【原著・基礎】
Tebipenem pivoxil の小児臨床試験におけるインフルエンザ菌の PCR 法による
耐性遺伝子解析と抗菌薬感受性
岸井こずゑ・千葉菜穂子・諸角美由紀・濱野(長谷川)恵子・生方 公子 北里大学大学院感染制御科学府病原微生物分子疫学研究室*
(平成
20
年9
月26
日受付・平成20
年12
月19
日受理)Tebipenem pivoxil(TBPM-PI)の小児を対象とした臨床第 II
相試験と第III
相試験において分離さ れ,原因菌と判定されたインフルエンザ菌は,急性中耳炎由来が112
株,急性副鼻腔炎由来が16
株,肺 炎由来が30
株,計158
株であった。全株についてβ
―ラクタム系抗菌薬耐性化にかかわるPBP3
をコード するftsI
遺伝子解析を行い,それらを遺伝子レベルで識別した。最も多かったのはgBLNAR
の51.9% で
あり,gLow-BLNARが8.2%, gBLPACR-I
とgBLPACR-II
がそれぞれ0.6% と 1.9% の割合で認められ
た。gBLNAS
は36.7% にすぎなかった。分離されたインフルエンザ菌の 94.9% は,莢膜を保持しない型
別不能株であった。TBPM
のgBLNAR
に対するMIC
90は1.0 µ g! mL
であり,経口抗菌薬のなかではcefditoren
(MIC90:0.25 µ g! mL)に次いで優れていた。他の経口抗菌薬の gBLNAR
に対するMIC
90は,ampicillinとfaro- penem
が8 µ g! mL,amoxicillin
とcefdinir
が32 µ g! mL
であった。ftsI
遺伝子の塩基解析では,次第にアミノ酸の置換数が増え始めていることが示された。また,無作為 抽出したgBLNAR
の全DNA
をSmaI
で切断後にPulsed-field gel electrophoresis
によってその切断パ ターンを比較した結果,著しい多様性が認められた。これらの成績から,TBPM-PIは小児の
gBLNAR
による急性中耳炎やその他の呼吸器感染症に対し て,優れた細菌学的効果を有していると結論された。Key words: tebipenem,Haemophilus influenzae,PBP-3,child,drug susceptibility
Haemophilus influenzae
(インフルエンザ菌)は,Streptococ-cus pneumoniae
(肺炎球菌)と同様に小児の急性中耳炎,急性副鼻腔炎や肺炎の主要な原因菌である。一般的に呼吸器感染 症や急性中耳炎由来のインフルエンザ菌は,その
9
割が莢膜 を保持しない型別不能株(NT株)であると報告されている1)。NT
株の病原性については諸説あるが,ヒトの気道表面が ウィルスなどの感染によりダメージを受けた際,そこにコロ ナイゼーションしているNT
株が病原性を発揮するというの が定説の一つとして挙げられる2)。一方,本邦において臨床上問題となってきた耐性インフル エンザ菌は,長い間
β-lactamase
産生菌のみで,1980年代に は分離菌の25〜30% を占めていた
2)。それらの耐性菌には,一 部のセフェム系抗菌薬が優れた抗菌力を発揮していたが,1998
年以降,β -lactamase
を産生しないインフルエンザ菌(β - lactamase-nonproducing ampicillin-resistant H. influenzae : BLNAR)が経年的に急速に増加し,その傾向は今でも持続し
ている3,4)。そして,臨床では本菌の耐性化による急性中耳炎の難治例 や反復例の増加が特に小児において問題となっている4)。
BLNAR
における耐性化は,β
―ラクタム系抗菌薬の作用標的である隔壁合成酵素―3(penicillin-binding protein 3:PBP
3)をコードする ftsI
遺伝子上に生じた変異に起因するもので,耐性菌では酵素活性上重要ないくつかのアミノ酸が他の アミノ酸へ置換している5)。なかでも,①
Asn
526Lys,② Arg
517His,③ Ser
385Thr
の3
カ所の置換が特に重要であることが明 らかにされている。①あるいは②のどちらかのアミノ酸置換 を有する株は,耐性度がやや低いことからgenotype
のg
を付して
gLow-BLNAR,①と③,あるいは②と③のアミノ酸置
換を有する株は
gBLNAR
として区別されている。gBLNAR の薬剤感受性の特徴は,ペニシリン系抗菌薬に対する感受性 も低下するが,PBP3に親和性の高いセフェム系抗菌薬に対 する感受性も大きく低下することである。本稿においては,TBPM-PIの小児を対象とした臨床第
II
相試験3
試験および臨床第III
相試験3
試験において原因菌 と判定されたインフルエンザ菌について,i)PCR
による耐性 遺伝子解析,ii)TBPM-PI
の活性本体であるTBPM
とその他 の抗菌薬感受性の比較,およびiii)ftsI
遺伝子の塩基解析を 行い,本菌に対するTBPM
の抗菌力について総合的に評価を*東京都港区白金
5―9―1
Ta bl e 1 . Re s i s t a nt t y pe s of Hae mo phi l us i nf l ue nz ae f or e a c h i nf e c t i on
Pne umoni a S i nus i t i s
No. of AOM s t r a i ns Ge not y pe
7 ( 2 3 . 3 ) 5 ( 3 1 . 3 )
4 6 ( 4 1 . 1 ) 5 8 ( 3 6 . 7 )
g BLNAS
1 ( 3 . 3 ) 0
