様式 C-19
科学研究費補助金研究成果報告書
平成21年 4月 10日現在
研究成果の概要:
本研究から、植物における Nudix hydrolase(AtNUDX)は多様な生理的役割を果たしているこ とが明らかになった。すなわち、シロイヌナズナ細胞質局在型の AtNUDX1 は酸化ヌクレオチド 浄化、AtNUDX2 は ADP-リボースのリサイクル、AtNUDX7 は NADH 代謝を介した PAR 反応制御によ り酸化ストレス防御に機能していることを明らかにした。また、推定オルガネラ局在型の AtNUDX15、14、19、および 23 の細胞内局在性および特異的基質も明らかにした。
交付額
(金額単位:円)
直接経費 間接経費 合 計
2007 年度 2,300,000 0 2,300,000 2008 年度 2,300,000 0 2,300,000
年度 年度 年度
総 計 4,600,000 0 4,600,000
研究分野:植物生理学
科研費の分科・細目:特定領域研究・オルガネラ分化
キーワード:Nudix hydrolase, ヌクレオシド 2-リン酸, DNA 突然変異, 酸化ストレス, 酸化ヌ クレオチド, ポリ(ADP-リボシル)化, ADP-ribose, NADH
1.研究開始当初の背景
Nudix hydrolase は、酸化ヌクレオチド (8-oxo-dGTP、2-OH-dATP など)、ADP-ribose、
NADH、FAD、CoA などの生体分子が含まれるヌ クレオシド 2-リン酸類縁体に対する加水分 解活性を有する酵素ファミリーである。高等 植物のシロイヌナズナ(At)には 27 種類の Nudix hydrolase 相同遺伝子(細胞質型:
AtNUDX1~11, 25、ミトコンドリア型:
AtNUDX12~18、葉緑体型:AtNUDX19~24, 26, 27)が存在している。したがって、それらは ミトコンドリア、葉緑体、細胞質などの種々 のオルガネラで多様な代謝反応の制御に深 く関与していると推測される。しかしこれま でに、本酵素が酸化ヌクレオチド以外のヌク レオシド 2-リン酸類縁体の代謝に関与する ことの生理的意義については全く明らかに なっていない。
研究種目:特定領域研究 研究期間:2007~2008 課題番号:19039032
研究課題名(和文) 植物オルガネラにおけるヌクレオシド2−リン酸類縁体の代謝
研 究 課 題 名 ( 英 文 ) Metabolisms of nucleoside 2-phosphate derivatives in plant organelles
研究代表者
重岡 成( SHIGEOKA SHIGERU ) 近畿大学・農学部・教授
研究者番号:80140341
研究者番号:80140341
2.研究の目的
Nudix ( Nucleoside Diphosphate linked some moiety X)hydrolase は、ヌクレオシド 2-リン酸類縁体に対する加水分解活性を有 する酵素ファミリーである。高等植物シロイ ヌナズナには細胞質 (12 種)、ミトコンドリ ア (7 種)、葉緑体 (8 種)などに計 27 種類も の Nudix hydrolase が局在していると推測さ れることから、各オルガネラでの種々のヌク レオシド 2-リン酸類縁体の代謝が酸化スト レス応答や代謝調節などの多様な細胞応答 に大きな影響を与えていることが示唆され る(図 1)。そこで本研究では、ミトコンドリ ア 型 (AtNUDX12 ~ 18) お よ び 葉 緑 体 型 (AtNUDX19~24, 26, 27) AtNUDX の分子特性、
および各オルガネラにおける AtNUDX による 種々のヌクレオシド 2-リン酸類縁体 (特に、
酸 化 ヌ ク レ オ チ ド , ADP-ribose, NAD(P)H, FAD, CoA など) の代謝制御の全容を明らかに することを目的とする。 シロイヌナズナNudix hydrolaseファミリーのアライメント
!"#$%&'()*+,-./0123*456"789:;<-=>?@!
!"#$%&'(!
!"#$%&')!
!"#$%&'*!
!"#$%&'+!
!"#$%&',!
!"#$%&'-!
!"#$%&'.!
!"#$%&'/!
!"#$%&(0!
!"#$%&('!
!"#$%&((!
!"#$%&()!
!"#$%&(*!
