3 3 3 3...ルーメンフローラ.ルーメンフローラルーメンフローラ解析ルーメンフローラ解析解析解析 1 1 1 1)))生菌剤)生菌剤生菌剤(生菌剤(BLCS)(( )))のののの影響影響影響 影響 ①目的 BLCS ルナシータは、日本エメラル株式会社により、米ぬか、小麦フスマ、大豆粕などを 吸着剤として枯草菌、乳酸菌、酵母などの微生物を混合発酵させて製造されたものである。 これを牛および羊に投与することにより、ルーメンでのメタン生成が抑制できることが報 告されている(高浦ら、第 106 回日本畜産学会)。しかし、BLCS ルナシータの効果がメタ ン生成菌に対して直接的に作用しているのか、あるいは他のルーメン細菌を介した間接的 な作用であるかは明らかにされていない。そこで本試験ではBLCS ルナシータを羊に投与 することにより生じるルーメンフローラの変化(メタン生成菌数の低下、BLCS ルナシータ に含まれる細菌の増加、水素消費細菌の増加など)を明らかにすることを目的とした。 ②試験対象サンプル 試験対象サンプルは帯広畜産大学殿から提供を受けたルーメン液とBLCS ルナシータであ る。ルーメン液は給与飼料のみを投与(対照区)、モネンシンを1 日当たり20mg 投与(モネ ンシン区)、BLCS ルナシータを1 日当たり4g 投与(4g 区)およびBLCS ルナシータを1 日当たり8g 投与(8g 区)したチェビオット種去勢羊から採取されたものである。 羊への給与 条件は以下の通りである。 対照区 給与飼料のみを投与 モネンシン区 モネンシンを1 日当たり20mg 投与 4g 区 BLCS ルナシータを1 日当たり4g 投与 8g 区 BLCS ルナシータを1 日当たり8g 投与 給与期間を20 日とし、14 日間の間隔を空けたものを1 サイクルとして、Ⅰ期からⅣ期ま での4 サイクルの試験にわたり、ルーメン液の採取が行なわれた。 ③試験方法 本試験ではDGGE 法、T-RFLP 法およびMPN-PCR 法の三種類を用いた。 DGGE(変性グラジエントゲル電気泳動)法を用いるルーメンフローラの解析対象として、真 正細菌、古細菌、真核生物とした。真正細菌の解析により、BLCS ルナシータ投与による BLCS ルナシータに含まれる細菌の出現および水素消費細菌の増減を把握することを目的 とした。メタン生成菌を含む古細菌の解析により、ルーメン内に含まれるメタン生成菌の 種類を特定することを目的とした。酵母を含む真核生物の解析により、BLCS ルナシータ に含まれる酵母がルーメン内における増加について明らかにすることを目的とした。 T-RFLP(制限酵素末端断片長解析)法を用いるルーメンフローラの解析対象は真正細菌とし
DNA バ ン ド が 単 一 に な る まで数回繰り返す。 た。DGGE 法の解析結果について、真正細菌の増減を定量的に裏付けることを目的として 実施した。 MPN-PCR 法(最確数-PCR)により、BLCS ルナシータがメタン生成菌数の低減に効果を有 しているかを定量的に確認することを目的とした。 ④ 試験1.PCR-DGGE 法によるルーメンフローラの解析 ④-1 PCR-DGGE 法のフロー 図4-1 にPCR-DGGE 法による微生物群集構造解析のフローを示す。 1. ルーメン液からのDNA 抽出・精製 ↓ 2. PCR 反応(プライマー:GC クランプ付) 1)a,b,c 真正細菌・古細菌・真核生物を対象としたPCR 条件(1 回目) 2)a,b,c 真正細菌・古細菌・真核生物を対象としたPCR 反応(GC クランプ付き) ↓ 3. アガロース電気泳動による16S rDNA の増幅の確認 ↓ 4. DGGE 解析 ↓ 5. DNA バンド切り出しとDNA 抽出・精製 ↓ 6. PCR 反応(プライマー:GC クランプの除去) ↓ 7. PCR 産物の精製 ↓ 8. シーケンス解析 ↓ 10.ホモロジー解析(微生物種の同定) 図4-1 PCR-DGGE 法による微生物群集構造解析のフロー ④-2 ルーメン液からのDNA 抽出・精製 ルーメン液にはDNA抽出が困難な細菌およびPCR阻害物質が存在すると考えられる。こ のため、本試験では強固な細胞壁を備えた微生物の物理的破壊工程、界面活性剤およびそ の他薬剤を65℃で作用させるDNA 抽出工程、腐植物質(或いは前駆物質であるリグニン) の除去工程を含む方法(Yoshida et al. 2005)を用いた。
④-3 PCR 反応
解析対象は真正細菌、古細菌および真核生物とした。
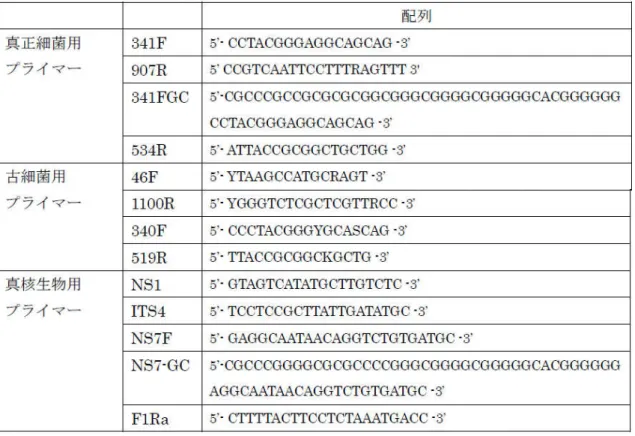
表 4-1 に真正細菌、古細菌および真核生物を対象としたプライマーの配列を示した。 表 4-1 真正細菌を対象としたPCR に用いたプライマーの配列
④-3-1a) 真正細菌を対象としたPCR 条件(1 回目) 抽出したDNA を鋳型として、真正細菌用のプライマー(341F、907R)を用いて、16SrDNA を増幅した。PCR 条件を図4-2 に示した。 [ PCR反応(Primer : 341F, 907R) ] 96℃ 5min 96℃ 1min 55℃ 30sec 25cycle 72℃ 1min 72℃ 5min 4℃ ∞ 図4-2 真正細菌の16SDNA を標的としたPCR 反応条件 ④-3-2a) 真正細菌を対象としたPCR 反応(GC クランプ付き) DGGE ゲルを用いた電気泳動を行うため、GC クランプ付(GC に富む配列)プライマー を用いたPCR を行った。図4-2 で得られたPCR 産物を鋳型として、真正細菌用GC ク ランプ付きプライマー(341FGC,534R)を用いてPCR を行い(図4-3)、アガロースゲル 電気泳動にてPCR 産物の増幅を確認した。 [ PCR反応(Primer : 341FGC, 534R) ] 96℃ 5min 96℃ 1min 55℃ 30sec 25cycle 72℃ 30sec 72℃ 5min 4℃ ∞ 図4-3 真正細菌の16SDNA を標的としたPCR 反応条件(GC クランプ付)
④-3-1b) 古細菌を対象としたPCR 条件(オプション) 抽出したDNA を鋳型として、古細菌用のプライマー(46F、1100R)を用いて16S rDNAを 増幅した。PCR 条件を図4-4 に示した。 〔PCR 反応(Primer: 46F, 1100R) 〕 94℃ 10min 92℃ 1min 53.5℃ 30sec 30cycle 72℃ 1min 72℃ 6min 4℃ ∞ 図4-4 古細菌の16S rDNA を標的としたPCR 反応条件 ④-3-2b) 古細菌を対象としたPCR 反応(GC クランプ付き) (オプション) DGGE ゲルを用いた電気泳動を行うため、真正細菌と同様、GC クランプ付プライマーを 用いたPCR を行った。図4-4 で得られたPCR 産物を鋳型として、古細菌用GCクランプ 付きプライマー(340FGC、519R)を用いて PCR を行い(図4-5)、アガロースゲル電気 泳動にてPCR 産物の増幅を確認した。 〔PCR 反応(Primer: 340F, 519R) 〕 94℃ 10min 92℃ 1min 65℃ 30sec 30cycle 72℃ 1min 72℃ 6min 4℃ ∞ 図4-5 古細菌の16SDNA を標的としたPCR 反応条件(GC クランプ付)
④-3-1c) 真核生物を対象としたPCR 条件(オプション)
抽出した DNA を鋳型として、真核生物用のプライマー(NS1、ITS4)を用いて 18S rDNA を増幅した。PCR 条件を図4-6 に示した。 〔PCR 反応(Primer: NS1, ITS4) 〕 96℃ 5min 96℃ 1min 40℃ 1min 30cycle 72℃ 1min 72℃ 5min 4℃ ∞ 図4-6 真核生物の18S rDNA を標的としたPCR 反応条件 ④-3-2c) 真核生物を対象としたPCR 反応(GC クランプ付き) (オプション) DGGE ゲルを用いた電気泳動を行うため、真正細菌、古細菌と同様、GC クランプ付プラ イマーを用いた PCR を行った。図 4-6 で得られた PCR 産物を鋳型として、真核生物用 GC クランプ付きプライマー(NS7-GC、F1Ra)を用いてPCR を行い(図4-7)、アガロ ースゲル電気泳動にてPCR 産物の増幅を確認した。 〔PCR 反応(Primer: NS7-GC, F1Ra) 〕 96℃ 5min 96℃ 1min 60℃ 30sec 30cycle 72℃ 30sec 72℃ 5min 4℃ ∞ 図4-7 真核生物類をターゲットとした nested PCR 条件
