新潟県の日本紅斑熱患者発生地域における
マダニ保有リケッチア群調査
新井礼子,加藤美和子,青木順子,池田菫1),田村務,Marcello Otake Sato2),サトウ恵1)
Investigation of Rickettsiae in Ticks Collected in the Area around the First
Confirmed Case of Japanese Spotted Fever Patient in Niigata Prefecture
Reiko Arai,Miwako Kato,Junko Aoki,Sumire Ikeda1),Tsutomu Tamura,Marcello Otake Sato2),Megumi Sato1)
Japanese spotted fever patient was confirmed first time in Niigata Prefecture in July 2014.It was suspected that ticks reserving
Rickettsia japonica had invaded and lived in Niigata Prefecture,therefore,we made investigation of Rickettsial DNA in ticks collected
in the area where Japanese spotted fever patient infected.As a result,535 ticks identified consisted of two genera and four species;
Haemaphysalis longicornis,Haemaphysalis flava,Ixodes ovatus,and Ixodes nipponensis.No ticks were found to be infected with Rickettsia japonica.However,spotted fever group Rickettsiae were found in 7 samples.
Keywords:Japanese spotted fever,Spotted fever group Rickettsiae,Tick 1 緒言 日本紅斑熱はRickettsia japonicaを保有するマダニに刺咬 されることによって感染,発症するリケッチア感染症で, 1984年に馬原らによって命名された1).マダニに刺咬後,2 ~8日後に発熱,発疹を主訴として発症する.日本国内では 主に西日本で毎年170名程度の患者が発生し,2007年から 2016年までの10年間に16名の死亡者が報告されている2). これまで東日本の日本海側では患者発生が確認されてい なかったが,2014年には福井県と新潟県で初めて患者が報 告された3,4).新潟県内で初めて確認された患者は疫学調 査の結果,自宅周辺でマダニに刺咬された可能性が高く, さらなる患者の発生が危惧された. また近年,R. japonica以外の紅斑熱群リケッチア(Spotted fever group Rickettsiae,以下,SFGR)の感染による紅斑熱 症例が国内で報告されている5)ことから,患者発生地域の SFGRの侵淫状況を把握するため,患者宅周辺におけるマダ ニの生息状況と保有リケッチアについて調査を行った. 2 材料と方法 2.1 調査地点と調査期間 調査地点として患者宅周辺の山野2ヶ所を選出した.2ヶ 所とも標高約15~20 mで,地点Aは患者宅から南東約2 km の運動公園周囲の緑地帯,地点Bは患者宅から西約7 kmの 林道脇草地とし,2015年6月,2016年5月,11月および2017 年3月の計4回にわたりマダニを採取した. 2.2 方 法 マダニの採取はフランネル製の白布を用いた旗ふり法6) により行い,白布に付着したマダニをピンセットで回収し た. 回収したマダニを採取日,調査地点,マダニ種,発育 ステージ及び性別ごとに分類し,一部のマダニ種の鑑別に ついては,後述の方法でマダニから抽出したDNAを用いて Takanoらの方法7)により遺伝子学的に同定した. 成虫は1個体を1検体,若虫及び幼虫については5個体を1 検体としてエタノールで洗浄後,メタルビーズ入りチュー ブに入れて滅菌リン酸緩衝液またはIsogen Ⅱ(日本ジー ン)を添加し細胞破砕機を用いてマダニ虫体を破砕後, 1)新潟大学医学部 2)獨協医科大学医学部
Target Gene Refarence Rr17k 1p (forward) Rr17k 539n (reverse) Rr17k 90p (forward) Rr17k 539n (reverse) RpCs 780p (forward) RpCs 1258n (reverse) RpCs 877p (forward) RpCs 1258n (reverse) RR 190-70 (forward) RR 190-70l (reverse) 190FN1 (forward) 190RN1 (reverse) rompB OF (forward) rompB OR (reverse) rompB SFG IF (forward) rompB SFG/TG IR (reverse) Primer
Table 1. Primer pair used for detection and typing of SFGR
