DOI: http://doi.org/10.14947/psychono.36.1
不快感に関する視覚の時間特性
吉 本 早 苗
中京大学
Temporal characteristics of visual discomfort
Sanae Yoshimoto
Chukyo University
Time-varying patterns such as flickering lights can cause discomfort and induce seizures in photosensitive ob-servers. An understanding of the temporal characteristics of visual discomfort is therefore important from both sci-entific and practical viewpoints. The purpose of this paper is to review existing studies on the impact of temporal characteristics on discomfort. Two related factors have been suggested as predictors of discomfort caused by time-varying patterns: (1) excessive contrast energy at the medium temporal frequencies to which the visual system is generally most sensitive, and (2) temporal deviations from the natural (1/f) statistical characteristic. These effects mirror the visual discomfort caused by spatial patterns, in some ways but not all. The interaction between spatial and temporal parameters needs to be investigated to clarify the factors underlying visual discomfort.
Keywords: visual discomfort, flicker, natural image statistics, temporal frequency
1. は じ め に 光源の明滅によるフリッカは時に不快感を喚起し,人 によっては頭痛や目眩,さらには痙攣や意識障害を伴う 光感受性発作を誘発することがある。1997年12月16日 に放映されたアニメ番組「ポケットモンスター」の視聴 者が不快感を訴え,光感受性発作と思わしき症状を発症 し,全国各地で約700名もの患者が病院に搬送された事 件は海外でも驚きをもって報道された(Takahashi & Tsu-kahara, 1998)。問題となったシーンでは,青色光と赤色 光の高速切り替えにより輝度が激しく変化する効果が使 用されていた。このいわゆる「ポケモン騒動」はその規 模の大きさからあまりに有名であるが,テレビ番組が光 感受性発作を誘発した事例は,実はそれ以前から存在す る。1980年代には,米国で放映されたテレビ番組「キャ プテン・パワー」を視聴していた男性が光感受性発作を 起こした。1993年に英国で放映された「ポットヌード ル」のテレビコマーシャルにより,視聴者3名が光感受 性発作を起こしたという事例もある。いずれも急激な輝 度変化や激しいフリッカを伴うシーンが映像に含まれて おり,それが原因で発作が生じたと考えられている (Fisher, Harding, Erba, Barkley, & Wilkins, 2005; Harding &
Harding, 1999)。フリッカが不快感や光感受性発作を喚 起する事例は,このような映像作品に留まらず,日常で 使用する室内照明においても見られる。一般的に使用す る蛍光灯の時間周波数は100∼120 Hzであり,明滅を感 じる最小の時間周波数(臨界融合頻度; critical fusion frequency, CFF)を上回るため,連続した一様の明るさ として知覚されるはずである。しかしながら,そのよう な高時間周波数でフリッカする光源が不快感や頭痛を誘 発することが報告されている(Poplawski & Miller, 2013; Wilkins, Nimmo-Smith, Slater, & Bedocs, 1989)。
フリッカなど光感受性発作を起こしやすい視覚パター ンの明るさ,コントラスト,色,大きさ,時間周波数な ど様々な視覚特性について,古くから調べられてきた (e.g., Harding & Harding, 1999; Jeavons & Harding, 1975;
Wilkins, 1995)。これまでの研究では,光感受性発作の 患者を対象に心理実験や脳計測に主眼が置かれていた。 一方で健常者を対象とし,映像の不快感に関して生体の 感覚・知覚のメカニズム解明を目指す知覚心理学的観点 Copyright 2017. The Japanese Psychonomic Society. All rights reserved. Corresponding address: School of Psychology, Chukyo
Uni-versity, 101–2 Yagoto Honmachi, Showa-ku, Nagoya-shi, Aichi 466–8666, Japan. E-mail: [email protected]. ac.jp
からアプローチした研究は少なく未だ十分とは言えない (So & Ujike, 2010)。光感受性発作を起こしうる映像は 往々にして健常者にとっても不快であることから,時間 的に変化するパターンがもたらす不快感の原因を究明す ることは重要な課題である。本稿では視覚の周波数特性 に焦点を当て,視覚的不快感(visual discomfort)の時間 特性に関する研究成果を概観し,今後の課題について議 論することを目的とする。 2. 視覚的不快感と周波数特性 視覚的不快感と周波数特性の関係については時間・空 間の両次元で検討されており,特に空間次元において知 見が蓄積されている(Cole & Wilkins, 2013; Fernandez & Wilkins, 2008; Juricevic, Land, Wilkins, & Webster, 2010; O’Hare & Hibbard, 2011, 2013; Wilkins et al., 1984)。 