④-4 DGGE 解析 広範囲のバンドが得られるように、ゲル中のDNA 変性剤の濃度勾配を35-60%(真正細菌、 古細菌)および30-70%(真核生物類)(変性剤100%とは、7M 尿素、40%ホルムアミドに相 当)として DGGE 解析を行った。また、重合剤の濃度は、低濃度に設定し、ゲルを 4 時 間固化させることにより、グラジエントの精度を高めたゲルを作成した。さらに、バンド の分離能を高めるため、電圧を低くし、泳動時間を長く設定した(表4-2)。 表4-2 DGGE ゲル濃度と泳動条件 ポリアクリルアミドゲル濃度 :10%(真正細菌、古細菌) :6%(真核生物) 変性剤濃度勾配 :35-60%(真正細菌、古細菌)、 :30-70%(真核生物) 重合剤濃度 10% Ammonium Persulfate :0.7% TEMED :0.07% 泳動電圧 :70V 泳動時間 :960min 泳動温度 :60℃ ④-5 DNA バンドの切り出しとDNA 抽出・精製 電気泳動後、エチジウムブロマイドで染色を行い、脱染色後、UV トランスイルミネータ ー上でバンドを検出した。検出したバンドを滅菌されたメスで切り出し、1.5ml チューブ に入れた。切り出したゲルに、100μl のTE buffer を加え、35℃で一晩振とうさせた後、 エタノール沈殿法によりDNA の抽出・精製を行った。 DGGE ゲルからのバンド切り出しDNA シーケンスを行う基準を以下に示す。 ・BLCS ルナシータ投与区において増減の見られるバンド ・BLCS ルナシータに含まれるバンドに近いもの、とした。 なお、分離能が低く、複数のバンドが混在している可能性のあるバンドについては、バン ドが単一になるまでGC クランプ付PCR およびDGGE 解析を繰り返した。
④-6 PCR 反応(GC クランプの除去) 各バンドから抽出した DNA を鋳型として、シーケンス反応を行うため、GC クランプを 外すPCR を行った。 1) 真正細菌のGC クランプ無しPCR 真正細菌のGC クランプを外すPCR の条件を図4-8 に示した。 〔PCR反応(Primer : 341F, 534R)〕 96℃ 5min 96℃ 1min 55℃ 30sec 25cycle 72℃ 30sec 72℃ 5min 4℃ ∞ 図4-8 GC クランプの除去に用いたPCR 反応条件(真正細菌) 2) 古細菌のGC クランプ無しPCR(オプション) 古細菌のGC クランプを外すPCR の条件を図4-9 に示した。 〔PCR反応 (Primer : 340F, 519R)〕 94℃ 5min 92℃ 1min 65℃ 30sec 30cycle 72℃ 30sec 72℃ 5min 4℃ ∞ 図4-9 GC クランプの除去に用いたPCR 反応条件(古細菌)
3) 真核生物のGC クランプ無しPCR(オプション) 真核生物のGC クランプを外すPCR の条件を図4-10 に示した。 〔PCR 反応(Primer : NS7F,F1Ra) 〕 96℃ 5min 96℃ 30sec 50℃ 30sec 30cycle 72℃ 30sec 72℃ 5min 4℃ ∞ 図4-10 GC クランプの除去に用いたPCR 反応条件(真核生物) ④-7 PCR 産物の精製 アガロースゲル電気泳動により、PCR 産物を確認し、次いで、アガロースから目的バンド を切り出して、精製キット(QIAquickⅡ Purification Kit; QIAGEN 社製)により、PCR 産物を精製した。
④-8 シーケンス解析
④ -7 で 精 製 を 行 っ た PCR 産 物 を 鋳 型 と し て 、 ダ イ タ ー ミ ネ ー タ ー 法 (Big Dye Terminator v1.1 Cycle Sequencing Kit;ABI 社製)によりシーケンシング反応を行った。 シーケンス解析は、ABI Prism310 Genetic Analyzer(ABI 社製)を用いて行った。
④-9 ホモロジー解析(微生物種の同定) 解読した各バンドのDNA シーケンスを基に、インターネットを利用したFASTA 検索 によりDDBJ (日本)、EMBL(ヨーロッパ)のデータベースから微生物種の同定を行った。 ④-10 PCR-DGGE 法による解析結果 ④-11 真正細菌のDGGE 解析結果 真正細菌のDGGE バンドパターンを図4-11(Ⅰ期およびⅡ期)および4-12(Ⅲ期およ びⅣ期)に示した。また、塩基配列を決定したバンドの位置を四角で囲った。
図4-11真正細菌DGGE 解析結果(Ⅰ期、Ⅱ期) (白黒反転画像)
※ BLCS: BLCS ルナシータ, con: 対照区, mone: モネンシン区, 4g: 4g 区, 8g: 8g 区を示 す。
図4-12 真正細菌DGGE 解析結果(Ⅲ期、Ⅳ期) (白黒反転画像)
※ BLCS: BLCS ルナシータ, con: 対照区, mone: モネンシン区, 4g: 4g 区, 8g: 8g 区を示 す。
④-12 古細菌のDGGE 解析結果(オプション) 古細菌のDGGE バンドパターンを図4-13 に示した。蛍光輝度のやや強いバンドが一本と 極めて薄いバンドが 2 本見られた。なお、塩基配列を決定したバンドの位置を四角で囲っ た。 図4-13 古細菌DGGE 解析結果 (白黒反転画像) ※ BLCS: BLCS ルナシータ, con: 対照区, mone: モネンシン区, 4g: 4g 区, 8g: 8g 区を示 す。
④-13 真核生物のDGGE 解析結果(オプション) 真核生物のDGGE 染色画像を図4-14 に示した。バンドが認められなかったため、塩基配 列の同定は行わなかった。 図4-14 真核生物DGGE 解析結果 (白黒反転画像) ※ BLCS: BLCS ルナシータ, con: 対照区, mone: モネンシン区, 4g: 4g 区, 8g: 8g 区を示 す。
④-14 シーケンス解析結果
シーケンス解析の結果決定した真正細菌の塩基配列を表4-3 に、古細菌の塩基配列を表4-4 に示した。
④-15.ホモロジー解析結果(微生物種の同定) 各バンドの塩基配列に基づいて DDBJ-FASTA 検索を行い、ホモロジーの高かった上位 5 種を表に示した(真正細菌: 表 4-5、古細菌: 表 4-6)。また、EMBL-FASTA 検索も行い、 ホモロジーの高い上位5 種を表4-7(真正細菌)および表4-8(古細菌)に示した。 また、それらのIdentity(相同性;%で表す)とOverlap した領域(nt:塩基対数)を表 記した。DDBJ-FASTA とEMBL-FASTA の結果を比較すると、全ての結果におい て、EMBL-FASTA の方が相同性が高いあるいは同等であった。 表4-5 真正細菌の同定結果(DDBJ-FASTA)
表4-8 古細菌の同定結果(EMBL-FASTA) ④-16 同定した細菌の生態的特徴 表4-9 に同定した真正細菌の生態的特徴を示し、表4-10 に同定した古細菌の生態的特徴を 示した。なお、4-15 に示した通り、DDBJ-FASTA と比較して高い相同性の結果が得られ たEMBL-FASTA による結果を用いた。 表4-9 真正細菌の生態特徴(EMBL-FASTA の結果に基づく)
④-17 真正細菌の出現結果
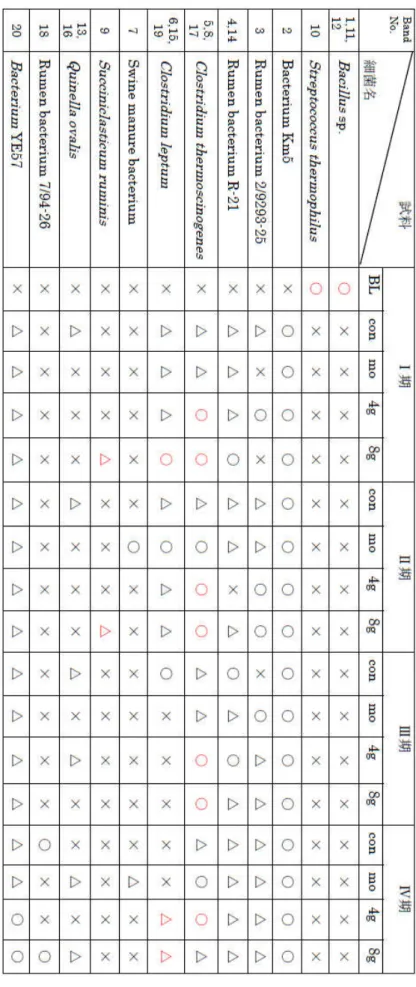
表 4-11 に DGGE 解析により得られたバンド輝度から試料に含まれる微生物量を判定 した結果を示した。細菌名の欄にはEMBL-FASTAのデータベース上に存在する細菌の16S rDNA の配列データと相同性が最も高いものを示した。
表4-11真正細菌の出現状況※BL: BLCS ルナシータ, con: 対照区, mo: モネンシン区, 4g 区8g 区
DGGE 法による解析の結果、BLCS ルナシータに含まれる細菌は BLCS ルナシータを投 与した羊のルーメン液中からは検出されなかった。第Ⅰ期においては、対照区と比較して BLCS ルナシータ4g 区でRumen bacteriumおよびClostridium thermoscinogenes のバ ンドの蛍光輝度が向上し、8g 区において Rumen bacterium、Clostridium thermoscinogenes、Clostridium leptum およびSucciniclasticum ruminis のバンドの蛍 光輝度の向上が認められた。一方、モネンシン区においては対照区と比較して大きな変化 は見られなかった。
