 の うち 87%(4.l x 104dPm)イ よ [HC]リ ン脂質,79%(37×
103 dP→ は [HC]ト リアシル グ リセ ロールであ つた。[HC]ジアシル グ リセ ロール及び[HCJアラキ ドン酸量は無視 し得 るものであ った(いずれ も1000 dPm以
下)。 標 識滑膜細 胞 をブラジキニ ン(10/M)ま
たはA23187(lμ
M)存
在下,あるいは非存在下 (コ ン トロール)で3h
培養 した場合,取
り込 まれた標 識 アラキ ドン酸の遊離は ブラジキ ニ ンで最大 (11.3%)と な りっコ ン トロー ルでは5.5%,A23187で
は9.7%で
あ った 。培 養液 中へ遊離 した全 放射活 性 に対 する遊 離 [“C]アラキ ドン酸量 は,ブ
ラジキニ ン刺激で82.2%,A23187刺
激で772%)刺
激剤無添加 (コン ト ロール)で641%と
な った 。従つて滑膜細胞か らの[“qァ
ラキ ドン酸遊離は,主 としてホスホ リパー ゼ による リン脂質の加水分解 によ り開始 され るもの と考 え られ る。 [MC]ア ラキ ドン酸 遊 離 に対 す るプラジキニ ン とヒアル ロ ン酸 の作 用 標識滑膜 細胞か らの [餡C]ア ラキ ドン酸 遊離 におけ るブラジキニン用量依 存性 を調べ た結果を Fig,1に 示 した。培養時間は3hと
した。標識 アラキ ドン酸遊離は10″Mブ
ラジキニンで最大に達 した。 標識滑膜細胞 を,ヒアルロン酸 (15 mg/ml,猛 2.0×106)存 在下 または非存在 下,10″Mブ
ラジ キニン添加 または無添加で培養 した。滑膜細胞か らの[HClアラキ ドン酸遊離タイムコースは6hま
で調べた(Fig 2)。 ブラジキニン無添加で も滑膜細胞 は 自発的に[HC]アラキ ドン酸を遊離 したが,ブラジキニンを添加すると培養後 15 minで 顕著な[HClアラキ ドン酸遊離が認め られた。ブラジキ ニン刺激 した細胞か らの
[Hqァ
ラキ ドン酸遊離量は無刺激細胞のそれに比較 し,培
養後 30 min及 び 3と で それぞれ38倍
及 び22倍
とな った。一 方,ヒ
アル ロ ン酸を共存させ る とっ細胞 か らの [HC]アラキ ドン酸遊離はブラジキニンの存在にかかわ らず培養後lhで
抑制された。 ヒアル ロン酸(15 mg/ml,猛 2.O x 106)で 前処理 した標識滑膜細胞 をっブ ラジキニン(10″M)添
加 または無添加 で45 min培養 した。この培養時間は,ヒ アル ロン酸が存在 した場合で も[HClアラ キ ドン酸遊離が抑制 されないことより決定 した(Fig 2)。 細胞か らの[HClアラキ ドン酸遊離はヒア ルロン酸前処理によ り抑制され,その効果は前処理時間に依存 した ばable l). ︵ め ︲ O r × Fこ Q つ ︶ 巧 ① の d 〇 一 〇 L ス 〓 > F Q C 〇 一つ 0 ∝0
υ
10 20 30
BradykininψM)
Fig.1.[14cIArachidonic acid release from prelabeled synovial
ce‖s in response to varying concentrations of bradykinin,
Ce‖s were exposed to various concentrations of bradykinin at
37°C for S h Each Pointrepresents the mean tt S D ofthree
6 4 2 ︵ ψ O r X , と α つ ︶ O ① り ヽ 〇 一〇 L ス 〓 > , O S 〇 一0 “ 匡 Incubation dme(h)
Fig.2. Effects of bradykinin and hyaluronan(HA)on the kinetics of〔 14c,1_
arachidonic acid release Prelabeled synovial∝ IIs were incubated with
(● IA1 0r WnhOut(OI△ )bradykinin(10州 )in the presence(▲ ,△ )Or
absence le,o)of1 5 mg/miHA(vr2 0 x 106)at 37° C ARer subiecting
the media to ditterentincubation periodsi radioactivity in the medium
was determined Each pOintrepresents the mean tt S D.ofthree orfour
separate experiments.
Table l. Efrect Of preincubation with hyaluronan(HA)on bradykinin‐ induced
ド
4claraCttdOntt acid rebase from syno創∝‖
sPreincubauOn dme Radioacti ty released(dpm× 10 3)
(h) COntrOl +Bradykinin Relauve tO cOntrol
0 24 48 1.16± 0.01 5.09± 0.32 1.38± 0.07 4.51 ±0.09 1.48± 0.10 4,41 ±0.12 4.39 3.26 2.98
Prelabeled syno al ce‖s preincubated wHh l.5 mg/mI HA(Aガ「2.0× 106)were
exposed to bradykinin(10μM)at 37°C for 45 min.Each value represents the mean tt S.D.ofthree separate experiments.
