東北日本におけるクマタカの繁殖と 分布に影響する要因
Factors affecting the reproduction and distribution of Mountain Hawk-Eagles (Nisaetus nipalensis) in Northeast Japan
岩 手 県 立 大 学 大 学 院 総 合 政 策 研 究 科
平成
25年
3月満期退学
氏
名: 葉 山 雅 広
指導教員: 金 子 与 止 男
目 次
第Ⅰ章 序論 ... 1
1.背景と目的 ... 1
2.本論文の構成 ... 4
第Ⅱ章 クマタカの生息地を評価する上での課題 ... 6
1.繁殖成功率に影響する要因 ... 6
2.潜在的分布適地に関する要因 ... 9
3.クマタカの生態と本研究で用いた用語について ... 11
1)分類 ... 11
2)分布 ... 11
3)繁殖 ... 11
4)行動域の広さ... 11
5)食性 ... 12
6)使用した用語... 12
第Ⅲ章 繁殖成功率に影響する環境要因 ... 14
1.はじめに ... 14
2.調査地と方法 ... 15
1)調査地 ... 15
2)繁殖状況の確認 ... 16
3)ブナの結実状況 ... 17
4)林相・土地利用 ... 17
5)気象 ... 18
6)統計解析 ... 18
3.結果 ... 22
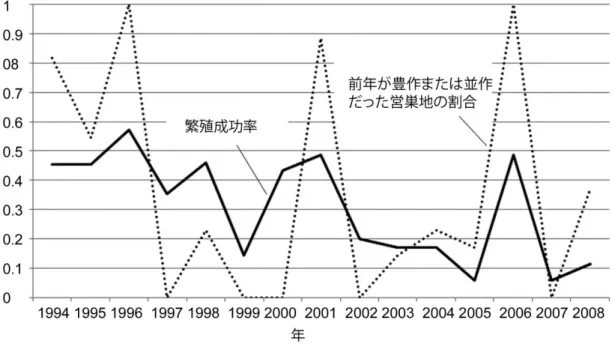
1)営巣地間の繁殖成功率等の比較 ... 22
2)繁殖成功率の年変化 ... 22
3)ブナの結実状況とクマタカの繁殖結果との関係 ... 23
4)クマタカの繁殖結果に影響する要因 ... 27
5)結実状況の違いによる繁殖結果に影響する要因の比較 ... 27
6)結実状況の違いによる繁殖成功率と景観の多様度との関係 ... 29
4.考察 ... 31
第Ⅳ章 分布に影響する環境要因 ... 36
1.はじめに ... 36
2.調査地と方法 ... 37
1)調査地 ... 37
2)解析に利用したデータセット ... 37
3)解析方法 ... 42
3.結果 ... 43
1)生息区での傾斜と植生別面積の関係 ... 43
2)クマタカの分布に影響している要因 ... 45
3)多雪地と少雪地との比較 ... 45
4.考察 ... 49
1)クマタカの分布に影響している要因 ... 49
2)多雪地と少雪地との比較 ... 50
第Ⅴ章 結論 ... 53
1.総合考察 ... 53
2.結論 ... 57
謝辞 ... 59
引用文献 ... 60
1
第Ⅰ章 序論
1.背景と目的
本研究は、森林性の猛禽類であるクマタカ Nisaetus nipalensis の生息地を保全するため に必要な情報を明らかにすることを目的としている。クマタカはインド亜大陸の南西部と セイロン島、ユーラシア大陸の東南部、インドシナ半島の北部から中央部と東部、マレー 半島北部、台湾島、日本列島に分布する猛禽類である。IUCNのレッドリスト(IUCN 2014)
では、クマタカは分布域が20,000km2よりも広く、個体数に減少傾向がみられるがVulnerable
(危急)とするほど急速な減少(10年間または3世代の間に30%以上減少)ではなく、個体 群もVulnerable の水準(成熟個体が10,000以下で10年間または3世代の間に10%以上減少し 続けている)に近づいているとは思われないとして、Least Concern(低懸念)とされている。
国内には亜種 N. n. orientalisが北海道から九州の山地の森林に生息し(森岡ら1998,日本鳥 学会 2012)、日本国内の最小推定個体数は約2,000羽と推定されている(環境省2012)。環境 省のレッドリスト(環境省2006)では絶滅危惧ⅠB類として掲載されており、「絶滅のおそ れのある種の保存に関する法律」により国内希少野生動物種に指定されている。クマタカ は行動圏が広いことからダムや高速道路の建設など、山地での大型事業ではクマタカに関 する調査や保全対策の立案が必要となることが多い。そのため、クマタカは絶滅危惧種の 鳥類の中ではイヌワシ Aquila chrysaetos と並んで比較的多く調査がされている種であり、
事業によっては影響の予測やミチゲーションのために営巣適地や採食環境の推定が行われ ているケースもある。クマタカの分布予測モデルなどは事業箇所の代替地を計画する際な ど、戦略的アセスメントにおいても有益である。これまでに、潜在的な分布予測に関して は北海道(鈴木ら2001)、山形県(杉山ら2009)、岩手県(伊藤ら2004)、鳥取県(伊藤ら2012)
で行われた先行研究がある。また、クマタカは広い行動圏をもつことからアンブレラ種と して着目され、国有林においては繁殖活動を阻害しないように施業の実施時期と位置に配 慮するとともに、長期的な視点から多様な種が生息できるよう林種転換と林分配置を検討 した地域もある。このように、クマタカは我が国の環境アセスメントにおいては常に注目 される種であるが、生息地が山間部であり行動域が広いため例数を確保するためには長期 間を要すること、繁殖が厳寒期から開始されることなどから基礎的な生態については不明 な点も多く、近年になってようやく個体識別や電波発信機による調査が端緒についたばか りである。また、猛禽類に関しては、質の低いテリトリーは質の高いテリトリーに比べ、
占有率が低く、遺棄される確率が高いという結果が示されており(Korpimäki 1988,Newton
& Marquiss 1991,Kostrzewa 1996)、その種を保護する上で生息地の評価は重要である。そ
のため、クマタカの基礎的な生態を明らかにすること、そして、その生息地の評価をより 適正に行う手法を検討することは、学術的な意義を有するとともに社会的な要請に応える 研究である。
そこで本研究では解決すべき課題として以下の2つを設定した。
2
①クマタカの繁殖成功率に影響する要因を明らかにすること
クマタカの繁殖成功率は近年低下している傾向があるとされる(環境省2012)(表 1)。 その原因として、しばしば生息環境の悪化が指摘されるものの、実際にデータに基づき検 討した事例はきわめて少ない(飯田ら2007)。そのため、クマタカの基礎的な生態を明らか にするために、クマタカの繁殖成否に影響すると思われるさまざまな要因を用いて検討を 行った。これまでに世界各地で多くの猛禽類を対象に繁殖結果を左右する要因に関する研 究が行われてきたが、年によって変化する食物量や気象などの影響を加味した研究例は少 数である。クマタカはアカネズミApodemus speciosusなどのネズミ類やニホンリスSciurus lis、
ヤマドリSyrmaticus soemmerringii、ノウサギ Lepus brachyurus、アオダイショウ Elaphe climacophoraなどを捕食する(森岡ら1998)。ブナFagus crenata林においては、これらの種の 多くはブナの種子生産量に生息密度が左右されていると思われる。そのため本研究では、