第Ⅱ期においては、対照区と比較して4g 区でRumen bacterium およびClostridium thermoscinogenes のバンドの蛍光輝度の向上が見られ、8g 区では Rumen bacterium、 Clostridium thermoscinoges および Succiniclasticum ruminis のバンドの蛍光輝度の向 上が認められた。モネンシン区においては対照区と比較して Clostridium thermoscinogenes、Clostridium leptum、Swine manure bacterium のバンドの蛍光輝度 の向上が認められた。
第Ⅲ期においては、対照区と比較して4g 区でClostridium thermoscinogenes のバンドの 蛍光輝度の向上が見られ、Clostridium leptum のバンドの蛍光輝度の低下が認められた。 BLCS ルナシータ 8g 区では Clostridium thermoscinogenes のバンドの蛍光輝度の向上 が見られ、Rumen bacterium、Clostridium leptum およびQuinella ovalis のバンドの蛍 光輝度の低下が見られた。モネンシン区においては Rumen bacterium のバンドの蛍光輝 度の向上とClostridium leptum のバンドの蛍光輝度の低下が認められた。
第 Ⅳ 期 に お い て は 、 対 照 区 と 比 較 し て 4g 区 で Clostridium thermoscinogenes、 BacteriumYE57 お よ び Clostridium leptum の バ ン ド の 蛍 光 輝 度 の 向 上 が 見 ら れ た 。 BLCS8g 区ではQuinella ovalis、Clostridium leptum およびBacterium YE57 の蛍光輝 度の向上が見られ、Rumen bacterium の蛍光輝度の低下が認められた。モネンシン区にお いては、Clostridiumthermoscinogenes およびSwine manure bacteriumのバンドの蛍光 輝度の向上が見られた。
④-18 PCR-DGGE 法によるルーメンフローラ解析に関する考察 1) 真正細菌の解析 1) -1 BLCS ルナシータに含まれる細菌 BLCS ルナシータに含まれる細菌として、Bacillus 属およびStreptococcus 属が同定され た 。Bacillus 属 と し て 同 定 さ れ た 細 菌 は ア ル カ リ 環 境(至 適 pH10.5)を 好 む も の で あ り (Krulwich et al. 1985)、ルーメン内は中性環境(高橋 2007)であることから、ルーメン内に おいて活発に増殖した可能性は低いと考えられる。しかし、ルーメン内に生存し続けて機 能を発揮していたと仮定すると、Bacillus 属の中にはフマル酸還元酵素を有する種もある ため(Schnorpfeil M et al. 2001)、水素を消費することが可能であり、また、酪酸が主要な 代謝産物であるため、その生成過程においても水素を消費することができると考えられる。 この様な水素の消費により、メタン生成菌が利用する基質を減少させ、メタン生成菌数を 低下させうると考えられる。また、BLCS ルナシータ投与区において酪酸濃度が上昇して いるが(高橋 2007)、その一部の生産を当該Bacillus 属が担っていた可能性も考えられる。 Streptococcus 属として同定された細菌は低pH 環境下(5.0 以下)で乳酸を産生するが、本 試験に用いた羊のルーメン内pH は中性(高橋 2007)であるため、乳酸を産生した可能性は 低いと考えられる。一方、Streptococcus 属の主要代謝産物はギ酸および酢酸であるが、ギ 酸の多くは水素と二酸化炭素に分解され、その後メタンに変換される。BLCS ルナシータ 4g 区および8g 区においてメタン生成が抑制される傾向を示したこと(高橋 2007)、および ル ー メ ン 内 の 酢 酸 濃 度 が 高 か っ た こ と(高 橋 2007)か ら 、BLCS ル ナ シ ー タ に 含 ま れ る Streptococcus 属がルーメン内で生存していた場合、ギ酸よりも酢酸が主要な代謝産物にな ったと考えられる。しかし、Bacillus 属およびStreptococcus 属のいずれもBLCS ルナシ ータ投与区ではDGGE のバンドとして検出されなかった。このため、これらの細菌がルー メン内で中心となってVFA 産生に貢献していた可能性は低いと考えられる。 1) -2 BLCS ルナシータ投与によるルーメンフローラの変化 BLCS ル ナ シ ー タ 投 与 4g お よ び 8g 区 に お い て は 、 第 Ⅰ 期 、 第 Ⅱ 期 、 第 Ⅲ 期 で Clostridiumthermosuccinogenes またはClostridium sp. FG4 のバンドの蛍光輝度が対照 区と比較して向上することが認められ、4g 区においては第Ⅳ期においても蛍光輝度の向上 が見られた。Clostridium thermosuccinogenes とClostridium sp. FG4 は相同性の数値が 同じであり、二つの菌は性質が近いと考えられる。Clostridium thermosuccinogenes は多 糖類のイヌリンを唯一の炭素源として利用可能であるが(Sridhar et al. 2000)、BLCS ルナ シ ー タ の 吸 着 剤 と し て 用 い た フ ス マ に イ ヌ リ ン が 含 ま れ て い る こ と が 報 告 さ れ て い る (Roland N et al.1995)。このため、フスマまたはイヌリンの羊への給与によりこれらの細菌 が増加した可能性も考えられる。しかし、Clostridium thermosuccinogenes が増加した原 因がBLCS ルナシータに含まれる細菌か吸着剤による影響なのかは不明である。これを明 ら か に す る た め に は 、BLCS ル ナ シ ー タ の 吸 着 剤 に 細 菌 を 添 加 し て い な い も の ま た は BLCS ルナシータを滅菌したものを対照区の羊に給与して、その時のルーメンフローラを 解析することが必要であると考えられる。
al. 1991)、この酵素の反応過程で水素を消費することができる。その結果、メタン生成菌の 基質を減少させ、メタン生成菌数を低下させた可能性も考えられる。さらに Clostridium thermosuccinogenes の主要代謝産物はコハク酸と酢酸であり(Drent WJ et al. 1991)、 BLCS ルナシータ投与区においてルーメン内で酢酸濃度が高かった結果(高橋 2007)に対 して本菌が寄与した可能性も考えられる。 Clostridium sp. FG4 の主要代謝産物は酢酸で あり、Clostridium thermosuccinogenesと高い相同性(99.8%)を有している(Kato S et al. 2005)。そのため、Clostridium sp. FG4はフマル酸還元酵素を有している可能性が高いと 考えられ、本菌も水素の消費に関与している可能性が高いと考えられる。 また、BLCS ルナシータ投与4g 区においては第Ⅳ期で、8g 区では第Ⅰ期および第Ⅳ期 で Clostridium leptum が 対 照 区 と 比 較 し て 増 加 し て い た こ と が 認 め ら れ て い る 。 Clostridium leptum は酪酸産生能を有しており、その代謝過程において水素を消費する。 その結果、メタン生成菌の基質を減少させ、メタン生成菌数を低下させた可能性が考えら れる。また、BLCS ルナシータ投与区においては対照区と比較して酪酸濃度が高いこと(高 橋 2007)の原因となっている可能性も考えられる。 Succiniclasticum ruminis も 対 照 区 と 比 較 し て BLCS ル ナ シ ー タ 投 与 8g 区 に お い て DGGE バンドの蛍光輝度の向上が第Ⅰ期および第Ⅱ期で見られた。