ヒアル ロ ン酸 の濃 度及 び分子量依 存的 な [MC]ア ラキ ドン酸遊離抑 制作 用 標識滑膜細胞に種々の濃度で分子量の異なるヒアル ロン酸(猛 20×106,1.4×
106,98x105及
び 28× 105)を 添加 し,ブ
ラジキニ ン(10″M)存
在下3h培
養 した。ヒアル ロン酸は滑膜細胞か らの [HC]アラキ ドン酸遊離 を05∼
15 mg/mlの範 囲で濃度依存的に抑制 した (Fig.3).高 分子量の ヒア ル ロン酸は低分子のものに比べ,強
い[Hqァ
ラキ ドン酸遊離抑制作用を示 した。ブラジキニン濃度 を,[HClア ラキ ドン酸遊離促進作用の飽和する10〃M以
上(Fig。 1)と して ヒアル ロン酸の影響を検 討 した。ヒアルロン酸は分子量20× 106と08∼
1.2×106の ものを用いぅ3h培
養 した。ブラジキニ ン10″Mを
添加 した ときに認め られ るヒアルロン酸の[HC]アラキ ドン酸遊離抑制パター ンは,ブ
ラジキニン濃度を20〃M及
び 30″Mへ
上げても変化 しなかつた(Fig 4)。 またこの場合 も,0.25∼ 1,O mg/mlの 範囲で高分子量のヒアル ロン酸は低分子のものに比べ,強
く[HClアラキ ドン酸遊離を 抑制 した。標識滑膜細胞 をA23187(1〃
M)で
3h刺
激 した場合 もヒアルロ ン酸は[14clァ ラキ ドン 酸遊離を濃度依存的に抑制 し,その作用は高分子量のものに強 く認め られた(Fig 5). 5 4 3 ︵ の b r x E α コ ︶ O ① ∽ ∞ ① 石 L 掛 一 ・ぢ≧ 硝 o 巧 側 ∝ 0PC O.5
1.0 1.5 Hyaluronan concentration(mg/mりFig.3. Ettects of varying the molecular weight of hyaluronan(HA)
on bradykinin‐ induced[14claraChidOnic acid release Prelabeled synovial cells were exPosed to bradykinin(10 μM)at 37°C for 3 h
in the absence(PCi positive control)o「 presence of HA of ditterent
mdecular wdghts(回
:28x1051囲 :98x1051囲
:1,4x106j■ :20x106)at0 5,1 01and 1 5 mg/mI.Each column represents the mean ttS D.of three separate experimOnts Statistical significancel
evaluated by the unpaired Studentls r‐ testi are ρく005(・),く001
10μM BK 20μM BK 30μM BK 2 0 8 6 ︵ めb r x E Q つ ︶ 釈 あ “ ⊇ e ス , 一 ・≧も “ ⊇ O s ∝ ψ 口C O.25 0.5 1 0 PC 0 25 05 1 0 PC 0 25 0.5 1 0 Hyaluronan concentration(mg/ml)
Fig.4. Effects of hyaluronan(HA)on[14claraChidOnic acid release in response to
va「ious concentrations of bradykinin(BK) Prelabeled synovial ce‖s were exposed
扉
違縄評
HFζ'解
躙話錦尉」
も
潔
:彦
T記
嘱
''甲
粘躍
I段格
)at 0 25,05,and1 0 mg/ml. Each column representsthe mean tt S D ofthree
早
眠階ξ
十
;E料
幹
')里甘
:‖F召
早
:階EISL∬
ittdbyheunpairedStudeNs
Hyaluronan concentration(mg/mり
Fig.5.Effects of hyaluronan(HA)on A23187-induced t14c〕 _araChidonic acid releasc Prelabeled synovial ceHs were exposed to A23187(刊 μM)
at 37°C for 3 h in the absence(PCi posiuve cOntrOり Or presencc of HA
of diffe「ent molecular weight(日 :08∼
12x106,■
:20x106)at 0 5,10,and 1 5 mg/mI Each column represents the mean tt S D ofthree separate experiments Statistical significancei evaluated by studentis
t‐testi are p<0,05(■)andく 0.01(**)
5 4 3 2 1 0 ︵ ∞ ︰O r X FL α O ︶ O ① ∽ ∞ ① 一Φ L ス 〓 > 甲 Φ ∞ 0 一O ω 匡 0.5 1.0 1.5
[“C]アラ キ ドン酸 遊 離 に対 す る ヒアル ロン酸 とメナ ル セ ル ロー ス の粘 度 の影 響 標 識滑膜 細胞 を
,異
な る分子量の ヒアル ロン酸 (1.5 mg/ml)及 びメチルセル ロース(04,1.5及
び 40 mg/ml)存在下 または非存在下,ブ
ラジキニ ン(10″M)添
加 または無添加 で3h培
養 した 。これ らの多糖溶 液 の粘度 と[Иqァ
ラキ ドン酸遊 離量 の関係 をFig.6に示 した 。8,l mPA・S以
