愛知県岡崎市の帰化タンポポの雑種化の研究
~遺伝子解析に基づく7年間の追跡調査~
愛知県立岡崎高等学校 2年 等百合佳 拜司さやか
1年 吉井章一郎 小山秀 二村隆征 藤原栞
※本調査はスーパーサイエンス部の 2003 年度~2009 年度の継続調査であり、全体をまと めた部分と私たちの行った部分が含まれる。私たちが行った 2008 年度、2009 年度の研究 については★を付した。
はじめに
岡崎高校の周辺には、ニホンタンポポ、セイヨウタンポポ、アカミタンポポの主に3種類のタン ポポが生育している。これらのうち、ニホンタンポポは「在来種」、セイヨウタンポポ、アカミタ ンポポは「帰化種」と呼ばれる。在来種は平野部の肥沃な土壌に恵まれた環境のよい郊外に多く 生育し、帰化種は都市開発によって土壌が不安定になった都市部を好んで生育しているという、 いわゆる「すみわけ」を行っている。また、形態にも特徴があり、図1に示すように、一般に、外総 苞片に注目すると在来種は反り返っていないが、帰化種は2種とも反り返っている。また、種子 に注目すると、ニホンタンポポ、セイヨウタンポポはクリーム色、アカミタンポポは赤褐色の種 子を持っている。このような生態的、生理的特性の違いや、タンポポの外見での見分けやすさか ら、タンポポは都市化の度合いを測る「環境指標生物」として機能してきた。
しかし近年、ニホンタンポポと帰化タンポポの雑種化が進行している(雑種化をしていない帰 化タンポポをこの研究では「真の帰化タンポポ」と呼んでいる)。これらの交雑によってできた 雑種性帰化タンポポは見た目がセイヨウタンポポやアカミタンポポに似ている。そのため、これ らのタンポポを見た目で見分けることが難しいとされる。また、ニホンタンポポの DNA を持つ 雑種性帰化タンポポは、真の帰化タンポポの生育域である市街地はもちろん、在来種の生育する 都市郊外にも生育できると考えられる。もしそうだとすれば、タンポポは今、環境指標生物とし て機能していないことになる。その真偽を確かめるべく、2003 年より調査を行ってきた。
本年度でタンポポ調査は7年目になり、毎年タンポポの分布を示すタンポポ地図を作ってきた 加えて、その年ごとにテーマを持って、タンポポの形態や生理的特性についての調査を行ってき た。
初めてタンポポ調査を実施した 2003 年度には、調査の結果からタンポポは環境指標生物とし ての機能を果たしていないという結論を導いた。このことが以降のタンポポ調査の礎となった 。 2004年度は雄核単為生殖という、雑種性帰化タンポポ形成時の特殊な受精方法に注目し、2005
1
年度は各種のタンポポの形態の違いに着目し、さまざまな点から各タンポポの比較を行った 。 2006、2007 年度にはニホンタンポポの持つ生理的特性である夏季休眠性について調査した 。 2008年度はタンポポの人工交配実験を行った。
そして今年度(2009 年度)は前年度までの研究結果から、タンポポの近縁同士の交配のしく みや、その交配で出来たタンポポの性質に注目し、形態の観察、人工交配実験、そして発芽実験な どを進めてきた。
外見での判別が可能
外見での判別が可能
外総苞片
外総苞片
図1 タンポポの種類
ニホンタンポポ、セイヨウタンポポ、アカミタンポポはこのような特徴を持っており、見た目で判断する ことができる。しかし近年、帰化タンポポの雑種化により個体の種の識別が困難になった。
生殖方法
1 有性生殖
