題 目
ゲノムワイド関連解析と fine mapping を用いた 痛風の感受性遺伝子の同定
崎山
さ き や ま真
ま さ幸
ゆ き(分子生体制御学専攻)
防衛医科大学校
平成 28 年度
略語一覧 1
用語集 3
第1章 背景 6
第2章 痛風のGWASによる新規関連遺伝子座の同定 10
2-1. 目的 10
2-2. 対象及び方法 10
2-2-1. 対象 10
2-2-2. 痛風・高尿酸血症の臨床病型分類 11
2-2-3. 遺伝子解析とそのquality control 11
2-2-4. 統計学的解析 12
2-3. 結果 13
2-3-1. GWAS 13
2-3-2. ABCG2遺伝子の機能低下型多型についての検討 14
2-3-3. リスクアレルの累積効果についての検討 15
2-3-4. 痛風の臨床病型の解析 16
2-4. 考察 17
第3章 MYL2-CUX2遺伝子領域における痛風関連遺伝子座の同定 21
3-1. 目的 21
3-2. 対象及び方法 21
3-2-1. 対象 21
3-2-2. SNPの選定 22
3-2-3. 遺伝子解析 22
3-2-4. 統計学的解析 23
3-3. 結果 23
3-3-1. 痛風の関連解析 23
3-3-2. 飲酒習慣を含めた検討 24
3-4. 考察 25
第4章 総括 27
第5章 結論 28
謝辞 29
付記 30
参考文献 33
図表 41
- 1 - 略語一覧
ABCG2; ATP–binding cassette (ABC), subfamily G, member 2 acetyl-CoA; acetyl-coenzyme A
ADH; alcohol dehydrogenase ALDH; aldehyde dehydrogenase AMP; adenosine monophosphate
AMPA; α-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate ATP; adenosine triphosphate
BMI; body mass index
CARMIL; capping protein ARP2/3 and myosin-I linker CI; confidence interval
CNIH-2; cornichon family AMPA receptor auxiliary protein 2 CP; capping protein
CUX2; cut-like homeobox 2
FEUA; fractional excretion of uric acid GCK; glucokinase
GCKR; glucokinase regulatory protein GLUT9/SLC2A9; glucose transporter 9 GWAS; genome-wide association study IBS; identity by state
J-MICC; Japan Multi-Institutional Collaborative Cohort LD; linkage disequilibrium
- 2 - LRRC16A; leucine-rich repeat-containing 16A
MAP3K11; mitogen-activated protein kinase kinase kinase 11 MRP4; Multidrug resistance-associated protein 4
MYL2; myosin, light chain 2, regulatory, cardiac, slow
NPT1/SLC17A1; sodium-dependent phosphate cotransporter type 1 OAT4/SLC22A11; organic anion transporter 4
OR; odds ratio
PACS1; phosphofurin acidic cluster sorting protein 1 QOL; quality of life
Q-Q plot; quantile-quantile plot RHUC; renal hypouricemia ROL; renal overload RUE; renal underexcretion
SNP; single nucleotide polymorphism SUA; serum uric acid
URAT1/SLC22A12; urate transporter 1 UUE; urinary urate excretion
- 3 - 用語集
一塩基多型(SNP; single nucleotide polymorphism)
ヒトの遺伝子は4種類の塩基(A; adenine、T; thymine、G; guanine、C; cytosine)の配 列によって成り立っているが、その配列は個人間で少しずつ異なっている。このような 個人差のうち、集団内で1%以上の頻度でみられる一塩基の違いだけによるものを一 塩基多型(SNP)と呼び、約1,000塩基に一か所の頻度で存在する。
ゲノムワイド関連解析(GWAS; genome-wide association study)
ヒトゲノム全体に渡って50万個以上の一塩基多型(SNP)をタイピングし、アレル頻度と 疾患もしくは血液検査結果などの量的形質との間に関連があるかどうかをみる遺伝統 計学に基づいた研究手法で、2002年に理化学研究所が初めて発表した。有意水準 は一般的に多重検定を考慮し、p < 5.0 × 10-8とされる。しかし、たとえこの厳しい水準 に達したとしても、実際にタイピングをしたSNPが真の原因SNPではなく、連鎖不平衡
(LD)にあるSNPのタグとして間接的な関連を示しているにすぎないこともある。この場 合はその領域を中心としたfine mappingが必要となる。
国際HapMap計画