下 の粘度 では,[HClア ラキ ドン酸遊離に対す るヒアル ロン酸 とメチルセル ロースの抑制作用は明 らかに異なっ ていた。特 に ブラジキニ ン存在 下で頭著であつた 。メチルセル ロースは 0,4 mg/ml(3.l mPA・S)及 び15 mg/ml(76 mPA・S)で は粘度 の増加 にもかかわ らず[14clァラキ ドン酸遊離抑制作用 を示 さな か ったのに 対 し,ヒ
アル ロン酸は効果的 に[Hqァ
ラ キ ドン酸遊 離 を抑制 した。しか しなが ら129
mPAas以
上 の粘度では,[Hqア
ラキ ドン酸遊離 量 における ヒアル ロン酸 とメチルセ ル ロースの差 はほ とん ど認め られなか った。 6 4 2 ︵ 甲 O r X FL α O ︶ O O ∽ “ ① 一ω ﹄ ム 〓 > F O ω O 一 0 “ 匡15
12
ViscosКy(mPA・
S)Fig.6. Effects of varying the viscosly of hyaluronan(HA)and methyI
ce‖ulose(MC)Samples On bradykinin‐ induced and spontaneous[14cI_
arachidonic acid releasesI Prelabeled synovial ce‖ s were incubated
wnh(0,O[▲ :pOSuve cOntroり Or without(■ ,□〕△i negauve contrOり
bradykinin(10 μM)at 37°C for 3 h in the absence(▲,△:∞ntrols)or
presence of HA(o,日 )Of different molecular weights at刊 5 mg/ml or
MC(Ol口)ato 4,15,and 4 0 mg/mI Each pointrepresents the mean 土S D ofthree separate experiments(except the positive controli n=9),
Signlficant ditterences of pく 005(4)jく001(・ ホ),andく0001∈ホう
[HC]ア ラキ ドン酸 取 り込み に対 す る ヒアル ロ ン酸 の影響 ヒアルロン酸が培養液 中へ遊離 され た[HC]アラキ ドン酸の滑膜細胞への再取 り込み を促進させ ることにより
,培
養液 中の遊離[“C]アラキ ドン酸 レベルを低下させている可能性が考え られ る。こ のため滑膜細胞の[HC]アラキ ドン酸取 り込みに対するヒアルロン酸の影響を検討 した。細胞は,ブ
ラジキニンに誘導された[HC]アラキ ドン酸遊離に対 して最大の抑制効果を与えた ヒアル ロン酸 (15 mg/ml,猛 20× 106)存 在下 または非存在下,細
胞か らの [HC]アラキ ドン酸遊離 レベルに比べ 過剰量である 0 6 nmoleの [HC]アラキ ドン酸(7× 104 dpm)と ともに培養 した。この結果,ヒ アル ロ ン酸は22hま
で細胞への[MC]アラキ ドン酸取 り込み量に影響 しなかった(Table 2)。Table 2. Effects of hyaluronan(HAl on inCOrparation oflabeled arachidonic acid(AA)in synO al celis
Time(h)
Residual radioadi ty in medium(dpm× 10 4)aRerincubauon cOntrol(wnhOut HAl incubation wК
h HA n
0 6.95 2 5.78±0.09 6 4.29■0.04 22 2.59± 0.01 6.94 5.74±0.03 4.55±0.09 2.62±0.01 2 3 3 3
紹
lttnぎ
詣
│ボ:帰ド
:蹴
獣緊∬
協
I牌
鉄材イ
'l騨!
Atthe end ofincubation,the residual radioacti ty was measured. Each value represents the mean tt S.D.(except O h).
[3H]プ ラ ジ キ エ ン結 合 特 性 滑膜細胞へ の[3H]ブラジキニ ン結合 タイムコースを 180 minま で調べた (Fig 7).ブ ラ ジキニ ン レセ プター に対 して親和 性 の異な るサ ブタイプの存在が示 されてい る[17,20]こ とよ り
,添
加 す る [3H]ブラジキニ ンは低濃度(0.7nM)と
高濃度(35 nM)で検 討 した がぅいずれ も全結合 は反応 後 60minで
平衡 に達 した 。これ よ リブラジ キニ ン結合実 験 は原則 的に 60 minの 反 応 時間で行 な った。 添加 す る[3H]ブラジキニ ン濃度 を変化 させた場合,滑
膜細胞へ の[aH]ブラジキニ ンの特異的な結合 が認 め られ,お
よ そ10 nMで
リガ ン ドの特 異的結合 が飽 和 した (Fig 8).特 異 的結合 デー タを Scatchard分 析 す る と,滑
膜細胞 の ブ ラジキニ ン結合部位 は1種類 と考 え られ)平
衡解 離定数(Kd)イよ
078 nM)全
結合 部位数(Bmax)は 7 6 fmole/1 05 cellSと な つた(Flg.8 1nseり。これは細胞 当 り9×5 4 3 2 1 0 ︵∽ 〓① o めO rヽ 甲 O r X ″L α 巧 ︶ コ E コ O n C 一E 一望ス 0、 ﹂∞ 一工 め︼ 120 150 180 incubation time(min)
Fig.7. ‐Πme course ofi3H〕 bradykinin(BK)binding by synoviai ce‖ s.
