DOI: http://dx.doi.org/10.14947/psychono.35.10
“違う”視えから見える世界
―比較錯視研究の意義―
中 村 哲 之
東洋学園大学Visual world revealed by “different” perception:
The importance of comparative research on visual illusions
Noriyuki Nakamura
Toyo Gakuen University
Research on visual illusions in different animal species as compared to human beings is fruitful to know the adaptive significance of the illusory phenomena. Comparative studies on visual illusions suggest the precedence of
global features in adult human visual perception and the precedence of local features in non-human animal visual
perception. Similar results are also reported by a developmental study (Doherty, Campbell, Tsuji, & Philips, 2010), suggesting the precedence of local features in human child visual perception. The importance of the comparative work on visual illusions in two perspectives (cross-species and developmental) is discussed.
Keywords: visual illusions, comparative cognition, cognitive development
1.知覚発達研究と比較知覚研究との 関係について 日本基礎心理学会第 34回大会のシンポジウム2:「錯 視と発達」のなかで,ヒトを含めた動物の錯視を種間比 較する立場から話題提供をおこなった。筆者の専門は動 物の認知発達研究ではなく,少なくともこれまでの研究 対象は,主にオトナの鳥類であった。なぜ,「発達」が テーマのシンポジウムに動物研究なのか。発達研究と動 物研究との間に接点を見出すことが困難な方もいるかも しれないので,はじめに両研究の親和性の高さについて 触れておきたい。 知覚発達研究と比較知覚研究との間には,少なくとも 2つの類似点が存在すると筆者は考える。1つ目は「研 究手法の類似」である。一般的に広義の発達心理学研究 では「“ゆりかご前”から“墓場前”まで」,すなわち胎 児期から老齢期に至るまでの心の変化を取り扱うとされ るが,多くの研究は乳児期から幼児期あたりの心の変化 を対象とするものである。特に知覚の発達は新生児期か ら乳児期にかけて著しい変化があるため,当該の時期を 対象とした研究の重要性が高い。しかし,この時期の大 半は言語獲得前であることから,ヒトのオトナを対象と した言語教示を前提とする研究手法を適用することはで きない。そのため,さまざまな研究手法に工夫が必要と なる(詳しくは,同号の山口,2016; 金沢,2016を参 照)。同様のことは,動物の知覚研究にもあてはまる。 ヒトを含めた動物は適切な環境認識を実現するために, 各自の生活様式に合った知覚を進化の過程のなかで獲得 してきたと考えられる。つまり,動物種間で必ずしも同 じとは限らない知覚の多様性を明らかにすることは,知 覚の適応的意義を検討する点において重要性が高い。し かし,基本的に動物もヒトの言語を用いることはないた め,ヒトのオトナを対象とした研究手法を直接適用する ことはできず,これまでに様々な研究手法が開発されて きた。ヒトの新生児・乳児とヒト以外の動物では身体能 力や体サイズの違いなどもあるため,全く同じ研究手法 を両者に適用できるわけではないが,互いに参考になる 視点や考え方があると思われる。 2つ目は「ヒトのオトナの知覚との比較」が根底に存 在するという類似点である。多くの知覚発達研究では, Copyright 2016. The Japanese Psychonomic Society. All rights reserved. Correspondence address: Faculty of Human Sciences, Toyo