Haplotype Mapping Projectのことで、日本人を含む様々な人種の全ゲノムにおけるハ
プロタイプマップ構築を目指し、米国国立衛生研究所(NIH)や理化学研究所をはじ め、世界中の複数の研究機関によって2002年から実施された。現在はその研究成果 が公開されており、ホームページ(https://hapmap.ncbi.nlm.nih.gov/)から一塩基多型
- 4 -
(SNP)やハプロタイプ等のデータが閲覧、利用できる。
多重検定
複数の検定を行うと、偶然に有意な結果が得られる確率が上昇する。例えば有意水 準がp < 0.05の時、X群, Y群, Z群の3群で平均体重を比較するにあたり、X群対Y 群、X群対Z群、Y群対Z群と3回検定を行うと、真に差が無い場合でも偶然に1つ の検定で有意となる確率が1 - (1 - 0.05) ×3 = 0.14となり、αエラー(第Ⅰ種の過誤)
が増大する。この問題を解決するためには様々な手法があり、本研究では有意水準を
「p < 0.05/検定数」に調節するBonferroni法を用いている。
連鎖不平衡(LD; linkage disequilibrium)
1本の染色体上の2座位間のアレルの関連のことで、「LDの関係にある」ことは「その 2座位間では組み換えが起きにくく関連が強い」ことを意味する。LDの指標としては D’値とr2値がよく用いられ、いずれも0から1の値を取り、値が大きいほど強いLDに あることを意味し、片方のアレルが判るともう一方のアレルが推測できることになる。逆 に数値が0に近い場合には2座位間での組み換えが起きやすく、片方のアレルが判 ってももう一方は予測困難である。
Fine mapping
ゲノムワイド関連解析(GWAS)等の全ゲノムにわたる解析は、それだけで疾患感受性 遺伝子が同定出来ることもある一方で、染色体上のある領域への絞り込みに留まるこ ともしばしばある。この場合、その領域についてさらに細かく遺伝子解析や統計解析を
- 5 -
行うことで、疾患感受性遺伝子を探索することができる。これをfine mappingと言い、本 論文の第3章がこれに該当する。
Hardy-Weinberg平衡
ある遺伝子座のアレルA、aが、それぞれ頻度p、1-pで存在する時、各遺伝子型の頻 度がA/A = p2、A/a = 2p(1-p)、a/a = (1-p)2となる法則のこと。この法則が成り立たない 場合(Hardy-Weinberg平衡の検定が統計学的に有意な場合)には、解析対象集団が 任意交配されていない可能性や遺伝子解析のエラーの可能性を考える必要がある。
- 6 - 第1章
背景
高尿酸血症(血清尿酸値が7.0 mg/dLを超える1)は、尿酸結晶の沈着による痛風 発作(激痛を伴う関節痛)の原因となるほか、高血圧、腎臓病、心臓病、脳卒中などの リスクとなることが知られている2。
生活習慣病のひとつである痛風は、「贅沢病」や「帝王病」などとも言われ、ルイ14 世、アイザック・ニュートン、レオナルド・ダ・ヴィンチも患ったとされる。我が国では、明 治初期にドイツ人医師のエルヴィン・フォン・ベルツが、「日本には痛風患者がいない」
と記録している3。しかし、今日では食生活が豊かになった影響もあり、我が国でも痛 風・高尿酸血症患者は近年増加傾向にある。現在、成人日本人男性の2~3割、すな
わち約1,000万人以上に高尿酸血症を認め、そのうちの一部が痛風を発症し、その数
は約100万人と言われている。また、痛風は中年以降の男性が多く発症する病気であ るとこれまでは考えられてきたが、最近は20歳代以下での発症も見られている3, 4。こ の原因として、食生活の欧米化や肥満などの環境要因が考えられるが、同じような生 活習慣にもかかわらず、痛風を発症する人としない人が存在することから、遺伝的要 因も推定されてきた(図1)。しかし、痛風・高尿酸血症が発症頻度の高い一般的な疾 患であるにもかかわらず、原因遺伝子としてこれまでに知られていたのは、
Lesch-Nyhan症候群やvon Gierke病など痛風を伴う稀な先天性疾患から同定された
ものに限定されていた。
近年、遺伝子解析技術が進歩し、ゲノムワイド連鎖解析やゲノムワイド関連解析
- 7 -
(GWAS; genome-wide association study)などによるヒトゲノム全体を網羅的に解析す る手法が登場したことにより、痛風や尿酸値の変動にかかわる遺伝子がいくつか同定 されてきた。例えばATP–binding cassette (ABC), subfamily G, member 2
(ABCG2/BCRP)遺伝子は尿酸輸送トランスポーター(輸送体)をコードしており、痛風 の主要な病因遺伝子であることが報告された5, 6。また、urate transporter 1
(URAT1/SLC22A12)遺伝子7やglucose transporter 9(GLUT9/SLC2A9)遺伝子8の機 能低下型変異が腎性低尿酸血症の原因となることが報告されている。しかし、痛風や 尿酸値の変動にかかわる遺伝子の全貌はいまだ明らかではない。
尿酸は、核酸やアデノシン三リン酸(ATP; adenosine triphosphate)などの構成成分 であるプリン体の代謝産物である。マウスを含む多くの哺乳類では尿酸分解酵素であ るウリカーゼにより尿酸が分解・代謝されるのに対し、ヒトではウリカーゼが欠損してい るため、尿酸はヒトにおけるプリン体の最終代謝産物となる9。肝臓を中心に1日あたり
約700 mg産生された尿酸は、約2/3が腎臓から排泄され、残り約1/3は主に腸管から
排泄される。腎臓では、尿酸は糸球体においてほぼ全量が濾過された後、近位尿細 管を中心に再吸収と分泌の両方が行われ、最終的に糸球体で濾過された尿酸の約6
