関節リウマチ患者マスト細胞は
変形性関節症患者マスト細胞に比較して 有意に prostaglandin D 2 を産生する
日本大学大学院医学研究科博士課程 外科系整形外科学専攻
三嶋 信太郎 修了年 2017 年
指導教員 德橋 泰明
関節リウマチ患者マスト細胞は
変形性関節症患者マスト細胞に比較して 有意に prostaglandin D 2 を産生する
日本大学大学院医学研究科博士課程 外科系整形外科学専攻
三嶋 信太郎 修了年 2017 年
指導教員 德橋 泰明
目次
略語 ・・・・ 1 概要 ・・・・ 4 緒言
1. 関節リウマチ ・・・・ 6 2. 関節リウマチとマスト細胞 ・・・・ 6 3. 関節リウマチと prostaglandin ・・・・ 8 4. 関節リ ウマチ 患者の 関節液 中の prostaglandin ・・・ ・ 9 5. microRNA と 関 節 リ ウ マ チ ・ ・ ・ ・ 9
目的 ・・・・ 11 対象と方法
1. 使用抗体、試薬、物品 ・・・・ 12 2. ヒト滑膜マスト細胞の分離と培養 ・・・・ 12 3. 線維芽細胞の分離と培養 ・・・・ 13 4. OA マスト細胞と RA 線維芽細胞との共培養 ・・・・ 13 5. 滑膜マスト細胞の活性化 ・・・・ 13 6. total RNA 抽出、相補的 DNA (cDNA) への逆転写、定量的 RT-
PCR ・・・・ 14
7. DNA chip 解析 ・・・・ 14
8. microRNA (miRNA) の網羅的発現解析 ・・・・ 15
9. メディエーターの測定 ・・・・ 15
10. 関節液の採取 ・・・・ 16
11. 統計処理 ・・・・ 16
結果
1. DNA chip 解析による OA 、 RA マスト細胞における遺伝子発現
量の比較 ・・・・ 17
2. 定量的 RT-PCR 解析による OA 、 RA マスト細胞における脂質メ ディエーター合成酵素の発現の比較 ・・・・ 17
3. OA 、 RA マスト細胞における IgE 依存性脱顆粒 反応および脂 質代謝物産生量の比較 ・・・・ 18
4. OA 、 RA マスト細胞における IgG 依存性脱顆粒反応および脂質 代謝物産生量の比較 ・・・・ 18
5. OA 、 RA 線維芽細胞における PTGS1 、 PTGS2 、 TBXAS1 および LTC4S mRNA 発現量の比較と OA マスト細胞と RA 共培養によ る遺伝子発現量への影響 ・・・・ 18
6. O A 、 R A マ ス ト 細 胞 の m i R - 1 9 9 a - 3 p 発 現 量 の 比 較 ・ ・ ・ ・ 1 9 7. OA 、 RA マスト細胞における miR-199a-3p と PTGS2 の発現の 相関 ・・・・ 19
8. O A 、 R A 患 者 の 関 節 液 中 の P G D 2 と P G E 2 量 の 比 較 ・・・・ 20
考察 ・・・・ 21
まとめ ・・・・ 28
謝辞 ・・・・ 29
表
1. マ ス ト 細 胞 欠 損 マ ウ ス ご と の マ ス ト 細 胞 の 関 節 炎 へ の 関 与 ・・・・ 30 2. 関節炎モデルマウスと PGD 2 ・・・・ 31 3. OA マスト細胞において RA マスト細胞と比較して 3 倍以上発
現量が高い microRNA の発現網羅的解析結果 ・・・・ 32 4. OA 、 RA 患者の検査結果、治療情報 ・・・・ 33
図
1. アラキドン酸カスケード ・・・・ 35
2. DNA chip 解析を用いた OA 、 RA マスト細胞における遺伝子発
現量の比較 ( クラスター解析 ) ・・・・ 36
3. DNA chip 解析を用いた OA 、 RA マスト細胞における脂質メデ
ィエーター関連遺伝子発現量の比較 ・・・・ 37
4. 定量的 RT-PCR 解析による OA 、 RA マスト細胞における脂質メ
ディエーター合成酵素の発現の比較 ・・・・ 38 5. OA 、 RA マスト細胞における IgE 依存性脱顆粒反応および脂質
代謝物産生量の比較 ・・・・ 39 6. OA 、 RA マスト細胞における IgG 依存性脱顆粒反応および脂質
代謝物産生量の比較 ・・・・ 40
7. OA 、 RA 線維芽細胞における PTGS1 、 PTGS2 、 TBXAS1 、 LTC4S
mRNA 発現量の比較と OA マスト細胞と RA 線維芽細胞の共培
養による PTGS1 、 PTGS2 、 TBXAS1 、 LTC4S mRNA 発現量への
影響 ・・・・ 41
8. O A 、 R A マ ス ト 細 胞 の m i R - 1 9 9 a - 3 p 発 現 量 の 比 較
・・・・ 42 9. OA 、 RA マスト細胞における miR-199a-3p の発現量と PTGS2
の発現の相関 ・・・・ 43 10. OA 、 RA 患者の関節液中の PGD 2 と PGE 2 量 ・・・・ 44
図説
1. アラキドン酸カスケード ・・・・ 44 2. DNA chip 解 析 を 用 い た OA 、 RA マ ス ト 細 胞 に お け る 遺
伝子発現量の比較 ( クラスター解析 ) ・・・・ 45 3. DNA chip 解 析 を 用 い た OA 、 RA マ ス ト 細 胞 に お け る 脂
質メディエーター関連遺伝子発現量の比較 ・・・・ 45 4. 定 量 的 RT-PCR 解 析 に よ る OA 、 RA マ ス ト 細 胞 に お け
る脂質メディエーター合成酵素の発現の比較 ・・・・ 46 5. O A 、 R A マ ス ト 細 胞 に お け る I g E 依 存 性 脱 顆 粒 反 応 お
よび脂質代謝物産生量の比較 ・・・・ 46 6. O A 、 R A マ ス ト 細 胞 に お け る I g G 依 存 性 脱 顆 粒 反 応 お
よび脂質代謝物産生量の比較 ・・・・ 46 7. OA 、 RA 線維芽細胞における PTGS1 、 PTGS2 、 TBXAS1 、 LTC4S mRNA 発現量の比較と OA マスト細胞と RA 線維芽細胞の共培養 による PTGS1 、 PTGS2 、 TBXAS1 、 LTC4S mRNA 発現量への影 響 ・・・・ 47 8. O A 、 R A マ ス ト 細 胞 の m i R - 1 9 9 a - 3 p 発 現 量 の 比 較
・・・・ 47
9. OA 、 RA マスト細胞のおける miR-199a-3p と PTGS2 の発現の相 関 ・・・・ 47 10. OA 、 RA 患 者 の 関 節 液 中 の PGD 2 、 PGE 2 量 の 比
較 ・・・・ 48
引用文献 ・・・・ 49
研究業績 ・・・・ 55
1
略語ALOX5: arachidonate 5-lipoxygenase
anti-CCP Ab: anti-cyclic citrullinated peptide antibody anti-TNF-α: anti tumor necrosis factor-α antibody CMA1: chymase 1
COX: cyclooxygenase CRP: C-reactive protein
CRTH2: chemoattractant receptor-homologous molecule on Th2 cells CYSLTR1: cysteinyl leukotriene receptor 1
DMARD: disease modifying antirheumatic drug DiHETE: dihydroxyeicosatrienoic acid
EETs: eicosatrienoic acids EIA: enzyme immunoassay
GPI: glucose-6-phosphate isomerase
H-PGDS: hematopoietic type-prostaglandin D synthase HDC: histidine decarboxylase
HETE: hydroxy eicosatetraenoic acid
HHT: hydroxy heptadecatrenoic acid
2 HPETE: hydroperoxyeicosatetraenoic acid IL: interleukin
IMDM: Iscove’s modified Dulbecco’s medium L-PGDS: lipocalin type-prostaglandin D synthase LSM: lymphocyte separation medium
LT: leukotriene
LTA4H: leukotriene A4 hydrolase LTC4S: leukotriene C4 synthase LX: lipoxin
MMP3: matrix metalloproteinase 3 MOX: methoxylamine
