九州大学学術情報リポジトリ
Kyushu University Institutional Repository
酸味受容体候補分子の電気生理学的解析
堀尾, 奈央
九州大学大学院歯学府
https://doi.org/10.15017/21993
出版情報:Kyushu University, 2011, 博士(歯学), 課程博士 バージョン:
権利関係:
酸味受容体候補分子の電気生理学的解析
堀尾 奈央
九州大学大学院歯学府歯学専攻 口腔常態制御学講座口腔機能解析学分野
指導:二ノ宮 裕三 教授
九州大学大学院歯学研究院
口腔常態制御学講座口腔機能解析学分野
目次
発表論文・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・1 要旨・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・5 緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・7 材料と方法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・10 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・20 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・50
謝辞・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・55 参考文献・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・56
発表論文
対象論文
Nao Horio, Ryusuke Yoshida, Keiko Yasumatsu, Yuchio Yanagawa, Yoshiro Ishimaru, Hiroaki Matsunami, Yuzo Ninomiya
Sour taste responses in mice lacking PKD channels PLoS ONE, 6: (5) e20007, 2011
本研究の一部は下記の学会において発表した。
味覚健康科学シンポジウム、大分、2010年3月
The 32th Annual Meeting of the Association for Chemoreception Sciences, St. Pete Beach, Florida, April, 2010
第33回神経科学会大会、兵庫、2010年9月 味と匂学会第44回大会、福岡、2010年9月
第52回歯科基礎医学会学術大会、東京、2010年9月
The 8th International Symposium on Molecular and Neural Mechanisms of Taste and Olfactory, Fukuoka, December, 2010
先端歯学スクール2011、神奈川、2011年9月
その他の発表論文
楠原庸子, 安松啓子, 大栗弾宏, 堀尾奈央, 前田勝正, 二ノ宮裕三 マウス鼓索神経挫滅後の再生過程におけるうま味応答の回復 日本味と匂学会誌, 15 (3): 289-292, 2008
Ryusuke Yoshida, Nao Horio, Yoshihiro Murata, Keiko Yasumatsu, Noriatsu Shigemura, Yuzo Ninomiya
NaCl RESPONSIVE TASTE CELLS IN THE MOUSE FUNGIFORM TASTE BUDS Neuroscience, 159: 795-803, 2009
Tadahiro Ohkuri, Keiko Yasumatsu, Nao Horio, Masafumi Jyotaki, Robert F. Margolskee, Yuzo Ninomiya
Multiple sweet receptors and transduction pathways revealed in knockout mice by temperature and gurmarin sensitivity
Am. J. Physiol. Regul. Integr. Comp. Physiol, 296: (4) R960-971, 2009
Keisuke Sanematsu, Nao Horio, Yoshihiro Murata, Ryusuke Yoshida, Tadahiro Ohkuri, Noriatsu Shigemura, Yuzo Ninomiya
Moduration and Transmission of Sweet Taste Information for Enagy Homeostasis New York Academy of Sciences, 1170: 102-106, 2009
Ryusuke Yoshida, Keiko Yasumatsu, Shinya Shirosaki, Masafumi Jyotaki, Nao Horio, Yoshihiro Murata, Noriatsu Shigemura, Kiyohito Nakashima, Yuzo Ninomiya
Multiple receptor systems for umami taste in mice New York Academy of Sciences, 1170: 51-54, 2009
Keiko Yasumatsu, Nao Horio, Yoshihiro Murata, Shinya Shirosaki, Tadahiro Ohkuri, Ryusuke Yoshida, Yuzo Ninomiya
Multiple receptors underlie umami taste responses in mice.
Am. J. Clin. Nutr., 90: (3) 747S-752S, 2009
安尾敏明, 吉田竜介, 堀尾奈央, 重村憲徳, 二ノ宮裕三 マウス味細胞におけるGABAの機能解析
日本味と匂学会誌, 16: (3) 323-326, 2009
Nao Horio, Masafumi Jyotaki, Ryusuke Yoshida, Keisuke Sanematsu, Noriatsu Shigemura, Yuzo Ninomiya
Nutrient sensors in the gastrointestinal tract: Modulation of sweet taste sensitivity by leptin
Journal of Pharmacological Sciences, 112 (1): 8-12, 2010
Ryusuke Yoshida, Tadahiro Ohkuri, Masahumi Jyotaki, Toshiaki Yasuo, Nao Horio, Keiko Yasumatsu, Keisuke Sanematsu, Noriatsu Shigemura, Yamamoto T, Margolskee RF, Yuzo Ninomiya.