~10%の量が尿中に排泄される。ここでの尿酸輸送機構には複数の尿酸トランスポー ターが関与している。我々の研究グループでは、これらの尿酸輸送に関連する分子に 着目し、その遺伝子の一塩基多型(SNP; single nucleotide polymorphism)解析を行い、
尿酸輸送に関わる新規遺伝子の同定をこれまで行ってきた。
URAT1は腎臓の近位尿細管上皮細胞の管腔側の細胞膜に局在する尿酸トランス
ポーターで、尿酸の再吸収に関わっており、腎性低尿酸血症1型の原因遺伝子として 知られている7。我々は、大規模な症例・対照研究を行うことによりURAT1遺伝子の尿
- 8 -
酸輸送機能消失型変異(Trp258Stop及びArg90His変異)が痛風・高尿酸血症の発症 に保護的に働くこと、変異の個数と血清尿酸値の低下の程度、血清尿酸値の男女差
の形成にURAT1が大きく関与していることなどを、明らかにしてきた10。
一方、ABCG2は近位尿細管上皮細胞の他、小腸上皮細胞の管腔側にも局在して おり、尿酸の体外への排泄を担っている5, 11。我々は、尿酸輸送機能低下をきたす ABCG2遺伝子の多型が、(1)若年型痛風の主な原因であること3, 4、(2)腎排泄低下 型高尿酸血症と腎外排泄低下型高尿酸血症の両者の主要な要因であること12、(3)
欧米人と比べ日本人に多いこと13などを発見した。
さらに腎近位尿細管に発現するorganic anion transporter 4(OAT4/SLC22A11)と sodium-dependent phosphate cotransporter type 1(NPT1/SLC17A1)の遺伝子多型が 腎排泄低下型痛風の発症に関連することも明らかにした14, 15。NPT1遺伝子で解析を 行ったSNPはrs1165196(Ile269Thr)であり、これは機能獲得型(gain of function)変異 であったが、その尿酸輸送機能の上昇には、トランスポーターのturnover rateの増加 が寄与している可能性も報告した16。
ヒトの腎近位尿細管の刷子縁膜側においてこれらの尿酸トランスポーターは、PDZ 蛋白質により束ねられて複合体を形成し、生体膜における物質輸送の機能単位となっ ている17。これをトランスポートソームと呼び、アクチンフィラメントがこのトランスポートソ ームの足場となっている(図2)。Capping protein(CP)はアクチンフィラメントの重合を 制御しているが、leucine-rich repeat-containing 16A(LRRC16A)遺伝子がコードする capping protein ARP2/3 and myosin-I linker(CARMIL)は、このCPを阻害することによ り18、トランスポートソームに影響を及ぼしている可能性がある。我々はLRRC16A遺伝 子の多型が痛風の発症に関連することを発見し、トランスポートソームと疾患の関連を
- 9 - 遺伝学的に初めて明らかにした19。
以上のように、我々の研究グループではこれまでに複数の痛風・高尿酸血症関連 遺伝子を同定してきたが、これらの候補遺伝子解析で同定できる原因遺伝子は、
ABCG2、URAT1、NPT1やOAT4など主に尿酸の輸送に関わるものに限定されていた。
そこで本研究では、まず痛風のGWASを行った。SNPアレイを用いてゲノム全体にわ たる遺伝子を解析し、アレル頻度が患者群と対照者群で異なるSNPを網羅的に探索 することで、痛風・高尿酸血症の病態形成に関わる遺伝子のさらなる同定を目指した。
さらに、GWASで同定された痛風関連遺伝子領域のうち、GWASのみでは真に痛風 の発症と関連する遺伝子が絞り込めなかった領域については、fine mapping(精細マ ッピング)を行うことで、その同定を目指した。
なお本研究は、ヘルシンキ宣言に従い、防衛医科大学校倫理委員会の承認を得 て実施した。
- 10 - 第2章
痛風のGWASによる新規関連遺伝子座の同定
2-1. 目的
近年、痛風のGWASが複数報告されているが20, 21、それらには臨床所見が不十分 である自己申告による痛風症例が含まれており、適切な病型分類にもとづいた遺伝子 変異の実態はいまだ不明のままである。そこで、我々は医師による臨床診断を受けた 痛風症例のみを対象としたGWASを世界に先駆けて実施して、さらに臨床病型との関 連も評価した。
2-2. 対象及び方法
2-2-1. 対象
痛風患者946人と対照者1,213人を用いて一次解析を実施し、さらに別の痛風患者
1,048人と対照者1,334人を用いて二次(再現)解析を行った(表1)。痛風症例は、み
どりヶ丘病院(大阪府)、京都工場保健会診療所(京都府)及び両国東口クリニック(東 京都)を受診した患者のうち、米国リウマチ学会の診断基準22をもとに医師が痛風であ ると確定診断した日本人男性症例のみを用いた。対照者はバイオバンク・ジャパン23 及び浜松市での健診における日本多施設共同コーホート研究(J−MICC Study; Japan Multi-Institutional Collaborative Cohort Study)24, 25の参加者で、痛風・高尿酸血症に 罹患していない日本人男性を対象とした。検体の採取にあたっては、インフォームド・
- 11 - コンセントを行い、同意書を取得した。
2-2-2. 痛風・高尿酸血症の臨床病型分類
血清の尿酸値とクレアチニン値および蓄尿による尿中の尿酸値とクレアチニン値を 測定し、尿中尿酸排泄率(FEUA; fractional excretion of uric acid)と尿中尿酸排泄量
(UUE; urinary urate excretion)を算出した。FEUA < 5.5%を腎排泄低下(RUE; renal underexcretion)型、UUE > 25 mg/h/1.73 m2(600 mg/day/1.73 m2)を腎負荷(ROL;
renal overload)型と分類した11(図3)。なお、FEUAとUUEの計算式は以下のとおりで ある。
FEUA =
)