1st 2nd 1st 8) 9) 2nd 17kDa gltA 9~11) 12) 2nd 1st 2nd ompB 1st ompA
QIAamp DNA Mini Kit(QIAGEN),Isogenome(日本ジー ン)またはHigh Pure PCR Template Preparation Kit(Roche) を用いてDNAを抽出し,nested-PCR法によりSFGR遺伝子を 検出した. 使用したプライマーはTable 1のとおりで,まず,SFGR の共通抗原である17kDaタンパク質をコードする遺伝子領 域(以下,17kDa)を検出し,陽性となった検体について Rickettsia属共通のクエン酸合成酵素遺伝子領域(以下, gltA)を検出した.両領域とも陽性となった検体をSFGR陽
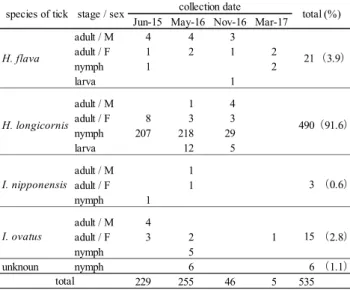
性とした.さらに,Outer membrane protein A遺伝子領域(以 下,ompA)及びOuter membrane protein B遺伝子領域(以下, ompB)の2ヶ所について検出し,17kDa,gltAの増幅産物と ともにダイレクトシークエンス法により塩基配列を解析し てリケッチア種の同定を試みた. 3 結果 3.1 マダニ生息状況 2地点延べ8回の調査で採取したマダニは535個体で,その うち490個体(91.6 %)がHaemaphysalis longicornis(フタ トゲチマダニ)であった.他にHaemaphysalis flava(キチ マダニ)(3.9 %),Ixodes ovatus(ヤマトマダニ)(2.8 %) 及び Ixodes nipponensis(タネガタマダニ)(0.6 %)が採取 された.また,若虫6個体については同定することが出来な かった.4回の調査のうち,2016年5月に採取されたマダニ が255個体と最も多く,次いで2015年6月の229個体,2016 年11月の46個体で,この3回の調査ともH. longicornisの若虫 が最も多く採取された.2017年3月にはすでに積雪はなかっ
たが,H. flava 4個体とI. ovatus 1個体が採取されたのみであ
った(Table 2). 3.2 マダニのSFGR保有状況 535個体のマダニから抽出したDNA153検体のうち,7検 体からSFGRを検出した(Table 3).Ixodes属の2種では14 検 体 中4 検 体 か ら 検 出 さ れ た . Haemaphysalis属 で は H. longicornis 117検体中2検体から検出され,H. flava 20検体か らは検出されなかった.同定できなかったマダニから1検体 検出された.SFGRが検出された7検体のうち,ompBは全て の検体で検出されたが,ompAは4検体のみ検出された. 3.3 SFGRの塩基配列解析 検出された増幅産物の塩基配列に基づく系統樹解析を行 った結果(Fig.1),R. japonica と一致するものはなかっ たが,Rickettsia asiaticaと推定された検体が2検体,Rickettsia monacensisが3検体,既知のリケッチア種のいずれかと推定 することが出来なかったものが2検体であった(Table 4). 4 考察 マダニは生息の至適温度や湿度,主たる吸血源動物が種 によって異なるため,生息する地域や環境がそれぞれ異な ることが知られている13). また,これまでに日本紅斑熱の原因であるR. japonicaの 検出が報告されているマダニは地域により異なっており, 比較的温暖な地域で生息するHaemaphysalis. hystricis(ヤマ アラシチマダニ),Haemaphysalis formosensis(タカサゴチ マ ダ ニ ) ,Haemaphysalis cornigera(ツノチマダニ), Dermacentor taiwanensis(タイワンカクマダニ)と,広く全 国に生息するH. longicornis及びH. flavaである5). 今回の調査で採取されたマダニのうち,H. longicornis,
H. flava,I. ovatus及びI. nipponensisは全て広く全国に生息し
ている種である.比較的温暖な地域を好むマダニ種は調査 期間を通じて採取されなかったことから,調査地点の優位 種ではないと考えられた. また,採取したマダニからR. japonicaは検出されなかっ たため,2014年に当県で確認された日本紅斑熱患者の媒介 マダニ種を推測することは出来なかったが,採取されたマ ダニの種類と割合から,H. longicornisまたはH. flavaであっ た可能性が高いと考えられる. SFGRについては,Haemaphysalis属よりもIxodes属から検 出される割合が高い傾向にあった.I. nipponensis 2検体と同 定不能マダニ1検体からR. monacensisと推定されたSFGRが 3株検出された(Table 4).R. monacensisはヨーロッパでは 紅斑熱症の原因として確認されており,広くIxodes属から 検出されるSFGRである14).ompAの系統樹において韓国お よび中国由来の株とヨーロッパ由来の株とではそれぞれ異 なるクラスターを形成する(Fig.1C).今回検出された Table 2. Identification of the ticks collected in Niigata prefecture.