0 1 ( 0 . 6 ) g BLPAR
4 ( 1 3 . 3 ) 0
9 ( 8 . 0 ) 1 3 ( 8 . 2 )
g Low- BLNAR
1 7 ( 5 6 . 7 ) 1 0 ( 6 2 . 5 )
5 5 ( 4 9 . 1 ) 8 2 ( 5 1 . 9 )
g BLNAR
0 1 ( 6 . 3 ) 0
1 ( 0 . 6 ) g BLPACR- I
1 ( 3 . 3 ) 0
2 ( 1 . 8 ) 3 ( 1 . 9 )
g BLPACR- I I
3 0 1 6
1 1 2 1 5 8
Tot a l
The numbe r i n ( ) i ndi c a t e s t he % i n e a c h di s e a s e
行ったので報告する。I. 材 料 と 方 法 1.対象菌株
0
歳以上15
歳以下の小児を対象として,2005年10
月〜2008年3
月 の 間 に 延 べ150
施 設 で 実 施 さ れ たTBPM-PI
の臨床第II
相試験3
試験および臨床第III
相 試験3
試験において分離され,原因菌であると判定され たインフルエンザ菌,合計158
株を対象とした。対象疾 患の内訳は急性中耳炎が112
株,急性副鼻腔炎が16
株,肺炎が
30
株であった。2.PCR
による耐性遺伝子解析PCR
による菌種同定と耐性遺伝子検索は,すでに報告 した方法に準じて行った6)。インフルエンザ菌の解析には,①インフルエンザ菌に 特異的な
P6
膜蛋白をコードするp6
遺伝子の検索用7),②
TEM
型β -lactamase
をコードするbla
TEM-1遺伝子の検 索用8),③ROB
型β -lactamase
をコードするbla
ROB-1遺伝 子 の 検 索 用9),④PBP3
を コ ー ド す るftsI
遺 伝 子 上 のAsn
526のLys
へのアミノ酸置換の検索用(PBP3-S)10),⑤Asn
526Lys
とSer
385Thr
の2
カ所のアミノ酸置換の検索用(PBP3-BLN)5),そして⑥
type b
インフルエンザ菌識別 用11)の計6
組のプライマーを使用した。これらの遺伝子解析により,それぞれの耐性にかかわ る遺伝子を保持する場合は,genotypeの
g
を付した。すなわち,遺伝子学的変異の見いだされなかった株は
gBLNAS,②あるいは③を保持する β -lactamase
産生株 はgBLPAR,④の変異を有する β -lactamase
非産生am- picillin
(ABPC)軽度耐性株はgLow-BLNAR,⑤の変異
を 保 持 す るβ -lactamase
非 産 生ABPC
耐 性 株 はgBLNAR,gLow-BLNAR
株 のβ -lactamase
産 生 株 はgBLPACR-I,gBLNAR
株のそれはgBLPACR-II
と表記 した。3.薬剤感受性測定
薬 剤 感 受 性 の 測 定 は,Muller-Hinton agar(Becton
Dickinson and Company, Sparks, MD)を基礎培地とし,
馬血液(最終濃度:2%)および
β -NAD(最終濃度:15 µ g! mL)を加えた寒天平板希釈法によって実施した
12)。 微量液体希釈法を用いなかった理由は,broth法では発 育の劣る菌がしばしば認められることによる。感受性測定の対象薬剤は,
TBPM, penicillin G
(PCG),ABPC,amoxicillin
(AMPC),cefdinir(CFDN),cefdi-toren(CDTR),faropenem(FRPM),clarithromycin
(CAM),azithromycin(AZM)の経口薬
9
薬剤と,ce-fotaxime(CTX), panipenem(PAPM),meropenem
(MEPM)の注射薬
3
薬剤,計12
薬剤とした。それぞれの薬剤は,当該企業から力価の明らかな原末 の提供を受けた。MIC測定時には標準株の
H. influenzae ATCC49247
およびH. influenzae ATCC 49766
をコント ロール株として用いた。4.ftsI
遺伝子の塩基解析PCR
による耐性遺伝子解析の結果,gLow-BLNAR,gBLNAR,gBLPACR-I,gBLPACR-II
と判定された99
株を対象に,ftsI遺伝子の塩基解析を行った。先ず,ftsI 遺伝子の約1.0 kbp
をsense-primer 5ʼ-GTTGCACATATCTCCGATGAG-3ʼと reverse-primer 5ʼ-CAGCTGCTTCAGCATCTTGC-3ʼを
用いて増幅した。得られたPCR
産物はQIA quick PCR purification kit
(Qiagen,Tokyo,Japan)にて精製後,BigDye Terminator cycle sequencing kit version 3.1
(Applied Biosystems,Foster City,CA)を用いてシー クエンス反応を行った。
DNA
シークエンスにはApplied Biosystems 3130! 3130xl Genetic Analyzer
(Applied Bio-systems)を使用した。
5.PFGE
解析インフルエンザ菌に対する
PFGE
解析は,すでに報告 された方法に若干の変更を加えて実施した13)。被験菌を混合した寒天ブロックを作成後,
30 U
の制限 酵素SmaI
にて30℃,16
時間の酵素反応を行い,DNA を切断した。電気泳動はCHEF MAPPER
(Bio-Rad Labo-ratories,Hercules,CA,USA)を使用し,14℃,6 V!