そ こ で,まずは視覚的不快感の空間周波数特性に関して得ら れている知見を紹介し,それとの比較において視覚的不 快感の時間周波数特性について議論する。 2.1. 空間次元 視覚の空間特性を表す方法の一つとして,正弦波状に 輝度が変化する格子パターンを刺激とし,検出閾の逆数 をパターンの空間周波数の関数として表したコントラス ト感度関数がある。これは視覚の空間周波数特性を表す ことになる。なお,視覚刺激のコントラストを時間的に 変調させて閾値を求めれば,視覚の時間周波数特性を定 義することができる。明所視下で静止した正弦波パター ンを用いた場合,コントラスト感度関数は約3 cycles per degree (c/°)に感度のピークを持つ帯域通過型(バンド パス型)の形状を示す(Campbell & Robson, 1968)。ピー ク感度に近い空間周波数を示す格子パターンは,不快感 や頭痛,光感受性発作を喚起することが知られている (Chatrian, Lettich, Miller, & Green, 1970; Marcus & Soso,
1989; Wilkins, 1995; Wilkins et al., 1984)。
Fernandez & Wilkins (2008)はランダムドットパター ンを用い,フーリエ変換して二乗余弦フィルタにより 3 c/°をピークに±3オクターブの範囲で振幅を高めた画 像とそうでない画像の不快感を測定した。その結果, 3 c/°をピークに振幅を高めた画像は,そうでない画像よ りも不快と判断される傾向にあることがわかった。同様 にランダムドットパターンを用いた研究から,3 c/°の 3 オクターブの範囲にある中域の空間周波数(特に 0.375∼1.5 c/°)の振幅を高めると不快感が上昇すること が報告されている(O’Hare & Hibbard, 2011)。彼らは, 感度の高い中域の空間周波数帯域を過度に刺激するよう
な視覚情報に対して強い神経応答が生じ,それが不快感 を生成する可能性があると指摘している。これは,高コ
ントラストパターンが不快感を喚起しやすく(Haigh,
Cooper, & Wilkins, 2015),それによって生じた偏頭痛と 第一次視覚野(V1)における神経活動量に関連性が見
られたことからも支持される(Huang, Cooper, Satana,
Kaufman, & Cao, 2003)。
蓮の花托など多数の穴や物体が密集した画像は多くの 人が気持ち悪いと感じ,集合体恐怖症(トライポフォビ ア)という用語を耳にするようになって久しいが,集合 体をはじめヘビやクモの画像から一部の現代アートま で,不快感を覚える画像は往々にしてピーク感度を示す 空間周波数(約3 c/°)の振幅が相対的に高いことが報告 さ れ て い る(Cole & Wilkins, 2013; Fernandez & Wilkins, 2008)。 2.2. 時間次元 時間周波数特性についても空間周波数特性と同様のこ とが言える。コントラストを時間的に変調した正弦波パ ターンの検出閾を測定してコントラスト感度関数を求め た場合においても,コントラスト感度関数は帯域通過型 を示す。ピーク感度を示す時間周波数は視覚刺激の大き さにより異なり,直径2°の円形刺激を用いるとピーク感 度を示す時間周波数は約 8 Hzとなるが,円形刺激の直 径を 17°まで大きくするとピーク感度は約15 Hzにシフ トする(de Lange Dzn, 1958; Kelly, 1961)。
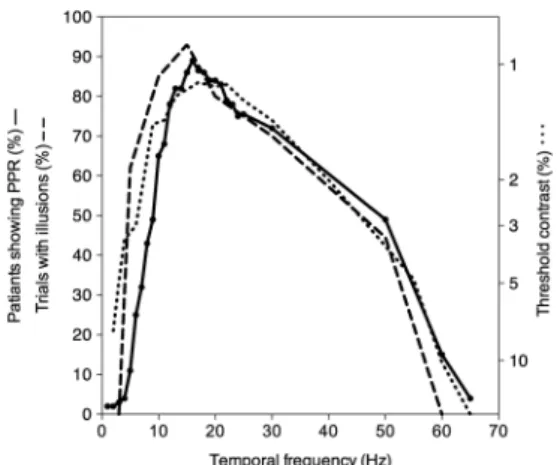
Lin, Hsieh, Chang, & Chen (2014)は18°×15°のLEDディ スプレイを用い,フリッカの不快感を測定した。その結 果,視覚刺激が大きいときのピーク感度となる約15 Hz で不快感が高まることがわかった。また,「ベンハムの 独楽」(Benham, 1894)は運動により白黒の無彩色パター ンから色覚が生じる錯視として知られているが,色覚が 最も生じやすい時間周波数も約15 Hzである。この錯視 は不快感や頭痛を喚起しうる現象であり,光感受性発作 が生じる可能性すらある(Wilkins et al., 1984)。このよ うに,運動する視覚刺激に対して健常者が不快感を感じ やすい時間周波数はコントラスト感度関数のピーク付近 にあり,脳波測定により光突発反応(photoparoxysmal response, PPR)が見られるフリッカの時間周波数とも合 致する(Harding & Harding, 1999; Harding & Jeavons, 1994)。 PPR とは光刺激によって生じる突発性異常脳波反応 であり,光感受性発作の診断に用いられる。コントラ スト感度関数により推定される時間周波数特性,運動す る無彩色パターンから色覚が生じる頻度,及び PPRが 認められた患者の割合を時間周波数毎にまとめたものを
Figure 1に示す。Figure 1から見て取れるように,いずれ もピークを示す時間周波数は約15 Hzである。以上の実 験的事実から,時間的に変化するパターンに関しても静 止したパターンと同様,感度の高い中域の時間周波数帯 域を強く刺激することが過剰な神経応答を誘発し,結果 として不快感に結びつくと考えられる。 3. 視覚的不快感と自然情景統計量 3.1. 空間次元 視覚的不快感と周波数特性に関連して,自然情景統計 量が不快感生成に関与するという説を紹介する。自然情 景をフーリエ変換すると,振幅の強度は空間周波数の逆 数に比例する傾向にあることが知られている(Burton & Moorhead, 1987; Field, 1987; Tolhurst, Tadmour, & Chao, 1992)。これは空間周波数をfとして「1/f」と表される。 つまり,フーリエ変換された画像の空間周波数スペクト ルの振幅成分を対数スケールでプロットすると,傾きは −1 となる。Juricevic et al. (2010)はランダムドットパ ターンやモンドリアンパターンを用い,その空間周波数 成分の振幅を表す傾きが 1/fを示す−1よりも急な場合 (<−1)と緩やかな場合(>−1)とで不快感が異なるか 検討した。振幅の傾きが−1よりも急な場合には,低空 間周波数成分が相対的に増加し,空間的ブラー(ぼけ) が生じる。一方で振幅の傾きが−1よりも緩やかな場合 には,高空間周波数成分が相対的に増加し,エッジが鮮 明になる。実験の結果,視覚パターンを構成する空間周 波数成分の振幅の傾きが−1 (1/f)の時に不快感が最も低 く,傾きが−1から離れるにつれて不快感が上昇するこ とが明らかになった。画像はぼけていても,鮮明すぎて も不快感が高まると言える。 振幅スペクトルが1/fを示さない画像に対して不快感 を感じることは,視覚系の反応特性が自然情景に対して 最 適 化 さ れ て い る と い う 仮 説(Atick, 1990; Atick & Redlich, 1992; Barlow, 1981; Field, 1987)に基づけば,うま く説明できる。ここで言う「最適化」とは,視覚情報を 少数の神経細胞により効率的に表現するスパースコー ディング仮説に基づく。時空間的に絶えず変化する莫大 な感覚情報から,なるべく少ない(スパースな)神経細 胞の発火により本質的な情報のみを表現することができ れば,複雑な情報を早く的確に上位の階層に伝えること が可能となるため,階層構造を持つ視覚系にとって効率 がよい。モデル研究及び神経生理学的研究から,自然情 景に関する視覚情報は少ない神経活動量で処理されるこ と が報 告 さ れ て い る(Lennie, 2003; Olshausen & Field, 2004)。