Gakuen University, 1–26–3 Hongo, Bunkyo-ku, Tokyo 113– 0033, Japan. E-mail: [email protected]
乳幼児が視ている世界がどのような過程を経てヒトのオ トナの知覚世界へと変化していくのかといった点に興味 があると思われる。あるいは,養育者が適切な対応を取 るために自身の知覚世界との違いを知ることが重要だと いった視点もあるかもしれない。いずれにしても,乳幼 児の知覚世界それ自体だけに興味の対象がとどまってい るわけではない。比較知覚研究についてもさまざまな立 場の研究者がいるが,少なくとも筆者の興味は,研究対 象としている動物の知覚世界自体だけでなく,ヒトのオ トナとの比較において何がわかるのかといった点にもあ る。このように考えると,比較知覚研究で明らかになっ たことを,知覚発達研究を進めていく際のヒントとして 活用していく可能性は十分にあるといえるのではないだ ろうか。逆に,知覚発達研究における知見を比較知覚研 究に活かすという方向性もあるだろう。 以上のことから,「錯視と発達」というテーマのなか で,比較錯視研究の成果を報告し議論することは,知覚 発達研究と比較知覚研究双方の発展において有益である といえる。 2.比較錯視研究の方法例 ヒトのオトナを対象とした実験方法にも多くの種類が あるのと同様に,言語を用いた教示ができない場合の実 験方法にも幾つか方法があるが,今回の講演では,ハト を被験体としたミュラー・リヤー錯視図形の知覚研究 (Nakamura, Fujita, Ushitani, & Miyata, 2006)を例に,比較
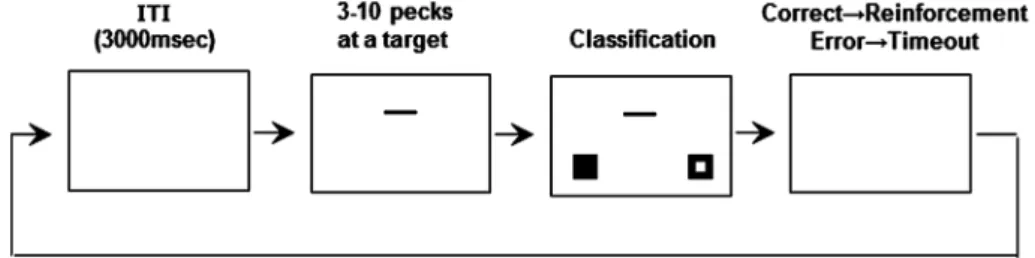
錯視研究方法を説明した。 この研究では,「モニター画面上に呈示される線分に つつき反応すること」「 呈示装置から を食べること」 を既に学習したハトに対し,2選択課題によって線分の 長さを報告させることを初めに訓練した(Figure 1)。 3秒の試行間間隔の後に,パソコンで制御した液晶モニ ター画面の中央に,30または60ピクセル(100ピクセル は29.7 mm)の水平線分2種類のうちのどちらか一方を 呈示した。モニター画面に呈示された線分に対して,被 験体がつつき反応を3∼10回(個体の学習進度を確認し ながら反応回数を操作した)おこなうと,線分の左下と 右下に正方形のアイコン(■と□)が呈示された。ある 個体は,短い線分が呈示される試行では■,長い線分が 呈示される試行では□に対して1回のつつき反応があれ ば正答とみなし, もしくは光(二次性強化子)による 強化を与えた。逆側の正方形アイコンにつつき反応が あった場合には,強化は与えず,タイムアウトを課すこ とで軽い罰を与えた。線分長と正解位置の関係性という 要因で結果が説明されてしまう可能性を避けるために, 別の個体に対しては,線分長と正解位置の関係を逆にす ることでカウンターバランスをとった。なお,最初の訓 練段階で矢羽を呈示せずに線分のみを呈示したのは,線 分の長さ以外の手がかりを用いて被験体がこの課題を遂 行してしまう危険性を避けるためであった。正答率 90%の学習基準達成の後,線分の長さを6種類(30, 36, 42, 48, 54, 60ピクセル)とし,短い方から3本を“短い”, 長い方から3本を“長い”と報告する2選択課題に移行 した。線分長に基づいた高精度な弁別ができていること をやさしい条件で確認しつつ,正答率が高くなり過ぎな い難しい条件を加えることで,後のテスト段階におい て,同じ線分長でも矢羽向きの違いによって「長い」と 反応する率が変化する余地を残すことが必要なためで あった。正答率 80%の基準に到達した段階で,線分長 を報告できるようになったとみなした。 次に,水平線分に加え矢羽をモニター画面上に呈示す る段階へと移行した。ただし,矢羽を含めた線分長を報 告しないようにさせるため,最初は線分長判断への影響 が少ないと考えられる線分の上下に矢羽を呈示した。突 然現れた矢羽によって,被験体の反応が乱れることを避 けるため,最初は薄い灰色の矢羽を呈示し,その後矢羽 の色を黒にした。矢羽なしのときと同様に,線分長の2 選択課題をおこなった。 最終訓練段階では,同方向を向いた矢羽を線分の両端 に呈示した(Figure 2(A): Judd図形)。この図形は,少な くともヒトでは長さの錯視は生じないことが報告されてい る(e.g., Glazebrook, Dhillon, Keetch, Lyons, Amazeen,