( 尿中クレアチニン濃度
) 血清尿酸濃度(
)
( 血清クレアチニン濃度
) 尿中尿酸濃度(
mg/dl mg/dl
mg/dl mg/dl
×
× ×100%
UUE =
100 min
60 ml mg/dl
×
×
×
) 蓄尿時間(
) 尿量(
) 尿中尿酸濃度(
×体表面積(m2) 73
. 1
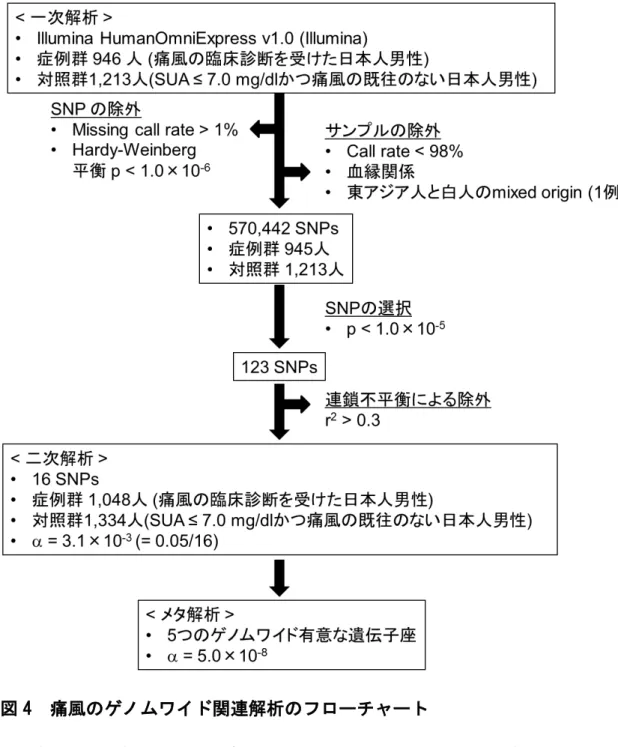
2-2-3. 遺伝子解析とそのquality control
DNAは全血から抽出した。一次解析では、痛風患者946人と対照者1,213人の検
体を、HumanOmniExpress v1.0(Illumina社)を用いて727,413 SNPsをタイピングした
(図4)。このうちタイピング成功率が99%未満のSNPとタイピング成功率が98%以下
の検体は、以後の解析からは除外した。また対照群におけるHardy-Weinberg平衡の 検定結果が統計学的に有意(p < 1.0 × 10-6)なSNPも除外した。さらにこの
HumanOmniExpress v1.0(Illumina社)のタイピング結果と国際HapMap計画phase
- 12 -
II26のデータをあわせて主成分分析27, 28を行い、東アジア人と白人のmixed originと 判断された1検体も除外した(図5)。なお同一(IBS; identity by state)アレル数の計算 も行ったが、血縁関係にある検体は見いだされなかった。加えてquantile-quantile plot
(Q-Q plot)も作成し、バイアスとなりうる集団構造化がないかどうかも検討した(図6)。
以上のquality controlを実施し、最終的に痛風患者945人と対照者1,213人の症例・
対照研究として、570,442 SNPsについて関連解析を実施した(図4)。
次に、一次解析でp < 1.0 × 10-5の基準を満たした123 SNPsについて、互いに連 鎖不平衡(LD; linkage disequilibrium)の関係にあるSNPがないか検討した。LDの指 標としてはr2値を用いた。r2値は0から1の値をとり、値が大きいほど強いLDにある、
すなわち2座位間での組み換えが起きにくいことを意味する。今回はr2 > 0.3という中 等度以上のLDの関係にあるSNPについては、一次解析で最もp値が低いSNPの みを二次解析の候補SNPとして選択した。その結果16 SNPsが二次解析の対象となり、
一次解析とは別の痛風患者1,048人と対照者1,334人をTaqMan法(Life
Technologies社)にてタイピングし、関連解析を実施した(図4)。二次解析ではタイピン
グ成功率が98%以下のSNPは存在せず、Hardy-Weinberg平衡の検定結果が統計学 的に有意(p < 0.05)なSNPも存在しなかった。
最後に一次解析と二次解析の結果をあわせたメタ解析を実施することで、評価を行 った。
2-2-4. 統計学的解析
関連解析は、症例群、対照群それぞれの疾患リスクアレル頻度を比較するカイ二乗 検定で実施し、オッズ比(OR; odds ratio)、95%信頼区間(95% CI; confidence
- 13 -
interval)とともに評価した。一次解析と二次解析の結果をあわせたメタ解析29を行うに
あたっては、Cochran’s Q検定30のp値とI2値31, 32で一次解析と二次解析の間に異 質性がないかどうかをまず判定した。Cochran’s Q検定のp < 0.05、もしくはI2 > 50%
の場合は異質性があると判断し、メタ解析にはランダム効果モデル
(DerSimonian-Laird法)33を用い、それ以外の場合は固定効果モデル
(inverse-variance法)を用いた。多重検定を考慮し、一次解析とメタ解析ではp < 5.0
× 10-8、二次解析ではBonferroni法による補正を行ったp < 3.1 × 10-3(= 0.05/16)を、
統計学的に有意な水準とした。
ABCG2遺伝子の3 SNPsの解析および痛風の病型分類ごとの解析では多変量ロジ スティック回帰分析、リスクアレルの累積効果の解析ではカイ二乗検定、痛風関連遺 伝子が臨床検査情報(FEUAとUUE)に与える影響については線形回帰分析、病型分 類と臨床検査情報(FEUAとUUE)の解析にはPearsonの相関係数の検定を用いた。
SNPの染色体上の位置は、NCBI human genome reference sequence Build 37.4の情 報に基づいて記した。全ての統計解析はSTATA v.11.0(Light Stone社)、SPSS v.22.0J
(IBM社)、R v.3.0.234とそのGenABELパッケージを用いて実施した。
2-3. 結果 2-3-1. GWAS
一次解析でゲノムワイド有意水準(p < 5.0 × 10-8)に達したのは、ABCG2遺伝子領 域、SLC2A9遺伝子領域、MYL2(myosin, light chain 2, regulatory, cardiac, slow)遺伝 子とCUX2(cut-like homeobox 2)遺伝子の間(MYL2-CUX2)領域の、合計3か所であ
った(図7、8)。それぞれの領域で最もp値が低かったSNPは、ABCG2領域では
- 14 -
rs2728125(p = 1.5 × 10-27; OR = 2.05; 95% CI = 1.80-2.34)、SLC2A9領域では rs3775948(p = 6.7 × 10-15; OR = 1.64; 95% CI = 1.45-1.86)、MYL2-CUX2領域では rs2188380(p = 5.7 × 10-13; OR = 1.78; 95% CI = 1.52-2.08)であった(表2)。
一次解析でゲノムワイド有意であった3つの遺伝子領域は二次解析でも有意であり、