Endocannabinoids selectively enhance sweet taste.
Proc. Natl. Acad. Sci. USA, 12;107 (2): 935-939, 2010
堀尾 奈央, 日下部 裕子, 河合 崇行, 二ノ宮 裕三 Gα14ノックアウトマウスにおける味覚神経応答解析 味と匂学会誌, 18: (3) 213-216, 2011
その他の学会発表
日本味と匂学会第42回大会, 富山 (富山市民プラザ), 2008.09.17-20 日本味と匂学会第43回大会, 北海道 (旭川市民文化会館), 2009.09.02-04 第60回西日本生理学会, 福岡 (福岡市歯科医師会館), 2009.11.06-07
第87回日本生理学会大会, 岩手 (盛岡市民文化ホール,いわて県民情報交流セン ター), 2010.05.19-21
ECRO, England, 2011.09.07-10
第34回神経科学会学会, 神奈川, 2011.09.14-17
第52回歯科基礎医学会学術大会, 岐阜, 2011.09.30-10.02 第45回味と匂学会, 石川, 2011.10.05-07
第62回西日本生理学会, 佐賀, 2011.10.14-15
The 9th International Symposium on Molecular and Neural Mechanisms of Taste and Olfactory, Fukuoka (Kyushu University), 2011.11.04-06
要旨
味覚は、食を通じて健康を維持する上で不可欠の感覚である。腐敗物は酸味 を呈し、動物は摂取を拒否する。生体防御にとって重要なシグナルとなる酸味 の受容体については、未だその詳細は不明である。
酸味受容体候補分子として polycystic kidney disease 1-like 3 (PKD1L3) と polycystic kidney disease 2-like 1 (PKD2L1) が知られている。両分子は、舌咽神経 支配領域である舌後方部有郭乳頭味細胞では共発現するが、鼓索神経支配の舌 前方部茸状乳頭味細胞ではPKD2L1のみ発現する。味細胞は形態学的に I、II、
III型および基底細胞 (IV型) に分類されるが、PKD2L1は酸味応答を示すIII型 味細胞に発現し、両分子を強制発現させたHEK細胞では、酸を取り除いた後に 起こるオフ応答が確認されている。
本研究では、酸味受容における両分子の生体での役割を明らかにするため、
PKD2L1 と PKD1L3 の単独および両分子の遺伝子ノックアウト (KO) マウスを
作出し、電気生理学的解析を行った。
鼓索神経全神経束応答解析の結果、PKD2L1-KOマウスと両分子ダブルKOマ ウスではワイルドタイプ (WT) マウスと比べ、酸味応答が有意に減少したが、
その減少量は平均値レベルで25-45%程度だった。舌咽神経全線維束応答解析で は、酸味応答はいずれの KO マウスにおいても WT マウスとの差が見られなか った。塩味・苦味・甘味・うま味応答は両神経ともにマウス間に差が無かった。
また、舌咽神経での酸味オフ応答は、いずれの KO マウスにおいても有意に減 少した。
さらに、PKD2L1-KOマウスとWTマウスでの鼓索神経単一神経線維応答を解
析した結果、PKD2L1-KOマウスで酸味応答は有意に減少した。また、III型細胞 のマーカーである GAD67 を発現する細胞を緑色蛍光タンパク質 GFP により可 視 化 し た マ ウ ス (GAD67-GFP) と PKD2L1-KO マ ウ ス を 交 配 さ せ
GAD67-GFP+PKD2L1-KO マウスを作出し、ルーズパッチ法で茸状乳頭 III 型味
細胞応答を記録した結果、GAD67-GFP+PKD2L1-KOマウスで酸味応答は有意に 減少した。
これらの結果から、舌前方部鼓索神経領域の酸味応答に PKD2L1 が一部関与
し、PKD2L1を介する酸味応答は茸状乳頭でGAD67を発現する酸味応答味細胞
と、鼓索神経酸味応答単一神経線維を経由して伝達される可能性が示唆された。
また、舌後方部舌咽神経領域の酸味オフ応答にPKD1L3 とPKD2L1が関与する ことがKOマウスを用いた実験でも明らかとなった。
緒言
味覚は、消化管の入り口にある食調節センサーとして、生体恒常性を維持す るうえで極めて重要な役割を果たしている。甘味・塩味・うま味はカロリー源・