本菌はコハク酸をプロ ピオン酸に代謝する(van Gylswyk NO 1995)。この代謝過程においては水素の授受は関与 していないが、BLCS ルナシータ投与区においてSucciniclasticum ruminis が増加したこ とは、水素消費に関与するフマル酸還元酵素によるコハク酸のプロピオン酸への代謝が行 われていたことを示している可能性がある。 BLCS ルナシータには吸着剤として米ぬか・フスマ・大豆粕などの易分解性タンパク質 が使われている(高橋 2007)にも関わらず、8g 区でルーメンからの尿中への窒素流出は減少 する傾向が示されている(高橋 2007)。したがってBLCS ルナシータ投与により増加した細 菌(Clostridium thermosuccinogenes、Clostridium leptum、増加時期および投与区は前出) により菌体タンパク質の合成が促進された可能性が考えられる。モネンシン区においては、 VFA 濃度の低下、アンモニア態窒素濃度の低下、メタン発生量の低下、プロトゾア数の低 下が見られている(高橋 2007)。BLCS ルナシータ投与区とはメタン発生量の低下のみが共 通し、他は異なる結果が得られている。DGGE の解析結果においては、モネンシン区で第 Ⅱ期および第Ⅳ期でSwine manure bacterium が増加し、BLCS ルナシータ投与区とは異 なるルーメンフローラを構築していることが明らかとなった。このため、BLCS ルナシー タによるメタン発生の低下はモネンシンによるメタン発生の低下とは異なるメカニズムに よるものであると考えられる。 2) 古細菌の解析 羊のルーメン内におけるメタン生成菌をクローンライブラリーにより測定した過去の研究 例では、メタン生成菌の約 96%が Methanobrevibacter 属であることが報告されている (Wright AD et al. 2004)。本試験においても出現した主な DGGE バンドの同定結果は Methanobrevibacter sp.で あ っ た こ と か ら 、 過 去 の 報 告 と 一 致 す る も の で あ っ た 。 Methanobrevibacter sp.は水素資化性メタン生成菌であることから、BLCS ルナシータを
投与した羊において、水素の授受に関与する真正細菌の増減が生じたことがルーメン内の メタン発生に密接に関連していたものと考えられる。 3) 真核生物の解析 DGGE 法による真核生物の解析対象として、元々ルーメン内に生息しているものとBLCS ルナシータに含まれる酵母が考えられる。しかし、本試験のDGGE 解析の結果、全くバン ドが認められなかった。その原因として次の二点が考えられる。その一つは検出感度の問 題 で あ る 。 ル ー メ ン 内 に は 約 104 個/ml の 真 核 生 物 が 存 在 し て い る と さ れ て お り(板 橋 2007)、PCR-DGGE の検出感度(103 個/ml)に近似した数値である。BLCS ルナシータに は酵母が含まれており、増殖すれば検出されたと考えられるが、検出されなかった結果か ら、酵母がルーメン内において活発に増殖しなかった可能性がある。 対照区と比較して BLCS ルナシータ 4g 投与区において繊維消化率(NDF、ヘミセルロー ス)がやや低下する傾向が見られた(高橋 2007)が、8g 区においては対照区と同程度に回復 していることが認められた(高橋 2007)。BLCS ルナシータ4g 区においては吸着剤の影響 により繊維消化率が低下したと思われる。酵母発酵培養物(Saccharomyces cerevisiae)の牛 ルーメン液への添加についてin vitro 試験を行った過去の報告では、VFA 生成とプロピオ ン酸生成が高まり、繊維分解菌数が増加して繊維消化率が約 10%高まることが報告されて いる(Lila et al. 2006)。さらに、BLCS ルナシータの投与によりVFA 生成とプロピオン酸 生成が高まる結果が得られている(高橋2007)。そこで、8g 区においても繊維消化率が回復 した理由として、BLCS ルナシータに含まれる酵母がルーメン内において機能していた可 能性が考えられる。 これらを明らかにするためには、ルーメン液に含まれる酵母を計数することが必要と考 えられる。また、BLCS ルナシータに用いる細菌の吸着剤として、繊維消化率に影響を与 えない物質を検討する必要があると思われる。 ⑤.試験2. T-RFLP 解析によるルーメンフローラの解析 ⑤-1 T-RFLP 法のフロー 図5-1 にT-RFLP 法によるルーメンフローラ解析のフローを示す 1. ルーメン液からのDNA 抽出・精製 ↓ 2. PCR 反応 ↓ 3. T-RFLP 解析 ↓ 4. T-RF データを利用した細菌の推定 図5-1 T-RFLP 法によるルーメンフローラ解析のフロー
1) ルーメン液からのDNA 抽出・精製 試験1 でルーメン液から抽出した DNA 溶液を用いた。また、Ⅱ期以降は前の期の試験の 影響を受けている可能性があるため、Ⅰ期の試料のみを用いた。 2) PCR 反応 試験1 で抽出したDNA 溶液(Ⅰ期のみ)を用いてPCR を行った。 試 験に用いたプライマーは
FAM-27f :AGAGTTTGATCCTGGCTCAG[5' 末端を6'-carboxyfluorescein(FAM)で標識] および1492r:GGCTACCTTGTTACGACTT である。 PCR 増幅反応条件を図⑤-2 に示す。 94℃ 2min ↓ 94℃ 30sec 55℃ 30sec 30cycles 68℃ 90sec ↓ 68℃ 7min 4℃ ∞ 図5-2 T-RFLP 解析のためのPCR 条件 3) T-RFLP 解析 PCR 増 幅 産 物 5μl を T-RFLP 解 析 に お い て 一 般 的 に 用 い ら れ て い る 制 限 酵 素 AluI(Nippon Gene)、HhaI(Nippon Gene)およびMspI(Nippon Gene)0.5μl を用いて37℃ で10 時間処理した後、95℃で10 分処理することで酵素を失活させた。失活処理液1μl、 サイズスタンダード(Gene Scan 500LIZ, Applied BioSystems 社)0.3μl、HiDi ホルムアミ ド(Applied BioSystems 社)12μl を混合し、95℃で2 分処理し、DNA を一本鎖にした。 得られた反応液をABI PRISM 310DNA シーケンサを用いてPOP-4 polymer ラン(変性系 電気泳動)を行い、制限酵素断片長(以下T-RF と略す)の蛍光強度をGene Scan により読み 込んだ。Gene Mapper ソフトウェア(Applied BioSystems 社)によりT-RF の蛍光強度の 定量解析を行った。
4) T-RF データを利用した細菌の推定
試験1 においてEMBL-FASTA 解析により同定した細菌の16S rDNA 配列をGene Bank から入手し、三種類の制限酵素の切断サイトを検索することにより、同定した細菌の理論 上のT-RF を求めた。理論上のT-RF の数値とT-RFLP 解析により得られたT-RF の数値