Jun-15 May-16 Nov-16 Mar-17 adult / M 4 4 3 adult / F 1 2 1 2 nymph 1 2 larva 1 adult / M 1 4 adult / F 8 3 3 nymph 207 218 29 larva 12 5 adult / M 1 adult / F 1 nymph 1 adult / M 4 adult / F 3 2 1 (2.8) nymph 5 unknoun nymph 6 6 (1.1) 229 255 46 5 535 I. ovatus 15 total I. nipponensis
species of tick stage / sex collection date H. flava H. longicornis total (%) (91.6) (0.6) 21 490 3 (3.9)
Table 3. Detection of SFGR's DNA from ticks
species of tick no. tested no. positive by nested-PCR
H. flava 20 0 H. longicornis 117 2 I. nipponensis 3 2 I. ovatus 11 2 unknown 2 1 total 153 7
A B
C D
Fig.1.Phylogenetic trees for identification of species of Rickettsiae based on 450 nucleotide of 17kDa gene(A),382 nucleotide of gltA gene(B),540 nucleotide of ompA gene(C),and 426 nucleotide of ompB gene(D).Black dots indicate positive samples detected from ticks,and white circles(A,B,and C) indicate R. japonica identified from Japanese spotted fever patient in Niigata Prefecture in 2014.
AF445384 R. sibirica CP003375 R. slovaca str. D-CWPP U17088 R. parkeri
AE006914 R.conorii subs. conorii Malish7 CP001612 R. africae ESF-5 HF935073 R. peacockii CAA501 AF027124 R. honei
14-973 17kDa D16515 R. japonica YH
AB473988 R. heilongjiangensis Sendai-16 CP010969 R. raoultii Khabarovsk
AB114804 Rickettsia sp. Hf332 T16-58 17kDa
AB114810 Rickettsia sp. Hj126 KJ534311 R. amblyommii isolate IL4
T15-110 17kDa
AB699873 Rickettsia sp. clone: Saga006 CP013133 R. rhipicephali strain HJ-5 M74042 R.australis
T15-117 17kDa T15-118 17kDa AB114798 R. asiatica IO-1 AB114800 Rickettsia sp. IO-25
GU827035 R. helvetica isolate 102 T15-164 17kDa
T16-54 17kDa T16-71 17kDa
LN794217 R. monacensis strain IrR Munich AB795127 R. tamurae HM-1
CP004889 R.prowazeki Breinl 0.01
KU587806 R. rickettsii strain InDRE 1098/Mexico/2013 14-973glt
CP002912 R. heilongjiangensis 054 AP017602 R. japonica strain YH-M CP010969 R. raoultii strain Khabarovsk
T16-58 gltA AY578114 R. principis
CP015012 R. amblyommatis isolate An13 CP003375 R. slovaca str. D-CWPP
T15-110glt DQ105664 R. helvetica
AB297808 R. asiatica strain: Huangshan-3(China:Mt.Huang-shan) AB297810 R. asiatica strain: Jeju-2(South Korea: Jeju Island) AB114799 Rickettsia sp. IO-25
AF394901 R. asiatica strain IO-1 /Japan/2001 T15-117 gltA
T15-118 gltA AB812551 R. tamurae strain HM-1 EU665235 R. monacensis(Central China) KC993860 R. monacensis strain CN45kr(South Korea)
T16-71 gltA T16-54 gltA
T15-164 gltA
KP215387 R. monacensis strain Est1623 LN794217 R. monacensis strain IrR/Munich/USA
U59715 R. prowazekii KC469611 R. typhi
0.01
FJ172358 R. parkeri JF700255 R. sibirica
DQ150681 R. rickettsii strain 1989CO1 CP003375 R. slovaca str. D-CWPP