Weeks, & Elliott, 2005)。さらに本研究でも,ハトの実験 で用いた図形をヒトでテストしたところ,この図形に対 して長さの錯視はほとんど生じていないことを確認して いる。テストでは,Judd 図形を呈示する試行とミュ ラー・リヤー錯視図形を呈示する試行を3 : 1の割合で実 施した。ミュラー・リヤー錯視図形を呈示する試行で は,実際に知覚された線分長を答えてもらうため,長・ 短いずれの反応に対しても正答とした。訓練同様に分化 強化でテストしてしまうと,例えば,42ピクセルの水 平線分を持つ内向矢羽(><)図形が呈示された際,も しハトでもヒトと同方向に錯視が生じ,水平線分長が 50ピクセル程度に知覚されたとすれば,ハトは「長い」 と反応する可能性が高い。しかし,物理的に42ピクセ ルの水平線分であるからという理由で罰を与えてしまう と,それ以降の試行で行動の調整が生じ,知覚された線 分長に基づいた反応が期待できなくなる危険性がある。 3.錯視辞典に掲載されていない 錯視 の発見 上述した方法で実験をおこなった結果,ミュラー・リ ヤー錯視図形に対しては,ハトでもヒトと同方向の錯視 が生じていることが示唆された(Figure 2(B)で下の図 の水平線分のほうが長く見える)。ハトが水平線分の長 さ以外の手がかりを用いていた可能性(例えば,矢羽を 含めた図形全体の長さ)についても検討したが,そうし Figure 2. Examples of geometric illusory figures. (A) Judd figures. (B) Müller–Lyer illusory figures. (C) Ebbinghaus–
Titchener illusory circles. (D) Zöllner illusory figures. (E) Reversed Ebbinghaus–Titchener illusory circles. (F) Various possible perceptions of the pattern at the top. (G) Various possible perceptions of the pattern at the top. (H) Navon figures.
た手がかりでは本実験の結果を説明することはできな かった。さらに先行研究から,オマキザル(Suganuma, Pessoa, Monge-Fuentes, Castro, & Tavares, 2007),アカゲザ ル(Tudusciuc & Nieder, 2010),ヨウム(Pepperberg, Vici-nay, & Cavanagh, 2008),セキセイインコ(Watanabe, Nishi-moto, Fujita, & Ishida, 2014),ヒヨコ(Winslow, 1933),ハ エ(Geiger & Poggio, 1975)でも,ミュラー・リヤー錯視 が生じていると報告されている(ただし,被験体が線分 長以外の手がかりを用いていた可能性が検討されていな い実験報告も含む)。もしこれらの研究結果が正しいも のであるとするならば,この錯視は多くの動物で同じよ うに生じている現象である可能性が高いと結論すること ができる。 しかし,他の錯視図形についても同様の方法で研究を 進めていくうちに,全ての錯視が動物間で共通に生じて いるわけではないことがわかった。例えば,エビングハ ウス・ティチェナー錯視図(Figure 2(C))に対して,左 右それぞれの図形の中心にある円の大きさを比べた場 合,ヒトには小さな円に囲まれた左側の中心円のほうが 大きく見える。Murayama, Usui, Takeda, Kato, & Maejima (2012)によれば,ハンドウイルカ(Tursiops truncatus) でも同様の錯視が生じていることが示唆されている。と ころが,Paron & Fagot (2007)によれば,ヒヒには左右 の円は同じ大きさに見えていると報告されている。さら に我々の研究から,ハトやニワトリでは,大きな円に囲 まれた左側の円のほうが大きく見えていること,つま り,ヒトとは逆方向の錯視が生じていることが示唆され た(Nakamura, Watanabe, & Fujita, 2008, 2014)。