を比較することにより細菌を推定した。 ⑤-2 結果 表 5-4 にEMBL-FASTA により同定した細菌の16S rDNA を三種類の制限酵素処理した 時の理論上のT-RF を示した。DDBJ-FASTA と比較してEMBL-FASTA による検索にお いては相同性が同等もしくはそれ以上であるため、表5-4 においてはEMBL-FASTA の結 果を用いた。Ⅰ期の試料を用いたAluI 処理によるT-RFLP のチャートを図5-3 に示し、 HhaI処理およびMspI 処理による T-RFLP のチャートを図 5-4、図 5-5 にそれぞれ示し た。また、T-RF、ピークの高さ・面積を示した表について、AluI 処理の結果を表5-5 に、 HhaI 処理およびMspI 処理による結果について表5-6、表5-7 にそれぞれ示した。 AluI 処理の結果において、対照区と比較してBLCS ルナシータ投与区(4g 区および8g 区) でピーク高が増加した主なT-RF について述べる。なお、括弧内には表5-4 に示したT-RF と近似する細菌名を示す。増加した主なT-RF は、43~45bp、57bp(Bacterium Km5、8g 区のみ)、68bp、82bp、87bp(Clostridium leptum、8g 区のみ)、109bp、110bp、134bp、 282bp(8g 区のみ)、283bp(8g 区のみ)および436bp(Quinell ovalis、8g 区のみ)であった(図 5-3 に赤矢印で示す)。一方、対照区と比較して4g 区および8g 区でピーク高が低下した主 な T-RF は 70bp の み で あ っ た(図 5-3 に 青 矢 印 で 示 す)。 ま た 、BLCS ル ナ シ ー タ の T-RFLP結果で得られたピークの中で、4g 区または8g 区においても同じT-RF のサイズ で出現したピークは認められなかった。 HhaI 処理の結果において、対照区と比較してBLCS 投与区(4g 区および8g 区)でピー ク高が増加した主なT-RF は67bp(Bacterium Km5、8g 区のみ)、110bp、126bp、127bp、 189bp、209bp(8g 区のみ)、241bp(8g 区のみ)、383bp(Rumen bacterium、8g 区のみ)お よび560bp(Clostridium thermosuccinogenes、8g 区のみ)であった(図 5-4 に赤矢印で示 す)。一方、対照区と比較してBLCS投与区でピーク高が低下した主なT-RFは571bpであ った(図5-4 に青矢印で示す)。 MspI 処理の結果については、対照区と比較してBLCS 投与区(4g 区および8g 区)でピー ク高が増加した主なT-RF は77bp、89bp、307bp(8g 区のみ)、465bp(Bacterium Km5、 8g区のみ)であった(図5-5 に赤矢印で示す)。一方、対照区と比較してBLCS 投与4g およ び8g 区でピーク高が低下した主なT-RF は52bp、58bp、70bp、95bp、156bp、168bp、 169bp、170bp、237bp(Clostridium leptum)、268bp、461bp、481bp、493bp および495bp であった(図5-5 に青矢印で示す)。
⑤-3 T-RFLP 解析によるルーメンフローラの解析に関する考察
各制限酵素処理において BLCS ルナシータと BLCS ルナシータ投与区で出現したピーク の中で同一と思われるT-RF は認められず、投与したBLCS ルナシータに含まれていた細 菌がルーメン内で活発に増殖した可能性は低いと考えられ、DGGE 解析の結果を裏付ける も の と な っ た 。 ま た 、BLCS ル ナ シ ー タ 投 与 8g 区 に お い て は 対 照 区 と 比 較 し て 、 Clostridium leptum、Clostridium thermosuccinogenes と予測されるピーク高が増加する ことが認められ、DGGE の蛍光輝度の結果と一致するものであった。 ⑥.試験3. MPN-PCR 法を用いたメタン生成菌の定量 ⑥-1 MPN-PCR 法によるメタン生成菌の定量のフロー 図6-1 にMPN-PCR 法によるメタン生成菌の定量のフローを示した。 1. PCR 反応に関する予備検討 ↓ 2. MPN-PCR 法 ↓ 3. 最確数表によるメタン生成菌の定量 図6-1 MPN-PCR 法によるメタン生成菌の定量 ⑥-2 PCR 反応に関する予備検討 ル ー メ ン 内 に 存 在 す る 全 て の メ タ ン 生 成 菌 を 定 量 す る こ と を 目 的 と し て 、 Methanobacteriaceae 目 、 Methanococcales 目 、 Methanosarcinales 目 、 Methanomicrobiales 目および Methanosaeta 属に特異的なプライマーを用いて PCR を 行った。表⑥-1 にメタン生成菌を定量するために用いたプライマーの配列を示す。なお、 フォワードプライマー(109F)は古細菌に共通した配列であり、リバースプライマーは各メ タン生成菌に特異的な配列とした。図6-2 にMethanobacteriaceae 目を増幅させるPCR 条件を示した。各メタン生成菌をPCR で増幅するための最適条件を明らかにするための予 備試験を行い、Methanococcales 目および Methanosarcinales 目はアニーリング温度を 58℃ に 設 定 し 、Methanomicrobiales 目 およ び Methanosaeta 属 は ア ニ ーリ ン グ 温 度 を 56℃に設定した。なお、アニーリング温度以外のPCR 増幅条件はMethanobacteriaceae 目 と同一である。 PCR 反応を行うための鋳型として用いた DNA 溶液は 1) 試験 1 で用いたもの(0.5ml か ら抽出)、2) Ultra clean soil DNA kit(MoBio 社製)を用いて試料0.5ml から抽出を行った もの、3) 試験 1 では 0.5ml の試料から DNA 抽出を行ったが、高感度で検出を試みるた めに 4ml の試料から DNA 抽出を行ったもの、とした。また、PCR 阻害物質の除去を目 的として、Power clean DNA clean-up kit(MoBio 社製)およびBSA(5μg/μl)の添加(0.8μ l/PCR反応液40μl)について検討を行った。
表6-1メタン生成菌の定量に用いたプライマー 94℃ 4min 94℃ 45sec 50℃ 1min 30cycles 72℃ 1min 72℃ 7min 4℃ ∞ 図6-2 Methanobacteriaceae 目を増幅させるPCR 条件 ⑥-3 MPN-PCR 予備検討の結果を受けて DNA 抽出液の 10 倍希釈液、100 倍希釈液および 1000 倍希釈 液を用いて各希釈液につき5 反復のPCR を行い、PCR 増幅産物(40μl)をエタノール沈殿 により10μl に濃縮し、全量を1%アガロースゲル電気泳動に供すことにより増幅産物の確 認を行った。また、10 倍希釈液でバンドが見られない試料については、DNA 抽出液の原 液およびエタノール沈殿による10 倍濃縮液を調製してPCR を行った。 ⑥-4 最確数表によるメタン生成菌の定量 MPN-PCR法で出現したバンド数に基づいて最確数表からメタン生成菌の定量を行った。
⑥-5 結果
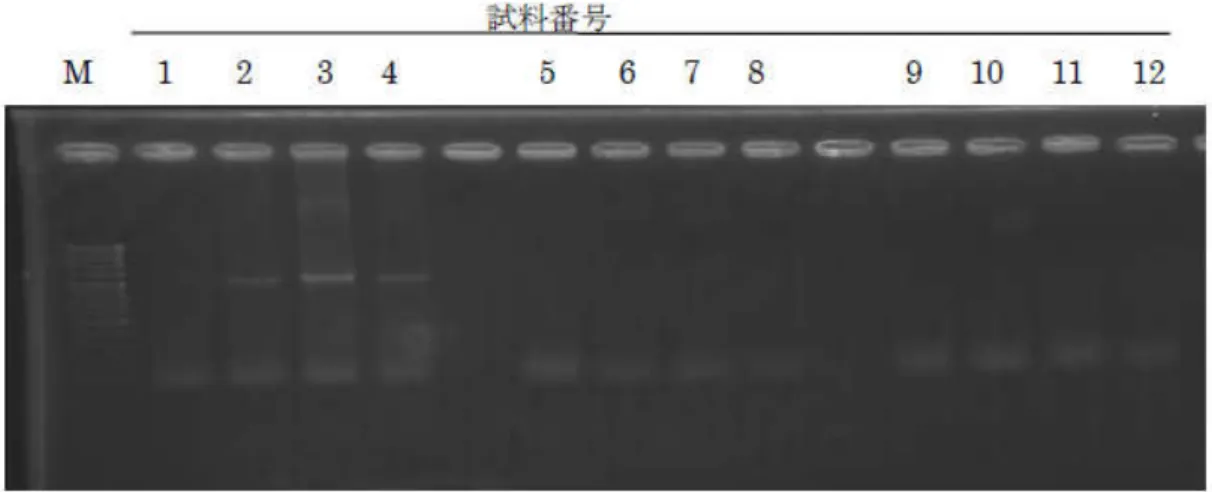
図 6-3 に Methanococcales 目、Methanomicrobiales 目、Methanosarcina 目、 Methanosaeta 属 お よ び Methanobacteriaceae 目 の 増 幅 結 果 を 示 す 。 そ の 結 果 、 Methanobacteriaceae 目のみで検出された。
図6-3 メタン生成菌の検出結果
図 6-4 に PCR 阻害物質除去による効果および試料濃縮による影響についての結果を示す (図中のM はマーカーを示す)。また、図6-4 の各レーンに流した試料の条件を表6-2 に示 す。DNA 抽出キットおよび試料量を4ml に増やしてDNA を抽出した方法では、BSA を 添加してもPCR 増幅産物は認められなかった。しかし、試験 1 で抽出したDNA を鋳型 としてBSA を添加してPCR を行うことにより、PCR 増幅産物が認められた。
図6-4 PCR 阻害物質除去による効果および試料濃縮による影響
表6-2 図⑥-4 に示した電気泳動画像における各レーンに流した試料の条件
表 6-3 にMPN-PCR 法で検出されたバンド数を示す。なお、10 倍濃縮液およびDNA 抽 出液の原液はPCR阻害物質が増加することや濃縮時にDNAロスが生じた可能性を考慮し、 参考データとした。
表6-3 MPN-PCR で検出されたバンド数
表 6-3 に示したバンド数から最確数表に基づいて細菌数を算出した結果を表6-4に示した。 Ⅰ期およびⅡ期においては対照区と比較して全ての試験区でメタン生成菌数の低下が認め られた。Ⅲ期においては4g 区においてメタン生成菌数が対照区と比較してやや多い結果と なった。Ⅳ期においては全ての試験区で検出限界以下となった。