Celis were incubated wnh o 7 nM(o)。「35 nM(o)Ofi3HIBK at 37° C. Each pOint represents the mean tt S D.ofthree separate experiments.
「 ● 02468 Bradykh bound(fmde/10Scdlsl
0.5
20
[3HIBradykinin concentration(nM)Fig.8. Binding of i3Hibradykinin(BK)to synOVial ceIIs increasing concentratlons ofi3HIBK were
incubated whh ce‖s at 37°C lor 60 min SpecWic i3HIBK binding is defined as the direrence between
binding in the absence(tOtaりand presence(nonSpecific)of 5 μM uniabeled BK, Each pOint represents
the mean tt s D ofthree separate experiments. Inset represents Scatchard transformation of the specific binding data ︵ ∽ 一一〇 〇 め O r ヽ 甲 O r × 声L α O ︶ O C コ O n C 一 E 一望 ス O ω ﹂ ∞ 一〓 め ︺
2.0
1.5 Φ O ﹂ L ヽ つ E コ 0 ∞30
10
ブラジキニ ン結合 部位 の特異 性 を調べ るため
,ブ
ラジ キニ ンレセ プター…アゴニス ト及び ―ア ンタゴニス トを用いて っ滑膜細 胞への[3H]ブラジキニ ン結合 に対 す る競合効 果 を検討 した(Fig.9)。
滑膜細胞 は濃度 を変 えたアゴニス トまたは ア ンタゴニ ス ト存在下
,3nM[3H]ブ
ラ ジキニ ン(この濃度では非特異 的 結合 を無視 し得 る;Fig 8)と ともに37°C,60 minイ ンキュベー トした。[Lysyl]―ブ
ラジキニ ンは強 く[3H]ブラジキニ ンの結合 を阻害 し,その作用は未標識 ブラジキニ ンと同等であっ
た 。また B2 ブラ ジキ ニ ン レセ プ ター ア ンタ ゴニ ス トで あ る[Thf.8,》Phe7]_ブラジ キニ ン及 び
[》 Arg°,Hyp3,ThiiS,♭Phe7]_ブラジ キニ ンも結合 阻害作 用 を示 した 。しか しなが らBl―ブ ラジキニ
ンレセ プターアゴニス トであ る[des―Arg9]―ブラジキニ ンはほ とん ど競合効果 を示 さなか った。これ
らの結果 よ り
,滑
膜 細胞 の ブラジキニ ンレセ プターはB2ク ラスに属 するもの と考 え られ る141-4司.FIg.9. cOmpetition of binding of i3Hibradykinin(BK)to synOVial ceIIs by BK analogues.
Celis were incubated at 37° C for 60 min whh 3 nM 1 3HIBK and varying concentrations of
uniabeted BK(o),ILysyI卜 BK(o),ID‐ ArgO,H坤3,Thi5,8,D‐Phe7卜BK(▲
),IThi5,8,D‐Phe7卜BK
(A),and ideS‐ Arg91_BK(口 ) Each pOint represents the mean tt S,D.of three separate
experiments [3H]プ ラ ジキニ ンの結合 に対 す る ヒアル ロ ン酸 の影響 滑膜細胞 を,ヒ アル ロン酸(15 mg/ml,猛 20× 106)存 在下 または非存在下)30 nM[3H]ブ ラジキ ニンとともにインキュベー トした。滑膜細胞への[3H]ブラジキニン結合は1∼180 minの 範囲で調 べたがぅいずれの時間ポイン トにおいても,ヒ アルロン酸は特異的[3H]ブラジキニン結合量に影響 4。0 20。 。 ︵ の 一一〇 0 ﹂ O r 卜 L 住 0 ︶ O C D Q O ⊂ 一⊂ 一X ス 0 、 ﹂ m 一 正 ﹂ 10‐8 10‐ 7 10‐6 Peptide concentration(M)
しなかつた(Fig 10)。 また滑膜細胞 をヒアル ロン酸 (1.5 mg/ml,猛 2.O X 106)で30∼180 min前処 理 しても[3H]ブ ラジキニン結合量に影響 を与えなかった(Fig,■).ヒアル ロン酸 (1.5 mg/ml,猛 2.O X 106)ま たは ヒアル ロン酸 と同様に高粘度の溶液 を与え るメチルセル ロース (1.5 mg/ml)で 30
min前
処理 した滑膜細胞 を,さ らに同濃度のこれ らの多糖存在下,8nM[3H]ブ
ラジキニ ンととも に 60 minイ ンキュベ ー トした。この場合 も,い