錯視に関する膨大な研究をまとめたハンドブックや著 書(e.g., Coren & Girgus, 1978; 後藤・田中,2005; 今井, 1984; Robinson, 1998)には,様々な錯視現象に関する解 説が分かりやすく説明されている。例えば,エビングハ ウス・ティチェナー錯視の項目を参照すると「中くらい の円が複数の大きい円に囲まれるとより小さく見え,複 数の小さい円に囲まれるとより大きく見える(後藤・田 中,2005, p. 59)」といった具合である。しかし,錯視辞 典に書かれているこうした解説は「ヒトという種(さら に厳密にいえば,健常なヒトのオトナ)において生じる エビングハウス・ティチェナー錯視」に関するものであ り,この世に存在する数多くの“エビングハウス・ティ チェナー錯視”のごく一部に過ぎないということが,先 に述べたこの錯視の多様性の事例から分かる。ツェル ナー錯視図(Figure 2(D))に対しても,アカゲザルでは ヒトと同方向の錯視が生じるという報告がある(Agrillo, Parrish, & Beran, 2014)一方で,ハトやニワトリでは錯視
が生じる方向がヒトと逆であるということが報告されて いる(Watanabe, Nakamura, & Fujita, 2011, 2013)。さらに 他の錯視図についても,こうした多様性が生じている可 能性は十分に考えられる。 過去の動物を被験体とした錯視研究において,錯視と はヒトの錯視を意味することがほとんどであった。つま り,「ヒト以外の動物においても,(ヒトで生じる)錯視 が生じているのかを確かめる」ことが研究目的となって おり,それ以外の結果が生じることには興味がない,あ るいは想定すらされていない。そして,ヒトの錯視から 予想されるものとは異なる結果が確認された場合,それ は実験上の何らかの不備による結果として扱われること さえあった(例えば,Dücker, 1966)。こうした歴史的経 緯からも,錯視辞典に掲載されていない“錯視”は,“エ ビングハウス・ティチェナー錯視”や“ツェルナー錯視” 以外にもまだまだ数多く存在しているのかもしれない。 4.種間で異なる結果が意味するもの 錯視に多様性が存在するのはなぜだろうか。その理由 は,視覚のメカニズムを考えれば説明がつく。「視る」 という現象は,「光情報が眼球の奥側にある網膜で光信 号に変換され,脳内で情報処理結果として生じる主観的 な体験」であり,脳が生み出した“虚構(フィクショ ン)”である。「視る」ことが主観的な経験であることの 事例としては,モンシロチョウの体色が例として挙げら れる。紫外線を色として認識しない我々にとってはオス とメスは同じ色に見えるが,紫外線を色として認識する 動物にとっては全く違って見えているという(小原, 2003)。すべての動物が“ありのままの世界”を認識し ていればこうした問題は生じないのだが,残念ながら, 動物の神経系の情報処理速度では外界に存在する膨大な 情報を全て処理することはできない。すべての情報を処 理することなく外界を認識するために,ヒトを含めた動 物は,環境内の膨大な情報の中から自身が生活する際に 必要な情報だけを選択的に処理する戦略をとっているの である(例えば,ヒトは紫外線を色として認識するため の情報処理をしない)。動物が外界の情報の一部だけを 処理することで環境認識を実現している以上,外界の物 体が持つ物理的性質と視覚システムによって復元された “物体”がもつ知覚的な性質の間にはズレが生じている。 視覚システムの優れた復元能力のおかげで,日常生活に おいてそうしたズレを実感する機会は少ないが,ある特 定の状況下においては,そのズレが顕著に現れることが ある。それが錯視と呼ばれる現象である。錯視は,当該 の生体が有しているものの見方そのものの表れであり,
「適応的かつ合理的な視覚機能の副産物」といえる。動 物種や発達段階の違いによって生活環境から取り出すべ き情報に多様性があるならば,その情報を基にして脳内 で再構成される視覚世界やその表れである錯視にも多様 性が存在することになる。 このように考えると,比較錯視研究によって種間で異 なる結果が得られた場合,そこにはそれぞれの動物が有 する視覚世界の特徴を明らかにするためのヒントがある といえる。ここでは,ヒトにとって重要な情報とは何 か,他の動物と比べたときに,ヒトが見ている“幻想世 界”にはどのような特徴があるのかについて考察してい くことにする。 先に述べたエビングハウス・ティチェナー錯視につい て,ヒヒでは錯視が生じないという結果については,中 心円の大きさ知覚において周囲円が影響を与えなかっ た,すなわち,彼らが中心円に重きを置いた情報処理, すなわち「局所志向的な情報処理」をしていることを示 唆する結果であったと解釈できる。