cm
の条件で20
時間行った。II. 結
果1.PCR
による耐性遺伝子型急性中耳炎由来
112
株,急性副鼻腔炎由来16
株,肺炎 由来30
株, 計158
株の耐性遺伝子型はTable 1
に示す。急性中耳炎由来株では
gBLNAR
が55
株(49.1%),gLow-BLNAR
が9
株 (8.0% ), gBLPACR-II
が2
株(1.8%)であり,合計すると耐性菌の占める割合は
58.9%
となった。急性副鼻腔炎由来株では
gBLNAR
が10
株(62.5%),gBLPACR-Iが
1
株(6.3%)で,耐性菌の占め る割合は68.8% であった。
一方,肺炎由来株で最も多かったのは
gBLNAR
の17
株(56.7%),次 い でgLow-BLNAR
の4
株(13.3%),gBLPACR-II
の1
株(3.3%),gBLPARの1
株(3.3%)で あり,耐性菌が全体の76.6% を占めた。
疾患により耐性菌の占める割合にはばらつきが認めら
Fi g . 1 . Re l a t i on be t we e n pa t i e nt s ’ a g e a nd t he r e s i s t a nc e g e ne t y pe .
gBLNAS gBLPAR
gLow-BLNAR gBLNAR gBLPACR-I gBLPACR-II 35
30 25 20 15 10 5 0
20
15
10
5
0
gBLNAS gBLPAR
gLow-BLNAR gBLNAR gBLPACR-I gBLPACR-II (A) AOM+Sinusitis
(B) Pneumonia (n)
(n)
AOM: acute otitis media
>10y
<1y 1y 2y 3y 4y 5y 6y 7y 8y 9y 10y
>10y
<1y 1y 2y 3y 4y 5y 6y 7y 8y 9y 10y
れたが,いずれの疾患においてもその割合は
50% 以上と
きわめて高頻度であった。なお,これらの原因菌のうち
150
株(94.9%)は莢膜を 保持しない型別不能株であり,残りの7
株が莢膜b
型,1
株が莢膜f
型であった。2.症例の年齢と耐性遺伝子型との関係
Fig. 1(A)には急性中耳炎例と急性副鼻腔炎例,Fig.
1(B)には肺炎例の年齢分布と原因となったインフルエ
ンザ菌の耐性遺伝子型との関係を示す。急性中耳炎例と急性副鼻腔炎例における発症年齢を
1
歳以下,2〜5歳,および6
歳以上に区別して耐性菌の割 合をみると,1歳以下で耐性菌による発症例が有意に多 かった(χ
2=14.5930,P=0.0007(**))。肺炎例では症例 数が少なく統計学的解析はできなかった。3.耐性遺伝子型と薬剤感受性
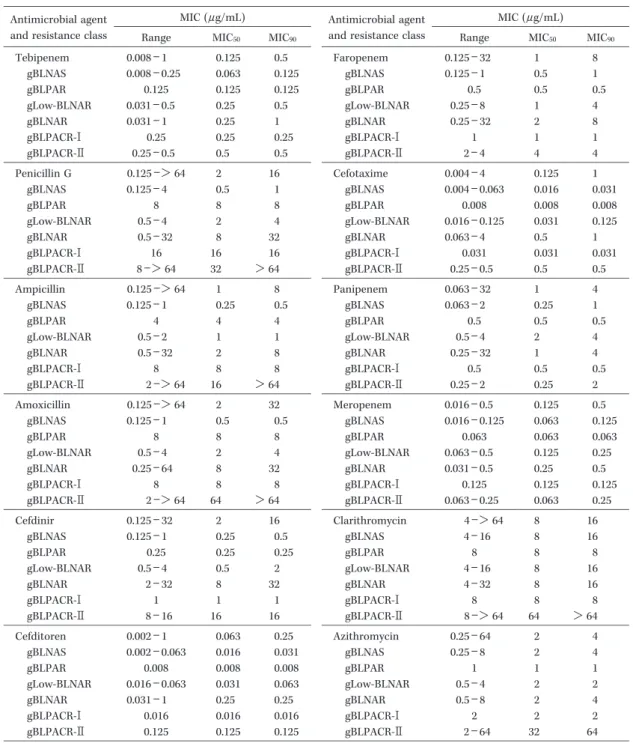
耐性型ごとに分類されたインフルエンザ菌に対する
TBPM
と 経 口 抗 菌 薬9
薬 剤,お よ び 注 射 薬3
薬 剤 のMIC
50,MIC
90, ならびにMIC range
をTable 2
に示す。また,Fig. 2には
TBPM,Fig. 3
にはその他のβ
―ラク タム系抗菌薬のMIC
分布に耐性遺伝子の成績を重ね合 わせて示した。菌株全体の耐性型の内訳は,gBLNAS が58
株(36.7%),gBLPAR
が1
株(0.6%),gLow-BLNAR
が13
株(8.2%),gBLNAR
が82
株(51.9%),gBLPACR-I
が1
株(0.6%),そしてgBLPACR-II
が3
株(1.9%)で あった(Table 1)。ftsI
遺伝子変異の影響を強く受けるセフェム系抗菌薬 の,感受性分布は2
峰性を呈し,gBLNARは耐性側に1
つのピークを形成しており,gBLNAS