主に在来種(シロバナタンポポは例外)が備わっているもので、ニホンタンポポは、この有性生 殖を行う。有性生殖では 2 つの細胞の接合によって新たな遺伝子の組み合わせを持つ個体が生 じる。配偶子の形成に際しては減数分裂が行われ、染色体の乗り換えや選択が生じ、配偶子の遺 伝子型は多様なものとなる。その配偶子の組み合わせで生じる接合子はさらに多様な遺伝子の 組み合わせを持つことになる。このような過程を経て、生物の多様性を生み、ひいては種分化や 進化の一因となると考えられている。
2
2 無融合生殖(無性生殖)
これは帰化種やシロバナタンポポ等が行う生殖方法である。これは、簡単にいえば、クローンの 作成である。親の特徴が次世代に保存的に受け継がれ、個体群の単一化をまねき、また、同種の別 クローン由来の個体群と形態的に異なり、生殖的にも隔離されることも多い。
3 単為生殖
これも帰化タンポポに備わっている。帰化種は本来、有性生殖、無性生殖も可能だが、3 倍体や 4倍体なので、有性生殖をするのには不都合なので、受粉をせずに発生をする、つまり、有性生殖 器官を強引に無性生殖的に用いてしまうわけである。作成方法にも、2 種類あり、前者は減数分 裂をしないという方法である。また、後者は一回減数分裂をした後に、融合をするという方法で ある。しかし、後者は厳密に言えば、クローンではなく、クローン個体との近親交配と似た結果と なる。しかし、どのような生殖方法をとっているかはよく分からない。
4 雄核単為生殖による雑種化
これはセイヨウタンポポとニホンタンポポの交雑の際に、精核が卵核と合体することなく、卵 核を排除してしまう受精方法である。これにより、核と細胞質が完全に違うタンポポができてし まう。
5 核の交雑による雑種化
帰化種は減数分裂で n や2 n の花粉ができてしまう。その時に、帰化種と交雑をしてしまう。無 融合生殖を行う帰化種は減数分裂で花粉はでき、それがニホンタンポポに受粉することで雑種 ができると考えられているので、雑種の母親はニホンタンポポとなる(図2)。
図2 雑種化の形成
核の交雑による、一般的な雑種のモデル
3
採取方法
岡崎高校では、2003 年度から 2009 年度までの7年間、毎年春頃に岡崎市のタンポポ調査
(岡崎高校周辺のタンポポの実態調査)を愛知県内の調査(渡邊、1998)に倣って行ってきた 調査の範囲は岡崎高校周辺の2 km 四方の区域内とし、その区域内でタンポポをスーパーサイ エンス部員全員(毎年 20 名程度)で採取した。採取については、2 km 四方の調査範囲を、地形 図を用いて8×8の形64の小枠(メッシュ)に均等に区分し、各メッシュから、一般的な見か け上の判別方法を用いて、ニホンタンポポ、セイヨウタンポポ、アカミタンポポを各々1本ずつ 採取した。尚、帰化タンポポは無融合生殖を行うため、クローン個体の2つ以上の採取を避ける ために、同種のタンポポの採取地点は150 m 以上離れたところで採取するようにした。
乙川
調査範囲
2 km
豊橋市
名古屋市
R1
図3 調査範囲の詳細
岡崎高校周辺の 2km 四方を調査範囲とした
4
遺伝子解析
1 アロザイム酵素多型
見かけ上の判断で採取された3種のタンポポには、雑種性タンポポが含まれている。しかし、外 見上の判断ではそれを区別することができない。そこで、アロザイム酵素多型という方法を用い て、タンポポの遺伝子解析を行った(渡邊、1997)。
酵素には機能が同じでも活性中心以外のアミノ酸配列が少しずつ異なる場合があり、この状態 を酵素多型と呼ぶ。また、1つの遺伝子座に複数の対立遺伝子が存在している状態をアロザイム
(allo=対立、 zyme=酵素)という。アロザイム酵素多型解析は直接 DNA を調べるのではな く、種内に蓄積した酵素の変異を調べる方法で、この方法で種内、種間の識別を行うことができ る。この方法は全てのタンポポから酵素を抽出し電気泳動をする必要があるが、酵素は長時間健 在できず、加えて大量の処理が集中するため、愛知教育大学の渡邊研究室の方々に解析の協力を していただいた。