表6-4 メタン生成菌数に及ぼすモネンシンおよびBLCS ルナシータの効果 ※ 1) DNA 抽出液を10 倍濃縮することにより、PCR 阻害物質が増加することや濃縮時に DNA ロスが生じる可能性を考慮し、10 倍希釈以降のデータを使用して定量値を算出した。 ⑥-6 MPN-PCR 法を用いたメタン生成菌の定量に関する考察 牛ルーメン内においてメタン生成菌は約 105~106 個/ml 存在することが報告されている が(小原 2006)、羊を用いた本試験においては約104~105 個/ml と数値が近似していた。 反芻動物である牛と羊のルーメン内には同程度の密度でメタン生成菌が生息していると考 えられることから、今回用いたDNA 抽出法およびPCR 条件はメタン生成菌の定量に適し たものであると考えられる。BLCS ルナシータ投与区においては、Ⅰ期およびⅡ期におい て対照区よりもメタン生成菌数の低下が見られており、BLCS ルナシータの効果によるも のである可能性が考えられた。BLCS ルナシータの 8g 区ではⅡ期以降メタン生成菌数は 検出限界以下となり、より強力にメタン生成菌数を低減させた可能性が示唆された。一方、 4g 区のⅢ期においては対照区よりもメタン生成菌数が多いという結果となった。BLCS ル ナシータ 4g 投与では効果が弱かったことやⅡ期試験の影響が残っていた可能性もあるか もしれないと考えられる。 Ⅳ期においては、全ての区においてメタン生成菌数は検出限界以下になっていた。同様 にⅢ期試験の影響が残っていたことも可能性の一つとしてあるのかもしれない。メタン生 成菌数の全ての試験区の平均値は第Ⅰ期より第Ⅱ期が少なく、第Ⅱ期よりも第Ⅲ期が少な く、第Ⅲ期よりも第Ⅳ期が少ない結果となっている。このため、試験間隔を決めるための 検討を行うことが今後の課題と考えられる。 ま た 、BLCS ル ナ シータ 8g 区 にお いて は、4-18. 1)-2 に 示 し た通 り、酪 酸 を産 生す る Clostridium leptum が増加することが明らかとなった。高橋らの実験(2007)ではBLCS ル ナシータ8g 区においてルーメン内の酪酸濃度が上昇し、それに対応するようにルーメン内 pH が低下している(高橋 2007)ことが示されている。メタン発生量はpH が0.1 低下する と10%程度低下することが示されているが(Russell JB 1998)、BLCS ルナシータ8g 区に おいて、ルーメン内 pH の経時的変化を測定した結果、対照区と比較してルーメン液採取 後4時間から8 時間ではpH が0.1 程度低下したことが示されている(高橋 2007)。ルー メン内 pH が低下することにより、メタン生成菌またはメタン生成菌が付着するプロトゾ アの数が8g 区において活動低下または死滅し、ルーメンからのメタン発生量が低下した可 能性が考えられる。すなわち、BLCS ルナシータの投与で、酪酸を産生する Clostridium
leptum が増加してpH が低下し、メタン生成菌が死滅してメタン発生が低下した可能性が 考えられる。 モネンシン区と4g 区においては、対照区と比較してメタン生成菌数の低下はわずかなも のであり、メタン発生量の低下(高橋 2007)と一致するものであると考えられる。一方、8g 区においては対照区と比較してメタン生成菌数は 1/10 以下に低下していることが示され たが、メ タン発生量は対照区と比 較し て 5~10%低減する に留まっていた(高橋 2007)。 BLCSルナシータに含まれる物質がPCR を阻害してBLCS ルナシータ8g 投与後のルー メン内のメタン生成菌数の検出に影響を与えていたのではないかと考えられる。 モネンシン区においてはプロトゾア数の低減が認められ(高橋 2007)、プロトゾアは繊維分 解活性を有することから繊維消化率の低下が予想されたが、中性デタージェント繊維(NDF)、 酸性デタージェント繊維(ADF)の消化率は対照区と同じ程度であった(高橋 2007)。これに 対してはプロトゾア数の低下が繊維消化率に及ぼす影響が小さかったことと、繊維分解菌 数が増加したことの2 つの可能性が考えられる。 ⑦まとめ BLCS ルナシータに含まれる細菌は投与区においてルーメン液から検出されず、ルーメン 内において中心的に機能している可能性は低いことが示された。しかし、他の細菌の活性 化を通して機能を発揮している可能性は残されている。BLCS ルナシータ投与区において は、Clostridium thermosuccinogenes、Clostridium leptum、Succiniclasticum ruminis が増加しており、いずれの細菌も水素を消費する代謝経路を有することから、メタン発生 の低減と関係している可能性があると考えられる。 BLCS 投与区においては多くの場合、対照区と比較してメタン生成菌数の低下が認められ た。 ⑧. 提言 ルーメンフローラの変化がBLCS ルナシータに含まれる細菌か吸着剤による影響であるの か 明 らか にす るた め 、BLCS ル ナシ ータ の吸 着剤 に 細菌 を添 加し て いない も の、 また は BLCS ルナシータを滅菌したものを対照区の羊に給与する必要があると考えられる。 BLCS ルナシータに含まれる酵母がルーメン内において機能しているか明らかにするため、 培養法により計数することや繊維分解菌をMPN-PCR 法等の手法を用いて定量することが 必要と考えられる。 本試験で生息が確認された Methanobrevibacter は培地中の増殖速度が真正細菌の数十 分の 1 程度と遅く、ルーメン内における増殖速度は明らかにされていないことから、試験 間隔を決めるためにBLCS ルナシータおよびモネンシン投与後のメタン生成菌数を経時的 に測定することが必要であると考えられる。また、ルーメンからのメタン発生は粗飼料多 給により低下することが報告されている(Russell JB 1998)ことから、給与飼料の違いがメ タン生成菌数に及ぼす影響を明らかにし、メタン生成菌数が高まる飼料を用いて試験を行
うことが必要であると思われる。BLCS ルナシータ 4g 区において繊維消化率が低下して いることから、BLCS ルナシータの吸着剤として繊維消化率に影響を与えない物質を検討 する必要も考えられる。
⑨.参考資料
Drent WJ, Lahpor GA, Wiegant WM, Gottschal JC (1991) Appl EnvironMicrobiol (1991) 57:455-462.
板橋久雄(2007) ルーメン研究会報18: 33-39.
Kato S, S Haruta, ZJ Cui, M Ishii, Y Igarashi(2005) Appl Environ Microbiol. 71:7099-7106.
Krulwich TA, Aqus R, Schneier M, Guffanti AA (1985) J Bacteriol. 162:768-772. Lila ZA, N Mohammed, T Takahashi, M Tabata, T Yasui, M Kurihara, S Kanda and H Itabashi (2006) Anim Sci J. 77:407-413.
小原(2006)ルミノロジーの基礎と応用. 農文協.
Roland N, Nugon-Baudon L, Andrieux C, Szylit O (1995) Br J Nutr. 74:239-249. Russell JB (1998) J Dairy Sci. 81:3222-3230.
Schnorpfeil M, Janausch IG, Biel S, Kroger A, Unden G (2001) Eur J Biochem. 268:3069-3074.
Sridhar J, Eiteman MA, Wiegel JW (2002) Appl Environ Microbiol. 66:246-251. 高浦一希ら (2006) 第 106 回 日本畜産学会大会
高橋潤一(2007) 生菌剤(BLCS ルナシータ)がめん羊のルーメンメタン生成、飼料消化率 及び窒素・エネルギー出納に及ぼす影響.
Van Gylswyk NO (1995) Int J Syst Bacteriol. 45:297-300.63
Wright AD, Williams AJ, Winter B, Christophersen CT, Rodgers SL, Smith KD (2004) Appl Environ Microbiol. 70:1263-1270.