!"#$%&(,!
!"#$%&(-!
Nudix motif!
1!&&&&2&!3!&&&&&&&!43$!&!33!1!&!
!!
!
"
#
$
%
&
' ( )
* + , -
図 1. シロイヌナズナ Nudix hydrolase ファ ミリーの Nudix モチーフ近傍のアライメント
* 選択的スプライシングによりペルオキシ ソーム (PTS1)局在型
3.研究の方法
シロイヌナズナにおける推定ミトコンド リア型 (AtNUDX12~18)および葉緑体型 (AtNUDX19~24, 26, 27) AtNUDX の分子特性 を明らかにするために、大腸菌を用いてリコ ンビナントタンパク質を生成した。得られた リコンビナントタンパク質は Ni アフィニテ ィークロマトグラフィーにより単一に精製 した。
AtNUDX の細胞内局在性を明らかにするた めに、GFP 融合タンパク質発現プラスミドを 構築した。得られたプラスミドはアグロバク テリアを用いて植物培養細胞(タバコ BY-2 株、シロイヌナズナ緑色培養細胞 T-87 株)
に形質転換した。
AtNUDX1, 2, 6, 7, 19, 23 の生理機能を明 らかにするために、それぞれの遺伝子破壊株 を SALK より入手した。また、それらの過剰 発現株は CaMV プロモーターを用いた定法に 従って作製した。
4.研究成果
(1) シ ロイヌナズナ全 Nudix hydrolase (AtNUDX1~27)の分子特性 の解析
これまでに我々は、細胞質局在型 AtNUDX の分子特性を明らかにしている(Ogawa et al.
J. Biol. Chem. 280: 25277-25283,2005) 。 す な わ ち 、 そ れ ら は 酸 化 ヌ ク レ オ チ ド
(AtNUDX1)、ADP-ribose および NADH(AtNUDX2, 6, 7, 10)、CoA(AtNUDX11)、ApnA(AtNUDX25)
に対して特異的な加水分解活性を有してい た 。 そ こ で 、 推 定 ミ ト コ ン ド リ ア 型 (AtNUDX12~18)および葉緑体型(AtNUDX19~
24, 26, 27) AtNUDX の分子特性を明らかにす るために、それらのリコンビナントタンパク 質を用いた酵素学的性質の解析を行った。そ の結果、AtNUDX14 は ADP-ribose、AtNUDX15 は CoA および種々の CoA 誘導体、AtNUDX19 は NADPH、AtNUDX 23 は FAD を特異的基質とする ことが分かった(図 2)。
次に、これらの AtNUDX の細胞内局在性を GFP 融合タンパク質を用いて解析した。その 結果、AtNUDX15 はミトコンドリア、AtNUDX14、
AtNUDX19 および 23 は葉緑体に局在すること が 確 認 さ れ た ( Ogawa & Yoshimura et al.
Plant Physiol. 2008 )。
図 2. AtNUDX の細胞内局在性と推定される機 能
(2) 種々 の ヌクレオ シド 2-リン酸 類縁体 代謝 の生理 的 意義 の解明
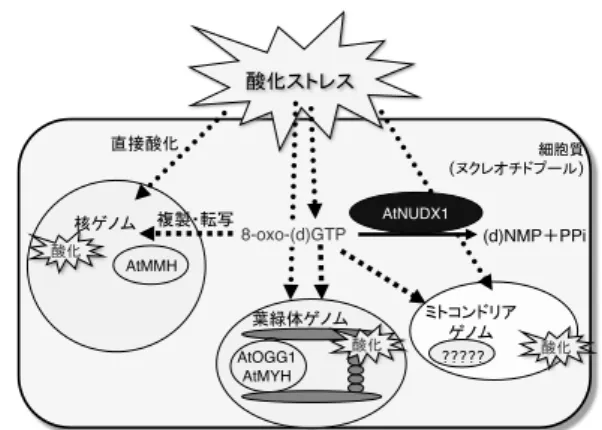
① DNA/RNA 酸化 傷害に対 する防御 機構 AtNUDX の分子特性から、シロイヌナズナに は細胞質局在型の AtNUDX1 が唯一の酸化ヌク レオチド加水分解酵素として存在すること が示された。大腸菌を用いた相補試験の結果、
AtNUDX1 は大腸菌mutT欠損株における DNA の 突然変異および mRNA の転写エラー発生頻度 を抑制した。また、シロイヌナズナ nudix1 破壊株では、正常および酸化ストレス条件下 での酸化塩基(8-oxo-dG)量が顕著に増加し ていた(図 3)。これらのことから、AtNUDX1 は細胞質のヌクレオチドプールの浄化によ り、酸化ヌクレオチドの核、ミトコンドリア および葉緑体ゲノムへの取り込みを防いで いることが示された(図 4)(Yoshimura et al.