EU716649 R. conorii
AP017602 R. japonica strain: YH_M 14-973 rOmpA
CP002912 R. heilongjiangensis 054 T15-110 rOmpA DQ372955 R. antechini
GQ223392 R. tasmanensis strain T152 AH015610 R. raoultii strain Khabarovsk JN043510 Rickettsia sp. GL162
JX972178 R. monacensis strain MT34 (Korea/2012) EU665232 R. monacensis (China: Anhui) KJ588273 R. monacensis strain CHO11 (Korea/2014)
T16-71 rOmpA T16-54 rOmpA T15-164 rOmpA
DQ100169 R. monacensis strain IrR/Munich /USA KT119437 R. monacensis isolate Est1623 /Estonia/ FJ919640 R. monacensis isolate N19 /Italy/2009 DQ103259 R. tamurae strain AT-1
0.02
AE006914 R. conorii str. Malish 7 CP003375 R. slovaca str. D-CWPP CP001227 R. peacockii str. Rustic
AF123708 R. conorii subsp. caspia A-167 HM050273 R. sibirica strain RH05
T15-110 rOmpB T16-58 rOmpB
AP017602 R. japonica strain: YH_M GQ223393 R. tasmanensis strain T152
KJ663752 R. raoultii strain 4
EU883092 R. monacensis strain CN45Kr(South Korea) LN794217 R. monacensis strain IrR/Munich
T16-71 rOmpB T16-54 rOmpB T15-164 rOmpB
DQ113910 R. tamurae strain AT-1 DQ110870 R. asiatica strain IO-1
T15-118 rOmpB T15-117 rOmpB
KP866151 R. helvetica isolate Komi
株も東アジア地域の株と相同性が高く,ヨーロッパ地域か ら検出される株とは遺伝子学的に若干の違いが見られる可 能性が示唆された. また,I. ovatusの2検体からR. asiaticaと推定された株が検 出さ れた .R. asiaticaは,1993年にFujitaらが福島県でI. ovatusから分離した株で15)その後も国内各地で同様の株が 分離され,ニホンジカの血液からも検出されている14).今 回使用したプライマーではompBの領域は検出されたが, ompAについては検出することが出来なかったこともFujita らの報告と同様の結果となった. H. longicornisから検出した2株はいずれもリケッチア種 が推定できなかった(Table 4).17kDaの系統樹(Fig.1A) においてNo. T15-110と塩基配列が一致したRickettsia sp. clone Saga 006(AB699873)は佐賀県で野生のアライグマ の脾臓から検出された株である16).またNo. T16-58につい
てはgltAの領域(Fig.1B)で極東ロシアや中国東北部で
Haemaphysalis japonica(ヤマトチマダニ)から検出されて
いるCandidatus Rickettsia principis(AY578114)14)と一致し
た.同様の株は国内で広く検出されている17)ことから,こ れら2検体と同じようなSFGRは広く国内に生息していると 示唆された.この2検体については,さらに詳細な解析を実 施して行く必要があると考えている. 今回は日本紅斑熱患者発生地域で調査を実施したが,R. japonicaは検出されなかった.しかしR. japonicaを媒介する と指摘されているH. longicornis及びH. flavaが採取された ことや ,病原 性が 不明な が らもR. japonica以外の4種の SFGRが検出されていることから,この地域において紅斑熱 が発生する可能性は否定できず,今後も監視を継続するこ とが必要と考えられた.また,当県は南北に長く,また海 岸部と山間部では気象条件も大きく異なることから,広く 県内で同様の調査を実施し,新潟県全体のマダニの生息状 況と病原体の保有状況を把握する必要がある. 5 結語 2014年に新潟県内で初めて日本紅斑熱患者の発生を確認 したことから,患者宅周辺の山野においてマダニ生息状況 と保有SFGRの調査を実施した.H. longicornis,H. flava,I. ovatus及びI. nipponensisの2属4種のマダニと同定不能マダ ニ計535個体が採取された.これらのマダニからR. japonica は検出されなかったが,他のSFGRが7検体から検出された. 本報文は,経常研究「新潟県のマダニにおけるリケッチ ア等の検索」(2015~2016年度)及び科学研究費助成事業 基盤研究(C)「マダニ棲息分布調査と次世代シーケンシ ングによる保有病原体の網羅的検出」(2016~2018年度) (研究代表者:新潟大学医学部サトウ恵助教)の成果であ る. 6 文献 1)馬原文彦,古賀敬一,沢田誠三等:わが国初の紅斑熱リ ケッチア感染症,感染症誌,59,1165-1172(1985). 2)国立感染症研究所,病原微生物検出情報月報,38,110-112 (2017). 3)高田伸弘,清水達人,五十嵐一誠等:福井県南部で初確 認できた紅斑熱の速報,若狭湾地方の環境要因と症例比 較から,衛生動物,69,60(2015). 4)新井礼子,加藤美和子,青木順子等:新潟県の日本紅斑 熱患者発生地域におけるマダニ保有リケッチア調査,第 68回日本衛生動物学会東日本支部大会プログラム・講演 要旨2016,29(2016). 5)安藤秀二,藤田博巳:国内における紅斑熱群リケッチア 症を媒介するマダニ類と病原体との多様な関係,Med. Entomol.Zool.,64,5-7(2013). 6)高田伸弘:病原ダニ類図譜,金芳堂(1990). 7) Takano A,Fujita H,Kadosato H,et al.:Construction of