再現性が確認できた。これらに加え、glucokinase regulatory protein(GCKR)遺伝子領 域のrs1260326(p = 2.8 × 10-6; OR = 1.32; 95% CI = 1.18-1.49)とcornichon family AMPA receptor auxiliary protein 2(CNIH-2)遺伝子領域のrs4073582(p = 1.6 × 10-4; OR = 1.55; 95% CI = 1.23-1.96)も二次解析では有意(p < 3.1 × 10-3)であった(表 2)。
一次解析と二次解析をあわせたメタ解析では、先述の5つの遺伝子領域(ABCG2、
SLC2A9、MYL2-CUX2、GCKR及びCNIH-2)のSNPであるrs2728125(pmeta = 7.2 × 10-54; OR = 2.04; 95% CI = 1.86-2.23)、rs3775948(pmeta = 5.5 × 10-27; OR = 1.61; 95%
CI = 1.47-1.75)、rs2188380(pmeta = 1.6 × 10-23; OR = 1.75; 95% CI = 1.57-1.96)、
rs1260326(pmeta = 1.9 × 10-12; OR = 1.36; 95% CI = 1.25-1.48)、rs4073582(pmeta = 6.4
× 10-9; OR = 1.66; 95% CI = 1.40-1.96)の全てが有意であり(図8、表2)、最終的にこ れらを痛風関連遺伝子領域と考えた。また、mitogen-activated protein kinase kinase kinase 11(MAP3K11)遺伝子領域のrs10791821は、メタ解析でpmeta = 1.0 × 10-7とゲ ノムワイド有意水準(p < 5.0 × 10-8)に近かったため(表2)、痛風関連遺伝子領域の 可能性があると考えた。
2-3-2. ABCG2遺伝子の機能低下型多型についての検討
尿酸トランスポーターABCG2は、2つのSNPsの組み合わせによって機能低下が生
- 15 -
じ、痛風の発症のしやすさに関与していることが解明されている5。すなわち、日本人 痛風男性症例の約8割に尿酸輸送能が消失するrs72552713(Gln126Ter)もしくは尿 酸輸送能が半分に低下するrs2231142(Gln141Lys)のリスクアレルがみられ、発症リス クを約3倍以上高めていることが、これまでに報告されている5。このため、今回の GWASにおいてABCG2遺伝子領域で最も痛風と関連の強いSNPとして同定された rs2728125は、尿酸輸送機能低下をきたすこれら2つのSNP(rs72552713と
rs2231142)のsurrogate markerである可能性を考えた。
これら2つのSNPsはHumanOmniExpress v1.0(Illumina社)に搭載されておらず、
一次解析ではタイピングされていなかった。そこで、TaqMan法(Life Technologies社)
により追加でこれら2つのSNPをタイピングしたところ、2つとも痛風と有意な関連を認 めた(表3)。さらに、rs2231142はrs2728125と強いLD(r2 = 0.76)にあることが判明し た。このためrs72552713、rs2231142およびrs2728125の3つのSNPsによる多変量ロ ジスティック回帰分析を行うことで互いに補正し、再度評価を行った。その結果、
rs72552713とrs2231142は単変量解析と同じく統計学的に有意(それぞれp = 1.8 × 10-18、1.9 × 10-12)であったが、rs2728125はp = 0.19となり有意性は消失した(表3)。
以上より、ABCG2遺伝子領域の真に痛風発症と関連するSNPはrs72552713と
rs2231142であると考え、以後の解析にはこれら2つのSNPsを用いた。
2-3-3. リスクアレルの累積効果についての検討
今回同定した痛風関連SNPs(ABCG2領域のrs72552713とrs2231142、SLC2A9領 域のrs3775948、MYL2-CUX2領域のrs2188380、GCKR領域のrs1260326、CNIH-2
領域のrs4073582)のリスクアレルをひとりの人が同時に複数有した場合、痛風のリスク
- 16 - がどの程度高まるかについて検討した。
有するリスクアレルが4個以下の群を基準(= 1)とした場合のORは、5個有する群 では1.79(p = 3.5 × 10-3)、6個有する群では3.16(p = 2.3 × 10-10)、7個有する群で は5.10(p = 9.7 × 10-21)、8個有する群では10.1(p = 5.3 × 10-39)、9個有する群では 18.6(p = 3.6 × 10-45)であった(図9)。
2-3-4. 痛風の臨床病型の解析
「2-3-1. GWAS」の対象症例のうち、FEUAとUUEの臨床検査情報が得られた痛風 症例を病型分類し(375例が腎負荷型、509例が腎排泄低下型)、GWASで同定した SNPsの痛風発症に与える影響が病型ごとに異なるか、解析を行った(表4)。
ABCG2遺伝子の2つのSNPs(rs72552713とrs2231142)のORは、腎負荷型では それぞれOR = 4.35、3.37、腎排泄低下型ではそれぞれOR = 1.28、1.88となり、腎負 荷型でより影響力が強いという結果であった。この病型によるORの差異は、統計学的 にも有意であった(それぞれp = 2.4 × 10-5、1.0 × 10-7)。一方、SLC2A9遺伝子の rs3775948は、腎負荷型ではOR = 1.38、腎排泄低下型ではOR = 1.94であり、腎排泄 低下型で有意に影響力が強いという結果であった(p = 2.7 × 10-4)。その一方、他の4 つの遺伝子領域(MYL2-CUX2、GCKR、CNIH-2及びMAP3K11)のSNPsに関しては、