MS4A2: membrane-spanning 4A gene 2 MTX: methotrexate
n.d: not done
NSAIDs: non-steroidal anti-inflammatory drugs OA: osteoarthritis
PE: phycoerythrin PG: prostaglandin
PGDS: prostaglandin D synthase
PLA
2: phospholipase A
23 PSL: prednisolone
PTGDR: prostaglandin D receptor PTGER: prostaglandin E receptor PTGES: prostaglandin E synthase PTGIS: prostaglandin I synthase PTGS: prostaglandin synthase RA: rheumatoid arthritis
RANKL: receptor activation of NF-κB ligand RF: rheumatoid factor
TBXAS1: thromboxane A synthase 1 Th: T helper
TNF: tumor necrosis factor
TPSB2: tryptase beta 2
Tx: thromboxane
WBC: white blood cells
4
概要
背景:関節リウマチ
(rheumatoid arthritis; RA)
は、複数の遺伝的要因に環境因子が加 わり自己免疫応答が惹起され、これらの結果として慢性炎症性病態が複数の関節に 対称性に生じ、進行性の破壊性関節炎に至る疾患と考えられている。RA
患者の血清 中には抗シトルリン化蛋白抗体などの自己抗体が検出され、これらが病態発症との 関連を示す全身性の自己免疫疾患である。RA
の病態においては関節炎マウスモデル や滑膜の組織染色を用いた研究からマスト細胞はRA
の発症や炎症の増悪に関与す るいわゆる“
悪玉”
として考えられてきた。しかし、ここ1-2
年のRA
患者の関節滑膜 組織中のマスト細胞関連遺伝子発現や関節液中のマスト細胞由来のメディエーター がRA
の重症度や炎症マーカーと逆相関することが報告され、“
善玉”
の可能性が示 唆されていたが、その機序は不明であった。TNF- α阻害薬、 IL-6
受容体阻害薬など の生物学的製剤を使用して関節破壊を抑制し、寛解状態に持ち込める確率が向上し たが、生物学的製剤は、高価であり、生物学的製剤に反応しない患者も存在し、よ り効果的で安価な治療法が望まれる。目的:
RA
患者由来培養マスト細胞(
以下RA
マスト細胞と呼ぶ)
がどのようにし て、RA
の炎症を抑制するかその機序を明らかにし、新規治療薬の開発に資する研究 を行うことを目的とした。方法:関節滑膜は
RA
患者およびOA
患者に施行された人工膝関節置換術の際に採 取された。各患者由来の滑膜からマスト細胞を培養し樹立した。DNA chip
を用いてOA
およびRA
由来マスト細胞の遺伝子プロファイルを網羅的に調べた。OA
患者由 来マスト細胞(
以下OA
マスト細胞と呼ぶ)
とRA
マスト細胞における遺伝子発現量は定量的
RT-PCR
法で測定した。IgE
およびIgG
依存的な活性化に違いが見られるかを検証するために、
OA
およびRA
マスト細胞からのアラキドン酸代謝物の産生とヒ スタミン遊離量をenzyme-immunoassay (EIA)
法で測定した。関節滑膜からOA
患者5
由来線維芽細胞
(
以下OA
線維芽細胞と呼ぶ)
とRA
患者由来線維芽細胞(
以下RA
線維芽細胞と呼ぶ)
を培養し、遺伝子発現量を定量的RT-PCR
法で測定した。OA
マ スト細胞とRA
線維芽細胞を共培養することで変化するOA
マスト細胞の遺伝子発 現量を定量的RT-PCR
法で測定した。OA
マスト細胞とRA
マスト細胞の性質を決定 する因子として、microRNA (miRNA)
に着目し、両マスト細胞のmiRNA
のプロファ イルを網羅的に調べた。OA
マスト細胞とRA
マスト細胞のprostaglandin synthase 2 (PTGS2 = COX-2: cyclooxygenase-2)
の発現量とmiR-199a-3p
の発現量を定量的RT- PCR
法で比較し、相関性を調べた。患者の関節液中のPGD
2、PGE
2量はEIA
法で測 定した。統計学的解析は、Mann-Whitney
のU
検定によって行われた。結果:
DNA chip
の結果、prostaglandin synthase 1 (PTGS1 = COX-1: cyclooxygenase- 1)
、prostaglandin synthase 2 (PTGS2 = COX-2)
、thromboxane synthase 1 (TBXAS
1)
、leukotriene C4 synthase (LTC4S) mRNA
の発現量はOA
マスト細胞と比較してRA
マス ト細胞の方が有意に高かった。また、IgE
依存性刺激において、PGD
2産生量はRA
マスト細胞のほうが有意に高かった。一方LTB
4産生量はOA
マスト細胞で高い傾向 にあった。IgG
依存性刺激においてPGD
2産生量はRA
マスト細胞のほうが有意に高 かった。したがって、OA
およびRA
マスト細胞は、異なった性質を有していること が明らかになった。RA
線維芽細胞においてPTGS1
、PTGS2
、TBXAS1
、LTC4S mRNA
の発現量はOA
線維芽細胞と同程度であった。OA
マスト細胞をRA
線維芽細 胞と共培養してもPTGS1
、PTGS2
、TBXAS1
、LTC4S mRNA
の発現量に変化は見ら れなかったことから、両マスト細胞の性質の違いは、線維芽細胞に起因しないこと が示唆された。次にmiRNA chip
の結果、OA
マスト細胞の方が、RA
マスト細胞よ り3
倍以上発現量が高いmiRNA
を20
個見出した。これら20
個のmiRNA
のうちPTGS2
の発現制御に寄与するmiRNA
はmiR199a-3p
であった。miR-199a-3p
とPTGS2
の発現量の相関を調べたところOA
マスト細胞では相関がなかったが、RA
マスト細胞では負の相関がみられた。関節液中の
PGD
2量は、RA
の方が有意に高か ったのに対し、PGE
2量は両群間に有意な差は見られなかった。結論:
PGD
2は各種炎症モデルで炎症の抑制効果を持つことが示唆されており、この 報告と本研究の結果から、RA
マスト細胞が免疫複合体の刺激によって過剰なPGD
26
を産生することにより、
RA
の炎症を抑制し ている可能性が示唆された。7
緒言
1.
関節リウマチ(rheumatoid arthritis; RA)
RA
は、複数の遺伝的要因に環境因子が加わり自己免疫応答が惹起され、これら の結果として慢性炎症性病態が複数の関節に対称性に生じ、進行性の破壊性関節炎 に至る疾患と考えられている(1, 2)
。RA
患者の血清中には抗シトルリン化蛋白抗体(Anti-citrullinated peptide antibodies; ACPAs)
などの自己抗体が検出され、これらが病 態発症との関連を示す全身性の自己免疫疾患である(1, 2)
。RA
の炎症の主病変は関 節滑膜である。RA
では滑膜の毛細血管周囲に抗原提示細胞とT
細胞の浸潤が起こ り、続いてB
細胞の浸潤がみられる。T
細胞のなかでもTh17
細胞が病態発症に中心 的な役割を担っていることが知られている(3)
。Th17
細胞は、Interleukin (IL) -17
を 産生し、滑膜線維芽細胞を活性化して炎症を惹起する。滑膜線維芽細胞は炎症が進 展するにつれ増殖し、多層化、絨毛状になる。滑膜の深部では炎症細胞の浸潤と血 管の新生が起こり、肉芽組織が形成されその中で骨破壊を担う破骨細胞が活性化される
(1, 2)
。血管新生、滑膜表層細胞の増生、炎症細胞の活性化、骨・軟骨の破壊などの病態は、種々の細胞から分泌される種々のサイトカインやケモカインが複合的 に働いた結果であると考えられている。前述した
IL-17
や、マクロファージ、滑膜 細胞の作るReceptor Activation of NF-κB Ligand (RANKL)
、腫瘍壊死因子(tumor necrosis factor α; TNF-α)
、IL-1
、IL-6
などの炎症性サイトカインが病態形成の主要因 と考えられている(1, 2, 4)
。1980
年代にRA
に対するメトトレキサート(methotrexate; MTX)
の有効性が確立され、アンカードラッグとしてRA
治療の中心となってきた。また
MTX
の反応が十分でない症例にはTNF-α阻害薬、 IL-6
受容体 阻害薬などの生物学的製剤を使用して関節破壊を抑制し、寛解状態に持ち込める確 率が向上した(4)
。しかし生物学的製剤は、高価であり、生物学的製剤に反応しない 患者も存在し、より効果的で安価な治療法は望まれる。2.