とは比較的明瞭に 区別されていた。ペニシリン系抗菌薬やカルバペネム系Ta bl e 2 . MI C di s t r i but i ons a nd r e s i s t a nc e g e ne s i de nt i f i e d by PCR i n Hae mo phi l us i nf l ue nz ae MI C ( μ g / mL) Ant i mi c r obi a l a g e nt
a nd r e s i s t a nc e c l a s s MI C ( μ g / mL)
Ant i mi c r obi a l a g e nt
a nd r e s i s t a nc e c l a s s Ra ng e MI C
50MI C
90Ra ng e MI C
50MI C
908 1
0 . 1 2 5 ― 3 2 Fa r ope ne m
0 . 5 0 . 1 2 5 0 . 0 0 8 ― 1
Te bi pe ne m
1 0 . 5
0 . 1 2 5 ― 1 g BLNAS
0 . 1 2 5 0 . 0 6 3
0 . 0 0 8 ― 0 . 2 5 g BLNAS
0 . 5 0 . 5
0 . 5 g BLPAR
0 . 1 2 5 0 . 1 2 5
0 . 1 2 5 g BLPAR
4 1
0 . 2 5 ― 8 g Low- BLNAR
0 . 5 0 . 2 5
0 . 0 3 1 ― 0 . 5 g Low- BLNAR
8 2
0 . 2 5 ― 3 2 g BLNAR
1 0 . 2 5 0 . 0 3 1 ― 1
g BLNAR
1 1
1 g BLPACR- I
0 . 2 5 0 . 2 5
0 . 2 5 g BLPACR- I
4 4
2 ― 4 g BLPACR- I I
0 . 5 0 . 5
0 . 2 5 ― 0 . 5 g BLPACR- I I
1 0 . 1 2 5 0 . 0 0 4 ― 4
Ce f ot a x i me 1 6
2 0 . 1 2 5 ―> 6 4 Pe ni c i l l i n G
0 . 0 3 1 0 . 0 1 6
0 . 0 0 4 ― 0 . 0 6 3 g BLNAS
1 0 . 5
0 . 1 2 5 ― 4 g BLNAS
0 . 0 0 8 0 . 0 0 8
0 . 0 0 8 g BLPAR
8 8
8 g BLPAR
0 . 1 2 5 0 . 0 3 1
0 . 0 1 6 ― 0 . 1 2 5 g Low- BLNAR
4 2
0 . 5 ― 4 g Low- BLNAR
1 0 . 5
0 . 0 6 3 ― 4 g BLNAR
3 2 8
0 . 5 ― 3 2 g BLNAR
0 . 0 3 1 0 . 0 3 1
0 . 0 3 1 g BLPACR- I
1 6 1 6
1 6 g BLPACR- I
0 . 5 0 . 5
0 . 2 5 ― 0 . 5 g BLPACR- I I
> 6 4 3 2
8 ―> 6 4 g BLPACR- I I
4 1
0 . 0 6 3 ― 3 2 Pa ni pe ne m
8 1
0 . 1 2 5 ―> 6 4 Ampi c i l l i n
1 0 . 2 5 0 . 0 6 3 ― 2
g BLNAS 0 . 5
0 . 2 5 0 . 1 2 5 ― 1
g BLNAS
0 . 5 0 . 5
0 . 5 g BLPAR
4 4
4 g BLPAR
4 2
0 . 5 ― 4 g Low- BLNAR
1 1
0 . 5 ― 2 g Low- BLNAR
4 1
0 . 2 5 ― 3 2 g BLNAR
8 2
0 . 5 ― 3 2 g BLNAR
0 . 5 0 . 5
0 . 5 g BLPACR- I
8 8
8 g BLPACR- I
2 0 . 2 5 0 . 2 5 ― 2
g BLPACR- I I
> 6 4 1 6