病型による影響力に有意な差はみられなかった(表4)。
さらに、SNPと臨床検査値(FEUAとUUE)との関連についても評価を行った。その結 果、ABCG2遺伝子の2つのSNPs(rs72552713とrs2231142)のリスクアレルを有すると FEUAとUUEを上昇させ、SLC2A9遺伝子のrs3775948のリスクアレルを有するとFEUA
とUUEを低下させることが分かった。
- 17 -
以上の各SNPが痛風の臨床病型に与える影響力を横軸、臨床検査データ(FEUAと
UUE)に与える影響力を縦軸にとり、図示すると図10のようになる。臨床病型とFEUA
に与える影響力の相関係数はr = 0.96(p = 4.8 × 10-4)、臨床病型とUUEに与える影 響力の相関係数はr = 0.96(p = 5.0 × 10-4)であり、いずれも強い相関がみられた。
2-4. 考察
痛風のGWASを実施し、痛風関連遺伝子領域として5つの遺伝子領域(ABCG2、
SLC2A9、MYL2-CUX2、GCKR及びCNIH-2)を同定した。このうちABCG2、SLC2A9 はいずれも尿酸トランスポーターをコードする遺伝子で、過去の欧米のGWASでも痛 風との関連性が同定されていた20, 21。一方、MYL2-CUX2、GCKR、CNIH-2の3つの 遺伝子領域は、GWASによる厳しい基準下に痛風発症との強い関連性を示すものと して世界で初めて見いだされた。
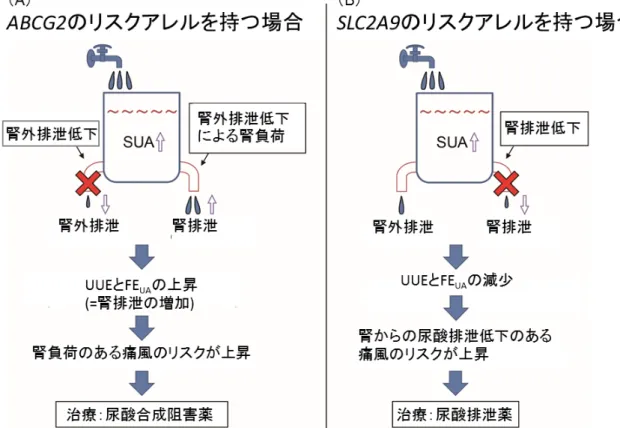
ABCG2は尿酸排泄トランスポーターであり、腎近位尿細管の他、小腸上皮細胞の
管腔側にも局在している。「2-3-2. ABCG2遺伝子の機能低下型多型についての検 討」でも述べたように、ABCG2遺伝子の2つのSNPs(rs72552713とrs2231142)の組み 合わせによってその尿酸輸送機能は0~75%に低下し、痛風の発症頻度に関与して いる5。ABCG2遺伝子の機能低下型多型は、腸管を主とする腎外の尿酸排泄低下を もたらし、腎負荷型の高尿酸血症をきたすことが知られている11, 35。このことは、「2-3-4.
痛風の臨床病型の解析」で、ABCG2遺伝子の2つのSNPsが腎排泄低下型痛風より も腎負荷型痛風への影響が強いという結果に適合する。 またABCG2遺伝子の機能 低下型多型は、若年性痛風発症のリスクでもあり4、さらに「ABCG2の尿酸輸送機能が
25%低下する遺伝子多型」の影響力は、「身長170 cmの人が5.7 kg太る(BMIとして
- 18 -
1.97 kg/m2の上昇)」ことや「毎週1.7 Lのウィスキーを飲む(純エタノール量に換算して
552.1 g)」ことに相当することが示され36、血清尿酸値に対して極めて大きな影響力を
持つことも分かっている。この2つのSNPsによるABCG2の尿酸輸送機能低下は、コ ーカサス人種(白人)やアフリカ系アメリカ人(黒人)では約15%であるのに比し、日本 人では約半数にみられるという人種差があることも特徴である13。
SLC2A9は、主に腎近位尿細管上皮細胞の血管側の細胞膜に位置する尿酸トラン
スポーターであり、尿酸の再吸収に関わっている。SLC2A9遺伝子は腎性低尿酸血症 2型(RHUC2; renal hypouricemia type 2)の原因遺伝子であり、rs121908321
(Arg380Trp)やrs121908322(Arg198Cys)などの変異が尿酸輸送機能を著しく低下さ せることが報告されている8, 37, 38。「2-3-4. 痛風の臨床病型の解析」で、SLC2A9遺伝
子のrs3775948は腎負荷型痛風よりも腎排泄低下型痛風への影響が強いという結果
を示したが、このことは上記の知見と矛盾しない。
GCKRはグルコキナーゼ(GCK; glucokinase)の活性を調整する制御因子である。
GCKは、解糖系の最初の段階であるグルコースからグルコース6-リン酸への変換を触 媒する酵素であり、膵β細胞においてはインスリン分泌におけるグルコースセンサーと して機能しており、糖代謝の重要な役割を果たしている39, 40。GCKRがGCKへ結合す るとGCKの活性は低下し、乖離するとGCKの活性は上昇し解糖系は促進される41。 解糖系で産生されたグルコース6-リン酸からは、ペントースリン酸経路によって核酸の 合成に必要なリボース5-リン酸が産生される。この代謝経路を介して、GCKR遺伝子が 痛風の発症と関連している可能性が考えられる。また、今回のGWASで同定された
SNPであるrs1260326(Leu446Pro)のリスクアレルは、空腹時血糖値を低下させるとい
う報告41, 42がある一方、血清中性脂肪値41, 42や血清尿酸値を上昇させるという報告が
- 19 -
ある20, 43, 44。これらのことから、GCKR遺伝子のrs1260326が糖代謝の個人差を生み出
し、肥満や糖尿病等を介して、痛風の発症に関与している可能性も考えられる。
CNIH-2は神経細胞やグリア細胞に発現し、AMPA
(α-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate)型グルタミン酸受容体に結合 する補助サブユニットとして機能しており、グルタミン酸シグナリングに関連する45, 46。こ
のCNIH-2遺伝子が痛風発症に影響を及ぼす機序は現在のところ不明であるが、
AMPA型グルタミン酸受容体は中枢神経系に広く分布し記憶や学習に関与すること から、CNIH-2遺伝子多型が食生活や飲酒習慣の個人差に影響することで、痛風の発 症に関与している可能性も考えられる。今回同定されたCNIH-2遺伝子のSNP