ハトやニワトリが示 した,ヒトと錯視方向が逆になるという結果は,ヒトで もある状況下では同様の逆錯視が生じることが報告され ている(盛永,1956; Weintraub, 1979)。Figure 2(E)に示 した図に関して,中央に位置する円の大きさを比べた場 合,右側の円が大きく見えるというヒトが多いのだが, 物理的には左右ともに同じ大きさである。Figure 2(E) 右の図は,左の図内で中心円を取り囲んでいる円の中心 から近い部分 1/4のみだけを残し,残りの3/4を削った ものである。つまりこの錯視現象は,中心に位置する円 の近傍のみしか知覚できない状況を強制的に作り出す と,ヒトでも錯視の生じる方向が逆になりうることを示 している。つまり,ハトやニワトリも,処理される空間 範囲に違いはあるものの,ヒヒと同様に中心の円に重き を置いた処理をしていると考えられる。こうした処理に 対し,ヒトのエビングハウス・ティチェナー錯視は,図 形全体を知覚的に体制化することによって生じる現象で あるという報告がある(Roberts, Harris, & Yates, 2005)。 すなわち,Figure 2(C)の図に対して,ヒトは「図形全 体との関係性」のなかで中心の円を見るといった「全体 志向的な情報処理傾向」が他の動物に比べて強いことが 分かる。 このような情報処理の違いがエビングハウス・ティ チェナー錯視の多様性を生み出しているという仮説は, 複数種間で同方向の錯視が生じていることが報告されて いるミュラー・リヤー錯視図の知覚に関する結果に対し ても矛盾なく適用できる。ヒトに水平線分を等分するこ とを求めた実験結果から,この錯視は図形全体に働く相 互作用ではなく,図形部分間(線分と矢羽の接点付近) の局所的な相互作用によって生じていることが示唆され ている(Post, Welch, & Caufield, 1998)。もしこの仮説が ヒト以外の動物に対しても当てはまるものであるなら ば,ヒトと比べて局所志向的な情報処理傾向の強い種 (例えばハト)においても,ヒトと同方向のミュラー・ リヤー錯視が生じて不思議ではない。 「図形全体との関係性のなかでターゲットを処理する」 というヒトの視覚情報処理は,幾何学的錯視図以外に も,さまざまな図形認識において確認されている。 Figure 2(F)上の図に対して,多くのヒトでは,黒色正 方形の背後に45度回転した灰色の正方形の一部が“隠 れている”といったアモーダル補間が生じる(Figure 2 (F)左下)。しかし,ヒトが灰色“正方形”と認識した 図形について,ハトやニワトリはアモーダル補間をせ ず,切り欠け図形として認識する(Figure 2(F)の右下: Fujita & Ushitani, 2005; Nakamura, Watanabe, Betsuyaku, & Fujita, 2010)。類似の現象は,Figure 2(G)の図でも報告 されている。ヒトは「1本」の長い黒棒が灰色の帯図形
の“背後”を左右に動いていると認識するが,ハトは「2本」
の短い黒棒が動いていると認識する(Ushitani, Fujita, & Yamanaka, 2001)。Figure 2(F) や (G) の図形に対して, 見えない部分は手前の物体に“遮 ”されていると解釈 し,その“遮 ”部分を補うことによって図形全体を一 つのまとまりとして処理するのがヒト,見えない部分は 存在しないといった認識をするのがハトやニワトリであ るといえる。Figure 2(H)のような階層構造を持つ図形 認識においても,種間で異なる情報処理がされている。 ヒトは,局所(大きなSを構成する小さなHもしくは大 きなHを構成する小さなS)よりも全体を重視する傾向 が強いため,左からSHと答える(Navon, 1977)。一方で ハトは,特殊な状況を除き,全体よりも局所を重視する 傾向が強いため,左から HSと答える(Cavoto & Cook, 2001)。ヒトは「木よりも森を見る」傾向が強いのに対 して,ハトは「森よりも木を見る」傾向が強いといえる。 チンパンジー(Fagot & Tomonaga, 1999),ヒヒ(Deruelle & Fagot, 1998; Fagot & Deruelle, 1997),アカゲザル(Hop-kins & Washburn, 2002),フサオマキザル(Spinozzi, De Lillo, & Salvi, 2006; Spinozzi, De Lillo, & Truppa, 2003) と いった霊長類でも,このNavon型階層図形に対しては, ヒトに比べて全体より個々の要素に注意を向ける傾向が 強いことが報告されている。