2 ―> 6 4 g BLPACR- I I
0 . 5 0 . 1 2 5 0 . 0 1 6 ― 0 . 5
Me r ope ne m 3 2
2 0 . 1 2 5 ―> 6 4 Amox i c i l l i n
0 . 1 2 5 0 . 0 6 3
0 . 0 1 6 ― 0 . 1 2 5 g BLNAS
0 . 5 0 . 5
0 . 1 2 5 ― 1 g BLNAS
0 . 0 6 3 0 . 0 6 3
0 . 0 6 3 g BLPAR
8 8
8 g BLPAR
0 . 2 5 0 . 1 2 5
0 . 0 6 3 ― 0 . 5 g Low- BLNAR
4 2
0 . 5 ― 4 g Low- BLNAR
0 . 5 0 . 2 5
0 . 0 3 1 ― 0 . 5 g BLNAR
3 2 8
0 . 2 5 ― 6 4 g BLNAR
0 . 1 2 5 0 . 1 2 5
0 . 1 2 5 g BLPACR- I
8 8
8 g BLPACR- I
0 . 2 5 0 . 0 6 3
0 . 0 6 3 ― 0 . 2 5 g BLPACR- I I
> 6 4 6 4
2 ―> 6 4 g BLPACR- I I
1 6 8
4 ―> 6 4 Cl a r i t hr omy c i n
1 6 2
0 . 1 2 5 ― 3 2 Ce f di ni r
1 6 8
4 ― 1 6 g BLNAS
0 . 5 0 . 2 5
0 . 1 2 5 ― 1 g BLNAS
8 8
8 g BLPAR
0 . 2 5 0 . 2 5
0 . 2 5 g BLPAR
1 6 8
4 ― 1 6 g Low- BLNAR
2 0 . 5
0 . 5 ― 4 g Low- BLNAR
1 6 8
4 ― 3 2 g BLNAR
3 2 8
2 ― 3 2 g BLNAR
8 8
8 g BLPACR- I
1 1
1 g BLPACR- I
> 6 4 6 4
8 ―> 6 4 g BLPACR- I I
1 6 1 6
8 ― 1 6 g BLPACR- I I
4 2
0 . 2 5 ― 6 4 Az i t hr omy c i n
0 . 2 5 0 . 0 6 3
0 . 0 0 2 ― 1 Ce f di t or e n
4 2
0 . 2 5 ― 8 g BLNAS
0 . 0 3 1 0 . 0 1 6
0 . 0 0 2 ― 0 . 0 6 3 g BLNAS
1 1
1 g BLPAR
0 . 0 0 8 0 . 0 0 8
0 . 0 0 8 g BLPAR
2 2
0 . 5 ― 4 g Low- BLNAR
0 . 0 6 3 0 . 0 3 1
0 . 0 1 6 ― 0 . 0 6 3 g Low- BLNAR
4 2
0 . 5 ― 8 g BLNAR
0 . 2 5 0 . 2 5
0 . 0 3 1 ― 1 g BLNAR
2 2
2 g BLPACR- I
0 . 0 1 6 0 . 0 1 6
0 . 0 1 6 g BLPACR- I
6 4 3 2
2 ― 6 4 g BLPACR- I I
0 . 1 2 5 0 . 1 2 5
0 . 1 2 5 g BLPACR- I I
抗菌薬においては,gBLNARの
MIC
は耐性側に分布し たものの,感性株と耐性株とを明確に区別することは難 しかった。gBLNAR
に対するTBPM
のMIC
は,MEPMと同様に,
gBLNAS
に比してやや高いものの明確には区別できず,0.031〜1
µ g ! mL
の狭い濃度域に分布した。gBLNAR
に対するMIC
90が優れていた抗菌薬を順に 示すと,CDTR(0.25µ g! mL)>MEPM
(0.5µ g! mL)>
TBPM
(1µ g! mL)=CTX
(1µ g! mL)>PAPM
(4µ g!