関節リウマチとマスト細胞(a)
関節リウマチとマウスマスト細胞8
K/BxN
マウス血清中には抗glucose-6-phosphate isomerase (GPI)
抗体が含まれてお り、K/BxN
マウス血清を正常マウスに移入すると、関節炎(K/BxN-passive arthritis [PA])
が生じる(5)
。K/BxN-PA
はFc
受容体のγサブユニットやFc γ RIII
を欠いたマウ スでは発症しない(6, 7)
。抗II
型コラーゲンモノクローナル抗体cocktail
を用いた関 節炎モデルと抗原誘発性関節炎モデルの研究において、IgG
受容体の活性化が関節 炎発症に中心的な役割を果たしていることが明らかとなっている(8-10)
。マスト細 胞欠損マウスであるW/W
vマウスとSl/Sl
dマウスに、自己抗体を移入した実験から、K/BxN-PA
にはマスト細胞が必要であり(11)
、さらに自己抗体誘導性マスト細胞活性化には
Fc γ R
が関与していることが報告されている(12)
。W/W
vマウスと同様に末 梢血の好中球が少なく、Kit
に変異を有するマスト細胞欠損マウスであるPretty2
マ ウスにおいても、K/BxN-PA
は減弱した(13)
。また、マウスマスト細胞プロテアー ゼ6
欠損マウスはK/BxN-PA
の発症に抵抗性であるとも報告されている(14)
。一 方、W
shマウスやCpa3
cre/+マウスなどのマスト細胞欠損マウスでは関節炎は減弱しな かったと報告されている(15, 16) (
表1)
。これらのことから、RA
の病態におけるマ スト細胞の役割についてはいまだに議論がある。最新の報告ではK/BxN-PA
を用い て時期特異的にマスト細胞を欠損させると、発症前にマスト細胞を欠損させた時の み炎症を抑制したという報告がある(17)
。(b)
関節リウマチとヒトマスト細胞RA
患者の滑膜組織では、健常膝関節滑膜組織と比較して(18)
、あるいは、半月 板切除術後の半月板組織と比較してマスト細胞数が増加し、マスト細胞数と臨床的 な滑膜炎症の程度との相関すること(19)
、脱顆粒像を示すマスト細胞が増加し、マ スト細胞周囲にTNF- αや IL-1
が存在すること(20)
、RA
患者の関節液中においてヒ スタミンが検出されること(21)
、またMC
TC タイプ(tryptase
、chymase
の両方のプ ロテアーゼを持つマスト細胞)
のマスト細胞数は、OA
と健常膝関節滑膜組織と比較 して増加していること(22)
が報告されている。RA
患者の滑膜マスト細胞はOA
患 者の滑膜マスト細胞と比べ抗IgE
抗体(23)
、アナフィラトキシンC5a (24)
に反応し て多量のヒスタミンを遊離することが報告されている。これらの結果からマスト細 胞がRA
の病態、特に、炎症の増悪に関与していることが示唆されていた。2012
年、Lee
らは免疫複合体が高親和性IgG
受容体であるFcγRI
、低親和性IgG
受容体9
である
Fc γ RIIA
を介して関節滑膜マスト細胞を活性化することを報告したが、RA
と 変形性関節症(osteoarthritis: OA)
の関節滑膜マスト細胞表面のFc γ RI
、Fc γ RII
、FcγRIII
および高親和性IgE
受容体であるFcεRI
の発現量に差はなく、FcγRI
やFcεRI
の架橋によって遊離されるヒスタミン量やサイトカイン量には差がなかった(25)
。 最近、RA
患者の関節滑膜組織においてstem cell factor
の受容体であるKit
やトリプ ターゼなどのマスト細胞特異的遺伝子発現と血清c-reactive-protein (CRP)
などのRA
の重症度には負の相関が見られることから、マスト細胞は、RA
の関節炎において抗 炎症作用を有している可能性が示唆されている(26, 27)
。したがってヒト滑膜マスト 細胞がRA
の関節炎の抑制に関与していることが示唆されその機序の解明が治療に 結び付くと考えられる。3.
関節リウマチとprostaglandin
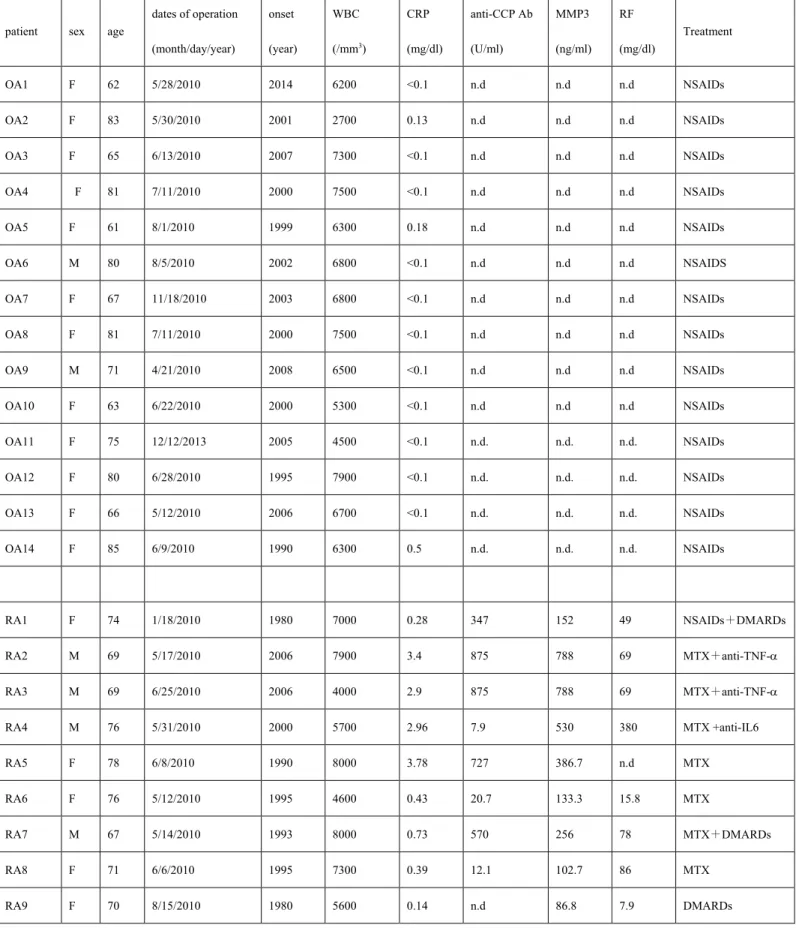
アラキドン酸カスケードはアラキドン酸を原料としてプロスタグランディン類
(prostaglandins: PGs)
やトロンボキサン類(thromboxanes; TXs)
やロイコトリエン類(leukotrienes: LTs)
などの脂質メディエーターを生合成する代謝経路である(
図1)
。 アラキドン酸は細胞膜のリン脂質に含まれ、活性化したphospholipase A
2(PLA
2)
に よって細胞内へ遊離される。その後、細胞膜にあるcyclooxygenase (COX)
によって 中間体を経て、PGD
2、PGE
2、PGF
2α、PGI
2およびTXA
2が合成される。また、アラ キドン酸はarachidonate 5-lipoxygenase (ALOX5)
によってLTA
2を介して最終的にLTB
4、LTC
4、LTD
4、LTE
4が生成される。prostaglandin
は、RA
の滑膜細胞で産生が亢進し、炎症惹起作用、血管透過性亢進、
T
細胞の遊走、マトリックスメタロプロテイナーゼの産生誘導など、炎症反応 の調整をする(28)
。さらに、滑膜の増殖や血管新生にも関与する最も重要なメディ エーターと考えられている(28)
。コラーゲンによって関節炎を誘導したDBA/1J
mice
に、PGD
2受容体のひとつであるDP1
のアンタゴニストを投与すると炎症反応 は増悪する一方で、PGD
2のもう一つの受容体であるDP2 (
別名chemoattractant
receptor-homologous molecule on Th2 cells; CRTH2)
のアンタゴニストを投与すると炎 症は不変であった(29)
。また、PGD
2を投与すると炎症は抑制され、DP1
アゴニスト を投与すると炎症は抑制された(29)
。これらの結果から、PGD
2はDP1
を介して炎 症を抑制する役割を担っていることが示された。コラーゲンによって関節炎を誘導10
した
DBA/1J mice
ではDP2
アンタゴニスト投与で炎症反応は不変であった(29)
。アジュバンド誘導性関節炎モデルマウス
(C57BL/6J mice)
を用い、炎症反応の変化を 調べた報告がある。その報告によると野生型マウスとCRTH2
ノックアウトマウスに 炎症反応を惹起させると、両方のマウスでPGD
2の産生が亢進し、炎症反応の増悪が 見られたがCRTH2
ノックアウトマウスの方が、より重度な炎症反応の増悪をみとめた
(30)
。いずれもPGD
2は関節炎に対し、抑制効果を有することが示唆された。以上のことを表
2
に示す。prostaglandin
の中でもPGE
2はRA
の炎症惹起に対して重要 な役割を担っている(31)
。PGE
2はRA
の滑膜組織において発熱、疼痛、血管透過性 亢進、炎症細胞遊走などを引き起こし、炎症を増悪させる。また、破骨細胞を活性 化する作用もあることが知られている(32)
。骨吸収促進による関節破壊はRA
の重 要な病態である。またRA
の滑膜組織ではPTGS2
が著明に発現している(33)
。これ らのことからRA
においてPTGS2
を抑制することが抗炎症をもたらす可能性が考え られる。4.