食物量の指標となるブナの豊凶と降水量などの年によって変化する変数を用いて、繁殖地 の質の違いが繁殖結果とどのように関係しているのかを解析した。分析の対象となる単位 は営巣地であり、個々のつがいの年ごとの繁殖成否に影響した可能性がある要因として、
林相別面積、気象条件、食物の多寡、前年の繁殖成否を調査し、解析を行なった。
表 1 繁殖成功率の変化
地域 年 繁殖成功率 出典
西中国山地 1981~1985 (n=21) 85.7% 環境省(2012)より広島ク マタカ生態研究会(未発表)
の結果を間接引用
1996 (n=12) 8.3%
山形県 2003 (n=24) 22.7% 杉山ら(2009)より猛禽類
調査会(2006)の結果を間 接引用
2004 (n=24) 18.1%
2005 (n=24) 16.6%
②クマタカの分布に影響する要因を明らかにすること
クマタカは森林性の鳥類であるが、どのような森林に繁殖分布しているのかを分析し、
これまでの先行研究で示されてきた結果と比較した。先行研究ではクマタカの分布には特 に地形が深くかかわっていることが示されてきた一方、植生に関してはあまり関係が示さ れていなかった。そのため、本研究では調査対象地を東北全域という広範囲とし、クマタ カのより一般的な分布特性を明らかにすることを目指した。また、クマタカの分布に影響 する要因の集計範囲を、営巣木の周辺から行動域全体を網羅すると考えられる範囲まで、
500mずつ拡大し、どのような違いが示されるのかに着目した。さらに、東北地方において は、雪の存在は地形とともに植生に対して強い影響を及ぼしており、降雪量によって森林 の樹種構成や景観が異なっている。したがって、降雪量の多寡によってクマタカの分布に
3
作用する要因は異なっていることが予測され、その検証を試みた。
また、クマタカは九州のイスノキ Distylium racemosum やアカガシ Quercus acuta などか ら構成される常緑広葉樹林から、北海道のミズナラ Quercus crispulaやエゾマツ Picea jezoensisなどが混生する森林など、多様な森林に分布している。食物としてはアカネズミな どのネズミ類、ニホンリス、ノウサギ、イタチ Mustela itatsiなどの小~中型哺乳類、アオダ イショウなどの爬虫類、ヤマドリなどの中型鳥類など、生息環境に応じて多様な種を採食 している。そのため、クマタカは地域によって適応的な行動をとっており、繁殖や分布に 影響する要因も異なっていることが予見される。そこで、本研究では東北地方に生息する クマタカを一つの個体群と想定して考察を行うこととした。なお、東北地方の特色の一つ である多雪であるということと、広大な森林地帯とその中心をなすブナ林の存在に注目す ることにより、東北地方の生態系のユニークな一面を明らかにすることも本研究をすすめ る上での副テーマとした。
4 2.本論文の構成
本論は次の構成でとりまとめた(図 1)。まず、第Ⅱ章では本研究と関係する先行研究の 要点と問題点の整理を行ない、本研究において解明すべき点を示した。また、本研究の対 象であるクマタカの生態に関して、本論を読んでいただく上で必要な事項を文献から引用 し、とりまとめた。
第Ⅲ章では、クマタカの繁殖結果に影響する要因を検討した。検討にあたって、東北地 方に広くみられるブナ林に生息するクマタカを調査対象とした。これは、ブナの結実が年 によって大きく変動することが知られており、このブナ林に生息するクマタカの繁殖状況 を調査することは、ひいてはブナ林に生息する多くの動物が関係するブナ林の生態系の特 徴を明らかにすることにも通じるものである。従来から、どのような行動圏をもつつがい の繁殖成功率が高いのかという研究は行われてきた(Newton 1991)。しかし、年によって変 動する要因(例えば、数年おきに大発生する北欧のネズミ類、北米の灌木林地帯の干ばつ など)に着目してその行動圏の環境を評価した事例は数少ない。一般的に好適と考えられ るなわばりを保持するつがいは、いつも多くの雛を巣立たせているのだろうか、それとも、
ある年は繁殖に有利な環境が別の年にはあまり利点が無い環境になってしまうことはない のだろうか?この疑問を解くことを本章のテーマとした。
第Ⅳ章では、クマタカの分布に影響している要因を検討した。クマタカの潜在的な分布 を予測するためのモデルは、都道府県といった行政界を単位としたものが従来多かったが、
クマタカの生態を明らかにするうえでは行政界にこだわらず、より多くのサンプルを用い て環境の比較を行うことが望ましいと考える。そのため、本研究では林野庁東北森林管理 局と関東森林管理局の協力により、東北6県のクマタカの営巣地の分布情報を基に分布予測 モデルを作成し、先行研究との対比を行なった。また、先行研究においては、地形が最も クマタカの分布に影響していることが示されており、植生に関連した要素の影響は比較的 弱いものであった。しかし、この結果には、一つの都道府県といった比較的同質な環境に おいてモデルを作成していることも影響しているのではないかと考えられる。クマタカの 生息環境である森林は、地形と土壌、気候、人為的活動などにより形作られたものであり、
東北地方のような降雪地域では降雪によって植生は大きな影響を受けていることが知られ ている。そのため、クマタカの生息環境を形成する森林は、地形とともに降雪の影響を受 けていると考えられることから、特に降雪量の多寡に注目し、クマタカの分布に関連する 要因の違いを検討した。
第Ⅴ章では総合考察を行い、今後の課題を整理した。そして、生息地の評価を行う上で 考慮すべき点などを整理し、結論とした。
5
図 1 本論文の構成 第Ⅴ章 結論
第Ⅲ章 繁殖成功率に影 響する環境要因
第Ⅳ章 分布に影響する 環境要因
第Ⅱ章 クマタカの生息地を評価する上での課題
第Ⅰ章 序論
6
第Ⅱ章 クマタカの生息地を評価する上での課題
1.繁殖成功率に影響する要因
猛禽類の繁殖に生息地の植生や景観要素が影響する事例は数多く報告されており、森林の タイプや土地利用区分ごとの面積割合(Widén 1989,Kostrzewa 1996,Sim et al. 2001,由井 ら2005,Greenwald et al. 2005,Moser & Garton 2009)、林縁長(Franklin et al. 2000)、森林の 断片化(Redpath 1995)、地形的要因(Lopez-Lopez et al. 2006)、土地利用の変化(Widén 1997)
などとの関係が報告されている。また、これらの要因は単独で作用するだけではなく、他 の要因と互いに関連しながら影響している(Sonerud 1986,Franklin et al. 2000,McClaren et al.