Plant Cell Physiol. 2007) 。
WT! KO-nudx1 (T5!")!
7 days!
0 day!
PQ treatment!
(3 µM)!
WT! KO-nudx1!
8-oxo-dG / 106 dG!
0!
1!
2!
0 day after the ! PQ treatment!
14 days after the!
PQ treatment!
0.5!
1.5!
2.5! a!
b!
b!
c!
図 3. AtNUDX1破壊株 (KO-nudx1)の酸化スト レス下でのゲノム中の 8-oxo-dG 蓄積 播種 7 日後の植物体を 3 µM パラコートを含 む培地で 7-14 日間栽培した。
!"#$!
AtOGG1!
AtMYH!
%&'"#$! ()*+,-.!
//"#$!
!!!!!! 01!
01!
01!
ヌクレオチドおよびDNAの酸化損傷とその修復機構
(d)NMP2PPi!
013)43!
8-oxo-(d)GTP!
AtNUDX1!
567/!
"894:;,<=>#!
?@01!
AtMMH!
ABCDE!
図 4. シロイヌナズナにおけるヌクレオチド および DNA の酸化損傷とその修復機構 AtMMH,AtOGG1,AtMYH:酸化塩基除去修復酵素
②ADP-ribose/NADH 代謝によるヌ クレオ チ ド リ サ イ ク ル お よ び poly (ADP-ribosyl)ation 反応の制御
シロイヌナズナには ADP-ribose および NAD(P)H pyrophosphatase 活性を示す Nudix hydrolase アイソザイム(細胞質型 AtNUDX2, 6, 7, 10、ミトコンドリア型 AtNUDX14、葉緑 体型 AtNUDX19)が存在することを示した。興 味 深 い こ と に 、 ADP/NADH 特 異 的 AtNUDX
(AtNUDX2 および 7)の発現は酸化ストレス 耐性能と相関していた(図 5)。過剰発現株お よび発現抑制株における ADP-ribose, NADH, NAD+, ATP, poly(ADP-ribose)量の解析の結果
(図 6, 7)、AtNUDX2 はポリ(ADP-リボシル) 化サイクルから生成する遊離 ADP-リボース のリサイクルにより、細胞内エネルギー状態 を維持することで酸化ストレス耐性に寄与 していることが示された(図 8)(Ogawa et al.
Plant J. 2009)。
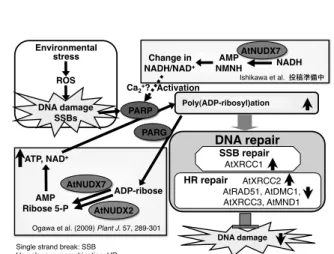
また、AtNUDX7 は生体内で AtNUDX2 とは異 なり ADP-ribose だけでなく、NADH 代謝を介 した PAR 反応制御にも関与することで酸化ス トレス防御に機能していることが示唆され た(図 8)。さらに、その PAR 反応制御により、
DNA 一本鎖損傷および DNA 相同組換え修復因 子の発現量が制御されていた(Ishikawa et al. in preparation )。
Cytosol NADPH
AtNUDX19 Chloroplast
Photosynthesis Fatty acid biosynthesis
β-oxidation Peroxisome Acyl-CoA
Acetyl-CoA etc. AtNUDX15a
ADP-ribose AtNUDX2, 10?
Acetyl-CoA etc.