2 2 2 2)))乳酸菌)乳酸菌乳酸菌の乳酸菌の産生のの産生産生産生するするするプロテアーゼするプロテアーゼ耐性抗菌性物質プロテアーゼプロテアーゼ耐性抗菌性物質耐性抗菌性物質耐性抗菌性物質(((PRA)( ))の)のの影響の影響影響影響 ①目的 味の素株式会社殿においては、ルーメンから発生する温室効果ガスであるメタンを抑制す る効果の見られる乳酸菌由来の抗菌物質の開発に取り組まれており、試験管レベルでメタ ンをほぼ完全に抑制する結果を得ている。抗菌物質をルーメン液に添加した際に結果とし て増加する細菌は、メタン生成菌の基質である水素の利用またはメタン生成菌に対する抗 菌効果を有している可能性が考えられる。このため、この様な機能を持つ細菌はメタン抑 制効果を有するプロバイオティクスとしての開発が期待できる。以上より本試験は、抗菌 物質をルーメン液に添加した時の細菌相の変化を明らかにすることを目的として行った。 ②試料 ルーメン液をホルスタイン種から採取し、緩衝液および基質を添加したもの(対照区)および 対照区に乳酸菌培養上清を10%添加したもの(添加区)を、それぞれ24 時間培養し、微生物 解析用試料とした。また、メタン抑制効果を有するモネンシン投与時の細菌相と比較する ため、チェビオット種去勢羊に給与飼料のみを投与したもの(コントロール区)および1 日当 たりモネンシン20mg を2 週間投与したもの(モネンシン区)から採取したルーメン液も微 生物解析用試料として用いた。
③方法 ③-1 PCR-変性グラジエントゲル電気泳動(DGGE)法 図3--1 にPCR-DGGE 法による微生物群集構造解析のフローを示す。 1. 試料からのDNA 抽出 ↓ 2. PCR 反応 1)真正細菌を対象としたPCR 反応(1 回目) 2)真正細菌PCR 反応(GC クランプ付き) ↓ 3. アガロース電気泳動による16S rDNA の増幅の確認 ↓ 4. DGGE 解析 ↓ 5. DNA バンド切り出しとDNA 抽出・精製 ↓ 6. PCR 反応(プライマー:GC クランプの除去) ↓ 7. PCR 産物の精製 ↓ 8. シーケンス解析 ↓ 10.ホモロジー解析(微生物種の同定) 図3-1 PCR-DGGE 法による微生物群集構造解析のフロー ③-2 試料からのDNA 抽出 ルーメン液にはDNA 抽出が困難な細菌およびPCR 阻害物質が存在すると考えられる。 このため、本試験では強固な細胞壁を備えた細菌の物理的破壊工程、腐植物質(あるいは前 駆物質であるリグニン)の除去工程を含む方法(Yoshida N et al. 2005)を用いた。 DNA バンドが単一にな るまで数回繰り返す
③-3 PCR 反応 表 3--1 にプライマーの配列を示した。 表 3--1 真正細菌を対象としたPCR に用いたプライマーの配列 ③-4 真正細菌を対象としたPCR 反応(1 回目) 抽出したDNA を鋳型として、真正細菌用のプライマー(341F、907R)を用いて、16SrDNA を増幅した。PCR 条件を図3-2 に示した。 〔PCR 反応(Primer : 341F, 907R) 〕 96℃ 5min 96℃ 1min 55℃ 30sec 25cycle 72℃ 1min 72℃ 5min 4℃ ∞ 図3-2 真正細菌の16SDNA を標的としたPCR 反応条件 ③-5 真正細菌を対象としたPCR 反応(GC クランプ付き) DGGE ゲルを用いた電気泳動を行うため、GC クランプ付(GC に富む配列)プライマー を用いたPCRを行った。具体的には図3-2 で得られたPCR産物を鋳型として、真正細菌 用GC クランプ付きプライマー(341FGC,534R)を用いてPCR を行い(図3-3)、アガロ ースゲル電気泳動にてPCR 産物の増幅を確認した。
〔PCR 反応(Primer : 341FGC, 534R) 〕 96℃ 5min 96℃ 1min 55℃ 30sec 25cycle 72℃ 30sec 72℃ 5min 4℃ ∞ 図3-3 真正細菌の16SDNA を標的としたPCR 反応条件(GC クランプ付) 3-6 DGGE 解析 広範囲のバンドが得られるように、ゲル中の DNA 変性剤の濃度勾配を 30-70%(変性剤 100%とは、7M 尿素、40%ホルムアミドに相当)としてDGGE 解析を行った。また、重 合剤の濃度は、低濃度に設定し、ゲルを 4 時間固化させることにより、グラジエントの精 度を高めたゲルを作成した。さらに、バンドの分離能を高めるため、電圧を低くし、泳動 時間を長く設定した(表3-2)。 表3-2 DGGE ゲル濃度と泳動条件 ポリアクリルアミドゲル濃度 :10% 変性剤濃度勾配 :30-70% 重合剤濃度 10% Ammonium Persulfate :0.7% TEMED :0.07% 泳動電圧 :70V 泳動時間 :960min 泳動温度 :60℃ ③-7 DNA バンドの切り出しとDNA 抽出・精製 電気泳動後、エチジウムブロマイドで染色を行い、脱染色後、UV トランスイルミネータ ー上でバンドを検出した。検出したバンドを滅菌されたメスで切り出し、1.5ml チューブ に入れた。切り出したゲルに、100μl のTE buffer(pH8.0)を加え、室温で一晩振とうさせ た後、エタノール沈殿法によりDNA の抽出・精製を行った。なお、分離能が低く、〔PCR 複数のバンドが混在している可能性のあるバンドについては、バンドが単一になるまでGC
クランプ付PCR およびDGGE 解析を繰り返した。 ③-8 PCR 反応(GC クランプの除去) 各バンドから抽出した DNA を鋳型として、シーケンス反応を行うため、GC クランプを 外すPCR を行った。 1) GC クランプ無しPCR GC クランプを外すPCR の条件を図3-4 に示した。 〔PCR反応 (Primer : 341F, 534R)〕 96℃ 5min 96℃ 1min 55℃ 30sec 25cycle 72℃ 30sec 72℃ 5min 4℃ ∞ 図3-4 GC クランプの除去に用いたPCR 反応条件 ③-9 PCR 産物の精製 アガロースゲル電気泳動により、PCR 産物を確認し、次いで、アガロースから目的バンド を切り出して、精製キット(MonoFas ; ジーエルサイエンス社製)により、PCR 産物を精 製した。 ③-10 シーケンス解析
3-9 で精製を行ったPCR 産物を鋳型として、ダイターミネーター法(Big Dye Terminator v1.1 Cycle Sequencing Kit;ABI 社製)によりシーケンシング反応を行った。
シーケンス解析は、ABI Prism310 Genetic Analyzer(ABI 社製)を用いて行った。
③-11 ホモロジー解析(微生物種の同定)
解読した各バンドのDNA シーケンスを基に、インターネットを利用したFASTA 検索によ りDDBJ のデータベースから微生物種の同定を行った。
④. 結果 DGGE バンドパターンを図4-1 に示した。また、塩基配列を決定したバンドを矢印で示し、 最も高い相同性を示した細菌名を示した。なお、出現した全ての細菌はルーメンからの分 離例が報告されている。 con: コントロール区、mo: モネンシン区 (チェビオット種去勢羊を用いた試験) 対照: 対照区、添加: 添加区 (ホルスタイン種を用いた試験) 図4-1 DGGE 解析結果(白黒反転画像)
④-1 シーケンス解析結果
シーケンス解析の結果決定した塩基配列を表4-2 に示した。
④-2 ホモロジー解析結果(微生物種の同定)
各バンドの塩基配列に基づいて DDBJ-FASTA 検索を行い、ホモロジーの高かった上位 5 種を表4-3 に示した。また、それらのIdentity(相同性; %で表す)とOverlap した領域(nt; 塩基対数)を表記した。
④-3 同定した細菌の生態的特徴
表4-4 に同定した細菌の生態的特徴を示した。 表4-4 同定した細菌の生態的特徴
⑤考察 1) 抗菌物質添加時のルーメンフローラ解析におけるDGGE の有用性 PCR-DGGE 法による解析により、抗菌物質添加時に無添加と比較してルーメン内で増減 する細菌を捉えることができた。表5-1 に抗菌物質添加時に増加した細菌、減少した細菌、 変化しなかった細菌について、最も相同性の高い細菌名およびグラム陽性・陰性の区別を 記載した。 表 5-1 抗菌物質添加時に増加した細菌、減少した細菌、変化しなかった細菌およびグラム 陽性・陰性の区別 ※増加した細菌・・・無添加と比較してDGGE バンドの蛍光輝度の上昇またはバンドの出 現(目 視観察による) ※減少した細菌・・・無添加と比較してDGGE バンドの蛍光輝度の低下またはバンドの消 失(目 視観察による) ※変化しなかった細菌・・・無添加と比較して DGGE バンドの蛍光輝度が変化しない(目 視観察 による)
2)-1 抗菌物質添加により増加した細菌によるメタン抑制について 抗菌物質投与時に増加する細菌は、メタン生成菌の基質である水素の利用またはメタン生 成菌に対する抗菌効果等を有している可能性が考えられる。このため、表 5-1 に示した抗 菌物質添加により増加した細菌の中に、メタン抑制に関与しているものが存在する可能性 がある。表 5-1 に示した抗菌物質添加時に増加した細菌の中で、メタン抑制に関与してい る細菌が存在すると仮定した場合、Streptococcus sp.およびButyrivibrio sp.の2 種類が最 も関与している可能性が高いと推察される。ルーメン内におけるこれまでの分離例(Tajima K et al. 2001b)から推察すると、これらの細菌の属種はそれぞれStreptococcus bovis およ び Butyrivibrio fibrisolvens であると思われる。この仮定が正しい場合のメタン抑制メカ ニズムの仮説を以下に示す。 ①Streptococcus bovis によるメタン抑制メカニズム(仮説) Streptococcus bovis LRC0476、HC5 株はバクテリオシンを産生し、HC5 株の産生するバ クテリオシンであるbovicin HC5 の抗菌メカニズムは不明であるが、in vitro で混合ルー メン液からのメタン産生を約50%抑制することが報告されている(Lee et al. 2002)。 