ずれの多糖 とも[3H]ブラジキニ ン結合量に影響 し なかつた(Table 3)。 0 30 60 90 120 150 180 incubation dme(min)Fig.lo, Ettect of hyaluronan(HA)on the kinetics of t3Hibradikinin(BK)binding
kjゞ
群
`
監釧乱箔低
,著写イ
ピ紹評
I習μ
誂男
T懇
汗
」
l排
ざ
毛
Ftte(・〕
indicated penods speclfic t3HIBK binding(o:0)iS derined as the d‖ ference between binding in the absence(totali□ ,日)and presence(nonSpecilc:△〕▲)of
5 μM uniabeled BK Each pointrepresents the mean tt S.D,of three separate
experiments 3.5 3 . 0 2 . 5 2 . 。 1 . 5 ︲ ・ 0 0 ・ 5 0 ︵ ∽ 〓 O O め O r ゝ Φ ︲ O r X ″ L α O ︶ O E コ o n C 一E 一 望 丞 つ ω よ Ш 甲 ﹂ め ]
3 2 1 ︵の 一一① O り O rヽ ∞ ︲ O r X , と α つ ︶ O C D O n E 一 ⊂ 一Y ス O s ﹂ m 一正 o 中
60 120 180
Preincubation time(min)砧妥
ピ
善
頑
見
1帝
│き0酔
齢銘器鉾想
選螢巻
M
difference between binding in the absence(totaり and presence(ROnSpecific)of 5 μM unlabeled BK Each pointrepresentsthe mean
土S D ofthree separate experiments
Table 3. Compattson of[3Hibradykinin(BК 9 binding in hyaluronan
(HAl‐and methyl cellulose(MC)‐ treated syno al cells
[3HIBK binding(dpm′1 05 ceWS)
Treatment Total Non‐specific Specific
Control
HA
MC
2122± 26 1996± 164 2037±157 220±5 212± 50 270± 112 1902 [1,01a) 1784 [0.941 1768 [0.931胎鉛ポ
1認
端鰍
!謝
縄温
と
好
il:鷺]眺監
駅
lilぎfor 60 min at37° C.SpeciHc[3HIBKbinding isdeHned asthe
difference between binding in the absence(totaり and presence (nOnSpecWic)of 5 μM uniabeled BK.Values are the mean tt S.D. of three separate experiments. a)Each value in square parenthesis
細胞 生存率及 び密 度 に対 す るヒアルロ ン酸 の影 響
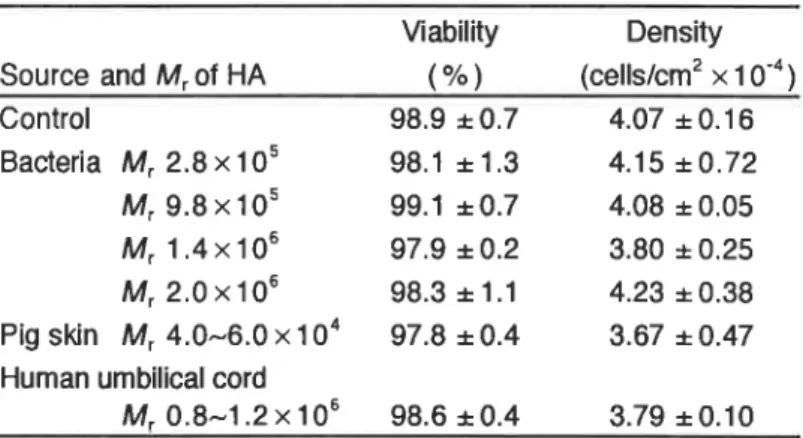
滑膜細胞 を,由来及び分子量の異な るヒアル ロン酸 (1.5 mg/ml)存 在下 または非存在下
,48h培
養 した。いずれの場合 も滑膜細胞の生存率は
98%以
上であった(Table 4)。 細胞密度(単位面積当 りの細胞数)にもヒアル ロン酸の影響はほとんど認め られなかった(Table 4).
Table 4. Effects of hyaturonan(HAl on the abi‖ty and the densКy
Of synO al ce‖s
Viab‖Ry Denshy
Source and Vrof HA (%) (celISん m2×10‐4)
Contro1 98.9± 0.7 4.07±0.16 Bacteria M「 2.8× 105 98.1 ±1.3 4.15± 0,72 ん49.8× 105 99.1± 0.7 4.08±0.05 ん牛 1.4×106 97.9± 0.2 3.80±0.25 ん42.0×106 98.3± 1.1 4.23± 0.38 Pig skin Mr 4.0∼ 6.0×104 97.8± 0.4 3.67±0.47 Human umbWical cord
ん40.8∼1.2×106 98.6±0.4 3.79±0.10
Synovial celis were incubated wК h or wHhout(contrOり 1.5 mgォコ
IHA
of different sources and molecular weights(Mr)at 37°C for 48 h.
Values are the mean tt S.D.oi three or six(cOntrOり Separate experiments, where each was performed in duplicate.