自然情景の振幅スペクトルは1/fを示す傾向にあるこ とから,そのような空間的特徴を持たない画像を処理す るためにはより多くの神経細胞が活動しなければなら ず,それが不快感に結びつくと考えられる(Fernandez & Wilkins, 2008; Juricevic et al., 2010)。この説明は,Hib-bard & O’Hare (2015)によるV1神経細胞の反応特性に関 するモデル研究からも支持される。彼らは様々な静止パ ターンを用い,不快度の高いパターンの振幅スペクトル は1/fを示さず,かつV1の神経活動量を増加させること を報告した。その一方で,近年のfMRI研究から,視覚 パターンを構成する各空間周波数成分の振幅の傾きを操 作すると,傾きが−1 (1/f)を示す時にV1を含む比較的 初期の視覚野のBOLD反応が最大になること,すなわち 神経応答量が増加することが示されている(Isherwood, Schira, & Spehar, 2017)。この点については「5.3. 神経基 盤」で別途論じる。
3.2. 時間次元
空間次元における不快感と自然情景統計量の関係は, 時間次元においても見られるであろうか?上述したよう に,不快感と時間周波数に関する研究はこれまでにも行 われているが(Harding & Harding, 1999; Harding & Jeav-ons, 1994; Lin et al., 2014; Wilkins, 1995),単一の時間周波 数との関連性に留まることが多く,自然情景統計量との 関連性は検討されていなかった。本節では,時間的に変 Figure 1. The effect of temporal frequency on the
per-centage of photosensitive patients showing a PPR plot-ted in solid line (Harding & Jeavons, 1994), the thresh-old contrast plotted in dotted line (Kelly, 1961), and the percentage of trials on which normal observers re-ported illusions of color plotted in dashed line (Wilkins, 1995). Figures are redrawn using data from the publications.
化するパターンを用いてこの問題に取り組んだYoshimo-to et al. (2016)の研究を中心に,時間次元での視覚的不 快感と自然情景統計量の関係について議論する。 自然情景の振幅スペクトルが1/fを示す傾向にあるの は,空間次元のみに限られているわけではない。自然情 景の時間成分をフーリエ変換すると,振幅の強度はやは り時 間 周 波 数 の 逆 数 に 比 例 し(Dong & Atick, 1995a; Snow, Coen-Cagli, & Schwartz, 2016; van Hateren & van de Schaaf, 1996),そのような特徴を持つ時間情報は少ない 神経活動量で効率的に処理される可能性がある(Dong & Atick, 1995b; van Hateren, 1993; van Hateren & Ruderman, 1998)。したがって,時間的に変化するパターンから喚 起される不快感に自然情景統計量が関与していても不思 議ではない。しかしながら,時間次元における自然情景 統計量は1/fを示さない眼球・頭部運動による影響を受 ける(Wilkins, 2016)。また,視覚情報処理機構は時空間 で異なる。例えば,時間・空間周波数特性はともに視覚 系に複数存在する時間・空間周波数チャンネルにより決 定されると考えられているが,空間周波数チャンネルが 6程度であるのに対し,時間周波数チャンネルは2∼3と 少ない(Snowden & Hess, 1992; Watson & Robson, 1981)。 そのため,時間次元においても空間次元と同様に自然情 景統計量によって不快感を説明することができるかは明 らかでない。
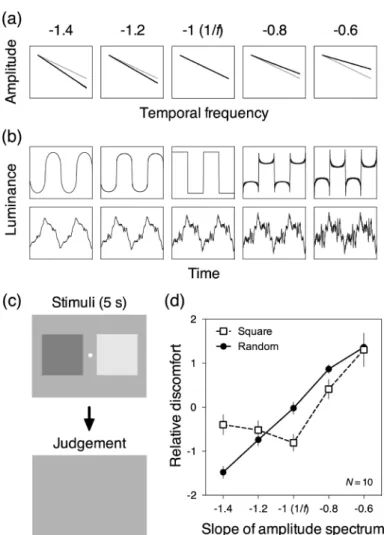
Figure 2. Schematic examples of (a) the amplitude spectra and (b) the waveform of the square wave (top) or random (bot-tom). (a) The amplitude is plotted as a function of the temporal frequency in log-log axis. The gray curves are plots of am-plitude at the slope of –1 (1/f). (b) The luminance of the waveform is plotted as a function of the time. (c) Schematic de-scriptions of the stimuli in a single trial. (d) The relative discomfort score is plotted as a function of the slope of the amplitude spectrum. The two curve represent ratings for the different phases (square wave or random). Error bars represent 95% confidence interval (CI). Figures are redrawn using data from Yoshimoto et al. (2016).