mL)=AZM
(4µ g ! mL)>ABPC
(8µ g ! mL)=FRPM
(8
µ g! mL)>CAM
(16µ g! mL)>PCG
(32µ g! mL)=
AMPC(32 µ g! mL)=CFDN(32 µ g! mL)であった。
4. β
―ラクタム系抗菌薬に対する感受性とftsI
遺伝子 上のアミノ酸置換との関係Table 3
に は,gLow-BLNAR(13株),gBLNAR(82 株),gBLPACR-I(1株),およびgBLPACR-II
(3株)の 計99
株におけるftsI
遺伝子上のアミノ酸置換および主 な薬剤のMIC
90値を示す。これらのグループ分類は,
Hasegawa
ら3)によって報告 された8
つのアミノ酸置換,すなわちVal-
329,Met-377,Ser-
385,Leu-389,Ala-502,Val-511,Arg-517,Asn-526の結果に 基づいている。なかでも3
カ所のアミノ酸置換,すなわ ち,①Arg
517His,② Asn
526Lys,③ Ser
385Thr
の置換を中 心に分類している。Fi g . 2 . Di s t r i but i on of Te bi pe ne m MI C of Hae mo phi l us i nf l ue nz ae ( n = 1 5 8 ) . (%)
( μ g/mL)
0.002 0.004 0.008 0.016 0.031 0.063 0.125 0.25 0.5 1 2 4
gBLNAS gBLPAR
gLow-BLNAR gBLNAR gBLPACR-I gBLPACR-II
MIC 50
40
30
20
10
0
gLow-BLNAR
株の13
株(13.1%)は,初期に比較的多 くみられたAsn
526Lys
を有する株であった。一方,gBLNAR株および
gBLPACR-II
では,保存性 ア ミ ノ 酸 配 列 のSer-Ser-Asn(SSN)周 囲 に 位 置 す る Met
377Ile, Ser
385Thr,および Leu
389Phe
のアミノ酸置換を 有する株が77
株と多く認められ,それらのうち71
株はAsn
526Lysを同時に有する株,残りの6株はArg
517Hisを
有する株であった。それ以外の株は最も重要なSer
385Thr
を 有 し て い た が,Met377Ile
お よ びLeu
389Phe
を 併 せ て 有 し て い な か っ た。Ser385Thr
とArg
517His,あ る い は Ser
385Thr
とAsn
526Lys
を有する耐性菌において,ABPC のMIC
90は16〜64
倍 低 下 し,セ フ ェ ム 系 抗 菌 薬 で は32〜128
倍低下していた。しかし,gBLNARに対するTBPM
のMIC
90の変動をみると,gBLNAS
に比較して8
倍程度の感受性低下におさまっていた。注目される変異株として,
β
―ラクタム系抗菌薬の結合 サイトであるSer
を含むSer-The-Val-Lys
(STVK)中にVal
329Ala
置換を有する株が2
株認められた。5.gBLNAR
のPFGE
解析Fig. 4
には無作為に抽出された中耳貯留液由来インフルエン ザ 菌 の
PFGE
の 成 績 を 示 す。制 限 酵 素 と し てSmaI
を用いた。DNA切断パターンには多様性が認められ,
gBLNAR
はさまざまなクローン由来株であることが示された。
III. 考
察著者らは,わが国において肺炎球菌の
PRSP
が問題と なり始めた頃から程なく,同じ呼吸器感染症の原因菌の 一つであるインフルエンザ菌においても,ABPCやce- faclor
(CCL)の感受性が低下した株が出現し始めたこと を報告した1)。それらの株についてβ
―ラクタム系抗菌薬 に対する感受性を詳細に比較すると,ABPCよりもセ フェム系抗菌薬の感受性低下が著明であった14)。この現象は,インフルエンザ菌に対してセフェム系抗菌薬が抗 菌力を発揮するうえで最も重要な,隔壁合成酵素の
PBP 3
に対する薬剤の結合親和性が変化していることを示唆 するものであった。この予測に基づいてさまざまな耐性 レベルのインフルエンザ菌を解析した結果,PBP3を コードするftsI
遺伝子上に共通して認められる変異を見 いだした。しかもそれらは,保存性アミノ酸配列の近く に位置し,かつ中性アミノ酸から塩基性アミノ酸へと置 換していた5)。その後,それらの変異がインフルエンザ菌 のβ
―ラクタム系抗菌薬耐性化にかかわっていることを 形質転換によっても証明している3)。PBP3とβ
―ラクタ ム系抗菌薬の結合を考えると,耐性菌ではPBP
の活性部 位における立体構造に著しい歪みが生じ,結果的にPBP
への薬剤の結合親和性が低下しているものと推定され る。一方,1998年以降の耐性化の状況をみると,BLNAR の増加傾向は持続し,2007年には遂に分離菌の
40% を
上回った。そして,臨床的には耐性インフルエンザ菌に よる肺炎,急性中耳炎,急性副鼻腔炎の難治例や反復例 の増加,さらにはHib
における耐性化が治療上最も解決 されなければならない緊急問題となっている。このようなわが国特有の急激な耐性化が進行した背景 には,耐性インフルエンザ菌に対して経口ペニシリン系 抗菌薬の抗菌力が本質的にそれほど優れておらず,しか も経口セフェム系抗菌薬においても
CDTR
以外は感受 性が非常に低下していたことにある。加えて,インフル エンザ菌は常在細菌の一面も有し,免疫学的に未熟な乳 幼児は本菌を保菌しやすく,そのことも耐性菌による感 染例を増加させる一因になったと考えられる。本来,インフルエンザ菌に対しては,ニューキノロン 系抗菌薬がきわめて優れた抗菌力と殺菌力を保持してい るが,本薬の小児感染症への適応は承認されていない。
Fi g . 3 . Re l a t i on be t we e n PBP3 g e ne mut a t i on a nd β - l a c t a m MI C i n Hae mo phi l us i nf l ue nz ae . Ampicillin