関節リウマチ患者の関節液中のprostaglandin
RA
患者の滑膜細胞ではPGE
2、PGF
2α、PGI
2、TXA
2などが産生され、関節液中で はPGD
2、PGE
2、PGF
2α、6-keto-PGF
1α、TXB
2、LTB
4などが検出される。RA
患者で はOA
患者と比較し、関節液中のPGE
2濃度が有意に高く、NSAIDs
を服用しているRA
患者ではステロイド服用患者よりもPGE
2濃度は低いということが報告されている
(34)
。関節内でのPGE
2産生細胞として関節液中マクロファージや滑膜細胞が挙げられる
(31)
。PGD
2の合成酵素にはL-PGDS (lipocalin type-prostaglandin D synthase)
とH-PGDS (hematopoietic type-prostaglandin D synthase)
があり、末梢組織ではH-PGDS
によって生合成される。H-PGDS
はマスト細胞、樹状細胞、Th2
リンパ球などの造血 系の細胞に発現している(35-37)
。関節液中で検出されるPGD
2はこれらの細胞から 産生されたものと考えられる。ヒト滑膜マスト細胞はPGD
2を産生することが報告されており
(38)
、RA
の滑膜マスト細胞が免疫複合体で刺激されPGD
2を産生し、局所の炎症を制御しているという仮説を立てた。
5. microRNA
と関節リウマチmicroRNA (
以下miRNA)
はゲノム上にコードされ、多段階的な生成過程を経て最11
終的に
20
から25
塩基長の微小RNA
となる機能性核酸である。特定のmRNA
に結 合し、その翻訳を阻害したり、不安定化して破壊に導いたりして当該分子の発現を 制御して免疫機能を調節している(39)
。非常に多くのmiRNA
が関節リウマチと関 係するものとして同定されており、バイオマーカーや、診断や予測、治療への応用 が期待されている(40)
。例えば、RA
のB
細胞ではmiRNA-155
が抗体産生を促進す る役割を果たし(41)
、RA
の線維芽細胞ではmiRNA-17
がTNF- αのシグナルの抑制
に関与している(42)
。本研究ではRA
とOA
マスト細胞から産生されるPGD
2の量の差は
miRNA
によって制御されているという仮説を立てた。12
目的
RA
患者由来培養マスト細胞がどのようにして、RA
の炎症を抑制するかその機序を 明らかにし、新規治療薬の開発に資する研究を行うことを目的とした。13
対象と方法
1.
使用抗体、試薬、物品以下の抗体、試薬、物品は、それぞれ下記の会社から購入した。
HistoDenz solution
、ウシ血清アルブミン(bovine serum albumin ; BSA)
はSigma-Aldrich (Tokyo, Japan)
、ヒトミエローマIgE
は、Calbiochem (San Diego, CA, USA)
、フィコエリスリ ン(phycoerythrin: PE)
、標識ヤギ抗マウスモノクローナル抗体は、BD Bioscience (San Jose, CA, USA)
、ウシ胎児血清、ペニシリン/
ストレプトマイシンは、GIBCO (CA, USA)
、リンパ球分離溶液(lymphocyte separation medium; LSM)
は、Organon Teknika (Durham, NC, USA)
、ヒトリコンビナントstem cell factor (SCF)
、ヒトリコン ビナントIL-6
は、PeproTech (Rocky Hill, NJ, USA)
、無血清Iscove’s methylcellulose medium
は、Stem Cell Technologies (Vancouver, BC, Canada)
、TaqMan
プライマーは全 て、Applied Biosystems (Tokyo, Japan)
、miRNeasy Mini kit
はQiagen (Hilden,
Germany)
、抗FcγRI F(ab')
2 フラグメント(F[ab’]
2αFcγRI, clone 10.1)
はID Labs (London, ON, Canada)
、マウスIgG1 F(ab')
2フラグメント(F[ab’]
2mIgG1)
および抗マ ウスIgG F(ab')
2フラグメント特異的ヤギF(ab')
2フラグメント(gF[ab’]
2αmF[ab’]
2)
はJackson ImmunoResearch (West Grove, PA, USA)
。2.
ヒト滑膜マスト細胞の分離と培養平成
22
年2
月8
日付けで関節滑膜組織の使用に際して、倫理委員会・臨床研究審 査委員会の承認番号RK-100115-4
を受けた。その後改訂版として平成23
年4
月12
日、平成24
年12
月17
日、平成25
年6
月25
日付けで追加承認を受けた。平成28
年2
月5
日付け承認番号RK-160112-2
を受けた。手術前にインフォームドコンセン トを患者とその家族に行い、承諾書を頂いた。その後、日本大学医学部附属板橋病 院にて行われた、人工膝関節置換術で切除された関節滑膜組織の一部を実験に供し た。滑膜組織を酵素的に処理し、細胞を単離した(
マスト細胞の純度は約5%)
。マス ト細胞の純度はキムラ染色(43)
を用いて算出した。マスト細胞の純度を上げるため に、単離された細胞を、2%
ウシ胎児血清と100 units/ml
ペニシリン/ 100µg/ml
ス トレプトマイシンを添加したIscove’s modified Dulbecco’s medium (IMDM; Invitrogen)
14
に再浮遊させた後、
22.5% HistoDenz solution
とリンパ球分離溶液(LSM: lymphocyte
separation medium)
を用いて比重遠心した。滑膜マスト細胞の前駆細胞と成熟マスト細胞は、その遠心により得られた沈殿層と
LSM
の境界面の細胞層より回収された(25)
。キムラ染色陽性細胞(43)
、すなわちマスト細胞の平均純度は43 ± 4% (9
検体 の平均±
標準誤差)
であった。比重遠心後に回収した滑膜マスト細胞を無血清Iscove’s methylcellulose medium
に200 ng/ml
のヒトリコンビナントSCF
と50 ng/ml
の ヒトリコンビナントIL-6
を添加したIMDM
で培養した 。42
日目にはmethylcellulose medium
をリン酸緩衝液(phosphate buffered saline; PBS)
で溶解し、0.1% BSA
、100 ng/ml
のヒトリコンビナントSCF
、50 ng/ml
のヒトリコンビナントIL-6
、100 units/ml
ペニシリン/
ストレプトマイシンを含有したIMDM (
マスト細胞[MC]
培地と呼ぶ。以下同様)
に再浮遊させ培養を継続した。3.
線維芽細胞の分離と培養前述の滑膜マスト細胞の分離と培養で記述した方法と同様に滑膜細胞を酵素処理 まで行った後に、
2%
ウシ胎児血清と100 units/ml
ペニシリン/
ストレプトマイシンを添加した
IMDM (
以下、線維芽細胞培地と呼ぶ)
に細胞を再浮遊させ、フラスコ内において
48
時間、37 °C
でインキュベートすると、フラスコの底面に線維芽細胞を認める。トリプシン溶液を加えて細胞をはがした後に
PBS
で回収し、より大きなフラ スコに移し、線維芽細胞培地で培養する。4. OA
マスト細胞とRA
線維芽細胞との共培養OA
マスト細胞とRA
線維芽細胞をcollagen coting
の24
穴プレートを用いてマスト 細胞培地で96
時間共培養する。まずRA
線維芽細胞をプレートに線維芽細胞培地と ともに加え、48
時間インキュベートしコンフルエントにする。その後培地をマスト 細胞培地に置き換え、一穴につき3×10
5個ずつOA
マスト細胞を加える。96
時間 後、マスト細胞のみ単離する。RA
線維芽細胞は底面に張り付くため、弱くピペッテ ィングするとOA
マスト細胞のみ単離可能であった。5.