Yoshimoto et al. (2016)はこの問題について,フリッ カを用いて検討した。フーリエ合成により生成した矩形 波とランダム波に基づいて輝度を変調させ,各時間周波 数成分の振幅を表す傾きを−1.4, −1.2, −1 (1/f),−0.8, − 0.6で操作した。空間次元と同様に,振幅の傾きが−1よ りも急な場合(<−1)には,低時間周波数成分が相対 的に増加し,時間的なブラーが生じる。つまり,時間的 な輝度変化が滑らかになる。一方で振幅の傾きが−1よ りも緩やかな場合(>−1)には,高時間周波数成分が 相対的に増加し,時間的な輝度変化が鮮明になる。基本 周波数は 1 Hzとし,各時間周波数成分(高調波)の位 相は等しくするか(矩形波),あるいはランダムに変化 するものとした(ランダム波)。生成したフリッカの振 幅スペクトルを Figure 2(a),輝度の波形を Figure 2(b) に示す。Figure 2(b)上段は矩形波で,下段はランダム 波の例である。一対比較法に基づいて振幅を操作したフ リッカを対提示し,実験参加者(N=10)は左右どちら のフリッカがより不快に感じるかを回答した。矩形波と ランダム波は別々の条件として実施した。刺激の模式図 と一試行の流れをFigure 2(c)に示す。刺激の大きさは 17°×17°であり,コントラストは平均50%(SD=10%) とした。刺激は5秒間提示された。 各フリッカの相対的な不快感を推定した結果をFigure 2(d)に示す。矩形波に基づくフリッカでは,振幅スペ クトルが1/fを示さない時に不快感が上昇した。これは, 空間次元と同様に,自然情景統計量(1/f)からのずれ がより多くの神経細胞を発火させ,結果として不快感を 喚起するという仮説により説明される。一方で,ランダ ム波に基づくフリッカではそのような傾向は見られず, 傾きが−1.4のときに不快感が最も低下し,傾きが緩や かになるにつれて不快感が上昇することがわかった。以 上の結果は,時間的なパターンによる不快感については 自然情景統計量に基づいて説明される場合とそうでない 場合があることを示唆する。また,振幅スペクトルが不 快感に及ぼす影響は各時間周波数成分の位相が等しい時 (矩形波)とランダムに変動する時(ランダム波)で異 なったことから,時間次元における視覚的不快感には振 幅だけでなく位相が関与する可能性がある。 4. 視覚的不快感と位相 4.1. 空間次元 静止パターンがもたらす不快感に関しては,自然情景 統計量(1/f)との相違性など振幅スペクトルが主要な 原因として考えられており,位相は不快感にはさして重 要でないとされている。Fernandez & Wilkins (2008)は,
不快度の高いパターンと低いパターンで振幅スペクトルは 変えずに位相スペクトルのみ交換し,不快度が変わるか 検討した。その結果,振幅スペクトルが元のパターンと同 じであれば,位相スペクトルが異なっていても不快度は変 わらないことを発見した。このことから,彼らは空間次元 において位相は不快感に影響しないと結論づけた。 4.2. 時間次元 時間次元においては空間次元とは異なり,位相が視覚 的不快感に影響する可能性が見出されたが,この問題は これまで取り組まれていなかった。そこでYoshimoto et al. (2016)は振幅スペクトルを 1/f としたフリッカを用 い,各時間周波数成分の位相をある角度(0°, 30°, 60°, 90°) で等しくするか,ランダムに変動させるかによってフ リッカの不快感が異なるか検討した。波形を Figure 3 (a)に示す。位相が0°の時には矩形波となる。Figure 2 (c)に示した方法と同様,一対比較法により実験参加者 (N=14)の位相を操作したフリッカに対する不快感を 推定した。刺激の大きさは17°×17°であり,コントラス トは平均50%(SD=10%)とした。刺激は5秒間提示さ れた。
Figure 3. (a) Schematic examples of the waveform for phases of 0°, 30°, 60°, 90° or for random phases, from left to right. The luminance is plotted as a function of the time. (b) The relative discomfort score is plotted as a function of the phase spectrum. Error bars represent 95% CI. Figures are redrawn using data from Yoshi-moto et al. (2016).
実験の結果,位相がランダムに変化する時に不快感が 著しく上昇することがわかった(Figure 3(b))。これは, 時間次元における不快感には振幅だけでなく位相も関与 することを示している。Fernandez & Wilkins (2008)に よると,空間次元では振幅スペクトルが不快感の程度を 左右し,位相は不快感に影響しない。彼らが用いた刺激 は位相がランダムに変化するパターンではなく現代アー トであるため,この実験結果との直接的な比較はできな い。しかし,位相が不快感に与える影響は時間次元と空 間次元で大きく異なる可能性がある。 時間次元で位相が不快感に影響しうる原因として,関 与する視覚情報処理機構が異なる可能性がある。各時間 周波数成分の位相が等しい場合には,輝度変化は時間的 な エ ッ ジ を反 映 し う る(Bilson, Mizokami, & Webster, 2005)。例えば,矩形波に基づく輝度変化は視覚的遮蔽 や眼球・頭部運動による時間的変化と一致する。そうで あれば,輝度変化が滑らかであっても鮮明であっても不 自然さが加わり,結果として不快感が上昇すると考えら れる(Figure 2)。なお,空間次元では低空間周波数成分 が強調されたブラー画像に対し不快感が上昇することが 報告されている(O’Hare & Hibbard, 2013)。この説明と して,ブラー画像へは焦点調節応答が変動する調節微動 が生じやすいことが不快感に結びつくと論じられている が,Yoshimoto et al. (2016)で用いられた視覚刺激は輝 度が一様に変化するフリッカであり,調節反応との関連 性が低いことから,輝度変化が滑らかな時間的ブラーに 対して生じる不快感には眼球運動の制御とは異なる視覚 情報処理機構が関与すると考えられる。 各時間周波数成分の位相がランダムに変化するフリッ カを用いた場合には,時間的ブラーが減少し輝度変化が 鮮明になるほど不快感は上昇する(Figure 2(d))。位相 が等しい場合には一定の間隔で出現するエッジが存在す るが,位相がランダムに変化する場合にはそれがないと いう点で両者は異なる(Figure 3(a))。ただし,それが なぜ不快感に影響するのかは明らかでない。ランダム波 に基づくフリッカが不快感を喚起する理由として,第一 にランダム波に基づく輝度変化は矩形波のような位相が 等しい波形に基づく輝度変化に比べ予測が困難であるこ とが挙げられる。スパースコーディングの観点からすれ ば,予測が困難な状況はより多くの神経活動量を要し, それが不快感に繋がると考えられる。この説明に基づけ ば,位相が等しい条件,すなわち輝度変化が単調で予測 しやすい場合には,少ない神経活動量で情報処理を行う ことができるため,不快感が上昇しないのだと言える。 第二に,中高域の時間周波数成分に対する抑制的な働 きの強さが原因になっている可能性がある。