(%)
Amoxicillin
Cefdinir Cefditoren
Faropenem Cefotaxime
Meropenem Panipenem
50 40 30 20 10 0
MIC
gBLNAS gBLPAR gLow-BLNAR gBLNAR gBLPACR-I gBLPACR-II
1 2 4 8 16 32 64
> 64
0.0020.0040.0080.0160.0310.0630.125 0.25 0.5 ( μ g/mL)
1 2 4 8 16 32 64
> 64
0.0020.0040.0080.0160.0310.0630.125 0.25 0.5 ( μ g/mL)
1 2 4 8 16 32 64
> 64
0.0020.0040.0080.0160.0310.0630.125 0.25 0.5 ( μ g/mL)
1 2 4 8 16 32 64
> 64
0.0020.0040.0080.0160.0310.0630.125 0.25 0.5 ( μ g/mL)
1 2 4 8 16 32 64
> 64
0.0020.0040.0080.0160.0310.0630.125 0.25 0.5 ( μ g/mL)
1 2 4 8 16 32 64
> 64
0.0020.0040.0080.0160.0310.0630.125 0.25 0.5 ( μ g/mL)
1 2 4 8 16 32 64
> 64
0.0020.0040.0080.0160.0310.0630.125 0.25 0.5 ( μ g/mL)
1 2 4 8 16 32 64
> 64
0.0020.0040.0080.0160.0310.0630.125 0.25 0.5 ( μ g/mL)
(%) 50 40 30 20 10 0
MIC 50 (%)
40 30 20 10 0
MIC
50 (%)
40 30 20 10 0
MIC 50 (%)
40 30 20 10 0
MIC
50 (%)
40 30 20 10 0
MIC 50 (%)
40 30 20 10 0
MIC
50 (%)
40 30 20 10 0
MIC
このような現状から,小児のインフルエンザ菌感染症に 対して確実に有効性が期待できる
β
―ラクタム系抗菌薬 の開発が望まれていた。本編で述べたインフルエンザ菌に対する
TBPM
の抗 菌活性は,CDTR
ほどではないものの比較的優れている ことが示された。特に,経口吸収性に優れているTBPM-
T ab le 3 . A n al ys is o f th e ft sI g en e en co d in g P B P 3 o f re si st an t H a emo ph il u s in flu en za e i d en ti fi ed a s ca u sa ti ve p at h o ge n MI C
90( μ g/ mL ) A mi n o a ci d s u b st it u ti o n N o o f st ra in s R es is ta n t ty p es K T G mo ti f
cSS N mo ti f
bST V K
aME P M C T X F R P M C D T R C F D N A MP C A B P C T B P M A sn -5 26 A rg -5 17 V al -5 11 A la -5 02 L eu -3 89 Se r- 38 5 Me t- 37 7 V al -3 29 0 .0 63 0 .0 08 0 .5 0 .0 08 0 .2 5 0 .5 0 .2 5 0 .1 25 - - - - - - - - gB L N A S
d0. 25 0 .1 25 4 0 .0 63 2 4 1 0. 5 L ys - - - - - - - 7 gL o w - B L N A R (n = 13 )
L ys - - (T h r) - - - - 2 L ys - - (V al ) - - - - 3 L ys - - (V al ) - - (I le ) - 1 ― ― ― ― ― ― ― ― L ys - - T h r - - Il e - 1 gB L P A C R - I 0. 25 1 4 0. 25 32 8 4 0. 25 - H is - - - T h r - - 1 gB L N A R (n = 82 ) - H is - - - T h r (I le ) - 1 - H is - - (P h e) T h r (I le ) - 6 0. 5 1 8 0. 25 32 32 8 1
L ys - - - - T h r - - 1 L ys - - - - T h r - (A la ) 2 L ys - - - - T h r (I le ) - 1 L ys - - (T h r) - T h r - - 1 0. 5 1 8 0. 5 32 32 16 0. 5 L ys - - - P h e T h r Il e - 66 L ys - (A la ) - P h e T h r Il e - 1 L ys - - (V al ) P h e T h r Il e - 2 ― ― ― ― ― ― ― ― - H is - - - T h r Il e - 1 gB L P A C R - I I ― ― ― ― ― ― ― ― L ys - - - P h e T h r Il e - 2
aST V K : S er
327-T h r- V al -L ys
bSS N : S er
379-S er -A sn
cK T G : L ys
512-T h r- G ly
dC o n tr o l: A T C C 49 76 6
Fi g . 4 . PFGE a na l y s i s of r e s i s t a nt Hae mo phi l us i nf l ue nz ae ( g BLNAR) i s ol a t e d f r om t he mi ddl e e a r f l ui d.