5.まとめ
紹介した。ヒトの錯視が唯一絶対のものではないという ことが明らかになりつつあるが,ここでいう「ヒト」と は,厳密には「ヒトのオトナ」を指している。ヒトのオ トナでは生じることがないような錯視も研究対象となり うる点において,比較錯視研究と錯視の発達研究には共 通点があるといえる。両研究領域の成果を共有していく ことで,錯視の多様性や視覚の多様性を明らかにするこ と,またヒトのオトナにおいて生じる錯視や視覚の特徴 を相対的な視点から検討することができるだろう。 例えば,種間で多様性が報告されているエビングハウ ス・ティチェナー錯視図の知覚に関しては,ヒトという 種内においても多様性があることが報告されている。ま ず,ヒトのオトナでも,中心円の大きさを見た目で判断 するように教示された場合には錯視が生じるが,中心円 を親指と人差し指でつかむように教示された場合(動作 で判断した場合)にはほとんど錯視が生じないことが報 告されている(Aglioti, DeSouza, & Goodale, 1995; Haffen-den, & Goodale, 1998)。この結果は,知覚に対する視覚 システムと動作に対する視覚システムがそれぞれ分かれ ていることを示唆する認知神経科学の知見とも一致する (Goodale & Milner, 1992; Milner & Goodale, 1995)。また,
ヒトのなかでも自閉症児では,ヒヒと同じように錯視が 生じないという報告がある(Happé, 1996; but see Ropar & Mitchell, 1999)。また,ヒトの4歳児ではこの錯視が生起 せず,年齢を重ねるとともに錯視量が増加するという報 告もある(Doherty, Campbell, Tsuji, & Phillips, 2010)1。同
じ図形を目の前にしても発達段階によって認識の仕方が 異なるといった現象は,Figure 2(G)のような図にも生 じる。この図に対して,動いている棒を“1本”と認識 するのは生後4カ月以降であること,さらに棒が静止し ている場合にそれを“1本”と認識するのは生後6カ月 以降だという(Kellman & Spelke, 1983)。比較錯視研究で は,これらに類似した錯視の種差を情報処理傾向の違い (全体志向的か局所志向的か)として解釈したが,もし かしたら同様の解釈をヒトの錯視発達現象に対しても適 用できるかもしれない。ただし,こうした検討はごく一 部の錯視図についてのみおこなわれているに過ぎないの が現状である。今後は,他の錯視図に関しても両領域で 研究を進めていき,そこで得られた情報を共有していく ことが必要であろう。また,ヒトのコドモやある特定の 動物においてのみ生じるような,錯視辞典には掲載でき ない“錯視”というものがあっても不思議ではない。今 後はこうした新しい錯視の発見を試みていくことも重要 であると思われる。 引 用 文 献
Aglioti, S., DeSouza, J. F., & Goodale, M. A. (1995). Size-con-trast illusions deceive the eye but not the hand. Current
Bi-ology, 5, 679–685.
Agrillo, C., Parrish, A. E., & Beran, M. J. (2014). Do rhesus monkeys (Macaca mulatta) perceive the Zöllner illusion?
Psychonomic Bulletin and Review, 21, 986–994.
Cavoto, K. K., & Cook, R. G. (2001). Cognitive precedence for local information in hierarchical stimulus processing by pi-geons. Journal of Experimental Psychology: Animal Behavior
Processes, 27, 3–16.
Coren, S., & Girgus, J. S. (1978). Seeing is deceiving: The
psy-chology of visual illusions. Hillsdale, NJ: Erlbaum.