空間次元に おいては,基本波と高調波の位相が等しい場合には中高 域の空間周波数成分に対する順応は起きにくい(Mizoka-mi, Paras, & Webster, 2004; Nachmias, Sansbury, Vassilev, & Weber, 1973; Tolhurst, 1972; Tolhurst & Barfield, 1978)。し かしながら,高調波の位相がランダムに変動するなどし て基本波の位相と一致しない場合にはその限りでなく, 中高域の空間周波数成分に対する抑制的な応答は見られ にくくなる(Klein & Stromeyer, 1980)。時間次元におい ても同様の処理機構が備わっているのであれば,ランダ ム波に基づく輝度変調パターンの不快感が高い原因とし て,感度の高い中域の時間周波数成分が抑制されないこ とが考えられる。位相が視覚的不快感に与える影響につ いては,今後さらなる検討が必要である。 5. 今後の展望 本稿の最後に,これまでの議論を総括する。静止した パターンを用いた場合,不快感に関連する要因として, 以下の2点が挙げられる(Wilkins, 2016)。 (1)中域の空間周波数帯域における過度の振幅 (2)自然情景統計量(1/f)からの乖離 時間的に変化するパターンがもたらす不快感について も,ある程度は上記の「空間周波数」を「時間周波数」 に置き換えて記述することができる。一方で,位相が視 覚的不快感に与える影響は時間次元と空間次元で大きく 異なる可能性がある。次節以降では,視覚的不快感にお ける時間特性と空間特性の相互作用を中心に,今後考え るべき問題について論じる。 5.1. ブラー 位相と関連して,まずはブラーを取り上げる。空間次 元においては,各空間周波数成分の位相をランダムに変 動させるとランダムドットパターンが生成される。ラン ダムドットパターンに対する不快感は,その振幅スペク トルが1/fを示さない時に上昇する。とりわけ低空間周 波数成分を強調し,空間的ブラーを強くすると不快感が 高まる(Juricevic et al., 2010; O’Hare & Hibbard, 2013)。一 方で,ランダム波に基づくフリッカへの不快感は,低時 間周波数成分を強調し,時間的ブラーを強くするほど低 下する。(Yoshimoto et al., 2016)。これは,時間的ブラー と空間的ブラーでは不快感に与える影響が異なることを 示している。 時間・空間の両次元で変化する運動パターンを刺激と して用い,双方で振幅スペクトルを操作すると,不快感 にどのような影響が見られるであろうか?空間次元では
ブラー成分が多いと不快感が上昇し,時間次元ではブ ラー成分が少なくエッジが鮮明であるほど不快感が上昇 するため,直観的には低空間周波数と高時間周波数を強 調することが強い不快感を誘発すると予測される。 一方で,こうした時空間次元で映像を変調した場合, それに対する知覚や眼球運動も考慮しなければならな い。例えば,運動によりブラー画像が鮮明に見える錯視 現象が知られており(Bex, Edgar, & Smith, 1995),実際に 自然映像に低域通過型空間周波数フィルタをかけて不鮮 明にすると,高域通過型空間周波数フィルタをかけた時 よ り も映 像 が 鮮 明 に 見 え る こ と が 報 告 さ れ て い る (Takeuchi & De Valois, 2005)。また,均一な輝度変調のみ のフリッカとは異なり,空間属性を持つ映像において は,その鮮明度が調節微動の生起に影響を及ぼす(Day, Gray, Seidel, & Strang, 2009)。視覚的な不快感にとっては 画像の物理的なブラーが重要なのか,あるいはブラーに よりもたらされる知覚が重要なのか,どちらの可能性も 考えられる。 こうしたブラーといった映像の変調を通して視覚の不 快感を検討することにより,不快感の生成機序において 視覚の時間特性と空間特性がどのように相互作用してい るのか,その点の理解が深まると考えられる。 5.2. 視覚的質感 視覚的不快感において時間・空間の両特性が関与する 興味深い問題として,水,煙,炎といった非剛体素材の 認識に関する質感知覚がある。Kawabe, Maruya, Fleming, & Nishida (2015)はノイズパターンを用い,輝度変調に より生成した運動情報から液体らしさの認識が可能であ ることを発見した。彼らは,オプティカルフローの滑ら かさが液体らしさの印象を高めることを示している。質 感知覚においては,視覚系には自然界に存在する素材の 物理特性に関する単純な視覚的特徴を利用して効率的に 処理するメカニズムが備わっていると想定されている (Fleming, 2014; Kawabe et al., 2015; Kawabe, Maruya, &
Nishida, 2015; Marlow, Kim & Anderson, 2012; Motoyoshi, Nishida, Sharan, & Adelson, 2007; Norman, Wiesemann, Nor-man, Taylor, & Craft, 2007)。これは,質感知覚がスパー スコーディングにより処理されていることを示唆する。 そうであるとすれば,例えば液体らしさの質感知覚に関 しては,オプティカルフローを操作し液体らしさの印象 が損なわれると脳は効率的に表現できなくなり,結果と して不快感を喚起する可能性がある。なお,静止した自 然物体の質感がもたらす視覚的不快感については,質感 の認知プロセスとは独立に低次の自然情景統計量から直 接的に処理される経路の存在が指摘されている(森・本 吉,2016)。時間的側面が関わる質感知覚がもたらす不 快感にも同様の処理経路が存在するか,さらなる研究が 必要である。 5.3. 神経基盤 視覚的不快感の生成機序として有力視されている説 は,振幅スペクトルが1/fに収束する自然情景統計量を 持つパターンはスパースコーディングにより効率的に処 理されるという仮定を前提とする。自然情景と類似の時 空間的特徴(1/f)を持たないパターンを処理するため にはより多くの神経細胞が応答しなければならず,それ が視覚的不快感を生成すると考えられている(Hibbard
& O’Hare, 2015; Juricevic et al., 2010; Wilkins, 2016)。また, 静止画が動いて見えるエニグマ錯視に関しては,錯視の 効果により喚起される不快感の程度が高いときに脳波反
応が増強することが報告されており(O’Hare, Clarke, &
Pollux, 2016),スパースコーディングに基づく説明が支 持されている。 一方で先に述べた通り,空間次元においては振幅スペ クトルが1/fを示すパターンに対してV1, V2, V3の神経 応答量が増加するという報告もなされている(Isher-wood et al., 2017),これは,少なくとも比較的初期の視 覚野においてはスパースコーディング仮説が支持されな い可能性を示している。なお時間的に変化するパターン がもたらす不快感と脳活動との関係は検討されておら ず,こうした議論が時間次元においても当てはまるのか どうかは明らかでない。視覚的不快感と神経応答量の関 係については,例えば脳のより高次領域における活動を 測定するなど,今後さらなる検討が必要である。 視覚的不快感の神経基盤に関して検討されていないそ の他の点として,慣れの問題がある。つまり,視覚的に 不快な時空間パターンに順応すると,不快な印象は変化 するのであろうか?この問題は,視覚的不快感の神経基 盤を検討する上で重要であると考える。順応には感度の 調整や符号効率化などの機能がある(Webster, 2014)。 視覚的不快感をもたらす時空間パターンへの順応前後に おける印象変化と脳活動変化を対応させることにより, 神経基盤を特定できる可能性がある 5.4. 眼球運動 本稿では周波数特性に着目し視覚的不快感の生成機序 について論じたが,照明やテレビ画面によるフリッカに 関しては主に眼球運動との関連性が検討されている。見 たい対象物に視線を向けるための急速な眼球運動(サッ