AtNUDX11
(ADP-ribosyl)ation reaction AtNUDX7 NADH
AtNUDX6 8-oxo-(d)GTP
AtNUDX1
Mitochondrion Acetyl-CoA
Succinyl-CoA etc. AtNUDX15 ADP-ribose AtNUDX14
TCA cycle (Oxidative phosphorylation)
ApnA AtNUDX26, 27 FAD
AtNUDX23
ApnA AtNUDX25 AtNUDXsの細胞内局在性と機能
!"#! AtNUDX2$%&'#! AtNUDX7$%&'#!
AtNUDX2, 7$%&'#()*AtNUDX6, 7+,#-PQ./0!
!"#! AtNUDX7+,#! AtNUDX6+,#!
図 5. AtNUDX2, 7 過剰発現株および AtNUDX6, 7 破壊株のパラコート耐性能
上段: 播種 7 日後の植物体を 3 µM(上段)も しくは 2 µM(下段)パラコートを含む培地で 7 日間栽培し、その後通常培地でさらに 7 日 間栽培した。
0!
2!
4!
6!
8!
10!
12!
14!
16!
18!
b! b!
b!
b!
c! c!
c! c!
d! d! d! d!
e! e!
ef! ef!
f! f!
a!a!a!
0!
1!
2!
3!
4!
5!
6!
7!
ADP-ribose (nmol/g FW)! a!
a!
b!
b!
d!d! d!d!
Ct! OX-1!OX-2!RNAi!OX-1!OX-2!KO!
AtNUDX7!
AtNUDX2!
a!
c!c! c! c! c!
0 day after the PQ treatment!
3 days after the PQ treatment!
7 days after the PQ treatment!
NADH (nmol/g FW)!
Ct!OX-1!OX-2!RNAi!OX-1!OX-2!KO!
AtNUDX7!
AtNUDX2!
酸化ストレス下におけるADP-ribose/NADH量
図 6. 酸 化 ス ト レ ス 下 に お け る ADP-ribose/NADH 量の変化
播種 7 日後の植物体を 3 µM パラコートを含 む培地で 3-7 日間栽培した。
0!
5!
10!
15!
20!
25!
30!
35!
40!
45!
50!
0!
1!
2!
3!
4!
5!
6!
7!
8!
9!
10!
!! !!!!!! !!!!!! !!
"!"!"! "!
"!"!
#!#! #!#!
!!
"!
"!
0!
50!
100!
150!
200!
250!
300!
Ct!OX-1!OX-2!
AtNUDX2!
OX-1!OX-2!
AtNUDX7!
RNAi! KO!
!! !!
"! "! "!"!
"! "!
#! #! #! #!
$! $! $! $! $!
%!%!
"!"!
0 day after the ! PQ treatment!
3 days after the ! PQ treatment!
7 days after the ! PQ treatment!
Ct!OX-1!OX-2!
AtNUDX2!
OX-1!OX-2!
AtNUDX7!
RNAi! KO! Ct!OX-1!OX-2!
AtNUDX2!
OX-1!OX-2!
AtNUDX7!
RNAi! KO!
ATP (nmol/g FW)!
NAD+ (nmol/g FW)! poly (ADP-ribose) (%)!
"!
"! ""!
"!
"! "!"!
"!"!
"!"!"! "! "!
#!#! #!
!!
!!!!
酸化ストレス下におけるpoly (ADP-ribose)、NAD+およびATP量の変化
図 7. 酸 化 ス ト レ ス 下 に お け る poly (ADP-ribose)、NAD+および ATP 量の変化 播種 7 日後の植物体を 3 µM パラコートを含
む培地で 3-7 日間栽培した。
Environmental!
stress!
ROS!
DNA damage!
SSBs!
Poly(ADP-ribosyl)ation!
ATP, NAD+!
ADP-ribose!
AtNUDX7!
AtNUDX7 ! Change in !
NADH/NAD+!
AtXRCC1!
AtXRCC2!
AtRAD51, AtDMC1, ! AtXRCC3, AtMND1 ! SSB repair! HR repair!
DNA damage!
Ca2+?!
DNA repair!
AMP!
Ribose 5-P!
Activation!
NADH!
AMP!
NMNH!
Single strand break: SSB!
Homologous recombination: HR!
Ishikawa et al. !"#$%!
PARP!
PARG!
AtNUDX2!
Ogawa et al. (2009) Plant J. 57, 289-301!