滑膜マスト細胞の活性化FcεRI
の架橋刺激実験では、マスト細胞に0.5 μg/ml
のヒトミエローマIgE
を添加15
し
37°C
で30
分間感作させ、一度洗浄した後に4-(2-hydroxyethyl)-1-
piperazineethanesulfonic acid (HEPES)
緩衝液も再浮遊させた。ヒトリコンビナントIgE
で感作されたマスト細胞を、ヒスタミン、PGD
2測定実験(
マスト細胞数:100 μl
あたり1×10
3個)
では抗IgE
抗体で30
分間刺激し、サイトカイン測定実験(
マスト 細胞数:100 μl
あたり1×10
5個)
では抗IgE
抗体で6
時間刺激した。Fc γ RI
を介したIgG
依存刺激を行った。Fc γ RI
の架橋には、まずマスト細胞を抗Fc γ RI F(ab')
2フラグ メント(clone 10.1)
、またはマウスIgG1 F(ab')
2フラグメントとともに、それぞれ10
μg/ml
の濃度で37 °C
で30
分間インキュベートした。そして洗浄後に、HEPES
緩衝液で再浮遊させ、得られたマスト細胞を
1
および3 μg/ml
の抗マウスIgG F(ab')
2フラ グメント特異的ヤギF(ab')
2フラグメントとともにヒスタミンおよびPGD
2アッセー(
マスト細胞数:100 μl
あたり1×10
3個)
では30
分間インキュベートした。6. total RNA
抽出、相補的DNA (cDNA)
への逆転写、定量的RT-PCR
RNeasy Mini Kit (QIAGEN, Valencia, CA, USA)
を用いて、OA
およびRA
滑膜由来 マスト細胞からtotal RNA
を抽出した。抽出したtotal RNA (100 ng)
を10 mM dNTP mix
、oligo-dT primer (Invitrogen Life technologies)
、nuclease-free water
を混合し、65ºC
で5
分反応後、4ºC
で急冷した。この混合液に5×First strand buffer
、0.1 M 1,4-
Dithiothreitol (DTT)
、RNase out
、Super Script II (Invitrogen Life technologies)
を添加 し、42ºC
で50
分、72ºC
で15
分反応させ、4ºC
で急冷し、total RNA
をcDNA
に逆転 写した。逆転写したcDNA (5 ng)
とTaqMan Universal master Mix II (Applied
Biosystems)
、FAM
標識された各遺伝子に対するプローブセット(Applied Biosystems)
もしくはVIC
標識されたGAPDH
とβ-actin
に対するプローブセット(Applied
Biosystems)
、Nuclease free water
を、96
穴プレートに添加し、定量的RT-PCR
を実施 した。このPCR
は、StepOne Plus (Applied Biosystems)
で行なった。定量的RT-PCR
データは、StepOne software v2.1 (Applied Biosystems)
で解析し、GAPDH
もしくはβ-
actin
の発現量を用いて各遺伝子の発現量を標準化した。7. DNA chip
解析OA
マスト細胞とRA
マスト細胞の発現遺伝子をDNA chip
を用いて網羅的解析を 行った。OA
マスト細胞とRA
マスト細胞からRNeasy Mini kit (QIAGEN)
を用いて16
total RNA
を抽出し、前述の方法でcDNA
に逆転写した。逆転写したcDNA
とbiotin
標識されたヌクレオチド三リン酸を用いて、ビオチン化相補的
RNA (Biotin-cRNA)
を合成した。Biotin-cRNA
とHuman Genome U133 (Affymetrix)
を45ºC
で16
時間反 応させ、ハイブリダイゼーションした。その後、streptavidin-phycoerythrin (PE)
と反 応させ、Hewlett-Packard Gene Array Scanner (Palo Alto, CA, USA)
を用いて蛍光強度を 読み取った。各プローブの蛍光強度は、GeneChip Analysis Suite 5.0 (Affymetrix)
で数 値化した。数値化したデータをGenespring software (Agilent Techologies)
を用いて解 析し、RA
マスト細胞における発現量がOA
マスト細胞と比較して高かった遺伝子群 を抽出した。8. miRNA
の網羅的発現解析OA
およびRA
マスト細胞(
それぞれ3
ドーナー)
からmiRNeasy Mini kit (Qiagen, Hilden, Germany)
を用いてmiRNA
を抽出した。抽出したmiRNA (100 ng)
に、miRNA Spile-In solution (Agient Technoligies [Santa Clara, CA, USA])
、CIP Master Mix
を添加し、37°C
で30
分間反応させ脱リン酸化させた。その後、DMSO
を添加し、100°C
で10
分間反応させ、氷上で冷却し反応を停止させた。脱リン酸化処理したmiRNA
溶液にCy3
を含んだLigation Master Mix (Agilent Technologies)
を添加し、16°C
で2
時間反応させた。Cy3
でラベルしたmiRNA
を精製し、miRNA Complete Labeling and Hyb Kit (Agilent Technologies)
と、55°C
で20
時間反応させ、ハイブリダ イゼーションさせた。その後miRNA
を抽出した。miRNA
の網羅的発現解析はヒトmiRNA Micro assay kit Release16.0 (Agient Technoligies [Santa Clara, CA, USA])
を用い て行った。9.
メディエーターの測定PGD
2の測定にはPGD
2-methoxylamine (MOX) enzyme immunoassay (EIA) kit
(Cayman Chemical [Ann Arbor, MI, USA])
を用いた。PGE
2の測定にはPGE
2EIA kit
(Cayman Chemical [Ann Arbor, MI, USA])
を用いた。ヒスタミンの測定にはEIA kit
(MBL ([Tokyo, Japan])
を用いた。ヒスタミン遊離率は、(
放出されたヒスタミン量/
未刺激のマスト細胞に含まれる総ヒスタミン量)×100%
として算出した。17 10.
関節液の採取先述の滑膜採取と同様に、日本大学医学部付属板橋病院で行われた人工膝関節置 換術の際、
OA10
例およびRA10
例より関節液を約10ml
採取した。OA14
例はすべて
NSAIDs
で治療されていた。RA3
例は抗TNF- α抗体あるいは、抗 IL-6
抗体で治療されていたが、手術の
2-4
週間前に生物学的製剤は中止した。11.