Deruelle, C., & Fagot, J. (1998). Visual search for global/local stimulus features in humans and baboons. Psychonomic
Bul-letin & Review, 5, 476–481.
Doherty, M. J., Campbell, N. M., Tsuji, H., & Phillips, W. A. (2010) The Ebbinghaus illusion deceives adults but not young children. Developmental Science, 13, 714–721. Dücker, G. (1966). Untersuchungen über geometrisch-
optische Täuschungen bei Wirbeltieren (Optical Illusions in Vertebrates). Zeitschrift für Tierpsychologie, 23, 452–496. (In German with English summary)
Fagot, J., & Deruelle, C. (1997). Processing of global and local visual information and hemispheric specialization in hu-mans (Homo sapiens) and baboons (Papio papio). Journal of
Experimental Psychology: Human Perception and Perfor-mance, 23, 429–442.
Fagot, J., & Tomonaga, M. (1999). Global and local processing in humans (Homo sapiens) and chimpanzees (Pan
troglo-dytes): Use of a visual search task with compound stimuli. Journal of Comparative Psychology, 113, 3–12.
Fujita, K., & Ushitani, T. (2005). Better living by not complet-ing: A wonderful peculiarity of pigeon vision? Behavioural
Processes, 69, 59–66.
Geiger, G., & Poggio, T. (1975). The Mueller-Lyer figure and the fly. Science, 190, 479–480.
Glazebrook, C. M., Dhillon, V. P., Keetch, K. M., Lyons, J., Amazeen, E., Weeks, D. J., & Elliott, D. (2005). Perception-action and the Müller–Lyer illusion: Amplitude or endpoint bias? Experimental Brain Research, 160, 71–78.
Goodale, M. A., & Milner, A. D. (1992). Separate visual path-ways for perception and action. Trends in Neuroscience, 15, 20–25.
後藤倬男・田中平八(2005).錯視の科学ハンドブック 東京大学出版会
Haffenden, A. M., & Goodale, M. A. (1998). The effect of pic-torial illusion on prehension and perception. Journal of
Cog-nitive Neuroscience, 10, 122–136.
Happé, F. (1996). Studying weak central coherence at low
lev-1 5∼8カ月児でもエビングハウス・ティチェナー錯視
が生じていることを示唆した研究もある(Yamazaki, Otsuka, Kanazawa, & Yamaguchi, 2010)。
els: Children with autism do not succumb to visual illu-sions. A research note. Journal of Child Psychology and
Psy-chiatry, 37, 873–877.
Hopkins, W. D., & Washburn, D. A. (2002). Matching visual stimuli on the basis of global and local features by chimpan-zees (Pan troglodytes) and rhesus monkeys (Macaca
mulat-ta). Animal Cognition, 5, 27–31.
今井省吾(1984).錯視図形―見え方の心理学 サイエ ンス社
Kellman, P. J., & Spelke, E. S. (1983). Perception of partly oc-cluded objects in infancy. Cognitive Psychology, 15, 483–524. 金沢 創(2016).錯視の発達 基礎心理学研究,35,
43–46.
Milner, A. D., & Goodale, M. A. (1995). The visual brain in
ac-tion. Oxford: Oxford University Press.
盛永四郎(1956).大きさ対現象の条件の吟味 日本心 理学会第70回大会発表論文集,53.
Murayama, T., Usui, A., Takeda, E., Kato, K., & Maejima, K. (2012). Relative size discrimination and perception of the Ebbinghaus illusion in a bottlenose dolphin (Tursiops
trun-catus). Aquatic Mammals, 38, 333–342.
Nakamura, N., Fujita, K., Ushitani, T., & Miyata, H. (2006). Perception of the standard and the reversed Müller–Lyer figures in pigeons (Columba livia) and humans (Homo
sapi-ens). Journal of Comparative Psychology, 120, 252–261.
Nakamura, N., Watanabe, S., Betsuyaku, T., & Fujita, K. (2010). Do bantams (Gallus gallus domesticus) experience amodal completion? An analysis of visual search perfor-mance. Journal of Comparative Psychology, 124, 331–335. Nakamura, N., Watanabe, S., & Fujita, K. (2008). Pigeons
per-ceive the Ebbinghaus–Titchener circles as an assimilation il-lusion. Journal of Experimental Psychology: Animal Behavior
Processes, 34, 375–387.