2002,Gil-Sáncheza et al. 2004,Sergio et al. 2006,Byholm et al. 2007)。我が国でもクマタカ やイヌワシの繁殖成功率の低下と植生の変化との関係が指摘されている(飯田ら2007,由 井ら2005)。
しかし、つがいが巣立たせた雛の多さをハビタットの質で説明できた事例と、その他の要 因のほうがより強く影響していた例がある。気象条件など年毎によって変化する要因が猛 禽類の繁殖に影響を与えていることはこれまでも数多く報告されており、土地利用よりも 降雨量などの気象条件のほうがより強く作用していた例も多い(McDonald et.al 2004,
Newton1986,Mearns & Newton 1988,Kostrzewa 1989,Olsen& Olsen 1989,Kostrzewa
& Kostrzewa 1990,Potapov 1997,Dawson & Bortolotti 2000, 2002,Rodríguez &
Bustamante 2003)。例えば、オオタカ Accipiter gentilis では天候は直接的な影響だけで
はなく、狩り行動や食物量を通して間接的に繁殖結果へ影響をもたらす可能性も示されて いる(Fairhurst & Bechard2005)。アメリカチョウゲンボウ Falco sparverius に関する調 査では、繁殖の成功には獲物の多さよりも天候の変動が影響していることが示され、天候 は食物の量ではなく食物の得やすさに影響し、親による雛への給餌活動に影響を及ぼして いた(Dawson & Bortolotti 2000)。そして、オオタカの巣立った雛数とハビタットの特徴 を調査したMcClarenら(2002)は、ハビタットの特徴などの空間的なパターンよりも地域 的な天候や餌動物の生息数の変動といった時間的パターンのほうがオオタカの繁殖に影響 していることを示した。
餌動物の生息数が変動することによる影響に関しては、北米のオグロジャックウサギ Lepus californicusや北欧のネズミ類 Microtus agrestis、M. rossiaemeridionalis、Clethrionomys
glareolus など、生息数が年によって大きく変動する種を利用している猛禽類はその繁殖に
大きな影響をうけていることが明らかにされている(Steenhof et al. 1997,Korpimäki 1994,
Salafsky et al. 2005,Rutz & Bijisma 2006,Millon et al. 2010)。さらに、このような環境下で 繁殖するつがいは、食物の多い年と少ない年とで繁殖成功率に強く作用する要因が異なる こと(Steenhof et al. 1997)、食物が少ない年は多い年よりも植生や景観要素の影響がより強 く表れることが示されている(Hakkarainen et al. 2003,Lõhmus 2003)。例えば、オグロジャ ックウサギの生息数がピークの年では、イヌワシのつがいあたりの巣立ち雛数は5月と6月
7
の高温の日数と負の相関があった一方、ウサギの生息数が低い局面では冬季の厳しさが最 も効いており、5月と6月の暑さはその次に影響していた(Steenhof et al. 1997)。エストニア のノスリ Buteo buteo では、ハビタットの違いによる巣立雛数への影響は、ネズミ類が多い 年よりも少ない年において比較的強く示され、ネズミ類が少ない年は不均質な景観の生息 地のつがいの生産力(巣立った雛の数)が高く、ネズミ類の数がピークの時にはネズミが 好む環境を広く含んでいる均質な景観の生息地のつがいで高い生産力が示された(Lõhmus
2003)。キンメフクロウ Aegolius funereus でも、生息地の景観構成はネズミ類の増加相より
も減少局面においてより強く繁殖成功率に影響していた。ただし、ネズミ類が少ない年に は、繁殖するキンメフクロウも少なくなり、繁殖成績は景観構成による影響をあまり受け ていなかった(Hakkarainen et al. 2003)。
このように、年によって変動する要因は猛禽類の繁殖に大きな影響を与えており、その生 息地を評価する上でも重要であるにもかかわらず、そこに着目した研究はごく少数である
(Lõhmus 2003)。クマタカの繁殖成否に影響する要因を探求するにあたっても、植生や気 象などの要因に加えて食物量の年変化を加味して検討することは非常に重要であると考え る。しかしながら、クマタカが食物として利用する種は幅広く、アカネズミ、ニホンリス やノウサギなどの小型から中型の哺乳類、ヤマドリなどの小型から大型の鳥類、アオダイ ショウなどのヘビ類である(井上・山崎 1984,森岡ら 1998,布野ら 2000,柏原・安田 2004,
Kaneda 2009)。そのうえ、アオダイショウのように個体数推定自体が難しい種もあることか
ら食物量の年ごとの変化を把握することは困難である。そこで、ブナ林に生息するクマタ カの個体群を対象に、ブナの豊凶を食物量の指標として利用することを検討した。ブナ林 では数年に一度、豊作と呼ばれる種子生産量が極端に大きくなる年がある一方、それ以外 の年はほとんど種子生産を行わない(前田 1988,Suzuki et al. 2005,Masaki et al. 2008)。ブ ナ以外にも、ナラ類、シデ類などの種では年によって種子生産の変動が大きく、地域的に 同調して変動する豊作が数年の間隔で発生することが知られている(Kelly 1994)。そのよう な森林では、ドングリという資源の変動が第1の波となり、その後にネズミ類などのドン グリの消費者の増加(第2の波)が起き、ジェネラリストの捕食者によるネズミ類への反応 が変化する(第3の波)カスケード効果が知られている(Schmidt and Ostfeld 2008)。ニ ュージーランドではナンキョクブナ科の樹木が種子を落した後、ネズミ類の密度が極端に 上昇したことに伴い、その捕食者であるオコジョ Mustela erminea が急増することが観察さ れている(King 1983,Murphy & Dowding 1995)。北アメリカでは、ブナやカシなど結実に 豊凶がある森林での種子生産とげっ歯類の生息数の変動が、げっ歯類に卵を食べられる鳴 禽類の個体群のサイズ(Schmidt 2003,Clotfelter et al. 2007,Schmidt et al. 2008,Schmidt &
Ostfeld 2008)や猛禽類の多さ(Schmidt & Ostfeld 2003,Clotfelter et al. 2007)に影響してい ることが示されている。このジェネラリストの捕食者による波は、数の増加だけではなく、
代替食物からのスイッチングといった捕食者の行動的な変化によるものも含まれる
(Schmidt & Ostfeld 2003, 2008)。我が国においてもブナの種子生産の変動がハタネズミ
8
Microtus montebelli やアカネズミの生息密度(箕口1988,1996)、昆虫の個体群動態(Yasaka et al. 2003,Kon et al. 2005)、ツキノワグマ Ursus thibetanus の食性(溝口ら1996,Oka et al.