考 察
ヒアル ロン酸は)ブ
ラジキニンまたはA23187で
刺 激 した滑膜細胞ぅ及び刺激 しない細胞か らの [HC]アラキ ドン酸遊離を抑制 した。細胞内アラキ ドン酸 レベルは,ア
ラキ ドン酸の細胞か らの遊離 と細胞膜 リン脂質への取 り込みの平衡によ り調節 され ることが報 告されているB5]。 従 つて,培
養 液 中のアラキ ドン酸 レベルが ヒアル ロン酸によ り低下する現象について,ふ
たつの機構 を想定でき る。先ず ,ヒ アル ロン酸は実際 に滑膜細胞か らのアラキ ドン酸遊離を抑制するとい うもの。次に, ヒアル ロン酸は滑膜細胞のアラキ ドン酸再取 り込みを促進するというものである。これ らの可能性 を検討するため,滑
膜細胞への[HC]アラキ ドン酸取 り込みに対するヒアル ロン酸の影響を調べた。 この結果,ヒアル ロン酸(猛 20× 106)は 15 mg/耐 の濃度で滑膜細胞への[HC]アラキ ドン酸取 り込 みに影響 しないことが示された(Table 2)。 またヒアルロン酸は滑膜細胞に対 して毒性 を示さなかっ た(Table 4)。 従 つて ヒアルロン酸による[HClアラキ ドン酸遊離 レベルの低下は,[LClア ラキ ドン酸遊離抑制 に基 づ くもの と考 え られ る。 ブラジキ ニ ン刺激 した滑膜細 胞の[HC]ア ラキ ドン酸遊離 に対 するヒアル ロン酸の抑制作用は, ヒアル ロン酸の濃 度及び分子量 に依存 していた(Flg,3)。 ヒアル ロン酸は濃度及 び 分子量の増加 に 伴 い,その水 溶液 の粘度が増大 す る [5].[Hqア ラキ ドン酸遊離 は高濃度(40 mg/ml)のメナル セル ロースに よ つて も抑制 され ることよ り(Fig.6),高 分子量の ヒアル ロン酸の抑制効 果はぅ部分的に は粘度 に起 因す るもの と考 え られ る。これは
,培
養液 の粘度が細胞表面 っあ るい は細胞表面 に極 め て近接 した細胞 と培養 液 の境界 領域か らの アラキ ドン酸分子の拡散性 に影 響す る可能 性 を否定 で きないためであ る。しか しなが らメテルセル ロー ス とは異 な り,ヒ アル ロン酸は低 い粘度で も顕 著 な[HC]アラキ ドン酸遊離抑制作用 を示 した 。この結果 よ り,細
胞 か らの[HC]アラキ ドン酸遊離 に対 す るヒアル ロン酸 の抑制効果 は,単
に培養液粘度 の増大に よる結果ではな く,粘
度以外 の要 因が 関 与 してい ることが示唆 され る。 [HC]アラキ ドン酸遊離抑制は,ヒ
アル ロ ン酸が ブラジキニ ン分子 を捕捉 するこ とに よ り,細
胞 表 面上 の ブラジキニ ン レセ プターヘ の ブラジキニ ンの結合 を阻害 す るこ とで も達成 され る。この可能 性 を検討す るため,滑
膜細胞へ の[3H]ブラジキニ ン結合 に対す るヒアル ロン酸の影響 を調べた 。こ の結果)最
も強 く[14qァ ラキ ドン酸 遊離 を抑制す るヒアル ロン酸 (1.5 mg/ml,猛 2.0× 106)を 添加 した場合 で も,1∼180 minの 範 囲で[3H]ブラジキニ ンの結合 は全 く影響 されなか った(Fig 10)。 こ の ときに加 えた[3H]ブラジキニ ン濃度 (30 nM)は,特
異的結合 が既 に飽和 してい る濃度であ ったた め (Flg.8),添加す る[3H]ブラ ジキニ ンを飽和結合 量以下 として検討 した 。しか しなが らこの場合 もヒアル ロン酸 は[3H]ブラジキニ ン結合 に影響 しなか つた。メナルセル ロース(1.5 mg/ )は [HClアラキ ドン酸遊離 を抑制 しなか つたがlFig 6))[3H]ブラジキニ ン結合 にも影響 しなか つた。従 つ て,検
討 した範 囲で の培養液 の粘度 は ブラジキニ ンの細胞への結合 に影響せず,また ヒアル ロ ン酸 分子鎖 中にブラジ キニ ン分子 が捕 捉 され るこ ともない と考 え られ る 。これ らの こ とは,A23187を
用いて)レ
セ プ ター を介 さずに [HClア ラキ ドン酸遊離 を促進 させた場合で も ヒアル ロン酸に よる 遊離抑制が示 され るこ と (Fig 5),ま た 添加 す るブラジキニ ンを[HC]アラキ ドン酸遊離促進作用が 飽和 す る濃度以上 と して も,ヒ アル ロン酸 による遊離抑制パ ター ンに変化 はない こと (Fig.4)で も 支持 され る。 ヒアル ロ ン酸は滑 膜細胞 か らの[HC]アラキ ドン酸 遊離 に対 して,ブ