Nakamura, N., Watanabe, S., & Fujita, K. (2014). A reversed Ebbinghaus–Titchener illusion in bantams (Gallus gallus
domesticus). Animal Cognition, 17, 471–481.
Navon, D. (1977). Forest before trees: The precedence of glob-al features in visuglob-al perception. Cognitive Psychology, 9, 353–383.
小原嘉明(2003)モンシロチョウ―キャベツ畑の動物行 動学 中公新書
Parron, C., & Fagot, J. (2007). Comparison of grouping abili-ties in humans (Homo sapiens) and baboons (Papio papio) with the Ebbinghaus illusion. Journal of Comparative
Psy-chology, 121, 405–411.
Pepperberg, I. M., Vicinay, J., & Cavanagh, P. (2008). Process-ing of the Müller–Lyer illusion by a Grey parrot (Psittacus
erithacus). Perception, 37, 765–781.
Post, R. B., Welch, R. B., & Caufield, K. (1998). Relative spatial expansion and contraction within the Müller–Lyer and Judd
illusions. Perception, 27, 827–838.
Roberts, B., Harris, M. G., & Yates, T. A. (2005). The roles of inducer size and distance in the Ebbinghaus illusion (Titch-ener circles). Perception, 34, 847–856.
Robinson, J. O. (1998). The psychology of visual illusion. Mine-ola, New York: Dover.
Ropar, D., & Mitchell, P. (1999). Are individuals with autism and Asperger’s syndrome susceptible to visual illusion.
Journal of Child Psychology and Psychiatry, 40, 1283–1293.
Spinozzi, G., De Lillo, C., & Salvi, V. (2006). Local advantage in the visual processing of hierarchical stimuli following manipulations of stimulus size and element numerosity in monkeys (Cebus apella). Behavioural Brain Research, 166, 45–54.
Spinozzi, G., De Lillo, C., & Truppa, V. (2003). Global and lo-cal processing of hierarchilo-cal visual stimuli in tufted capu-chin monkeys (Cebus apella). Journal of Comparative
Psy-chology, 117, 15–23.
Suganuma, E., Pessoa, V. F., Monge-Fuentes, V., Castro, B. M., & Tavares, M. C. H. (2007). Perception of the Müller–Lyer illusion in capuchin monkeys (Cebus apella). Behavioural
Brain Research, 182, 67–72.
Tudusciuc, O., & Nieder, A. (2010). Comparison of length judgments and the Müller–Lyer illusion in monkeys and humans. Experimental Brain Research, 207, 221–231. Ushitani, T., Fujita, K., & Yamanaka, R. (2001). Do pigeons
(Columba livia) perceive object unity? Animal Cognition, 4, 153–161.
Watanabe, S., Nakamura, N., & Fujita, K. (2011). Pigeons per-ceive a reversed Zöllner illusion. Cognition, 119, 137–141. Watanabe, S., Nakamura, N., & Fujita, K. (2013). Bantams
(Gallus gallus domesticus) also perceive a reversed Zöllner illusion. Animal Cognition, 16, 109–115.
Watanabe, S., Nishimoto, Y., Fujita, K., & Ishida, M. (2014). Budgerigars (Melopsittacus undulatus) perceive a Müller– Lyer illusion. The Japanese Journal of Psychonomic Science,
33, 117–118.
Weintraub, D. J. (1979). Ebbinghaus illusion: Context, contour, and age influence the judged size of a circle amidst circles.
Journal of Experimental Psychology: Human Perception and Performance, 5, 353–364.
Winslow, C. N. (1933). Visual illusions in the chick. Archives of
Psychology, 153, 1–83.
Yamazaki, Y., Otsuka, Y., Kanazawa, S., & Yamaguchi, M. K. (2010). Perception of the Ebbinghaus illusion in 5- to 8- month old infants. Japanese Psychological Research, 52, 33– 40.
山口真美(2016).錯視の基盤: 視知覚の定型・非定型 の発達をめぐって 基礎心理学研究,35, 47–52.