2004)など、ブナ林に生息するさまざまな生物に影響を与えていることが明らかになってい る。そして、井上(2006)はブナの豊凶とクマタカの繁殖成否に相関があることを示して いる。クマタカが捕食する種の多くはブナの堅果を直接利用する(ネズミ類、ニホンリス、
ヤマドリ)か、ネズミ類の捕食者(アオダイショウ、イタチ、テンなど)であることから
(図 2)、本研究ではブナの豊凶を指標として、クマタカの食物量の変化を繁殖成功率の評 価に盛り込むこととした。さらに、このような年によって変動する環境条件の下では、繁 殖成功率に影響している要因も年によって変化している可能性が考えられる。例えば、ブ ナが豊作のときにはブナ林の面積割合が高いつがいで繁殖成功率が高いが、ブナが種子を つけない年には草地の面積割合が高いつがいの繁殖成功率が高いというようなことがある かもしれない。
図 2 調査地における食物網のイメージ クマタカ
アカネズミ ニホンリス アオダイショウ
イタチ、テン
ブナの種子
ヤマドリ ノウサギ ムササビ カケス
9 2.潜在的分布適地に関する要因
クマタカの潜在的な分布を予測するためのモデルはこれまでにも検討され、地形による 効果が大きいことが示されている。クマタカの分布に関して分析を行なった先行研究は4例 あり、次のような結果が明らかにされている。①鈴木ら(2001)は北海道全域を5㎞メッシ ュに分割し、クマタカの分布情報があるメッシュとないメッシュを比較した。各メッシュ の30°以上の傾斜地面積などの地形に関する指標や広葉樹林面積などのほか、人為改変地 の面積や森林の分断化や森林多様度などの指標を用いてハビタット選好性を調査するとと もに、クマタカが分布する確率から予測マップを作成した。そして、その解析結果から、
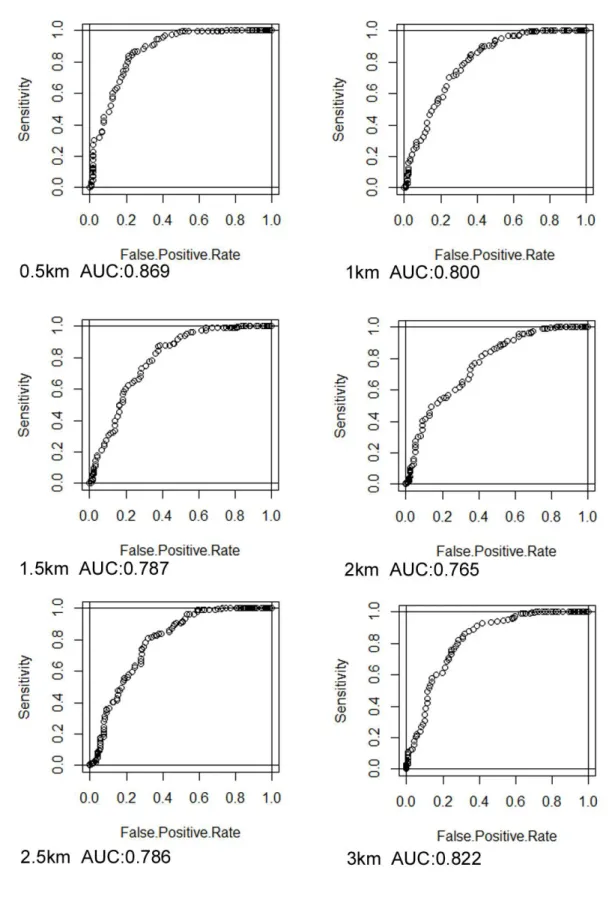
クマタカは生息環境の制限要因として地形に関する要因が影響している一方、森林の面積 などの資源量に関する要因はランドスケープレベルでは影響していないことが示唆された と述べている。また、判別分析によって正答率を求めたところ74%であった。②岩手県に おいては伊藤ら(2004)が岩手県全域を約5㎞のメッシュに分割し、クマタカの生息が確認 されたメッシュと未確認のメッシュの環境変数を比較した。その結果、標高が高く、傾斜 が急で、森林面積率が高い地域にクマタカは生息しているとの結果を得ており、分布推定 モデルの判別率は約80%であった。③杉山ら(2009)は山形県のクマタカの生息適地推定 モデルを作成しており、集計は巣からの距離が半径0.5㎞、1㎞、1.5㎞、3㎞の4通りの距離 ごとに行なっている。また、非生息地のサンプリングに市街地や大規模な農地を含めると、
山地の樹林地にクマタカは生息するという当然の結果になることが予測されるため、樹林 地からサンプリングを行っている。解析には地上開度と地下開度から算出した谷空間指数 や急傾斜地の面積割合などの地形に関するものと、針葉樹林の面積割合や伐開地と草地の 面積割合などの植生に関する指標を用い、すべての変数の組合せでロジスティック回帰分 析を行っている。その結果、最適モデルには、半径1㎞内の谷地形の分布状況と半径3km内 の樹林地の面積割合からなるモデルが選択された。4通りの集計距離を設けたにもかかわら ず半径1㎞内の谷空間指数が選択されたことに関して、この範囲はペア形成、産卵、育雛の ために必要な範囲(繁殖テリトリー)と概ね一致していることから、繁殖テリトリー内の 谷地形が重要ではないかと推測している。また、半径3㎞圏の樹林地面積割合が選択された ことに関して、多様な餌動物の生息が可能な森林が広く分布していることが好ましいのだ ろうと述べている。④伊藤ら(2012)は、鳥取県でクマタカの潜在的生息地と地域の保護 レベル、道路計画を重ね合わせたGap分析によって、攪乱危険地域の抽出を行なった。クマ タカの潜在的生息地図の作成にはロジスティック回帰が使用され、目撃情報の64%が説明 できたとされている。分析では、クマタカの目撃の有無を応答変数とし、説明変数には平 均標高、平均傾斜角、広葉樹・マツ混交林面積占有率、スギ・ヒノキ植林面積占有率、オ ープンエリア面積占有率が用いられた。
これまでの先行研究は、地形的要因がクマタカの分布に強く関係しているという点で一 致している。クマタカは営巣環境、採食環境ともに急傾斜地を利用するとされているが、
景観は水系分布などを含めた地形と土壌、植生、気候および微気象、そして人間と動物の
10
活動が互いに密接に絡み合って形成されている。地形的情報のみからクマタカの分布を予 測しうるということは、開発事業等を行う際には簡便に予測ができるという点で有益だと 思われる。しかし、クマタカやイヌワシ、オオタカなどの希少な猛禽類が生息している国 有林では、生息地としての質をより高めるための森林施業が試みられている地域もある。
そのような取り組みに関しては、植生がクマタカの分布にどのように影響しているのかが 明らかにされることが重要である。
また、降雪がある地域では、植生が雪から受ける影響は傾斜や雪に覆われている期間に よって異なっているが、クマタカの分布には多雪地も少雪地も同じ要因が影響しているの だろうか。これまでクマタカの分布予測を行うために作成されたモデルでは都道府県程度 の規模での分布予測であったが、クマタカの分布に関するより普遍的な生態的特徴を明ら かにするためには、より広域で検討を行う必要があると考える。本研究を行なった東北地 域は中央の脊梁山脈によって、降雪量が大きい日本海側と、降雪量が比較的少なく、冬期 の日照時間も長い太平洋側に大きく二分される。日本海側と太平洋側の植生の違いに関し てはブナとイヌブナFagus japonicaの分布域やチシマザサSasa kurilensisの分布の違いなど、
植生が異なっていることが知られている(梶本ら2002)。そのため、本研究では分布の予測 精度を高めることよりも、広域での生息地を保全し森林を管理してゆく上で、クマタカに とって潜在的に好適な環境はどのようなものかを検討することを目的とした。
11 3.クマタカの生態と本研究で用いた用語について
1)分類
本研究は、クマタカ Nisaetus nipalensis Hodgson, 1836 の日本における亜種 N. n. orientalis を対象に行った。クマタカは、タカ目 ACCIPITRIFORMES、タカ科 ACCIPITRIDAE、クマ タカ属 NISAETUS Hodgson に分類されている(日本鳥学会2012)。本亜種は日本鳥学会