bovicin HC5 はグラム陽性菌に対して抗菌作用を有することが報告されている(Houlihan AJ et al. 2006)ため、 ルー メン内 の主なメ タン生成 菌であ るグラム 陽性のメ タン生 成菌 (Methanobrevibacter ruminantium)に対して 直接抗 菌効果を有 している可能 性が考え ら れる。しかし、DGGE の結果からは真正細菌のグラム陽性菌の減少は見られなかったこと から、メタン生成菌に対する抗菌効果ではなく、メタン生成に関与する酵素活性を抑制し ている可能性も考えられる。また、本菌がバクテリオシンを産生していない株であるため に、メタン抑制とは全く関係が無い可能性もある。 ②Butyrivibrio fibrisolvens によるメタン抑制メカニズム(仮説) 本菌が産生するバクテリオシンは、グラム陽性菌に対して抗菌効果を有している(Jennifer et al 2002) 。 ル ー メ ン 内 に 存 在 す る 主 な メ タ ン 生 成 菌 で あ る Methanobrevibacter ruminantium はグラム陽性菌であり、このメタン生成菌に対して抗菌効果を示した可能性 もある。しかし、Streptococcus bovis と同様にDGGE の結果からは真正細菌のグラム陽 性菌の減少は見られていないため、メタン生成に関与する酵素活性を低下させる可能性も 考えられる。一方、本菌は不飽和脂肪酸に対する水素添加能を有しており、メタン生成菌 の基質である水素を消費するが、水素添加のために消費される水素はルーメン内の水素の 2%以内である(Czerkawski. 1986)。このため、不飽和脂肪酸に対する水素添加は、水素の 競合によるメタン発生の抑制に対する影響は極めて少ないと考えられる。本菌もバクテリ オシンを産生しない株である可能性はあるものの、メタン抑制に関与している場合は、メ タン生成に関与する酵素活性の抑制やバクテリオシンによるメタン生成菌に対する抗菌効 果である可能性が高いと推察される。 2)-2 メタン抑制効果を目指したプロバイオティクスの開発とその留意点について
今後の検討により、Streptococcus bovis やButyrivibrio fibrisolvens がメタン抑制に関与 している可能性が高いと結論される場合、全ての株にメタン生成酵素の阻害能やバクテリ
オ シ ン 産 生 能 が あ る わ け で は な い と 考 え ら れ る た め 、 細 菌 の 選 抜 時 に 注 意 を 要 す る 。 Streptococcus bovis の場合、多量に乳酸を産生することから、ルーメン内pH の低下が生 じ、病的状態であるルーメンアシドーシスを引き起こす危険性があることが指摘されてい る(Russell and Hino. 1985)。このため、抗菌物質の投与またはStreptococcus bovis をプ ロバイオティクスとして使用する場合には投与量に注意を払う必要があると考えられる。 また、ルーメン内乳酸の74%が Megasphaera elsdenii により利用されるという報告があ り(Counotte et al. 1981)、Megasphaera elsdenii をルーメン液に添加したin vitro の試験 では、乳酸蓄積量の低下とpH 低下の防止が見られている(Kung and Hession. 1995)。こ のため、Streptococcus bovis をプロバイオティクスとして使用する場合は、Megasphaera elsdenii 等の乳酸蓄積量を低下させる細菌との併用投与も検討する必要があると思われる。 3)-1 抗菌物質の抗菌作用について 表 5-1 に示した通り、抗菌物質添加により増加した細菌は全てグラム陽性菌であり、抗菌 物質添加により減少した細菌は全てグラム陰性菌であった。この結果から、抗菌物質はグ ラム陰性菌に特異的に抗菌効果を有している可能性が高いと考えられる。グラム陰性菌に 対する有効な抗生物質としてテトラサイクリン系があり、この抗生物質の抗菌効果はタン パク質合成を阻害することによるものである。また、テトラサイクリン系抗生物質は本試 験の結果でも減少の見られたPrevotella 属に対する抗菌作用を有しており、一つの可能性 として、本試験で用いた抗菌物質はタンパク質合成を阻害していることが考えられる。一 方、抗菌物質添加時に対照区と比較してバンドの蛍光輝度に変化の見られなかったバンド No.7 のBacterium L4M2 1-7 はグラム染色の結果は不明である。しかし、バンドNo.7 の ホモロジー解析の結果では、ホモロジーの高かった上位 5 種の中で本菌を除く全てがグラ ム陰性菌であった。このため、Bacterium L4M2 1-7 はグラム陰性菌である可能性が高い と推察されるが、本菌は抗菌物質による影響をほとんど受けていないことから、抗菌効果 がグラム陰性菌に対するものでは無い可能性もあると思われる。 3)-2 抗菌物質添加によるルーメン内揮発性脂肪酸(VFA)の変化について 味の素㈱殿の試験結果から、抗菌物質をルーメン液に添加して培養するとプロピオン酸濃 度が低下する場合が多いとの結果が得られている(味の素㈱. 2007 年. 未発表)。プロピオン 酸産生に関与しているPrevotella 属細菌や優占種ではないもののプロピオン酸を産生する グラム陰性菌が抗菌物質添加による影響で減少することにより、プロピオン酸濃度が低下 する可能性が考えられる。 3)-3 抗菌物質とモネンシンによる抗菌作用の比較 モネンシンはメタン抑制効果を有していることから、抗菌物質添加時に増減する細菌はモ ネンシン投与時に増減する細菌と類似している可能性を考えていたが、DGGE の結果から は、同じ挙動を示すバンドは見られなかった。逆に、抗菌物質添加で増加の見られたバン ドNo.3(Prevotella sp.)はモネンシン投与時には減少が見られた。また、抗菌物質添加で減 少 の 見 ら れ た バ ン ド No.8(Succinivibrio dextrinosolvens)お よ び No.12(Schwartzia
succinivorans)はモネンシン投与時には増加が見られた。この様な結果が得られた理由とし て、モネンシンが主にグラム陽性菌に対して抗菌効果を有しているのに対して、本試験の 対象とした抗菌物質は、主にグラム陰性菌に対して抗菌効果を及ぼしたことが考えられる。 3)-4 抗菌物質がメタン生成菌に及ぼす影響についての仮説 本試験は真正細菌の解析のみを行っており、メタン生成菌を含む古細菌の解析を実施して いないため、正確性に欠けると思われるが、メタン生成菌に対する抗菌物質の作用として 以下の仮説が考えられる。 ル ー メ ン 内 に は 主 要 な メ タ ン 生 成 菌 で あ る グ ラ ム 陽 性 の Methanobrevibacter ruminantium(Whitford et al. 2001)お よ び グ ラ ム 陰 性 の Methanomicrobium mobile (Tajima et al. 2001a)が存在することが知られている。抗菌物質添加により、メタン発生は ほぼ完全に抑制されている(味の素㈱2007. 未発表)ことから、抗菌物質は直接的・間接的に グラム陽性、陰性両方のメタン生成菌に対して抗菌効果を有している可能性が考えられる。 間接的なメタン生成菌に対する影響として、抗菌物質添加により増加した細菌の中に、メ タン生成菌との基質(水素)の競合や、抗菌物質の産生およびメタン生成に関与する酵素の阻 害物質の産生に関与しているものが含まれる可能性が考えられる。 4) まとめ 表5-2 に考察のまとめを示す。 表 5-2 考察まとめ ・抗菌物質は真正細菌のグラム陰性菌に対して抗菌効果を有している可能性が高い。 ・抗菌物質によるメタン抑制のメカニズムはモネンシンとは異なると推察される。 ・抗菌物質による抗菌効果だけでなく、抗菌物質添加により増加した Streptococcus 属ま たはButyrivibrio 属が産生するバクテリオシンがグラム陽性のメタン生成菌に対して抗菌 効果を有している可能性もある。 ・Streptococcus bovis は投与によりルーメンアシドーシスを引き起こす危険性があり、プ ロバイオティクスとして使用する場合には投与方法に注意を払う必要があると考えられ る。 ・グラム陰性菌の中で主にPrevotella 属の減少によりプロピオン酸の低下が生じている可 能性がある。 ⑥提言 本試験の結果、抗菌物質の添加によるルーメンフローラの変化を捉えることができたため、 今後は PCR-DGGE 法を用いて様々な条件下での抗菌物質添加時のルーメンフローラを解 析していくことが考えられる。
⑦引用文献
Counotte GH, RA Prins. Vet Res Commun. 5:101-115. 1981.
Czerkawski JW. Milligan LP, WL Grovum A Dobson (Eds.) pp.158-172. PrenticeHall, Englewood Cliffs, New Jersey.
Jennifer L. Rychlik and James B. Russell. Appl Environ Microbiol. 2002 March; 68(3): 1040–1046.
Houlihan AJ Russell JB. J Appl Microbiol. 100:168-174. 2006 Kung L Jr, AO Hession. J Anim Sci. 76:2190-2196. 1998.
Lee SS, JT Hsu, HC Mantovani, JB Russell. FEMS Microbiol Lett. 217:51-55. 2002. Russell JB, T Hino. J Dairy Sci. 67:1525-1531. 1985.
Tajima K, T Nagamine, H Matsui, M Nakamura, RI Aminov. FEMS Microbiol Lett. 200:67-72. 2001a.
Tajima K, Aminov RI, Nagamine T, Matsui H, Nakamura M, Benno Y. Appl Environ Microbiol. 67: 2766-2774. 2001b.
Whitford MF, RM Teather, RJ Forster, BMC Microbiol. 1:5. 2001.