ラジキ ニ ン存在下及び非存 在下 いずれの場合 も,培
養後 45 minま でほ とん ど影響 しなか った(Flg 2)。 ヒアル ロン酸が抑制効 果を発揮 す るには,滑
膜細胞 とヒアル ロン酸 がlh以
上接触 してい る必要があ るもの と考 え られ る。 滑膜細胞 を ヒアル ロ ン酸 (1.5 mg/ml,猛 2.0×106)で24hま
たは48h前
処理 す る と,ブ
ラジキニン刺激 による滑膜細胞 の[HC]アラキ ドン酸遊離応答 が低下 した(Table l)。 滑膜 細胞 を同 一の ヒア ル ロ ン酸で
48h培
養 して も細胞数 に変化 の なか つた こ と(Table 4)ぅ また滑膜細胞 をヒアル ロン酸 で前処理 して もブラジキニ ンの結合量 に影響 しなか ったこ と(Fig H)を
考慮 す る と)ヒ
アル ロン 酸 に よる[HClアラキ ドン酸遊離 抑制は,ヒ
アル ロン酸が滑膜細胞 に 直接作 用 した結果で あ ると考 え られ る。 細 胞 膜 に存 在 す る酵 素 は膜 流 動 性 な ど細 胞膜 の 物理 的堺 境 に影 響 を受 け る とい わ れ てい る 14・6,47].細胞膜 リン脂 質 を基 質 とす るホスホ リパー ゼ活性 も,リ ン脂質の物理状 態 に起 因す る細 胞 膜微細構造の変化 や細胞膜流動性 の変化 に影響 を受 け るこ とが報告 され てい る148-56].細胞表面上 には ヒアル ロ ン酸結合 タ ンパ ク質が 存在 す るが[10-Ⅷ,こ のタ ンパ ク質は細胞膜 を貫通 し,細
胞骨 格 と会合 してい るこ とが報告 されてい る 躊7-59].細胞表面上の この ような タ ンパ ク質 とヒアル ロン 酸が結合 した結果,膜
流動性 な どの細胞膜 の物理状態 に影響す ることが予想 され る(Scherna I 参照)。 ヒアル ロ ン酸 は巨大分子 であ るため,ひ とつの分子で 複数の ヒアル ロン酸結 合 タ ンパ ク質 と会合 す るもの と考 え られ るが,こ の状態は膜流 動性 の低下 を招 くことが推定 され る。膜流動性 の 低下 はホスホ リパーゼ活性低下 につなが るこ とが示 されてい る 辟9,判。また ヒアル ロン酸 (ヒ ト膳 帯 由来)は,04 mg/mlで
ブタ膵臓 由来 ホスホ リパーゼA2の
無細胞酵素 反応 に影響 しない ことが 報 告 されてい る 膵0].従 つて ヒアル ロ ン酸 に よる細胞 膜流動性 の低下 を仮定 すれば,ヒ
アル ロン酸 に よるアラキ ドン酸 遊離抑制作用 を説 明で きるもの と考 え られ る。Underhillと Toole[10]は)分
子量 45×105,95× 105及び13x106の
ヒアル ロン酸 を用い,線
維芽 細胞 への ヒアル ロン酸 の結合親和 性は分子量の増加 に依 存 して大 き くなるこ とを報告 してい る。また この現象 は,分子量の増加 に伴 っ て,ひとつの ヒアル ロン酸分子の 占有す る結合 タンパ ク質が多 くな ることに起 因す るものだ として い る。この報告 と,ヒアル ロン酸が滑膜細胞 か らのアラキ ドン酸遊離 を分子量依 存的に抑制 する結 果 とは よ く一致 す る。 近年 ,イ ンター ロイキ ンー1に
よる滑膜細胞や軟骨細胞 のプロス タグランジンE2産
生 を,ヒ アル ロン酸が 加ソj″οで抑制するこ とが報告された [61,62].プロスタグランジンE2はシクロオキシゲナー ゼ産物のひ とつであ るがっシクロオキシゲナーゼは小胞体 に存在 し,細
胞膜 には存在 しない “ 3,641. 外 部 よ り添加 した ヒアル ロン酸 は細胞膜 を通過 しない こ と[65]よ り,ヒ アル ロン酸 によるプロスタ グラ ンジ ンE2産
生抑制はシクロオキシゲナーゼに対す る直接作用 とは考 えに くい。イ ンター ロイ キ ンー1に
よって滑 膜細胞[6671]や軟骨細胞 『 2,73]の プロスタグランジンE2産
生が促進 され る過 程 にはホスホ リパーゼの活 性化 が伴 うこ とが数多 く報告 されてい る。また プロスタグラ ンジンE2 な どのエイコサ ノイ ド生合成 は,リ ン脂 質か らの遊離アラキ ドン酸生成 に よって律速 され るB6,49].これ らの ことを考慮 す ると,ヒ アル ロン酸が細胞膜 に結合 することによ リホスホ リパーゼの活性化 が抑制 され
,そ