2 酵素の解析
ニホンタンポポは、GOT(グルタミン酸オキサロ酢酸アミノ基転移酵素)において、対立遺伝 子 a、b または c バンドを持つ。これに対し、真の帰化タンポポは d バンドのみを持つので、在来 種と帰化種(非雑種性)を区別することができる。対立遺伝子 a~d の違いによりゲル上に現れ たバンド a~d を図4に示した。d バンドを持ち、かつ a、b、c バンドを持つものが雑種性帰化タ ンポポである。
また、6 PG(6ホスホグルコン酸脱水素酵素)において、アカミタンポポは特異的なバンドを 持つため、セイヨウタンポポ(アカミタンポポ以外)と区別することができる。6 PG のバンド のパターンも GOT 同様図4に示した。この2種類の酵素の対立遺伝子のバンドを組み合わせる ことで、①ニホンタンポポ、②真のセイヨウタンポポ、③雑種性セイヨウタンポポ、④真のアカミ タンポポ、⑤雑種性アカミタンポポの5種に分類した。(図4)
図 4 アロザイム酵素多型解析 5
タンポポ地図の作成方法
1 タンポポ地図作成プログラム
これは、ある範囲内の帰化種の割合を示すものであり、加重平均法(渡邊ら、1998)を用いて 作成した。元々は渡辺教授からいただいたプログラムを使用していたが、不慣れさから膨大な時 間を必要とした。そのため、本校部 活動内で生徒によりタンポポ地図作成プログラム
「MeshWorker」を開発し、毎年修正しながら使っている。これにより、
・ 色を自在に変化させることができる
・ メッシュの幅、ドット計測のための制限円の半径を自在に変化させることができる。
・ ベクトルデータで結果を出力しているため、拡大や縮小しても誤差が全く出ない 2 作成方法
図 5 の通り、タンポポ調査時に区切った 250 m四方のメッシュではなく、新しく 100m 四方に 区切る。次に、そのマスの色を決定するために正方形の中心から半径 500m の円を作成する。こ のことを「制限円」とする。これは、「そのマスの周りのタンポポが、影響を及ぼしている」とい う考えからである。そして在来種には"0"を、帰化種には"1"という得点をつける。何故、帰化種が
"1"というと、割合をだすためである。中心点から各々のタンポポ(暫定的に順に tnとつける)の 距離をはかり、式「そのマス=得点/距離/1/距離」に代入する。この式のスタイルは、加重平均法を もとにしている。そして出た数値をそれにあった色に、置き換える。
まとめをすると、
・500 mの範囲で平均化を行うので、たとえひとつのメッシュでタンポポが確認できなくても そのメッシュの周囲に個体があれば結果が推定できるという利点がある。
・加重平均法は、距離に応じて得点に重み付けをする。その結果、普通の平均法とは違い、計算地 点に近いタンポポほど計算結果への影響が大きくなり、計算結果もよりはっきりする。
図 5 タンポポ地図の作成方法
加重平均法によってタンポポ指数を求め、色分けをした。
タンポポ地図
岡崎高校周辺のタンポポ調査と遺伝子解析の結果から、MeshWorker を用いて(A)見かけの タンポポ地図、(B)真のタンポポ地図、(C)雑種のタンポポ地図の3つの地図を毎年作成し た。この地図では、東西に走る名鉄本線よりも北側の地域が市街地で、本線よりも南側の地域が 郊外となっている。平たくいえば、地図の右上が市街地で、左下が郊外である。
A 見かけのタンポポ地図