カード)を行っている間,網膜像が知覚されることはま ずない。これはサッカード抑制と呼ばれ,抑制的な応答 をする細胞の働きやサッカード前後の視対象によるマス
キングが原因で生じると考えられている(Bremmer,
Kubischik, Hoffman, & Krekelberg, 2009; Campbell & Wurts, 1978)。
Castet & Masson (2000)によると,網膜上における像 の移動速度が固視微動による像の移動速度を超えること でサッカード抑制が生じる。運動情報の処理に関与する 第 5次視覚野(V5/MT)へ選択的に投射するV1の大細 胞は固視微動による像の移動速度にピーク感度を示す が,その応答範囲にサッカード時の像の移動速度は含ま れないため抑制が起きると説明されている。サッカード 時の網膜上の像は連続していることから,サッカード抑 制は連続した像の運動に対して生じると考えられる。照 明やテレビ画面においては,たとえその時間周波数が CFFを上回っていたとしても視対象が時間的に不連続か つ断片的に網膜に投影される。そのため,通常の抑制が 生じずサッカード中の網膜像が知覚されることがあり, それが不快感に繋がると指摘されている(Roberts & Wilkins, 2013)。 実験室内では実験参加者がサッカードをしないよう注 視点を提示することが多いが,自然な状況ではサッカー ドをはじめとする眼球運動が頻繁に行われている。視覚 的不快感における眼球運動の影響と周波数特性の関連性 は未解明の問題であり,研究の進展が待たれるところで ある。 5.5. 個人差 最後に,視覚的不快感の個人差に言及する。本稿のは じめに,時間的に変化するパターンによる不快感や頭 痛,光感受性発作の事例を紹介したが,いずれも個人差 の大きいものである。例えば「ポケットモンスター」は 筆者も1997年当時視聴していたが,問題となったシー ンで光感受性発作はおろか特別不快に感じた記憶はな い。一方で,発作を起こして病院に搬送された者もい た。このような個人差をもたらす原因の一つとして, V1における抑制系の関与が挙げられる。
Meldrum & Wilkins (1984)は神経薬理学的研究から, 抑制性神経伝達物質であるγ-アミノ酪酸(GABA)の拡 散が不十分であることが光感受性発作を誘発する可能性 が あ る と指 摘 し た。Penacchio, Otazu, Wilkins, & Harris (2015)は視覚的不快感と V1 の神経活動量のモデル (Hibbard & O’Hare, 2015)に抑制系の働きを加えたモデ
ルを提案している。
近年,磁気共鳴の原理により代謝物を測定する磁気共 鳴分光法(magnetic resonance spectroscopy, MRS)により, 人間の脳内神経伝達物質濃度を測定することが可能に なった。MRS研究から,視覚的方位検出や曖昧図形の 双安定性知覚といった視知覚における個人差と神経伝達 物質濃度の関係性まで調べられることが明らかになり注 目を集めている(Edden, Muthukumaraswamy, Freeman, & Singh, 2009; van Loon et al., 2013)。Takeuchi, Yoshimoto, Shimada, Kochiyama, & Kondo (2017)は仮現運動による 錯視現象の一つである運動の同化・対比の個人差が前頭 前野における興奮性神経伝達物質グルタミン酸の濃度と 関係している可能性を示した。 こうしたMRS 研究から,成人よりも若年者の方が, 大脳基底核において興奮性神経伝達物質であるグルタミ ン酸濃度が高い傾向にあることが示されている(Boll-mann et al., 2015)。光感受性発作は15歳以下の若年者に 好発することが知られていることから(Eeg-Olofsson, Petersen, & Sellden, 1971; Petersen, Eeg-Olofsson, 1971),興 奮系の働きが強く,抑制系の働きが相対的に弱いことが 原因で若年者に光感受性発作や不快感が生じる可能性も ある。MRSにより脳の各領域における興奮性・抑制性 の脳内神経伝達物質濃度を測定することで,視覚的不快 感に見られる個人差の神経基盤を検討することができる と期待される。 昨今の映像メディア技術の革新的な発展に伴い,映像 利用の機会は増加の一途を辿っている。映像は時間・空 間属性に基づくため,時間・空間の両側面から視覚的不 快感をもたらす原因解明を目指すことは,視覚メカニズ ムを理解し映像の生体安全性を実現する上で必須とな る。今後は視覚的不快感の時間特性と空間特性,及び双 方の相互作用,そして個人差を対象とした研究の進展が 期待される。 引用文献
Atick, J. J. (1990). Could information-theory provide an eco-logical theory of sensory processing? Network, 22, 4–44. Atick, J. J., & Redlich, A. N. (1992). What does the retina know
about natural scenes? Neural Computation, 4, 196–210. Barlow, H. B. (1981). The Ferrier Lecture, 1980. Critical
limit-ing factors in the design of the eye and visual cortex. Pro-ceedings of the Royal Society of London, Section B, 212, 1–34. Benham, C. E. (1894). The artificial spectrum top. Nature, 51,
200.
Bex, P. J., Edgar, G. K., & Smith, A. T. (1995). Sharpening of drifting, blurred images. Vision Research, 35, 2539–2546. Bilson, A. C., Mizokami, Y., & Webster, M. A. (2005). Visual
of America A, 22, 2281–2288.
Bollmann, S., Ghisleni, C., Poil, S. S., Martin, E., Ball, J., Eich-Höchli, D., . . . O’Gorman, R. L. (2015). Developmental changes in gamma-aminobutyric acid levels in attention-deficit/hyperactivity disorder. Translational Psychiatry, 5, 1–8.