(2000)では Spizaetus nipalensis orientalis (Temminck & Schlegel, 1844)とされていたが、日 本鳥学会(2012)よりNisaetus nipalensis orientalis (Temminck & Schlegel, 1844)とされた。
山階(1986)ではクマタカ属 Spizaetus として、アフリカクマタカ Spizaetus africanus、
カワリクマタカSpizaetus cirrhatus、クマタカSpizaetus nipalensis、ジャワクマタカSpizaetus bartelsi、セレベスクマタカSpizaetus lanceolatus、フィリピンクマタカSpizaetus philippensis、
カオグロクマタカSpizaetus alboniger、ウォーレスクマタカSpizaetus nanus、クロクマタカ Spizaetus tyrannus、アカエリクマタカSpizaetus ornatus の10種が記載されていた。その後、
Ferguson-Lee & Cristie(2001)ではSpizaetusとして、S. andamanensis、S. floris、S. limnaeetus、
S.vanheurniの4種が加わり、14種として記載されていた。近年ではミトコンドリアDNAによ る検討が進められつつあり、Spizaetusを検討したHaring et al.(2007)はクマタカをNisaetus nipalensisとすることを提案した。
2)分布
クマタカはインド亜大陸の南西部とセイロン島、ユーラシア大陸の東南部、インドシナ 半島の北部から中央部と東部、マレー半島北部、台湾島、日本に分布している(森岡ら1998、
日本鳥学会2012)。亜種クマタカ(N. n. orientalis)は日本と朝鮮半島に分布・繁殖し、中国 北東部でも記録があるが、主な分布域は日本と考えられている(環境省2012)。日本国内で の確認記録は千葉県と沖縄県を除いて全国にあり、環境省(2012)では、ある程度自然が 保たれた標高300m以上のほとんどの山地にはクマタカが生息している可能性があると考 えたほうがよいであろうとしている。
3)繁殖
一夫一妻で、留鳥として一年をとおして同じ地域に定着して生息している。求愛期は11 月頃から始まり、本格的な巣作りは1~2月で樹上に巣を造る。一腹卵数は通常1で、3月上 旬~下旬に産卵する。抱卵期間は47日(森岡ら1998)で4月から6月に孵化する場合が多い。
巣立ちは孵化からおよそ70日後(森岡ら1998)で、7~8月に巣立つ(環境省2012)。巣立っ た幼鳥は、巣立ち後も親鳥が繁殖期に入る12~1月頃まで営巣林を中心とした数百mの範囲 から出ず、親から食物をもらっている(森本・飯田1992)。
4)行動域の広さ
行動圏の大きさは、鈴鹿山地でのテレメトリー調査の結果では28.25㎞2(メッシュでの集
12
計)、最外郭で35.75km2(山崎ら1995)、石川県白山地域での目視調査の結果では11.7㎞2(上 馬1989)であった。環境省(2012)では各地の調査結果から行動圏は狭いもので約10㎞2、 広いもので約35㎞2、場合によっては45㎞2を超えるとしている。
5)食性
クマタカは森林に生息するさまざまな小型~中型の哺乳類や鳥類、爬虫類を利用してい る。巣内育雛期に巣へ搬入される食物の構成は、つがいによって大きく異なる。哺乳類で はアカネズミやニホンリス、ムササビ Petaurista leucogenys、ノウサギ、イタチなど、鳥類 ではヤマドリやキジPhasianus colchicus、コジュケイBambusicola thoracicus、ライチョウ Lagopus muta、カケスGarrulus glandarius、ホシガラスNucifraga caryocatactes、トラツグミ Zoothera daumaなど、爬虫類ではヘビ類を捕食しており、特にアオダイショウがよく記録さ れている(井上・山崎 1984,久保上1987,山崎1994,森岡ら 1998,布野ら 2000,柏原・
安田 2004,Kaneda 2009)。
6)使用した用語
本研究で用いた用語の多くはSteenhof & Newton(2007)の定義にならい、日本語の表現 はその邦訳である山崎(2010)を参照した。主な用語は以下のとおりで「 」の記述は山 崎(2010)からの引用である。
営巣地:
本研究では、Steenhof & Newton(2007)および山崎(2010)での「営巣テリトリ ー:ペア形成しているペアの行動圏内で、1つ以上の巣を包含している、または過去 に包含していた場所。確認されている複数の巣を含む限定されたエリアで、通常は 継続的に存在し、1回の繁殖では1ペア以上は存在しない場所である。」とほぼ同義で 用いている。本論の第Ⅲ章では、繁殖期の平均的なホームレンジと同程度の面積と なるように巣を中心とした半径1.5㎞の円形の範囲をデータ集計の範囲とした。クマ タカは年によって巣の位置を変えることがあり、その場合にはホームレンジと想定 する範囲が年によって移動することから、各年のホームレンジと想定される範囲を すべて含む地域を便宜的に営巣地とよんだ。第Ⅳ章では植生ごとの面積などを集計 する範囲を巣から半径0.5㎞から3㎞まで変化させ、同じつがいの行動圏と想定され る地域に複数の巣がある場合には巣ごとに集計し、その平均値を解析に用いた。こ の距離区分はクマタカの行動に基づくものではなく、また、年によって巣の位置も 移動しているため、これらをすべて含む地域を便宜的に営巣地とよんだ。
巣立ち:
「羽毛が生えそろった雛が初めて自発的に巣を離れること。」
本研究では巣が造られている木から雛が離れ、初めて隣接する木へ移動した時点
13 を巣立ちとした。
繁殖成功とみなせる最低限の日齢:
「繁殖成功と考えることができる巣における巣内雛の標準的な日齢。雛は十分に 成長しているが、飛行にはまだ十分ではなく、研究者が安全に巣に入ることができ る時期であり、その後の実際の巣立ちまでの死亡率が最少となる日齢。多くの種に とっては、雛が自らの意志で巣を離れる日齢の80%である。ただし、巣立ち日齢が 相当異なる種または研究者による巣の確認時に時期尚早に巣を離れてしまう傾向の 強い種では、65~75%の日齢とする。」