この地図は、各年度に採取した全てのタンポポを対象に作成した地図で、採取時同様見かけだ けでのタンポポの判別に基づいて作られている。各ポイントの色はその地域周辺に於ける全タ ンポポにおける帰化タンポポの割合を示している。したがって、緑色に近ければニホンタンポポ の割合が多く、赤色に近ければ帰化タンポポの割合が多いことを示す。
2003年度から 2009 年度の7年間の分布地図を図 A に表した。2003、2004 年度の地図は右 上の辺りが赤く、左下の辺りが緑に近いことから、市街地に帰化タンポポ、郊外にニホンタンポ ポが多いという、いわゆる「すみわけ」が明確に現れている。しかし、翌年2005年度から 2009年度にかけて、タンポポ地図は全体的にオレンジ色が多い。つまり、ニホンタンポポの生息 域に帰化タンポポが多く見られることから、都市化が進行していると読み取ることができる。
図 A 見かけのタンポポ地図
2003年度 2004年度
2005年度 2006年度
2007年度 2008年度
Mesh Workerで計算した、色合い
2009年度 ★
B 真のタンポポ地図
この地図は遺伝子解析の結果に基づいて雑種性帰化タンポポであるものを除き、ニホンタンポ ポ、真のセイヨウタンポポ、真のアカミタンポポの3種類のタンポポを対象に作成された地図で ある。つまり、岡崎高校周辺の都市化の度合いを正確に測ることができるのである。各ポイント 周辺の対象の全タンポポにおける真の帰化タンポポの割合を色で表し、緑色に近ければニホン タンポポの割合が多く、赤色に近ければ真の帰化タンポポの割合が多いことを表している。しか し、調査の結果から、真のセイヨウタンポポは岡崎高校周辺にはほとんどないことが分かった。 そのため、ここでいう真の帰化タンポポのほとんどは真のアカミタンポポを示している。また、 アカミタンポポも年々増加傾向にある。
7年間のタンポポ地図を図 B に示した。2003、2004 年度は先ほどの見かけのタンポポ地図と 同様に、市街地に帰化タンポポが多く、郊外にニホンタンポポが多いことが分かる。しかし、それ 以降の5年間は、緑色の部分の分布にばらつきがある。特に、2006 年、2007 年度は全体的に緑 色が多く、ニホンタンポポが増加したように見える。だがこれは、雑種性アカミタンポポの増加 により真のアカミタンポポの数が減り、それに伴い相対的にニホンタンポポの割合が多くなっ たためである。これらから真の帰化タンポポは増加していないと読み取れるので、都市化は進行 していないといえる。しかし、この結論は先ほどの見かけのタンポポ地図の考察と矛盾してしま う。
図 B 真のタンポポ地図
2003年度 2004年度
2005年度 2006年度
2007年度 2008年度
Mesh Workerで計算した、色合い
2009年度 ★
C 雑種のタンポポ地図
この地図はニホンタンポポを除く真のセイヨウタンポポ、雑種性セイヨウタンポポ、真のアカ ミタンポポ、雑種性アカミタンポポの4種類のタンポポを対象に作成した。各ポイント周辺の対 象の全タンポポにおける真の帰化タンポポの割合を色で表し、黄色に近ければ雑種性帰化タン ポポが多く、赤色に近ければ真の帰化タンポポが多いことを表している。この地図においても、 真のタンポポ地図同様、真の帰化タンポポのほとんどは真のアカミタンポポが占めている。
7年間のタンポポ地図を図 C に表した。2007 年度まで年々地図のほとんどを黄色が占めてい る。このことから 2007 年度までは雑種性帰化タンポポ、特に雑種性アカミタンポポが増加して いることがわかる。ところが、2008、2009 年度の地図は全体的にオレンジ色をしている。これ は、雑種性アカミタンポポの数が減少し、真のアカミタンポポの数が増加したためと考えられる
図 C 雑種のタンポポ地図