統計処理OA
とRA
の2
群間の検定においては正規分布に従ってない分布の中心分布の差を 検定するため、Mann-Whitney
のU
検定を用いた。miR-199a-3p
とPTGS2
の発現量の相関は
Spearman
の順位相関係数を用いて相関の強さを検定した。解析には、GraphPad Prism 6 (MDF, Tokyo, Japan)
を用いた。p < 0.05
を統計学的に有意差がある とした。18
結果
1. DNA chip
解析によるOA
、RA
マスト細胞における遺伝子発現量の比較osteoarthritis (OA)
とrheumatoid arthritis (RA)
の滑膜中のマスト細胞の特徴を解析 するため、OA
患者(3
ドーナー)
とRA
患者(3
ドーナー)
の滑膜から、マスト細胞 を培養し、DNA chip
を用いて、遺伝子プロファイルを網羅的に解析した。図2
は、クラスター解析によって
RA
マスト細胞においてOA
マスト細胞と比較して発現が高 かった遺伝子群を示す。prostaglandin
、thromboxane
やleukotriene
の合成酵素であるPTGS1
、PTGS2
、TBXAS1
およびLTC4S
が、OA
由来培養マスト細胞よりもRA
由 来培養マスト細胞で高発現していた(
図2)
。これらのことから、OA
およびRA
マス ト細胞は、脂質メディエーターの生合成能に違いがあること示唆された。次に、OA
およびRA
由来培養マスト細胞のアラキドン酸カスケードにおける、脂質メディエ ーターの合成酵素の発現をより詳細にDNA chip
のデータを用い比較した。発現量はnormalize
したRA3
ドーナー、OA3
ドーナーの発現量の平均化で示した。図3A
ではprostaglandin
系代謝の酵素であるPTGS1
、PTGS2
、L-PGDS
、H-PGDS
、prostaglandin E synthase 1 (PTGES1)
、PTGES2
、prostaglandin I synthase (PTGIS)
、thromboxane
系代 謝の酵素であるTBXAS1
、leukotriene
系代謝の酵素であるLTA4H
、LTC4S
、ALOX5
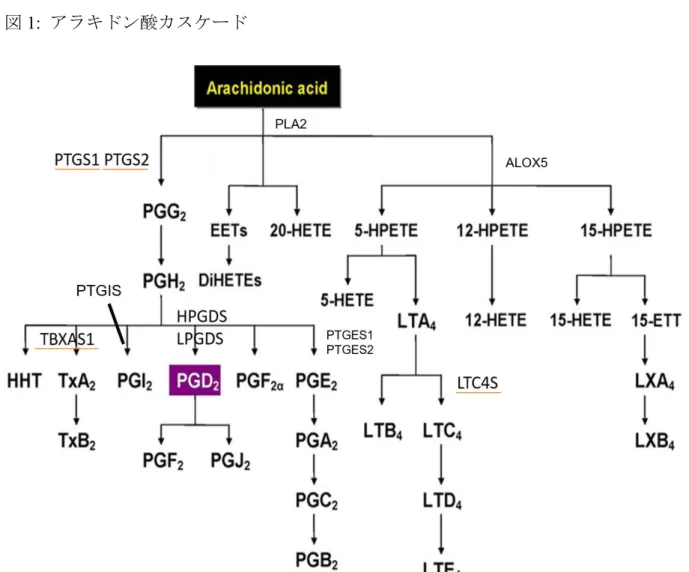
の発現量を示している。図3B
はprostaglandin
受容体である(prostaglandin D receptor) PTGDR
、CRTH2
、prostaglandin E receptor 2 (PTGER2)
、prostaglandin E receptor 3 (PTGER3)
、prostaglandin E receptor 4 (PTGER4)
、leukotriene
系受容体であるcycteinyl leukotriene receptor 1 (CYSLTR1)
の発現量を示している。結果、両マスト細胞間で、顕著に発現が異なる合成酵素は、
PTGS1
、PTGS2
、TBXAS1
やLTC4S
であった(
図3A and B)
。図3C
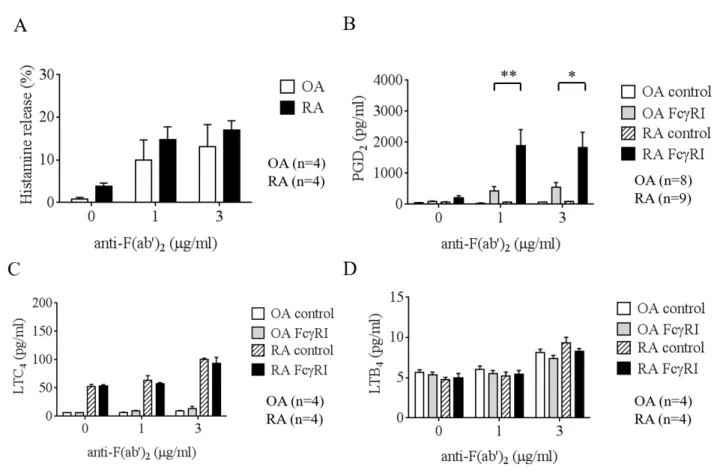
はマスト細胞特異的遺伝子群の発現量を示しており、マスト細胞特異的遺伝子群の発現量は、両マスト細胞間で、有意な差が認められなかったこと から、これらの合成酵素の発現が異なる原因は、マスト細胞の成熟度の違いによる ものではないことが考えられた。
2.
定量的RT-PCR
解析によるOA
、RA
マスト細胞における脂質メディエーター合成酵素の発現の比較
19
DNA chip
で得られた結果を確認するためにOA
、RA
マスト細胞間に差があるという
PTGS1
、PTGS2
、TBXAS1
およびLTC4S
の遺伝子発現量を定量的RT-PCR
法で解 析した。PTGS1
、PTGS2
、PTGES
、LTC4S
およびTBXAS1
の発現量を測定した結 果、RA
マスト細胞で有意に高かったのはPTGS1
、PTGS2
、LTC4S
およびTBXAS1
であった(
図4A)
。prostaglandin E synthase (PTGES)
の発現量は低く、有意差は見ら れなかった。造血型prostaglandin D synthase
であるH-PGDS
の発現量を測定したが有 意差は見られなかった(
図4B)
。3. OA
、RA
マスト細胞におけるIgE
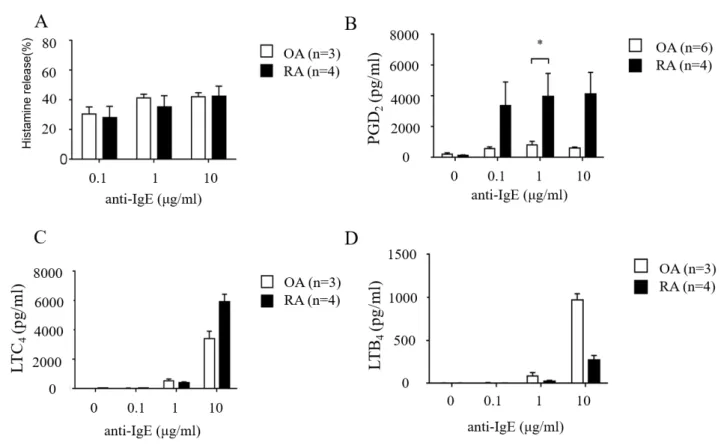
依存性脱顆粒反応および脂質代謝物産生量の比 較遺伝子発現量に有意差があった酵素により影響される脂質メディエーターに
PGD
2、LTC
4が挙げられる。OA
、RA
マスト細胞にIgE
依存性の刺激を行い、その 産生量を比較した。抗IgE
抗体刺激によるヒスタミン遊離量においてOA
、RA
マス ト細胞間に有意差はなかった(
図5A)
。抗IgE
抗体刺激により産生されるPGD
2の産 生量はRA
マスト細胞で有意に高く、LTC
4の産生量は、RA
マスト細胞で高い傾向 にあった(
図5B and 5C)
。一方、抗IgE
抗体刺激によるLTB
4の産生量はOA
で高い 傾向にあった(
図5D)
。4. OA
、RA
マスト細胞におけるIgG
依存性脱顆粒反応および脂質代謝物産生量の比 較IgG
依存性刺激によるヒスタミン遊離量およびLTB
4産生にOA
、RA
マスト細胞間 で有意差はなかった(
図6A and D)
。また、PGD
2産生量はRA
マスト細胞の方が有意 に高かった(
図6B)
。図6C
、6D
はLTC
4、LTB
4の産生量を示すが、ほとんど産生さ れなかった。5. OA
、RA
線維芽細胞におけるPTGS1
、PTGS2
、TBXAS1
、LTC4S mRNA
の比較とOA
マスト細胞とRA
線維芽細胞共培養によるPTGS1
、PTGS2
、TBXAS1
、LTC4S mRNA
発現量への影響OA
とRA
マスト細胞でみられたPTGS1
、PTGS2
、LTC4S
およびTBXAS1 mRNA
の発現量の差が線維芽細胞でもみられるかどうかを検討するためOA
患者、RA
患者20
の滑膜から単離、培養した線維芽細胞の遺伝子発現量を定量的
RT-PCR
法で測定し た。PTGS1
、PTGS2
、TBXAS1
およびLTC4S mRNA
の発現は、OA
、RA
線維芽細胞 間に有意差はなかった(
図7A)
。ALOX5 mRNA
の発現は感度以下であった(data not
shown)
。またOA
およびRA
線維芽細胞培養上清中のPGD
2量を測定したがEIA
検出感度以下であった
(data not shown)
。 マスト細胞の顆粒成熟や細胞表面分子の発現 は、微小環境に存在する線維芽細胞に影響を受けることが報告されている(44)
。こ れらの報告から、RA
とOA
マスト細胞の脂質メディエーター合成酵素発現の違い は、病態局所の滑膜を構成する線維芽細胞に依存しているという仮説を立てた。そ の仮説を検証するために、OA
由来培養マスト細胞とRA
由来線維芽細胞を共培養し た。マスト細胞と線維芽細胞間の相互作用によってPTGS1
、PTGS2
、TBXAS1
および
LTC4S mRNA
の発現量に変化があるかどうかを調べた。OA
マスト細胞とRA
線維芽細胞を
96
時間共培養し、OA
マスト細胞のみを単離し、遺伝子発現量を定量的RT-PCR
法で測定した。OA
マスト細胞は、RA
線維芽細胞と共培養してもPTGS1
、PTGS2
、TBXAS1
およびLTC4S mRNA
の発現は、亢進しなかった(
図7B)
。したが って、RA
の線維芽細胞はOA
マスト細胞のPTGS1
、PTGS2
、TBXAS1
およびLTC4S mRNA
の発現制御には影響を及ぼしていないことが明らかになった。6. OA
、RA
マスト細胞のmiR-199a-3p
発現量の比較遺伝子発現を制御する因子として、
miRNA