クマタカの孵化から巣立ちまでの期間は約70日(森岡ら1998)とされるため、本 研究では繁殖成功とみなせる最低限の日齢を、巣立ちに要する日数の80%である約 60日とした。
繁殖成功:
「少なくとも1羽の雛が、繁殖成功とみなせる最低限の日齢に達したこと。」 クマタカの一腹卵数が1であるため、本研究では雛の巣立ちをもって繁殖成功とし た。また、通常の巣立ち日齢の80%に達した雛を確認できた場合も繁殖成功とした。
繁殖成功率:
「あるシーズンに、少なくとも1羽の雛を、繁殖成功とみなせる最低限の日齢にまで 養育したペアの割合。たとえ1回以上の繁殖を試みた場合においても同様。テリトリ ーをもつペア当たり、または産卵したペア当たりについて報告されることが多い。」
本研究ではペアを構成する個体の識別が十分ではなかったため、そのシーズンにペ アが定着していた営巣地数に対する雛が巣立った営巣地の割合とした。また、この ほかに、ある特定の営巣地が一定期間に繁殖成功した割合に対しても‘繁殖成功率’
を用いた。
14
第Ⅲ章 繁殖成功率に影響する環境要因
1.はじめに
ブナ科の樹種などでは、種子生産が大きく年変動する(Kelly 1994)。そのような種子生 産に豊凶がある樹種で構成された森林では、豊作の年にはこの結実量の増加(第1の波)が ドングリを消費するネズミ類の増加(第2の波)を生じさせる。このネズミ類の増加はオコ ジョなどの捕食者の増加(第3の波)を引き起こす(Schmidt & Ostfeld 2008)。この第3の波 には捕食者数の増加だけではなく、ジェネラリストの捕食者が代替食物からネズミ類へス イッチングするといった行動的な変化によるものも含まれる。また、ドングリの消費者は ドングリが落下する前から個体数を増加させ始めていることが明らかにされており、Boutin et al.(2006)はアメリカアカリス Tamiasciurus hudsonicus と キタリス Sciurus vulgaris は 種子が落下する前に豊作を予測でき、繁殖を増加させていることを示した。日本では、種 子生産に大きな年変動がある種としてブナが知られており、5~7年間隔で豊作となるが、そ の間は結実しないか結実量が少ない年が続く(前田 1988,Suzuki et al. 2005)。日本におい てもブナが豊作となった後には、ハタネズミとアカネズミの生息密度が爆発的に上昇する ことが知られており(箕口 1988,1996)、ブナ林に生息するさまざまな種の生息密度はブ ナの結実状況によって影響を受けていると推測される。一般的に、猛禽類の繁殖率の年変 化は食物量の年変動が最も関連することから(Newton 1979)、クマタカにおいてもブナの結 実状況が間接的に繁殖結果に影響していることが予測される。もし、ブナの結実状況がク マタカの繁殖結果に影響しているのであれば、例えば、豊作の翌年には行動圏に占めるブ ナ林の割合が高いつがいは繁殖に成功する確率が相対的に高いが、凶作の翌年には草地な どのブナ林以外の植生の割合が高いつがいで繁殖成功率が高いというように、繁殖結果に 寄与する環境がブナの結実状況に応じて異なっているかもしれない。また、クマタカには 毎年繁殖せずに隔年で繁殖するつがいもいることから(森岡ら1998)、雛を巣立たせた翌年 の雌には生理的な状態が良くないものもいる可能性が考えられ、ある年の繁殖結果にはそ の前年の繁殖結果が関係しているかもしれない。しかし、このような年によって変化する 条件と空間的な生息環境の違いが、クマタカの繁殖に与える影響を複合的に検討した研究 事例は無い。
そこで本研究は、1990年から2008年にかけて山形県最上地方でクマタカの繁殖状況を調 査し、ブナの結実状況とクマタカの繁殖結果との関係を分析した。そして、(1)ブナの結実 状況とクマタカの繁殖結果との関連の有無、(2)関連があった場合、繁殖結果に影響を与え うる環境要因が何であるのかを検討した。
15 2.調査地と方法
1)調査地
調査地は山形県の北東部(39°01′N~38°28′N、140°43′E~139°46′E)に位置 し、南北約70 ㎞、東西約90 ㎞の範囲内の山地である(図 3)。本調査地は日本有数の豪 雪地であるが、夏期の気温は高く、降水量も多い。調査地の気象観測施設の中で山地に近 い肘折(38°36.4′N、 140°9.8′E、標高 330m)では、年降水量 2,808mm、年平均
気温8.8℃、累積降雪量は1,741㎝、最深積雪327㎝である(気象庁 気象統計情報 オン
ライン)。
本地域の自然植生はブナを中心とする落葉広葉樹林が主体をなし、部分的にミズナラ、
スギCryptomeria japonicaによって構成された天然林が分布する。尾根部の土壌が浅い岩場
にはキタゴヨウPinus parviflora var. pentaphylla、クロベThuja standishii、アカマツPinus
densifloraの常緑針葉樹が筋状の林分を形成する。また、雪崩常襲地や崩壊地では、ヒメヤ
シャブシAlnus pendula、タニウツギWeigela hortensisなどの低木やススキMiscanthus sinensis などの高茎草本が生育する。人工林のほとんどはスギの植栽林であり、その他にはアマカ ツが植栽された林がある(環境庁 1988)。
なお、詳細は後述するが、本研究では巣を中心とする半径1.5kmの円形の範囲をホーム レンジと仮定し、空中写真を用いて林相図を作成した。林相図から林相および土地利用区 分ごとの面積を集計した結果、2008年時点の35か所の営巣地でホームレンジに占める森 林率は平均84.3%(SD 9.6)、森林面積に占める人工林の面積割合は平均35.1%(SD 13.4)
であった。作成した林相図と現存植生図(環境省生物多様性センター 自然環境情報 GIS
(植生図)オンライン)を照らし合わせると、天然林の約80%がブナ林であった。本研究 では現存植生図の次の群落をブナ林として扱った。オオシラビソ-ブナ群落、チシマザサ
-ブナ群団、ブナ-チシマザサ群落、ブナ-チシマザサ群集、ヒメアオキ-ブナ群集、ク ロモジ-ブナ群集、マルバマンサク-ブナ群集、スギ-ブナ群落、ブナ-スズタケ群団、
ブナ-スズタケ群集、ヤマボウシ-ブナ群集、ブナ-ツクバネウツギ群集、ブナ-ミヤコ ザサ群集、ブナ-ヤマボウシ群集、ブナ-ツクシシャクナゲ群集、オオマルバノテンニン ソウ-ブナ群落、ブナ-オオモミジガサ群集、シラキ-ブナ群集、ブナ-ミズナラ群落。
16
図 3 調査地の位置とブナ林の分布.