(rs4073582)の近傍には、PACS1(phosphofurin acidic cluster sorting protein 1)遺伝子 のSNP(rs801733)があり(図8E)、これらは強いLDの関係にある(r2 = 0.97)。この
PACS1遺伝子は肥満との関連が報告されているため47、真の痛風関連遺伝子は
CNIH-2遺伝子ではなくPACS1遺伝子である可能性も考えられる。
MYL2-CUX2遺伝子領域に関しては、今回同定されたrs2188380は遺伝子間領域 のSNPであった(図8C)。このため、このSNPが標識する真の遺伝子を同定するため にさらなる解析を行ったが、その詳細は第3章で述べる。
MAP3K11はMAPキナーゼ(mitogen-activated protein kinase)の一種で、酸化ストレス や炎症性サイトカインなどに反応して様々な細胞の機能を発現させるc-Jun N-terminal
kinase(JNK)の活性化に重要な役割を果たしている48。興味深いことに、尿酸結晶を
単球が貪食すると、このJNK経路が活性化されることが報告されている49。また、今回 同定されたMAP3K11遺伝子のSNP(rs10791821)は単球におけるMAP3K11の発現 レベルにも関連することが分かっている50。このような過去の知見を考慮すると、今回
- 20 -
のGWASではrs10791821はゲノムワイド有意水準には達しなかったが、痛風関連遺
伝子の可能性があると考えられる。
- 21 - 第3章
MYL2-CUX2遺伝子領域における痛風関連遺伝子の同定
3-1. 目的
第2章で、痛風と関連する5か所の遺伝子領域を同定した。この5遺伝子領域のう ち、2つは尿酸トランスポーターとしてよく知られているABCG2遺伝子とSLC2A9遺伝 子であった。また、2つは糖代謝に関わるGCKR遺伝子とグルタミン酸受容体の補助 サブユニットとなるCNIH-2遺伝子で、これらの遺伝子上のSNPであった。一方、残る 1つのSNPは12番染色体のMYL2遺伝子とCUX2遺伝子の遺伝子間領域に存在し ており、真に痛風と関連する遺伝子は別に存在する可能性があったため、fine
mappingを行うことで、その同定を目指した。
3-2. 対象及び方法
3-2-1. 対象
痛風患者1,048人と対照者1,334人を用いて解析を行った(表5)。痛風症例は、京
都工場保健会診療所(京都府)及び両国東口クリニック(東京都)を受診した患者のう ち、米国リウマチ学会の診断基準22をもとに医師が痛風であると確定診断をした日本 人男性症例のみを用いた。対照者は浜松市での健診におけるJ−MICC Study24, 25の 参加者で、痛風・高尿酸血症に罹患していない日本人男性を対象とした。飲酒情報は アンケート用紙にて収集し、1か月に1回以上飲酒する人を飲酒者とした。検体の採
- 22 -
取にあたってはインフォームド・コンセントを行い、同意書を取得した。
3-2-2. SNPの選定
まず、第2章で同定されたMYL2-CUX2遺伝子間領域のSNPであるrs2188380を 中心として10,000,000塩基対(10 Mb)の範囲に存在するSNPを国際HapMap計画 phase III26のデータから抽出したところ、該当するものは8,595 SNPsであった(図11)。
これら8,595 SNPsの中からrs2188380とLDの関係にあるものが真の痛風関連SNP である可能性があるので、LDの指標であるr2値を計算したところ、中等度以上(r2 ≥ 0.3)のLDであるものは45 SNPsであった。次に、この45 SNPsにrs2188380を加えた 46 SNPsのr2値を全ての組み合わせ(1,035通り)で計算した(図12)。その結果、
rs2188380、rs7978484、rs16940688、rs2071629、rs11065783、rs3809297、rs4766566、
rs671、rs2555004の9 SNPsを選択すると、残りの37 SNPsがこの9 SNPsのいずれか とr2 ≥ 0.8という強いLDの関係にあり、37 SNPsをタグ付けられることがわかったため
(表6)、この9個のtag SNPsを遺伝子解析することとした。
3-2-3. 遺伝子解析
DNAは全血から抽出した。rs2188380は第2章のタイピング結果を用い、その他の
8 SNPs(rs7978484、rs16940688、rs2071629、rs11065783、rs3809297、rs4766566、
rs671、rs2555004)は、痛風患者1,048人と対照者1,334人をLightCycler 480(Roche Diagnostics社)を用いたTaqMan法(Life Technologies社)でタイピングした。また一部 の検体については、表7のプライマーと3130xl Genetic Analyzer(Life Technologies 社)を用いて直接シーケンス法でもタイピング結果に間違いがないことを確認した。な
- 23 -
おタイピング成功率は全て95%以上で、Hardy-Weinberg平衡の検定結果が統計学 的に有意(p < 0.05)なSNPも存在しなかった。
3-2-4. 統計学的解析
関連解析にはロジスティック回帰分析を用いた。統計学的有意水準はp < 0.05とし、
多重検定を行う場合はBonferroni法による補正を用いた。すなわち、9 SNPsの解析の 際はp < 5.6 × 10-3(= 0.05/9)、8 SNPsの解析の際はp < 6.3 × 10-3(= 0.05/8)を有意 水準とした。SNPの染色体上の位置は、NCBI human genome reference sequence Build 37の情報に基づいて記した。全ての統計解析はSPSS v.22.0J(IBM社)、
PLINK v.1.0751、R v.3.1.134とそのGenABELパッケージを用いて実施した。
3-3. 結果
3-3-1. 痛風の関連解析