2003年度 2004年度
2005年度 2006年度
2007年度 2008年度
Mesh Workerで計算した、色合い
2009年度 ★
以上3種類のタンポポ地図から、ニホンタンポポの生育する郊外にアカミタンポポが侵入し、 雑種化することで郊外に生育できるようになり、その結果、雑種と真のアカミタンポポの見分け がつかなくなり、種の特定が困難になったことで、見かけのタンポポ地図と真のタンポポ地図に 差が生じたと考えられる。よって、見た目で判断して作られたタンポポ地図はその意義を失い、 タンポポは環境指標生物として機能していないといえるだろう。
夏季休眠性
ニホンタンポポの生理的特性に夏季休眠性というものがある。夏季休眠性には、葉の夏季休眠 性と種子の夏季休眠性の2種類がある。タンポポ地図の考察から、雑種性帰化タンポポは雑種化 することで夏季休眠性を取り込みニホンタンポポの生育域に侵入してきたのではないかと考え 、 2006年度、2007 年度にそれぞれ、葉、種子の夏季休眠性についての実験を行った。
1 葉の夏季休眠性
葉の夏季休眠性とは、夏季に背の低いタンポポが他の雑草の陰になり、充分に光合成を行うこ とができなくなるため、葉を落とし根だけでひと夏を過ごす性質である。
2006年度に校内で生育しているタンポポから無作為に選んだ個体を対象に、葉の枚数の変化 を調べた。その結果をグラフに表したのが図6である。縦軸が葉の枚数を表し、横軸が日付を表 している。また、色1つ1つが1個の個体である。
ニホンタンポポのグラフを見てみると、どの個体も明らかに葉の枚数が減少していることがで きる。したがってニホンタンポポには夏季休眠性が備わっているといえる。対して真のアカミタ ンポポのグラフでは、葉の変化が多様で、一連の変化がなく、またどの個体も先ほどのニホンタ ンポポのような大きな変化が見られない。したがって、真のアカミタンポポには夏季休眠性が備 わっていないといえる。
そして、雑種性アカミタンポポのグラフに注目すると、葉の枚数が0になるまではいたらない が、ニホンタンポポと同じ傾向を持った個体があることが分かる。したがって、雑種性アカミタ ンポポの一部の個体には、夏季休眠性が備わっているのではないかと考えられる。
図 6 葉の枚数の変化の調査結果
2 種子の夏季休眠性
種子の夏季休眠性は、夏に発芽を抑制し、涼し くなった秋に発芽を再び促進させる性質であ
る。
2007年度に葉の夏季休眠性同様、校内で生育しているタンポポを無作為に選出しそこから種 子を取り出し、16℃(図 7 の青い線)と24℃(図 7 の赤い線)の環境に種子を等分し、それぞれ 発芽の様子を観察した。その結果を図7に示した。縦軸が分けた種子の発芽率、横軸が経過日数 を表している。
ニホンタンポポのグラフを見ると、16℃の温度下では半数以上の種子が発芽しているが、2 4℃の温度下ではほとんど発芽していない。したがって、高温化の環境での発芽が抑制されてい ることが分かり、夏季休眠性が備わっているといえる。真のアカミタンポポのグラフを見ると、 どちらの環境でもほとんどの種子が発芽していることが分かり、ニホンタンポポと大きく差が 生じていることが読み取れる。したがって、真のアカミタンポポには夏季休眠性が備わっていな いといえる。
そして2つの雑種性アカミタンポポのグラフに注目すると、片方はニホンタンポポに、もう片 方は真のアカミタンポポに近い特徴を持っていることが分かる。このことから、夏季休眠性が備 わっている個体と、備わっていない個体の両者が存在していることがいえる。
図7種子の発芽率の調査結果