図 8. 酸化ストレス下における AtNUDX2 およ び 7 の役割
③ NADPH、CoA および FAD の代謝制御
・ シ ロ イ ヌ ナ ズ ナ 野 生 株 の 葉 に お い て AtNUDX19 に 由 来 す る NADPH pyrophosphohydrolase 活性が認められた。ま た、nudix19 破壊株では細胞内 NADPH 量が増 加していた(図 9)。
・シロイヌナズナ遺伝子破壊株の解析から、
AtNUDX11 お よ び AtNUDX15 は 全 CoA pyrophosphohydrolase 活性の約 20%および 50%に寄与していることが示された(図 10)。
・ シロイヌナズナ葉の粗酵素液において、
FAD pyrophosphohydrolase 活性(135.3 5.8 nmol/min/mg protein)が検出された。また、
イオン交換クロマトグラフィーにより、リコ ンビナントタンパク質と同様の活性を示す、
単一のピークが認められた。AtNUDX23 発現抑 制 株 ( KD-nudx23 ) お よ び 過 剰 発 現 株
(OE-NUDX23)における FAD 加水分解活性は、
コントロール株と比較してそれぞれ 60%およ び 150%であった(図 11)。そこで、RF、FMN お よ び FAD レ ベ ル を 測 定 し た と こ ろ 、 OE-AtNUDX23 株のみではなく、KD-nudx23 株 においても各々の化合物レベルが減少して いた。
これらの結果から、AtNUDX11, 15, 19, 23 は細胞内での CoA, NADPH, FAD の代謝制御に 重要な役割を果たしていることが示唆され た。
* atg
KO-nudx19!
KD-nudx19!
AtNUDX19!
AtNUDX19!
Actin 8!
AtNUDX19遺伝子破壊株の単離
20!
15!
10!
0!
25!
NADPH pyrophosphohydrolase activity! (nmol/min/mg protein)!
5!
図 9. AtNUDX19 遺伝子破壊株(KO-nudx19)
の作出
T-DNA 挿入位置(左上)、RT-PCR(左下段)、
活性(右)
T-DNA insert!
AtNUDX11 (SALK_037735)!
100 bp!
AUG!
1! UGA!
933!
AtNUDX15 (SAIL_1255_G04)!
AUG!
1! UGA!
1473!
T-DNA insert!
AtNUDX11, 15破壊株 (KO-nudx11, 15)の解析
RT-PCR
WT! KO-!nudx11!
KO-!
nudx15!
Actin2!
AtNUDX15!
AtNUDX11!
CoA pyrophosphohydrolase活性 (%)
WT! KO-nudx11! KO-nudx15! 100!
46.2!
80.4!
Activity
図 10. AtNUDX11 および 15 遺伝子破壊株
(KO-nudx11, 15)の作出
T-DNA 挿入位置(左上)、活性(左下段)、RT-PCR
(右)
!"#$%&'()
!*+,)-)
!"#$%&'()
!"#$%&'(%)(*%)()&+'(,-*.$-/0123&$45$(6$/(73'(,8$
9::$
:$
;<:$
;::$
<:$
AtNUDX23過剰発現/発現抑制株の作出
RT-PCR!
Western!
図 11. AtNUDX23 遺 伝 子 発 現 抑 制 株
(KD-nudx23)の作出
RT-PCR(左上段)、ウエスタンブロット(左 下段)、活性(右)
5.主な発表論文等
(研究代表者、研究分担者及び連携研究者に は下線)
〔雑誌論文〕(計 8 件)
1. Ogawa, T., Ishikawa, K., Harada, K., Fukusaki, E., Yoshimura, K. and Shigeoka, S. (2009) Overexpression of an ADP-ribose pyrophosphatase, AtNUDX2, confers enhanced tolerance to oxidative stress onArabidopsis plants.(有) Plant J. 57, 289-301
2. Ogawa, T.*, Yoshimura, K.*, Miyake, H., Ishikawa, K., Ito, D., Tanabe, N. and Shigeoka, S. (2008) Molecular characterization of organelle-type Nudix hydrolases in Arabidopsis thaliana.( 有 ) Plant Physiol. 148, 1412-1424 *: These authors contributed equally.