が重要な役割を担っていることが知ら れている。OA
とRA
マスト細胞に発現量の差がみられた遺伝子群を制御するmiRNA
の発現を解析するために、OA3
ドーナーとRA3
ドーナーを用いてmiRNA
chip
解析を行った。その結果、OA
マスト細胞の方がRA
マスト細胞より3
倍以上発 現量が高いmiRNA
を20
個見出した(
表3)
。これまでに、miR-199a-3p
がPTGS2
の 発現に関与することが報告されている(45, 46)
。実際にmiR-199a-3p
の発現に、RA
およびOA
マスト細胞間で違いが見られるかを検証するために、15
ドーナーのOA
マスト細胞と9
ドーナーのRA
マスト細胞におけるmiR-199a-3p
の発現を定量的RT- PCR
法で比較した結果、OA
マスト細胞の方が、miR-199a-3p
の発現が高い傾向にあ ったが有意差を認めなかった(
図8)
。7. OA
、RA
マスト細胞におけるmiR-199a-3p
とPTGS2
の発現の相関21
OA
マスト細胞のmiR-199a-3p
の発現量とPTGS2
の発現の相関を調べた。OA
マス ト細胞において相関はなかったが(
図9A)
、RA
マスト細胞では、miR-199a-3p
とPTGS2
の発現量には負の相関が認められた(R = -0.7167
、p = 0.0369) (
図9B)
。したがって、
miR-199a-3p
はPTGS2
の発現量を負に制御していることが示唆される。8. OA
、RA
患者の関節液中のPGD
2とPGE
2量関節炎と脂質メディエーターの関係を検証するために、
OA
、RA
患者から採取し た関節液中のPGD
2、PGE
2量をEIA
法で測定した。患者の詳細は表4
に示す。OA
患者10
名、RA
患者10
名から採取した関節液ではOA
患者よりRA
患者の関節液で は、PGD
2濃度が有意に高かった(
図10A)
。OA
患者14
名、RA
患者10
名から採取 した関節液ではPGE
2濃度はOA
、RA
間に有意差はなかった(
図10B)
。以上より、PGD
2は、RA
の病態と関与がある可能性が示唆された。22
考察
RA
の病態においては関節炎マウスモデルや滑膜の組織染色を用いた研究からマス ト細胞はRA
の発症や炎症の増悪に関与するいわゆる“
悪玉”
として考えられてき た。しかし、ここ1-2
年のRA
患者の関節滑膜組織中のマスト細胞関連遺伝子発現や 関節液中のマスト細胞由来のメディエーターがRA
の重症度や炎症マーカーと逆相 関することが報告され、“
善玉”
の可能性が示唆されていたが、その機序は不明であ った。本研究ではRA
マスト細胞が免疫複合体の刺激によって過剰なPGD
2を産生す ることにより、炎症を抑制する可能性を世界で初めて示した。OA
マスト細胞とRA
マスト細胞に発現している遺伝子の網羅的遺伝子解析および定量的RT-PCR
解析に より脂質メディエーター合成酵素に関するPTGS1
、PTGS2
、TBXAS1
およびLTCS4 mRNA
がRA
マスト細胞でOA
マスト細胞と比較して有意に発現が高かった。その 結果としてIgE
依存性刺激およびIgG
依存性刺激によるマスト細胞から産生されるPGD
2量はRA
マスト細胞で有意に高かった。その要因としてmiRNA chip
解析を行 い、PTGS2
の発現に影響を与えるmiR-199a-3p
に注目した。miR-199a-3p
はOA
マス ト細胞とRA
マスト細胞間に有意な発現量の差は認めなかったもののRA
マスト細 胞においてmiR-199a-3p
とPTGS2
の発現量は有意な負の相関がありmiR-199a-3p
は 少なくともRA
マスト細胞においてPTGS2
の発現を制御している一因子であること が示唆された。PGD
2受容体およびPGD
2合成酵素の欠損マウスを用いた研究からPGD
2、特にHPGDS
由来のPGD
2は炎症の抑制効果を持つことが示されている(47-50)
。その機序としては①
PGD
2が樹状細胞表面のDP
1を介して樹状細胞の遊走と機能を抑制し、結果として
T
細胞機能を抑制すること(49)
、②PGD
2の分解産物である15-deoxy- Delta12,14-prostaglandin J
2(15d-PGJ
2)
がperoxisome proliferator-activated receptor
(PPAR) -γ
依存性および非依存性の系を介して好中球の遊走を抑制することによって炎症を抑制すること
(50)
、③ 炎症の際にT
細胞とB
細胞から産生されるIL-10
を増 加させ、マクロファージから産生される炎症性サイトカインTNF-α
を低下させること
(50)
が示唆される。実際、関節炎マウスモデルにおいて、PGD
2受容体の阻害薬(51)
、PGD
2受容体欠損マウス(30)
を用いた報告からもPGD
2は炎症抑制効果をもつ23
ことが示唆される。ヒトの
RA
においてPGD
2の阻害薬を用いた臨床研究はなく、PGD
2の直接作用は不明である。PTGS
の阻害薬であるNSAIDs
は抗炎症作用がありOA
とRA
に用いられている。NSAIDs
がRA
の病態を悪化させたという報告はないが、この理由としてNSAIDs
はPGD
2のみならずPGE
2やPGI
2の産生を抑制し、PGE
2は関節炎症を惹起している主要 因子であることから(52)
、抗炎症効果はPGE
2の抑制効果である(53)
。したがって マスト細胞が産生されるPGD
2を特異的に抑制した場合、炎症は増悪するかどうかは 今後の更なる検討が必要である。RA
患者の関節滑膜組織におけるKit
やtryptase
などのマスト細胞特異的遺伝子発 現量とRA
の重症度には負の相関が見られること(26)
やRA
患者の血清CRP
とtryptase
レベルにも負の相関が見られること(27)
より、マスト細胞は、RA
の病態において抗炎症作用がある可能性があると示唆されていた。本研究では、
RA
マスト細 胞からのPGD
2過剰産生がその抑制効果の一因である可能性が示唆された。実際、大 腸炎および大腸炎関連大腸がんマウスモデルにおいては、マスト細胞から産生され るPGD
2が大腸炎および大腸炎関連大腸がんの抑制因子であると報告されており(54)
、RA
においても同様の機構が想定されるが、関節炎マウスモデルを用いた検証 が必要である。最新の報告においてもK/BxN-PA
を用いて時期特異的にマスト細胞 を欠損させると、発症前にマスト細胞を欠損させた時のみ炎症を抑制したという報告がある
(17)
が、マスト細胞からはTNF-α
、IL-6
、ヒスタミン、プロテアーゼやPGD
2といった様々なメディエーターが遊離、産生されるため、本研究とは異なった 結果がでたものと考えられた。本研究では今後マスト細胞から産生されるPGD
2のみ を欠損させ関節炎マウスモデルで検討する予定である。Lee
らはヒトRA
とOA
の関節滑膜からマスト細胞を分離し、その表現型を解析し た 。その結果、免疫複合体がFcγRI
、FcγRIIA
を介して関節滑膜マスト細胞を活性 化することを報告した(25)
。しかしRA
とOA
の関節滑膜マスト細胞表面のFcγRI
、FcγRIIA
、FcγRIII
およびFcεRI
の発現量に差はなく、FcγRI
やFcεRI
の架橋に よって遊離されるヒスタミン量やサイトカイン量にも差は見られなかった。この時 点ではアラキドン酸代謝産物の産生量の違いは検討しなかったが、本研究の検討で その差が明らかとなった。RA
の炎症局所においてマスト細胞を活性化させる因子は 免疫複合体であり、実際にヒトの抗 シトルリン化蛋白抗体がヒトのマスト細胞を活24
性化させたという報告がある
(26)
。本研究のマスト細胞の刺激実験においてIgE
依 存性の系を用いた理由は、Lee
ら(25)
の検討で免疫複合体による刺激よりもマスト 細胞に強い活性化が起こり、Fc ε RI
、Fc γ RI
およびFc γ RIIA