これらの結果から、雑種性アカミタンポポの一部にはニ ホンタンポポの持つ夏季休眠性が備わっているのではな いかと考えられる。雑種化することにより、アカミタンポ ポは郊外に適応できるようになったといえるのである。
形態調査 ★
2005年度、2009 年度に花粉、葉の形状、外総苞片などの形態調査をした。その中で、最も種ご との違いがはっきりした、「外総苞片の反り返りの角度と先端の突起の厚み」に注目をした。
図8 形態調査をまとめた、分布図
図8には各個体の突起の厚さと外総苞片の角度を示した。
真のニホンタンポポは、ほとんどが外総苞片の反り返りが少なく、突起は分厚く、真の帰化タン ポポは外総苞片の反り返りが大きく、突起は薄い、あるいは全くないものが多い。
雑種性セイヨウタンポポは様々な形態をとる。それに対して雑種性アカミタンポポは分布が基 本的には真のアカミタンポポと似ているが、雑種性アカミタンポポの中にはこの平均的な分布 に沿わないものがあった。
このタンポポは、次の真のタンポポ地図(図9)では、赤い点で表した位置で発見された個体 である。この位置は、ニホンタンポポの割合が多く、雑種化したあともバッククロス、つまりもう 一度ニホンタンポポと交配したことによりニホンタンポポの遺伝子が濃くなったのではないか と考えられる。
図9 バッククロスの可能性のある個体を赤丸で示した。
人工交配実験 ★
1 交配実験、発芽実験
2008年度と 2009 年度には人工交配による次代のタンポポ作りを行った。理由は、野外の個
体にはばらつきが多く比較に限界を感じたことと、生殖方法自体にも興味があったためである。 四月、五月・・・蕾のあるタンポポを約50個体採集、室内で隔離して栽培し、開花してから花同 士を1,2回こすり合わせて人工授粉させて種子を得た。
六月・・・種子の結実率を調べ、結実した種子を使って発芽実験を行う。ピートモスという土に種 を蒔き、水のみを与える。土中温度は16~22℃で、pHは約6である。また、発芽実験を行う 際の、結実したかの判断は、肉眼で実のふくらみ具合を判別して行った。写真(図 10)の左の実 は結実していると判断し、右の実は結実していないと判断した。
図 10 判別の例
七月・・・発芽実験で発芽した個体とその両親を、DNAを用いた遺伝子解析にかけ、親子鑑定を 行った。
以上の手順で行った結果が図 11,12 のグラフである。
図 11 アカミタンポポ同士の交配結果
図 11 は、アカミタンポポ同士の交配の結果である。アカミタンポポは交配しなくても種子が 出来るが、比較のために行った。40個の頭花から、総種子数は1881個を採取した。この種子 の内、結実しているものといないものの割合を左の円グラフに示し、結実した種子の内、発芽し たものとしなかったものの割合を右の円グラフに示した。個々の頭花の結実率、発芽率は円グラ フの下に最大値、最小値、そして標準偏差を併記すると、結実率、発芽率はこのようになった。 ここで注目するのは、単為生殖をするアカミタンポポの結実率が100%ではないことである 単為生殖では花粉が付いたか付かなかったかに拘わらず、全ての種子が結実するはずであるか ら、アカミタンポポの中には単為生殖を行わないものがあるのかもしれない。よって有性生殖を する可能性、つまり帰化種が母親となり、雑種性タンポポを作る可能性があるかもしれない。
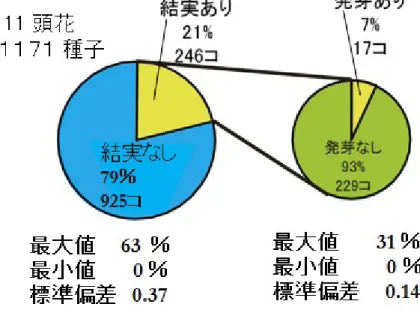
図 12 ニホンタンポポ(♀)と真のアカミタンポポ(♂)の交配結果
次に、ニホンタンポポ(♀)と真のアカミタンポポ(♂)の交配実験の結果を図 12 に示す。こ の実験に使用したニホンタンポポは、頭花の数は11個体、総種子数は1171個、結実率、発芽 率はグラフの下に示した。結実率も発芽率も非常に低いことから、ニホンタンポポからは、なか なか次代が生まれないことが分かる。