ミネラル源・タンパク質源の、苦味・酸味は毒物・腐敗物などのシグナルとし て感知され、動物に嗜好や忌避の行動をもたらす。これら味の受容により、快・
不快の情動や、唾液・消化液・ホルモンの分泌など様々な食調節系が働き、生 体の恒常性が維持されている。肥満、糖尿病、高血圧などの生活習慣病の増加 が問題となってきているが、食調節系の破綻は、それら生活習慣病をも導く。
近年の分子生物学的研究の進展に伴い、苦味受容体 (T2rs) (Adler et al., 2000;
Matsunami et al., 2000; Chandrashekar et al., 2000)、甘味・うま味受容体 (甘味:
T1R2+T1R3、うま味:T1R1+T1R3, mGluRs) (Nelson et al., 2001, 2002; Chaudhari et al., 1996, 2000; Damak et al., 2003; San Gabriel et al., 2005; Toyono et al., 2002, 2003;
Li et al., 2002)、塩味受容体 (ENaC) (Chandrashekar et al., 2010) が同定され、生体 の恒常性維持に働く味の受容や情報伝達の仕組みの理解が急速に進んできた。
しかし、生体での機能が同定された酸味受容体はまだない。
味細胞に発現する酸味受容体候補分子としてacid sensing ion channels (ASICs) (Ugawa et al., 2003) やhyperpolarization activated cyclic nucleotide gated potassium channels (HCNs) (Stevens et al., 2001)、potassium channels (Lin et al., 2004; Richter et al., 2004a)、5-nitro2-(3-phenylpropylamino)-benzoic acid (NPPB) 感受性 Cl- チャネ ル (Miyamoto et al., 1998)、polycystic kidney disease 1-like 3 and polycystic kidney disease 2-like 1 heteromers (PKD1L3+PKD2L1) (Ishimaru et al., 2006; Huang et al.,
2006) などが報告されている。これらのうち、マウス味蕾における網羅的な分子
発現解析 (Ishimaru et al., 2006) と、マウスゲノム配列情報をもとに味蕾特異的分 子のスクリーニング (Huang et al., 2006) により発見された transient receptor
potential (TRP) チャネルファミリーに属する PKD2L1 とその関連分子PKD1L3
(Clapham et al., 2003) は最も可能性の高い酸味受容体候補分子である。PKD1と
PKD2は、それらの遺伝子変異が常染色体優性多発性嚢胞腎の原因になることが 知られており (Delmas et al., 2004; Nauli et al., 2004)、PKD1とPKD2の二量体は、
機能的な受容体を形成するのに必要な形態である (Hanaoka et al., 2000)。これと
同様に、PKD1L3とPKD2L1も二量体として機能している可能性が考えられる。
実際に味細胞において、PKD1L3 と PKD2L1 の細胞表面への発現にその両分子 が必要となる (Ishimaru et al., 2006, 2010)。また、両分子を強制発現させたHEK 細胞は、カルシウムイメージングで酸味応答を示し、どちらか一方の分子のみ を発現させたHEK細胞では応答を示さない (Ishimaru et al., 2006; Inada et al., 2008; Ishii et al., 2009)。さらに、この両分子を発現するHEK細胞でおこる酸味応 答は刺激中に起こるオン応答ではなく、酸刺激除去後に起こるオフ応答であり (Inada et al., 2008)、そのオフ応答は過去の報告でも哺乳類での酸味刺激で引き起 こされることが示されている (DeSimon et al., 1995; Danilova et al., 2002; Lin et al.,
2002)。また、PKD1L3とPKD2L1の共発現の見られる有郭乳頭単離味細胞にお