3. Ogawa, T., Yoshimura, K., Shigeoka, S.
(2008) Functional analysis of an 8-oxo-7,8-dihydro-2'-deoxyguanosine 5'-triphosphate pyrophosphohydrolase, AtNUDX1, involved in repair of oxidative DNA damage in Arabidopsis thaliana.(無) Photosynthesis. Energy from the Sun 1323-1326
4. Yoshimura, K., Ogawa, T., Ueda, Y. and Shigeoka, S. (2007) AtNUDX1, an 8-oxo-7,8-dihydro-2ʼ-deoxyguanosine 5ʼ -triphosphate pyrophosphohydrolase, is responsible for eliminating oxidized nucleotides in Arabidopsis. (有) Plant Cell Physiol. 48, 1438-1449
5. Tanabe, N., Yoshimura, K., Kimura, A.
, Yabuta, Y. and Shigeoka, S.(2007)D ifferential expression of alternativel y spliced mRNAs of Arabidopsis SR prot ein homologues, atSR30 and atSR45a, in response to environmental stress. (有 ) Plant Cell Physiol. 48, 1036-1049 6. Yabuta, Y., Mieda, T., Rapolu, M., N
akamura, A., Motoki, T., Maruta, T., Y oshimura, K., Ishikawa, T. and Shigeok a, S. (2007) Light regulation of as corbate biosynthesis is dependent on t he photosynthetic electron transport c hain but independent of sugars in Arab idopsis. (有) Journal of Experimen tal Botany 58, 2661-2671
7. 吉村和也、薮田哲行、石川孝博、重岡 成 (2007)レドックス制御に関わるアスコル ビン酸およびヌクレオシド二リン酸類縁体.
(Nudix)(無)ビタミン 81, 83-93 8. 薮田行哲、小川貴央、吉村和也、重岡 成
(2007) 植物のレドックス制御機構と環境 ストレス応答への関与. (無) 蛋白質 核 酸 酵素 52, 578-584
〔学会発表〕(計 22 件)
1. 石 川 和 也 ら シ ロ イ ヌ ナ ズ ナ Nudix hydrolase, AtNUDX23 による FAD 代謝の制 御機構 2009 年 3 月 27-29 日 農芸化学学 会 2009 年度大会(福岡)
2. 吉村和也ら 2009 年 3 月 27-29 日 植物 における酸化ヌクレオチド蓄積の防御機 構の解明 農芸化学学会 2009 年度大会
(福岡)
3. 石 川 和 也 ら シ ロ イ ヌ ナ ズ ナ ADP-ribose/NADH pyrophosphohydrolase, AtNUDX7, に よ る poly (ADP-ribosyl)ation 反応の制御が酸化ス トレス耐性に及ぼす影響 2009 年 3 月 21-24 日 第 50 回日本植物生理学会年会
(名古屋)
4. 伊藤大輔ら CoA 及びその誘導体の代謝に おける CoA pyrophosphohydrolase の役割 2009 年 3 月 21-24 日 第 50 回日本植物生 理学会年会(名古屋)
5. 伊 藤 大 輔 ら シ ロ イ ヌ ナ ズ ナ CoA pyrophosphohydrolase による CoA 及びそ の誘導体の代謝制御 2008 年 12 月 8-12 日 第 31 回日本分子生物学会年会・第 81 回日本生化学会大会 合同大会(神戸)
6. 石川和也ら 酸化ストレス下でのシロイ ヌ ナ ズ ナ ADP-ribose/NAD(P)H pyrophosphatase に よ る poly (ADP-ribosyl)ation 反応の制御 2008 年 12 月 8-12 日 第 31 回日本分子生物学会年 会・第 81 回日本生化学会大会 合同大会(神 戸)
7. 伊 藤 大 輔 ら シ ロ イ ヌ ナ ズ ナ CoA pyrophosphohydrolase が CoA 及びその誘 導体の代謝に及ぼす影響 2008 年 9 月 13 日 日本農芸化学会 関西支部大会(京都)