a DNA datebase for ticks collected in Japan:application of molecular identification based on the mitochondrial 16S rDNA gene,Medical Entomology andZoology,65,13-21 (2014).
8)Mitsuhiro Ishikura,Shuji Ando,Yasuhiro Shinagawa,et al. :Phylogenetic analysis of spotted fever group Rickettsiae based on gltA,17-kDa,and rOmpA genes amplified by nested PCR from ticks in Japan,Microbiol.Immunol.,47,823-832 (2003).
Table 4. Presumed species of SFGR in ticks.
ompA ompB
T15-110
Jun-15 H. longicornis
nymph
+
+
unknown
T15-117
Jun-15 I. ovatus
adult / M
-
+
R. asiatica
T15-118
Jun-15 I. ovatus
adult / M
-
+
R. asiatica
T15-164
Jun-15 I. nipponensis
nymph
+
+
R. monacensis
T16- 54
May-16 I. nipponensis
adult / F
+
+
R. monacensis
T16- 58
May-16 H. longicornis
nymph
-
+
unknown
T16- 71
May-16 unknown
nymph
+
+
R. monacensis
9)Russell L.Reggnery,Cathrine L.Spruill,Brian D.Plikaytis :Genotypic identification of Rickettsiae and estimation of interspecies sequence divergence for portion of two rickettsial genes,J.Bacteriol,173,1576-1589(1991).
10)Fourier PE.,Roux V.,Raoult D.:Phylogenetic analysis of spotted fever group rickettsiae by study of the outer surface protein rOmpA,Int.J.Syst.Bacteriol.,48,839-849 (1998).
11)Christpher D.Paddok,John W.Sumner,James A.Comer, et al.:Rickettsia parkeri:A newly recognized cause of spotted fever rickettsiosis in the United States,CID,38, 805-811(2004).
12)Yeon-Joo Choi,Won-Jong Jang,Jong-Hyun Kim,et al. :Spotted fever group and typhus group rickettsioses in humans,South Korea,Emerging Infectious Deseases,11, 237-244(2005).
13)高野愛:マダニの生態とマダニ媒介感染症,山口獣医 学雑誌,42,1-8(1990).
14 ) Philippe Parola , Christopher D . Paddock , Cristina Socolovschi,et al.:Update tick-bone rickettsioses around the world:a Geographic approach,Clin.Microbiol.Rev., 26,657-702(2013).
15)Hiromi Fujita,Pierre-Edouard Fournier,Nobuhiro Takada, et al.:Rickettsia asiatica.sp.nov.,isolated in Japan, International Jounal of Systematic and Evolutionary Microbiology,56,2365-2368(2006).
16)Kenji Baba,Toshiya Kaneda,Hitoshi Nishimura,Hiroshi Sato:Molecular detection of spotted fever group Rickettsia in feral raccoons(Procyon lotor)in the western part of Japan, J.Vet.Med.Sci.,75,195-197(2013).
17)Gaowa,Norio Ohashi,Minami Aochi,et al.:Rickettsia in ticks,Japan,2007-2011,Emerging Infectious Disease, 19,338-340(2013).