調査地は山形県北部の山地で、北緯39度01分~38度28分、東経140度43分~139度46分の地域であ る。ブナ林の分布は、環境省生物多様性センターの自然環境情報 GIS(植生図)(オンライン)から作成 した。
2)繁殖状況の確認
調査期間は1990年から2008年で、調査方法と繁殖結果の判定等は Steenhof & Newton
(2007)にしたがい、次のように行なった。繁殖状況の調査は、産卵と雛の有無の確認、
雛の日齢および巣立ち時期の推定を目的に、抱卵期から巣内育雛期の間に巣を2回以上訪問 し、望遠鏡と双眼鏡で観察することにより行なった。使用されていない古巣に対しても、
新たに搬入された巣材の有無や巣の状態などを観察した。一般にクマタカの一腹卵数は1で あり(森岡ら 1998)、繁殖結果の判定は雛の巣立ちをもって繁殖成功とした。しかし、雛 の巣立ちを確認することは難しいため、通常の巣立ち日齢の80%に達した雛を確認できた ものは巣立ったとみなした。繁殖成功率は産卵の有無にかかわらず、その年につがいが定 着していた営巣地数に対する雛が巣立った営巣地の割合とした。分析の対象をつがいでは なく営巣地とした理由は、つがいを構成する個体の識別が十分ではなかったためである。
クマタカの行動圏の地形的な主な構成要素は大きな谷であり(森本・飯田1992)、別の谷に 巣を移動させることは多くない(クマタカ生態研究グループ 2000)。そこで、前年の巣の 位置から主要な尾根を越えて1km以上離れた別の谷に営巣した場合は、新たな営巣地として
17
扱った。なお、1か所の営巣地に定着しているつがい数は1である。調査は調査地のほぼ全 域で行ない、調査開始後も新たな営巣地の発見に努めた。調査対象の営巣地数は調査開始 時には6か所であったが、最終的には35か所となった。しかし、調査地内のクマタカの繁殖 分布を完全に把握することはできなかった。
3)ブナの結実状況
ブナの結実状況とクマタカの繁殖結果との関係性は、ある年の繁殖結果に対する1年前と 2年前のブナの結実状況のデータを用いて解析した。これは、ブナの果実は9月から11月に落 下し、クマタカは3月から4月に産卵するため暦上1年のずれが生じるからである。また、被 食者数の変動に対する捕食者の量的な変化はいくつかのパターンがあり(Begon et al. 2006,
Yang et al. 2008)、被食者の増加に対して捕食者数は時間差をもって増加することが知られ
ている(Schmidt & Ostfeld 2003,Clotfelter et al. 2007)。実際に、カシ類の豊作により増加し たネズミ類が、鳥類の個体群に2年間影響した事例(McShea 2000)があることから、2年前 の結実状況も解析に使用した。
ブナの結実状況は、独立行政法人森林総合研究所のブナ結実状況データベース(オンラ イン)によった。ブナ結実状況データベースでは、調査地点ごとにブナの結実状況を豊作、
並作、凶作、無結実に区分し、1989年以降の結実状況が示されている。豊作はほとんどの 木が結実、並作は大径木を中心に約半数の木が結実、凶作はわずかな木にのみ結実、無結 実は全く結実していない、という結実状況を表している。本研究では巣から最も近い地点 の結実状況を参照した。
4)林相・土地利用
林相と土地利用区分ごとの面積は、1987年から2001年の間に撮影された空中写真を用い て作成した林相図から集計した。林相図上で面積を集計した範囲は、ラジオテレメトリー 法による調査事例(クマタカ生態研究グループ 2000)を参考に、繁殖期の平均的なホーム レンジと同程度の面積となるように巣を中心とする半径1.5㎞の円形の範囲とした。クマタ カは年によって巣の位置を変えることがあるため、1か所の営巣地に複数の古巣が存在する ことがある。そのため、年ごとに繁殖に使用された巣を中心として各林相等の面積を集計 した。また、産卵は無かったものの造巣が確認された場合はその巣を、そして、産卵は無 かったが複数の古巣で巣材の運搬が確認された場合は、それらの中で直近の年次に産卵し た巣を中心とした。写真判読の際には、5m四方の大きさを林相および土地利用の最小区分 単位とした。林相は、現地で確認した林内の階層構造の発達状況および林冠と低木層との 間の林内空間の有無などと樹高の関係を参考にし、林分の平均樹高が10m未満の林分を若 齢とし、人工林、若齢人工林、天然林、若齢天然林(二次林)に区分した。景観の特徴を 表す指標は多数提案されているが、Turner et al.(2001)とCushman et al.(2008)による検討 結果を参考に、林分の分布パターンの指標としてclumpiness index を、林縁の複雑さの指標
18
として面積加重平均つきの fractal dimension index(O’Neill et al. 1988)を用いた。林相図の 作成および各林相の面積集計にはArcGIS 9.1(ESRI Inc. 2002)を用い、景観に関する指標は、
Fragstats 3.3(McGarigal et al. 2002)を使用して得た。
5)気象
猛禽類の繁殖と気象要因の関係については、降水量が年生残率(Franklin et al. 2000)、繁 殖成功率(Kostrzewa & Kostrzewa 1990,Kostrzewa & Kostrzewa 1991,Krüger et al. 2002,
McDonald et al. 2004)、被食動物の活動(Olsen & Olsen 1992)に影響していた事例、繁殖成 功率に気温(Elkins 1983,Kostrzewa & Kostrzewa 1991,Ontiveros & Pleguezuelos 2003,Fairhurst
& Bechard 2005)、降雪(Selås 2001)が影響していた事例などがある。そのため、本調査地
で平均的と考えられる営巣活動の進行段階に照らして、抱卵期の日平均気温(4月上旬から