Bremmer, F., Kubischik, M., Hoffman, K-P, & Krekelberg, B. (2009). Neural dynamics of saccadic suppression. Journal of Neuroscience, 29, 12374–12383.
Burton, G. J., & Moorhead, I. R. (1987). Color and spatial structure in natural scenes. Applied Optics, 26, 157–170. Campbell, F. W., & Robson, J. G. (1968). Application of
Fouri-er analysis to the visibility of gratings. The Journal of Physi-ology, 197, 551–566.
Campbell, F. W., & Wurts, R. H. (1978). Saccadic omission: Why we do not see a grey-out during a saccadic eye move-ment. Vision Research, 18, 1297–1303.
Castet, E., & Masson, G. S. (2000). Motion perception during saccadic eye movements. Nature Neuroscience, 3, 177–183. Chatrian, G. E., Lettich, E., Miller, L. H., & Green, J. R. (1970).
Pattern-sensitive epilepsy. I. An electrographic study of its mechanisms. Epilepsia, 11, 125–149.
Cole, G. G., & Wilkins, A. J. (2013). Fear of holes. Psychologi-cal Science, 24, 1980–1985.
Day, M., Gray, L. S., Seidel, D., & Strang, N. C. (2009). The re-lationship between object spatial profile and accommoda-tion microfluctuaaccommoda-tions in emmetropes and myopes. Journal of Vision, 9, 1–13.
de Lange Dzn, H. (1958). Research into the dynamic nature of the human fovea̶cortex systems with intermittent and modulated light. I. Attenuation characteristics with white and colored light. Journal of Optical Society of America, 48, 777–784.
Dong, D. W., & Atick, J. J. (1995a). Statistics of natural time-varying images. Network: Computation in Neural Systems, 6, 345–358.
Dong, D. W., & Atick, J. J. (1995b). Temporal decorrelation: A theory of lagged and nonlagged responses in the lateral ge-niculate nucleus. Network: Computation in Neural Systems, 6, 159–178.
Edden, R. A., Muthukumaraswamy, S. D., Freeman, T. C., & Singh, K. D. (2009). Orientation discrimination perfor-mance is predicted by GABA concentration and gamma os-cillation frequency in human primary visual cortex. Journal of Neuroscience, 29, 15721–15726.
Eeg-Olofsson, O., Petersen, I., & Sellden, U. (1971). The devel-opment of the electroencephalogram in normal children from the age of 1 through 15 years: Paroxysmal activity, Neuropadiatrie, 2, 375–404.
Fernandez, D., & Wilkins, A. J. (2008). Uncomfortable images in art and nature. Perception, 37, 1098–1113.
Field, D. J. (1987). Relations between the statistics of natural images and the response properties of cortical cells. Journal of the Optical Society of America A, 4, 2379–2394.
Fisher, R. S., Harding, G., Erba, G., Barkley, G. L., & Wilkins, A., (2005). Photic- and pattern-induced seizures: A review
for the Epilepsy Foundation of America Working Group. Epilepsia, 46, 1426–41.
Fleming, R. W. (2014). Visual perception of materials and their properties. Vision Research, 94, 62–75.
Haigh, S. M., Cooper, N. R., & Wilkins, A. J. (2015). Cortical excitability and the shape of the haemodynamic response. Neuroimage, 111, 379–384.
Harding, G. F. A., & Harding, P. F. (1999). Televised material and photosensitive epilepsy. Epilepsia, 40, 65–69.
Harding, G. F. A., & Jeavons, P. M. (1994). Photosensitive epi-lepsy. Cambridge: Cambridge University Press.
Hibbard, P. B., & O’Hare, L. (2015). Uncomfortable images produce non-sparse responses in a model of primary visual cortex. Royal Society Open Science, 2, 1–8.
Huang, J., Cooper, T. G., Satana, B., Kaufman, D. I., & Cao, Y. (2003). Visual distortion provoked by a stimulus in mi-graine associated with hyperneuronal activity. Headache: The Journal of Head and Face Pain, 43, 664–671.
Isherwood, Z. J., Schira, M. M., & Spehar, B. (2017). The tun-ing of human visual cortex to variations in the 1/fα ampli-tude spectra and fractal properties of synthetic noise imag-es. Neuroimage, 146, 642–657.
Jeavons, P. M., & Harding, G. F. A. (1975). Photosensitive epi-lepsy: A review of the literature and a study of 460 patients. In Clinics in developmental medicine (No. 56, pp. 1–121). London: William Heinemann Medical Books.
Juricevic, I., Land, L., Wilkins, A., & Webster, M. A. (2010). Visual discomfort and natural image statistics. Perception, 39, 884–899.
Kawabe, T., Maruya, K., Fleming, R. W., & Nishida, S. (2015). Seeing liquids from visual motion. Vision Research, 109, 125–138.
Kawabe, T., Maruya, K., & Nishida, S. (2015). Perceptual transparency from image deformation. Proceedings of the National Academy of Sciences, USA, 112, E4620–E4627. Kelly, D. H. (1961). Visual responses to time-dependent
stim-uli. I. Amplitude sensitivity measurements. Journal of the Optical Society of America, 51, 422–429.
Klein, S., & Stromeyer, C. F., III. (1980). On inhibition be-tween spatial frequency channels: Adaptation to complex gratings. Vision Research, 20, 459–466.
Lennie, P. (2003). The cost of cortical computation. Current Biology, 13, 493–497.
Lin, M. W., Hsieh, P. H. Chang, E. C., & Chen, Y. C. (2014). Flicker-glare and visual-comfort assessments of light emit-ting diode billboards. Applied Optics, 53, E61–E68. Marcus, D. A., & Soso, M. J. (1989). Migraine and
stripe-in-duced visual discomfort. Achives of Neurology, 46, 1129– 1132.
Marlow, P. J., Kim, J., & Anderson, B. L. (2012). The percep-tion and misperceppercep-tion of specular surface reflectance. Cur-rent Biology, 22, 1909–1913.
Meldrum, B. S., & Wilkins, A. J. (1984). Photosensitive epilep-sy: Integration of pharmacological and psychophysical evi-dence. In P. Schwatzkroin, & H. Wheal (Eds.), Electrophysi-ology of epilepsy (pp. 51–77). London: Academic Press.