のすべての受容体はcommon γ -chain (FcR γ )
とのダイマーであり、共通の細胞内シグナル伝達系を使うため
(55)
、一つの実験系として使用した。もちろんRA
の炎症局所でIgE
依存性の刺激でマスト細胞が活性化することは、考えにくい。
RA
マスト細胞をIgE
依存性の刺激を行うとLTC
4はRA
で有意に多量に産生されたことは
LTC4S
の発現が高いことで説明される(
図4A)
。一方LTB
4がOA
マスト細胞から有意に多量に産生される理由としてアラキドン酸カスケード
(
図1)
より、OA
においてはLTC4S
の発現量が低く、相対的にLTB
4の産生に傾いたものと考えら れる。また、IgG
依存性刺激ではLTC
4、LTB
4の産生量が低かった(
図6C
、6D)
。Fc ε RI
はαβγ2
のヘテロテトラマーであるがFc γ RI
とFc γ RIIA
はαγ2
のヘテロダイマー でβ
鎖をもたない(56)
。β
鎖はロイコトリエンの産生を上げることをNunomura
らは 示しており(57)
、IgG
依存性刺激ではβ
鎖が関与しないためと考えられる。したがっ て、RA
の炎症局所においてマスト細胞を活性化させる因子は免疫複合体であるので アラキドン酸代謝産物としてはPGD
2のみ過剰産生されると考えられる。OA
をRA
の対照疾患としていいかという問題がある。確かに健常人の滑膜組織を そのコントロールとすべきであるが、倫理的な問題があり、健常人から研究のため に滑膜組織を採取することは極めて難しい。RA
研究においては歴史的にRA
のコン トロール疾患としてOA
を用いてきた(21-24)
。OA
の発症年齢は通常50
歳以上で、女 性は男性の3
~4
倍多い点もRA
の患者の年齢と性差に有意差がつき難いという点(58)
、OA
では通常CRP
は陰性で病態が炎症ではない点、また、OA
患者で人工膝関節 置換術を施行される患者数も多く、検体が入手しやすい点も歴史的にOA
がコントロ ール疾患として使用されてきた理由であろう。Lee
らはヒトRA
とOA
の関節滑膜からマスト細胞を分離し、分離直後のマスト細 胞と培養滑膜マスト細胞の顆粒構造を電子顕微鏡による観察にて比較し同様の顆粒 構造であること、細胞表面に発現している受容体を比較し、Kit
、Fc γ RI
、Fc γ RIIA
、Fc γ RIII
およびFc ε RI
の発現量に差はなかったが、Fc γ RII Βの発現量に差異があったこ
とを報告した(25)
。すなわち、本研究で実験に供したマスト細胞は培養滑膜マスト 細胞であり、このマスト細胞は、滑膜組織の炎症局所に存在するマスト細胞と同一25
ではない。そのため現在、本研究で示した
PGD
2の産生量の差異について、ヒトRA
とOA
の関節滑膜からマスト細胞を分離し、分離直後のマスト細胞を用いて比較検 討している。さらに、本研究で使用した
RA
の滑膜組織は 人工関節を要した極めて重篤かつ進 行したものであり、 様々な治療介入後の検体のため初期や活動性のある時の病態で はマスト細胞は違う動きをしている可能性が十分考えうる。最近の報告では、活動 性のありなしのRA
の滑膜組織中のマスト細胞数を比較すると活動性のある場合に 有意に増加しており、寛解したままの患者と再燃した患者のRA
の滑膜組織中のマ スト細胞数を比較すると再燃した患者で有意に増加しているという(59)
。このよう にRA
の活動性の違いにより微小環境の影響を受けてマスト細胞は、動態を変化さ せると思われ、本研究のデータはあくまでも人工関節を要した極めて重篤かつ進行 したRA
の 滑膜組織 から樹立した培養マスト細胞の結果であるという限界が存在す る。また、
RA
のある患者は抗サイトカイン生物学的製剤で治療されていたが、英国お よび日本リウマチ学会のガイドライン(60, 61)
に従い術前2-4
週間前に中止した。インフリキマブでは、血中半減期は、
8.4
日であり、2-3
週間の休薬にて有効血中濃 度以下になり、エタネルセプトでは、血中半減期は4.6
日であり、1週間以上の休薬 にて有効血中濃度以下になるが(62)
、DMARDs
のみの使用RA
患者と比較して抗TNF
抗体薬剤使用RA
患者において手術部位の感染頻度が高いという報告があり(63)
、術前2-4
週間前に中止しても免疫に対する効果は無視できないため、データの 解釈も慎重であるべきであろう。また、OA
患者すべてにNSAIDs
が投与されていた がNSAIDs
はPTGS1
およびPTGS2
の酵素の阻害薬であり(64)
、PTGS2 mRNA
の発 現量には影響を及ぼさない。しかしながら、本研究で使用したRA
およびOA
の培 養滑膜細胞は少なくとも12
週間両者同じ条件で培養した後の細胞であり、培養前に 使用した 抗サイトカイン生物学的製剤やNSAIDs
の影響が結果を反映しているとは 考えにくい。12
週間以上同一の条件で培養しても両マスト細胞間に発現している特定の遺伝子 の発現量が異なるという事実は、マスト細胞の分化の違いやメチル化などepigenetic
な修飾が関与していることが想定される。実際Taketomi
らとの共同研究でIL-3
依存 性骨髄由来培養マスト細胞(
粘膜型で未熟なマスト細胞)
を線維芽細胞と共培養した26
時
PTGS1 mRNA
は1.4
倍、PTGS2 mRNA
は387
倍、発現量が上昇したため(45)
、 線維芽細胞との細胞間相互作用による成熟を想定したが影響はなかった。これは、滑膜マスト細胞が結合織型のマスト細胞でありすでに成熟度が高いマスト細胞であ ると考えた。次に
miRNA
に注目した。miRNA chip
解析を行い、miR-199a-3p
に注目 した理由は、OA
マスト細胞の方がRA
マスト細胞より3
倍以上発現量が高い20
個の
miRNA
のうち、唯一miR-199a-3p
が直接にPTGS2 mRNA
の発現を関節の軟骨細胞において抑制したという報告があったからである
(45, 46)
。miR-199a-3p
発現を検 討した実験ではRA
マスト細胞ではmiR-199a-3p
の発現とPTGS2
の発現との間に負 の相関が見られたことから、RA
マスト細胞ではmiR-199a-3p
によりPTGS2
発現が 制御されている可能性が考えられた。滑膜マスト細胞における詳細な機序を調べる ためには、OA
マスト細胞にanti-miR-199a-3p inhibitor (AM11779, has-mir-199a-3p;
Ambion, Austin, TX)
をトランスフェクションすることによりmiR-199a-3p
の発現を 抑制し(65) PTGS2
の発現が上がることと、RA
マスト細胞にmiR-199a-3p
を強制発現させ
PTGS2
の発現が下がることを証明しなければならない。今後の課題である。また、
RA
マスト細胞におけるPTGS2
発現調節に他のmiRNA
が関わっている可能性 も考えられる。ヒト肺がん細胞ではmiR-146a
がPTGS2
発現を制御しているという 報告されている(66)
。また、肝がん細胞 では正常肝細胞よりPTGS2
の発現が低 く、その制御にmiR-16
が関与していることが報告されている(67)
。今後はmiR chip
でOA
マスト細胞において高値を示した他のmiRNA
に関してPTGS2
の発現制御に どのように関わっているかを検討する必要がある。また本研究ではPTGS2
の発現制 御のみに着目したが、今後はRA
マスト細胞で高かった別の脂質代謝関連遺伝子に 関しても同様にmiRNA
による調節が起こりうるかを検討する必要がある。さらに、Kuwano
らの報告ではPTGS2
発現はIL-1β
やTNF-α
などのサイトカインによっても制 御されることが示されており(68)
、炎症性サイトカインによるPTGS2
および他の遺 伝子の発現制御の検討も今後の課題である。また本研究ではDNA
メチル化について は検討しなかったが今後の検討が必要である。最後に