の結果 として プロス タグランジンE2産
生が抑 制 された ことが推定 され る.ヒ アル ロン酸結合 タ ンパ ク質のひ とつであ るCD紹
分子 に対す るモノクローナル抗体 で滑膜細胞 を前処理 す る と,サ
イ トカイ ンに よ り刺激 した滑膜細胞 の プロスタグラ ンジンE2産
生 が 抑制 され るこ とが 報告 された 『 4],こ の報告は,細
胞膜 に結合 した ヒアル ロン酸 の ホスホ リパーゼ活 性 へ の影響 に関 す る仮説 を支持す るもの と考 え られ る。 本研究では,ヒ アル ロン酸の関節組織 における作 用を評価 するモデル として,ヒ ト培養滑膜細 胞 を用いた 。このモデル よ り得 られた結果は,関
節症 患者 関節液 中の プロスタグ ラ ンジ ンE2量
に 対 して,関
節 内投与 した ヒアル ロ ン酸 は抑制的であ る とい う近年 の知 見 『 5]と矛盾 しない 。関節 内に おいて高分子量 の ヒアル ロン酸が高濃度で存在 す ることは,滑
膜細胞 か らのア ラキ ドン酸遊離抑制 作用 に基 づ プロス タグ ラ ンジ ンE2産
生抑制効果 を もた らし,関
節 炎症 の鎮静化 に寄与 す るもの と 考 え られ る。Schema I. interaction of HA with the ce‖ surface
第
2章
ラ ッ ト好 中球 に よ る ウシ鼻 中隔 軟 骨 分 解 に対 す る ヒアル ロ ン酸 の抑 制 作 用序 論
コラーゲ ン及び プロテオグ リカ ンを主要成分 とす る関節軟骨は骨端 を被覆 し,荷
重 よ り骨 を保護 す る結合組織であ る。軟骨 の物理的強度 は,プ
ロテオグ リカ ンやコラーゲ ンの分解 によ り著 しく低 下す る 『 6,77].慢 性 関節 リューマチ を始め とす る炎症性 関節疾患は 関節軟骨 の分解が特 徴であ る が,部分 的に は炎症局所 に集 ま つた好 中球 に起 因 す る 『 8お0].好 中球に よる軟骨分解 には,主
と し て ライソゾー ム酵素,特
にセ リンプロテ アーゼであ るエ ラスターゼやカテ プシ ンGが
関与 す る とい われてい るが[8184]ぅ その機構 は よ く理解 されてい るわけではない 。刺激 され た好 中球は さ らに活 性酸素 を産生す るが,関
節液 の主成分のひ とつで あ るヒアル ロ ン酸 は活性酸素 に よ り分解 され,関
節液 の粘度低下 を招 くこ とが報告 されてい る ド5-87]。 そ こで好 中球 による軟骨分解 に対 して ヒアル ロン酸の分子量変化 がいかな る影響 を与 えるのか検討 した。本研究ではラ ッ ト腹腔好 中球 とウシ鼻 中隔軟骨 の共培養 系 をモデル として用いた 。また軟骨 の分解 に伴 い軟骨 よ り著 し くプロテオグ リカ ンが遊離 され るが ,コ ラーゲ ン遊離はほ とん ど見 られない ことよ り 『 6],軟 骨 よ り遊離す るプロ>
オグ リカ ン量 を硫酸化 グ リコサ ミノグ リカ ン量 と して 測定 し,軟
骨分解 の指標 と した 。好 中球の刺激 剤 と して, cytochalasin B存 在 下 洋formyl L― methionyl―L―leucyl―L―phenylalalline(FWIL⊃ ,
opsonized zymosan,12-0-tetradecanoylphofbol-13-acetate(TPA)及 び カ ル シウ ム ーイ オ ノ フ ォ ア



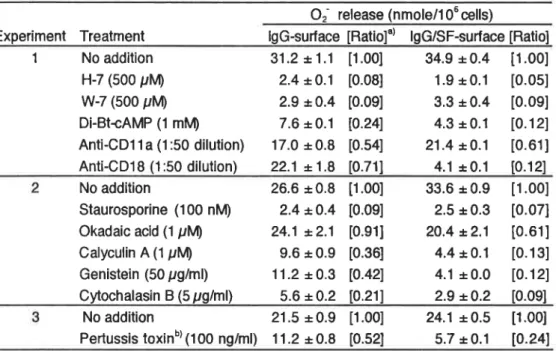

![Table ll.Effects of enzyme treatments of syno alttuid(SD On Oxidative response of neutroph‖ s Oす release(nmole/106 ce‖ S) Expettment Enzyme +pooled fracuon[Ratio]° ‐pooled frac∬ on lb) No addition 22.8± 0.5 11.431 15.9± 0.9 2C' No a](https://thumb-ap.123doks.com/thumbv2/123deta/5785655.1028191/59.892.167.690.139.402/effects-treatments-alttuid-oxidative-response-neutroph-expettment-addition.webp)