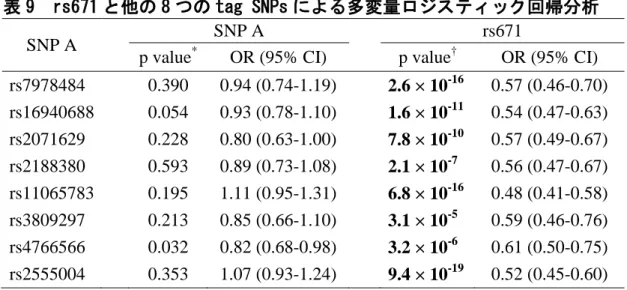
痛風患者1,048人と対照者1,334人における9個のtag SNPのタイピング結果を、
表8に示した。rs2555004を除く8 SNPsが、有意(p < 5.6 × 10-3)に痛風と関連してい た。有意であったSNPsの中では、aldehyde dehydrogenase 2(ALDH2)遺伝子のrs671
(Glu504Lys)のp値が最も低かった(p = 1.7 × 10-18; OR = 0.53; 95% CI: 0.46-0.61, 図13A、表8)。
次に最もp値が低かったrs671でその他の8 SNPsのp値を補正したところ、いずれ もp値は有意水準(p < 6.3 × 10-3)に達しなかった(図13B、表9)。一方、rs671は8 SNPsのいずれで補正をしても痛風との有意な関連が保たれていた(表9)。
- 24 -
以上の結果から、MYL2-CUX2遺伝子領域の真の痛風関連SNPは、ALDH2遺伝
子のrs671であると考えられた。
ALDH2はアルコール代謝で重要な役割を果たす酵素であり(図14)、rs671
(Glu504Lys)はその酵素機能に影響を及ぼす遺伝子多型としてよく知られている52。 ALDH2の酵素活性は、rs671がG/G(Glu/Glu)であるのに比べ、G/A(Glu/Lys)は 6.25%、A/A(Lys/Lys)はほぼ0%になる53。このことを踏まえると、rs671の解析は優性 モデル(Glu/Glu対Glu/Lys及びLys/Lys)で行うのがより適切であると考えられ、優性 モデルで関連解析を実施したところ、p = 2.9 × 10-21、OR = 0.44、95% CI: 0.37-0.52と 表8に示した遺伝子型モデルの解析結果よりもp値が低く(p = 1.7 × 10-18)、仮説に 矛盾しない結果であった。このため以降のrs671の解析は、優性モデルで検討を行っ た。
3-3-2. 飲酒習慣を含めた検討
ALDH2遺伝子のrs671は飲酒習慣に影響することが報告されており52、また飲酒は 痛風のリスクとなることも知られている。そこで「3-3-1. 痛風の関連解析」で見出された
rs671と痛風の関連が、飲酒習慣で補正した場合にどのようになるのか検討した。その
結果、飲酒習慣で補正した場合でもrs671と痛風の関連は統計学的に有意であった
(p = 4.3 × 10-12; OR = 0.52; 95% CI: 0.43-0.63; 表10)。飲酒習慣とrs671の間には 交互作用は認めなかった(p = 0.99)。非飲酒者のみでも解析を行ったところ、rs671と 痛風に有意な関連を認めた(p = 0.019; OR = 0.52; 95% CI: 0.30-0.90)。
- 25 - 3-4. 考察
第2章で痛風関連遺伝子領域として同定されたMYL2-CUX2遺伝子領域の真の痛 風関連SNPは、ALDH2遺伝子のrs671であることが分かった。日本人の約4割は rs671がG/A(Glu/Lys)もしくはA/A(Lys/Lys)である一方、白人や黒人ではこれらの遺 伝子型は極めて稀であり、ほぼすべての人がG/G(Glu/Glu)である54。これまでの白人 や黒人での痛風の遺伝子研究でALDH2遺伝子が同定されなかったのは、このため だと考える。
ALDH2は、アルコール代謝で重要な役割を果たす酵素である。体内に摂取された
アルコールは、まずアルコール脱水素酵素(ADH; alcohol dehydrogenase)によってア セトアルデヒドに代謝され、さらにアセトアルデヒトはアルデヒド脱水素酵素(ALDH;
aldehyde dehydrogenase)によって酢酸へと代謝される(図14)。ADH、ALDHともアイ ソザイムの存在が知られているが、その中で活性が高く、アルコール代謝に強くかかわ っているのは、ADH1BとALDH2である。酢酸はさらに代謝されてacetyl-coenzyme A
(acetyl-CoA)へと変換されるが、この反応の際、ATPからアデノシン一リン酸(AMP;
adenosine monophosphate)が産生される。細胞内のATP/AMP比は一定になるように 制御されているため、AMPの分解は促進され、イノシン、ヒポキサンチン、キサンチン を経て、尿酸へと代謝される55。以上の代謝経路が、飲酒が血清尿酸値を上昇させる と考えられている機序のひとつである。
「3-3-1. 痛風の関連解析」でも述べたように、ALDH2はrs671(Glu504Lys)によって 酵素活性に個人差が生じる。このため、rs671がG/G(Glu/Glu)で酵素活性が高い人 はアセトアルデヒドが早く代謝されるためアルコールに強く、rs671がG/A(Glu/Lys)や A/A(Lys/Lys)の人はアルコールに弱く、飲酒による顔面紅潮や頭痛、嘔気といったフ
- 26 - ラッシング反応を起こしやすい。
以上のことから、今回見出されたALDH2遺伝子のrs671と痛風の関連は、rs671が G/G(Glu/Glu)の人はG/A(Glu/Lys)やA/A(Lys/Lys)の人よりも多くアルコールを摂取 するために尿酸値が高くなりやすく痛風を発症しやすいという、飲酒による影響をみて いる可能性がある。このため飲酒習慣による補正を加えた解析も行ったところ、ORは1 に近づいた(補正なしのOR = 0.44に対し、補正ありのOR = 0.52)もののrs671と痛風 の関連は統計学的に有意であった。また、非飲酒者のみでの解析でも、rs671と痛風 の関連は統計学的に有意であった。今回用いた飲酒習慣に関する情報は、頻度のみ であり量は含まれていない。非飲酒者の解析は少数例であるため飲酒による影響を完 全に否定することは難しいが、ALDH2遺伝子が飲酒以外の機序で痛風発症へ影響 を及ぼしている可能性がある。