M: CHEF DNA size standard, lambda ladder 8.3―48.5 kb mixed digest of lambda
gBLNAR
M M
PI
は,乳幼児に対する4 mg! kg
の投与量で,3.46 µ g! mL
のCmax
が得られ,臨床においてgBLNAR
を十分にカ バーできるものと考えている。CDTR-PI
との小児比較試 験 に お い て,gBLNARに よ る 急 性 中 耳 炎 に 対 す るTBPM-PI
の細菌学的効果が,CDTR-PIと同等であった ことが,そのことを裏付けているものと推察された。今 後は,TBPM
と他薬剤について,gBLNAR
に対する殺菌 性,postantibiotic effect(PAE),あるいは形態変化を比 較し,TBPMの効果をより明確にする必要があろう。耐性インフルエンザ菌で懸念されることは,PFGE解 析の結果の項で述べたように,
gBLNAR
の特徴はクロー ンの異なる菌のPBP3
遺伝子に変異が生じていることで ある。つまり,常在細菌化しやすく,またbiofilm
を形成 しやすいインフルエンザ菌は,常にさまざまな抗菌薬の 選択圧を受けており,遺伝子変異を有する菌が容易に選 択されやすいともいえる。TBPM
耐性インフルエンザ菌を生じさせないために は,本薬の臨床への導入後において,不十分な投与量や エビデンスのない長期投与は厳につつしまねばならな い。そして,それらの厳密なコントロ―ルが望まれる。文 献
1) 生方公子,千葉菜穂子,小林玲子,長谷川恵子,紺野 昌俊:本邦において
1998
年から2000
年の間に分離 されたHaemophilus influenzae
の分子疫学解析―肺炎 球菌等による市中感染症研究会収集株のまとめ―。日 化療会誌2002; 50: 794-804
2) 生方公子:第
19
回日本臨床微生物学会総会,アナラ イザー・ワークショップ,2008; 513)
Hasegawa K, Kobayashi R, Nakayama E, Takada E, Ono A, Chiba N, et al: High prevalence of type b β - lactamase-nonproducing ampicillin-resistant Haemo- philus influenzae in meningitis: the situation in Japan where Hib vaccine has not been introduced. J An- timicrob Chemother 2006; 57: 1077-82
4) 生方公子:呼吸器感染症原因微生物の質的変化によ る薬剤耐性化。日化療会誌
2006; 54: 69-94
5)
Ubukata K, Shibasaki Y, Yamamoto K, Chiba N, Hasegawa K, Takeuchi Y, et al: Association of amino acid substitutions in penicillin-binding protein 3 with β -lactam resistance in β -lactamase-negative ampicillin-resistant Haemophilus influenzae. Antimi- crob Agents Chemother 2001; 45: 1693-9
6)
Hasegawa K, Yamamoto K, Chiba N, Kobayashi R, Nagai K, Jacobs M R, et al: Diversity of ampicillin- resistance genes in Haemophilus influenzae in Japan and the United States. Microb Drug Resist 2003; 9:
39-46
7)
Nelson M B, Apicella M A, Murphy T F, Vankeulen H, Spotila L D, Rekosh D: Cloning and sequencing of Haemophilus influenzae outer membrane protein P 6.
Infect Immun 1988; 56: 128-34
8)
Sutcliffe J G: Nucleotide sequence of the ampicillin resistance gene of Escherichia coli plasmid pBR 322.
Proc Natl Acad Sci U S A 1978; 75: 3737-41
9)
Juteau J M, Levesque R C : Sequence analysis and evolutionary perspectives of ROB-1 beta-lactamase.
Antimicrob Agents Chemother 1990; 34: 1354-9
10)Fleischmann R D, Adams M D, White O, Clayton R
A, Kirkness E F, Kerlavage A R, et al: Whole-genome random sequencing and assembly of Haemophilus in- fluenzae Rd. Science 1995; 269: 496-512
11)
Van Eldere J, Brophy L, Loynds B, Celis P, Hancock I, Carman S, et al: Region II of the Haemophilus influ- enzae type b capsulation locus is involved in serotype-specific polysaccharide synthesis. Mol Mi- crobiol 1995; 15: 107-18
12)