8. 吉村和也ら オルガネラにおけるヌクレ オシド-2 リン酸類縁体の代謝制御 日本 ビタミン学会 第 60 回大会 2008 年 6 月 13-14 日(仙台)
9. 石川和也ら 植物 ADP-リボース/NADH ピ ロホスファターゼの酸化ストレス下にお ける生理機能 2008 年 6 月 13-14 日 日本 ビタミン学会 第 60 回大会(仙台)
10. 小川貴央ら シロイヌナズナ葉緑体局在 型 Nudix hydrolase によるヌクレオシド 2- リン酸類縁体の代謝 2008 年 3 月 26-29 日 農芸化学学会 2008 名古屋大会(名古 屋)
11. 吉村和也ら シロイヌナズナミトコンド リア局在型 Nudix hydrolase によるヌクレ オシド 2-リン酸類縁体の代謝 2008 年 3 月 26-29 日 農芸化学学会 2008 名古屋大 会(名古屋)
12. 石川和也ら 酸化ストレス下におけるシ ロイヌナズナ Nudix hydrolase による ADP- リボース/NADH の代謝制御 2008 年 3 月 26-29 日 農芸化学学会 2008 名古屋大会
(名古屋)
13. 吉村和也ら シロイヌナズナ ADP-リボー ス/NADH ピロホスファターゼの酸化ストレ ス耐性への寄与 2008 年 3 月 20-22 日 第 49 回日本植物生理学会年会(札幌)
14. 小川貴央ら シロイヌナズナ葉緑体での
Nudix hydrolase による NADPH および FAD の代謝制御 2008 年 3 月 20-22 日 第 49 回日本植物生理学会年会(札幌)
15. 伊 藤 大 輔 ら シ ロ イ ヌ ナ ズ ナ CoA pyrophosphohydrolase の分子特性と機能 解析 2008 年 3 月 20-22 日 第 49 回日本 植物生理学会年会(札幌)
16. 吉村和也ら シロイヌナズナ AtNUDX1 に よる DNA/RNA 酸化損傷に対する防御機構 2007 年 12 月 11-14 日 第 30 回日本分子生 物学会年会・第 80 回日本生化学会大会 合 同大会(横浜)ワークショップ
17. 小川貴央、伊藤大輔、石川和也、吉村和 也、重岡成 2007 年 12 月 11-14 日 植物 に お け る オ ル ガ ネ ラ 局 在 型 Nudix hydrolase によるヌクレオシド 2-リン酸類 縁体の代謝 第 30 回日本分子生物学会年 会・第 80 回日本生化学会大会 合同大会(横 浜)
18. 小 川 貴 央 ら シ ロ イ ヌ ナ ズ ナ 8-oxo-(d)GTP pyrophosphohydrolase, AtNUDX1 によるヌクレオチドプールの浄化 2007 年 9 月 21-22 日 日本農芸化学会 関 西支部中部支部合同大会(愛知)
19. Yoshimura, K. et. al. Functional
analysis of Arabidopsis Nudix hydrolases under oxidative stress. ROS in Plants 2007. A SFFR Plant Oxygen Group meeting on reactive oxygen and nitrogen species. 2007.9.12-14 Signalling &
Metabolism, Oxidative stress, Antioxidants (September 12, 13 & 14 2007, Ghent, Belgium)
20. Ogawa,T. et.al. Functional analysis of an 8-oxo-7,8-dihydro-2'-deoxyguanosine 5'-triphosphate pyrophosphohydrolase, AtNUDX1, involved in repair of oxidative DNA damage in Arabidopsis thaliana.
2007.7.22-27 14th Photosynthesis Congress(22nd to 27th July 2007, at the SECC in Glasgow UK )
21. 吉村和也ら シロイヌナズナのオルガネ ラ局在型 Nudix hydrolase によるヌクレオ シド 2-リン酸類縁体の代謝 2007 年 5 月 24-25 日 日本ビタミン学会 第 59 回大会
(長崎)
22. 小川 貴央ら シロイヌナズナ細胞質型 ADP-リボース/NADH ピロホスファターゼの 生理機能の機能解析 2007 年 5 月 24-25 日 日本ビタミン学会 第 59 回大会(長崎)
6.研究組織 (1)研究代表者
重岡 成(SHIGEOKA SHIGERU)
近畿大学・農学部・教授 研究者番号:80140341 (2)研究分担者
吉村和也(YOSHIMURA KAZUYA)
中部大学・応用生物学部・講師 研究者番号:90379561