5月中旬)、巣内育雛期の降水量(5月下旬から8月上旬の日降水量の合計)、繁殖期間の降雪
量(11月上旬から4月下旬までの日降雪量の合計)を説明変数として用いた。気象に関する 情報は、最も巣に近い観測所または地域気象観測システム(AMeDAS: Automated
Meteorological Data Acquisition System)の観測情報を気象庁の気象統計情報(オンライン)
から利用した。
6)統計解析
調査年数は営巣地によって 8年から19年と異なっていることから、営巣地間で繁殖成 功率に差がないことを 35 か所の営巣地ごとに調査期間の繁殖成功数、失敗数を用いて
Fisherの正確確率検定により確認した。クマタカはモミAbies firma、ツガTsuga sieboldii、
コメツガTsuga diversifolia、アカマツ、ブナなどに巣を架けることが多い(森岡ら1998)
が、営巣した木が広葉樹か針葉樹かで繁殖成功率に違いがあるのかをFisherの正確確率検 定により確認した。そして、営巣地の密度の高さが繁殖活動に影響している可能性が考え られたため、各営巣地から半径2㎞、3㎞、4㎞、5㎞の範囲にある隣接する営巣地数と各 営巣地の調査期間の繁殖成功率との関係をそれぞれ一元配置分散分析により検証した。ま た、調査地全体での繁殖成功率が年によって異なっているのかを検証するため、調査を行 った1990年から2008年の各年の調査地全域の繁殖成功数と失敗数を用いてFisherの正確 確率検定を行なった。そして、繁殖成功率の経年変化の傾向を明らかにするため、各年の 繁殖成功率を対数変換し、線形回帰した後に逆変換することによりトレンド線を作成した。
本研究では調査地内のすべての営巣地を把握できていないため、調査期間に生息密度が 変化し、密度効果によって繁殖成功率が変化した可能性も考えられる。調査初期に調査地 で確認されていた営巣地は6か所であったが、その後の調査により35か所まで増加した。
2002年以降に新たに確認された営巣地は2か所だけであり、既知の営巣地に隣接する地域 では新たな営巣地がほとんど確認されなかった。そこで、少なくとも2002年以降は確認さ れている営巣地の周辺では生息密度に大きな変化がなかったと推測されることから、2002
19
年から2008年の期間について、営巣地間で繁殖成功率に差がみられるのかをFisherの正確 確率検定によって確認した。また、1990 年時点で確認されていた 6 営巣地と最近発見さ れた7営巣地とで繁殖成功率に差がみられるのか、2002~2008年の7年間の繁殖成功率 を比較した。
クマタカの繁殖結果とブナの結実状況との関係を確認するにあたっては、ある年の繁殖 結果に対し、その1年前の繁殖結果、1年前と2年前のブナの結実状況の三つの要因に注目し、
次の関係についてFisherの正確確率検定で調べた。①前年の繁殖成否と当年の繁殖結果、② 前年のブナの結実状況と当年の繁殖結果、③2年前の結実状況と当年の繁殖結果、④前年の ブナの結実状況と繁殖失敗事例に占める卵や雛が巣立ち前に死亡した事例の割合、⑤前年 のブナの結実状況と当年に産卵があった営巣地の割合。①では、調査期間に全営巣地から 得た各年の繁殖結果を前年の繁殖成否ごとに集計し、前年の繁殖に成功した場合と失敗し た場合の繁殖成功数・失敗数を用いて検定を行った。②では、全営巣地の各年の繁殖結果 を各地の前年の結実状況ごとに集計し、各結実区分の繁殖成功数・失敗数を用いて検定し た。その際に、前年の繁殖に成功した場合と失敗した場合とに分け、それぞれの場合で前 年の結実状況と繁殖結果との関係を調べた。③は②と同様の手順で2年前の結実状況ごとに 集計し、検定を行なった。④では、全営巣地の繁殖失敗事例について前年の繁殖成否ごと、
前年のブナの結実状況ごとに集計し、産卵したが巣立ち前に死亡した事例数と産卵が無か った事例数を用いた。⑤では、全営巣地での産卵の有無を前年の繁殖に成功した場合と失 敗した場合に分けて前年のブナの結実状況ごとに集計し、それぞれの当年に産卵した事例 数と産卵がなかった事例数を用いた。なお、各営巣地の調査初年のデータは前年の繁殖結 果がわからないため、1990年のクマタカの繁殖に関するデータは2年前のブナの結実状況が 不明なためすべての解析から除外した。
次に、繁殖結果に強く影響している要因を明らかにするため、営巣地周辺の土地利用区 分ごとの面積などを含め一般化線形混合モデル(以下、GLMM)を構築した。GLMMは、
同一の調査地点で繰り返し測定していることによる自己相関の回避に有効とされる
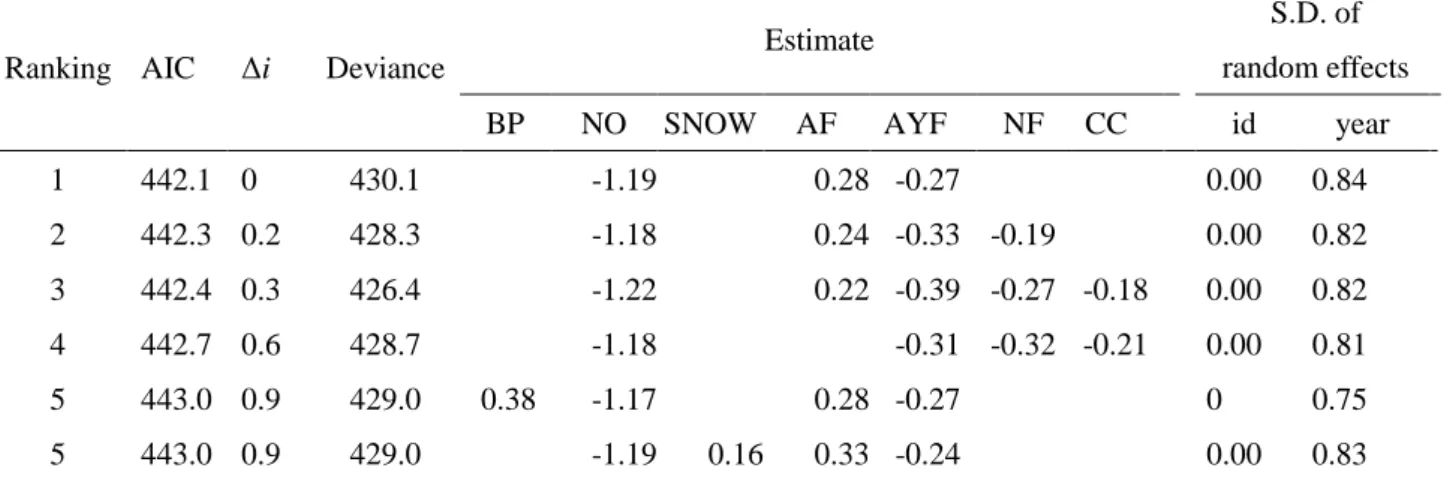
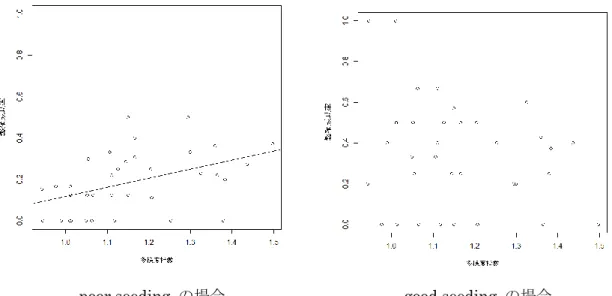
(Crawley 2005,Faraway 2006)。GLMMでは、応答変数を繁殖結果(巣立ち雛の有無)と し、表 2に示した15の変数を説明変数とした。GLMMでブナの結実状況を説明変数とし て用いる際には4つの区分を2区分に統合し、豊作または並作の場合を「good seeding」、 凶作または無結実の場合を「poor seeding」とした。これは、本研究が結実状況をクマタカ の食物条件を示す指標として使用しており、食物条件の良し悪しが繁殖結果に与える影響 の強さを他の説明変数と比較すること、食物条件が良い場合と悪い場合とで繁殖結果に強 く影響する要因が異なるのか比較することを目的としているからである。次に、good
seedingの営巣地のデータセットによるモデルとpoor seedingの営巣地だけのデータセット
によるモデルを GLMM により作成し、それぞれで繁殖結果に影響した要因に違いがみら れるのかを比較した。この比較は、表 2の15変数から前年のブナの結実状況を除いた14 の説明変数でモデルの作成を行なった。なお、食物条件が良い場合を「豊作と並作」の組