なお、今回の人工交配実験で作出した次代は、PCRと制限酵素を用いた、ITS領域のDNA 多型から判別をした。また、マイクロサテライトの DNA 分析からも確認をした。
2 実験方法
PCR(ポリメラーゼ連鎖反応)
まず、増幅対象の DNA 領域の両端の塩基配列を決定し、抽出した鋳型 DNA、プライマー、DNA ポリメラーゼと DNA 合成の基質である dNTP、そして酵素が働く環境を作るためのバッファー 溶液を混合し、サーマルサイクラーで自動増幅した。
※マイクロサテライト
反復配列で、とくに数塩基の単位配列の繰り返しである。中立で種系統内に固定されている。同 種内にも多様性があり、親から子へ染色体で伝わるため DNA 鑑定のための遺伝マーカーとして 利用されている。
図 13 特定のマイクロサテライトを増幅し、電気泳動によって可視化したもの
図 13 は自家不和合性のあるニホンタンポポで、あかみたんぽぽ(♂)と受精し、母親のニホン タンポポと同じ遺伝子を持つニホンタンポポが生まれたときの写真である。実は、ニホンタンポ ポには、他の個体の花粉が付着すると自家不和合性が崩れるという、メントール効果がある。ア カミタンポポの花粉が付着したことにより、メントール効果が起こり、ニホンタンポポ自身の花 粉と受精してしまったのかもしれない。(図 14 を参照)
図 14 自家不和合性が崩れる時の図
考察
タンポポ地図からは、ニホンタンポポと帰化タンポポが交雑し、雑種性タンポポが生じること で帰化タンポポの遺伝子がニホンタンポポの生育域に進入していることがわかる。また、ニホン タンポポと帰化タンポポは、ある程度の傾向を持ちながらもその分布域を変化させている。この ことは、多年生ではあるがタンポポが更新していることを示す。
雑種化することでタンポポが中間的な形態や生理的特性を備えることは、環境指標生物として の意義を失わせていると考えられる。
夏季休眠性の調査から、一部の雑種性アカミタンポポはニホンタンポポの持つ夏季休眠性を取 り入れ、ニホンタンポポの生育する郊外に進入できたと考えられる。また、形態の観察の結果か らは、外総苞片の角度と突起の厚さではニホンタンポポと帰化タンポポに差があり、その雑種は 中間的な性質を持つことが分かった。外総苞片が反り返り、突起の厚さが厚い雑種性タンポポは 雑種がニホンタンポポと戻し交配をしたものではないかと考えた。
本年度の人工交配実験から、無融合生殖をするはずのアカミタンポポの結実率は 100%ではな かった。アカミタンポポの中には、花粉を受け付け受精による生殖を行うものがあるのかも知れ ないと考えた。また、人工授粉によってニホンタンポポの自家不和合性が崩れることが確認出来 た。そして、雑種性タンポポ作出の可能性は非常に低いということが分かった。
展望
今後の展望としては、この人工交配実験を続け雑種性タンポポを作出し、形態や生理的特性が、 親のタンポポとどの様に違うのかを調べ、帰化タンポポの雑種化による分布域の拡大に関わる ことまで調べていきたいと思っている。また、先述のバッククロスについても調査していきたい と思っている。
謝辞
最後に、毎年タンポポ調査のご指導をいただいている、愛知教育大学の渡辺幹男教授、に謝意を 表します。
引用文献
渡邊幹男 1997 酵素多型で判別した雑種タンポポ―強奪種としてのタンポポ― 植物学学 会誌 21:43:47
渡邊幹男・榊原洋子・櫛田敏宏・神崎護・市川雅美・丸山由香里・内藤敬江・溝口忍・山倉拓 夫・芹沢俊介 1998. 加重平均法によるタンポポ地図の作成 植物学学会誌 15:147-150