Mizokami, Y., Paras, C., & Webster, M. A. (2004). Chromatic and contrast selectivity in color contrast adaptation. Visual Neuroscience, 21, 359–364.
森 詩織・本吉 勇 (2016).視覚的質感の情動的価値 に関わる潜在的情報処理過程.日本基礎心理学会第 35回大会プログラム,80.
Motoyoshi, I., Nishida, S., Sharan, L., & Adelson, E. H. (2007). Image statistics and the perception of surface qualities. Na-ture, 447, 206–209.
Nachmias, J., Sansbury, R., Vassilev, A., & Weber, A. (1973). Adaptation to square-wave gratings: In search of the elusive third harmonic. Vision Research, 13, 1335–1342.
Norman, J. F., Wiesemann, E. Y., Norman, H. F., Taylor, M. L., & Craft, W. D. (2007). The visual discrimination of bending. Perception, 36, 980–989.
O’Hare, L., Clarke, A. D. F., & Pollux, P. M. J. (2016). VEP Re-sponses to Op-Art Stimuli. PLoS ONE, 10, 1–18.
O’Hare, L., & Hibbard, P. B. (2011). Spatial frequency and vi-sual discomfort. Vision Research, 51, 1767–1777.
O’Hare, L., & Hibbard, P. B. (2013). Visual discomfort and blur. Journal of Vision, 13, 1–12.
Olshausen, B. A., & Field, D. J. (2004). Sparse coding of senso-ry inputs. Current Opinion in Neurobiology, 14, 481–487. Penacchio, O., Otazu, X., Wilkins, A. J., & Harris, J. (2015).
Uncomfortable images prevent lateral interactions in the cortex from providing a sparse code. Perception, 44, 67–68 (ECVP Supplement).
Petersen, I., & Eeg-Olofsson, O. (1971). The development of the electroencephalogram in normal children from the age of 1 through 15 years: Non-paroxysmal activity, Neuropadi-atrie, 2, 247–304.
Poplawski, M. E., & Miller, N. E. (2013). Flicker in solid-state lighting: Measurement techniques, and proposed reporting and application criteria. In Proceedings of the CIE Midterm and Centenary Conference, Paris, April 2013.
Roberts J. E., & Wilkins, A. J. (2013). Flicker can be perceived during saccades at frequencies in excess of 1 kHz. Lighting Research and Technology, 45, 124–132.
Snow, M., Coen-Cagli, R., & Schwartz, O. (2016). Specificity and timescales of cortical adaptation as inferences about natural movie statistics. Journal of Vision, 16, 1–27. Snowden, R. J., & Hess, R. F. (1992). Temporal frequency
fil-ters in the human peripheral visual field. Vision Research, 32, 61–72.
So, R. H., & Ujike, H. (2010). Visually induced motion sick-ness, visual stress and photosensitive epileptic seizures: What do they have in common?: Preface to the special issue. Applied Ergonomics, 41, 491–493.
Takahashi, T., & Tsukahara, Y. (1998). Pocket Monster inci-dent and low luminance visual stimuli: Special reference to deep red flicker stimulation. Acta Paediatrica Japonica, 40, 631–637.
Takeuchi, T., & De Valois, K. K. (2005). Sharpening image mo-tion based on the spatio-temporal characteristics of human
vision. Proceedings of SPIE-IS&T Electronic Imaging: Human Vision and Electronic Imaging X, 5666, 83–94.
Takeuchi, T., Yoshimoto, S., Shimada, Y., Kochiyama, T., & Kondo, H. M. (2017) Individual differences in visual mo-tion percepmo-tion and neurotransmitter concentramo-tions in the human brain. Philosophical Transactions of the Royal Society B, 372, 20160111.
Tolhurst, D. J. (1972). Adaptation to square-wave gratings: In-hibition between spatial frequency channels in the human visual system. Journal of Physiology, 226, 231–248.
Tolhurst, D. J., & Barfield, L. P. (1978). Interactions between spatial frequency channels. Vision Research, 18, 951–958. Tolhurst, D. J., Tadmour, Y., & Chao, T. (1992). Amplitude
spectra of natural images. Ophthalmic Physiological Optics, 12, 229–232.
van Hateren, J. H. (1993). Spatiotemporal contrast sensitivity of early vision. Vision Research, 33, 257–267.
van Hateren, J. H., & Ruderman, D. L. (1998). Independent component analysis of natural image sequences yields spa-tio-temporal filters similar to simple cells in primary visual cortex. Proceedings of the Royal Society of London, Section B, 265, 2315–2320.
van Hateren, J. H., & van der Schaaf, A. (1996). Temporal properties of natural scenes. Proceedings of SPIE, 2657, 139– 143.
van Loon, A. M., Knapen, T., Scholte, H. S., St John-Saaltink, E., Donner, T. H., & Lamme, V. A. (2013). GABA shapes the dynamics of bistable perception. Current Biology, 23, 823– 827.
Watson, A. B., & Robson, J. G. (1981). Discrimination at threshold: Labelled detectors in human vision. Vision Re-search, 21, 1115–1122.
Webster, M. A. (2014). Probing the functions of contextual modulation by adapting images rather than observers. Vi-sion Research, 104, 68–79.
Wilkins, A. J. (1995). Visual stress. Oxford: Oxford University Press.
Wilkins, A. J. (2016). A physiological basis for visual discom-fort: Application in lighting design. Lighting Research & Technology, 48, 44–54.
Wilkins, A. J., Nimmo-Smith, I., Slater, A., & Bedocs, L. (1989). Fluorescent lighting, headaches and eyestrain. Light-ing Research and Technology, 21, 11–18.
Wilkins, A., Nimmo-Smith, I., Tait, A., McManus, C., Della Sala, S., Tilley, A., Arnold, K., Barrie, M., & Scott, S. (1984). A neurological basis for visual discomfort. Brain, 107, 989– 1017.
Yoshimoto, S., Garcia, J., Jiang, F., Wilkins, A. J., Takeuchi, T., & Webster, M. A. (2016). Visual discomfort and temporal flicker. The 31th International Congress of Psychology, RC-11–169.