いてもこのオフ応答が観察されている (Kawaguchi et al., 2010)。さらに、PKD2L1 発現味細胞をジフテリア毒素によって特異的に死滅させる遺伝子を導入したマ ウスでは鼓索神経酸味応答がほぼ消失することが報告されている (Huang et al.,
2006)。ヒトにおいては、酸味味盲患者の舌前方部味蕾にはPKD1L3とPKD2L1
の発現が消失している (Huque et al., 2009)。これらの報告はPKD2L1が酸味受容 に重要な役割を持つことを示唆している。
味覚の受容を担う味細胞は形態学的にI、II、III型および基底細胞 (IV型) に 分類される (Delay et al., 1986; Farbman, 1965; Murray, 1971, 1973) 。味細胞では、
PKD2L1は serotonin (5-HT) や neural cell adhesion molecule (NCAM)、ubiquitin
carboxy-terminal transferase (PGP9.5) といったIII型味細胞のマーカーと共発現し (Kataoka et al., 2008)、III型味細胞は酸味応答を示すことが知られている (Huang et al., 2008; Yoshida et al., 2009)。PKD2L1は舌咽神経支配領域である舌後方部の 有郭乳頭味細胞と、鼓索神経支配の舌前方部の茸状乳頭味細胞の両方で発現が 見られるが、PKD1L3は茸状乳頭味細胞では発現が見られず有郭乳頭味細胞のみ で発現が見られるため (Ishimaru et al., 2006; Huang et al., 2006)、PKD1L3は舌前 方部では機能していないと考えられる。よって、PKD2L1は舌前方部と後方部の、
PKD1L3は舌後方部の酸味受容に関与する可能性が示唆される。
そこで本研究では、酸味受容におけるPKD1L3 とPKD2L1の生体での機能解 明を目的とし、PKD2L1 と PKD1L3 の単独および両分子の遺伝子ノックアウト
(KO) マウスを作出し、免疫組織化学的手法、および RT-PCR により遺伝子が
KOされていることを確認後、味刺激に対する鼓索・舌咽神経応答解析と、茸状 乳頭III型味細胞を行った。
材料と方法
動物
実験動物にはPKD1L3-KOマウス (8-16週齢、体重20-32g) (Ishimaru et al., 2010)、
PKD2L1遺伝子のエクソン3からエクソン9を欠損させて作出したPKD2L1-KO
マウス (8-16 週齢、体重 20-32g)、両者の交配により作出した PKD1L3/PKD2L1 ダ ブ ル KO (PKD1L3/2L1-DKO) マ ウ ス (8-16 週 齢 、 体 重 20-32g) 、 PKD1L3/PKD2L1ダブルワイルドタイプ (WT) マウス (8-16週齢、体重20-32g) を用いた。また、GAD67 (GAD1) のプロモーター領域制御下に緑色蛍光タンパ ク質green fluorescent protein (GFP) を発現するGAD67-GFPマウス (8-16週齢、
体重20-32g) (Tamamaki et al., 2003) と、PKD1L3-KOマウス、PKD2L1-KOマウ スをそれぞれ交配させて作出した GAD67-GFP+PKD1L3-KO マウス (8-16 週齢、
体重20-32g)、 GAD67-GFP+PKD2L1-KOマウス (8-16週齢、体重 20-32g) を用 いた。なお、すべての実験は、九州大学歯学部・東京大学・Duke大学動物実験 委員会の許可のもとで、動物実験に関する指針に基づき遂行した。
サザンブロッティング
マウス尾先端 2~3 mm をチューブに回収し、DNeasy Blood & Tissue Kit
(QIAGEN、Hilden、Germany) を用いて DNA を抽出した後、制限酵素 BamHI
(Takara, Tokyo, Japan) で37 °C一晩処理し、DNAの断片化を行った。0.8 %アガ ロースゲルを用いて20Vで14~20時間電気泳動を行った後1 μg/mlのエチジウム ブロマイド溶液 (Wako Pure Chemical Industries, Osaka, Japan) をゲルの上にかけ、
15分間静置した。アルカリ塩溶液 (0.5 M NaOH、1.5 M NaCl) を入れたバットに ゲルを移し、30 分間振盪させて DNA の変性を行った後、中和溶液 (0.5 M
Tris-HCl [pH=7.2]、1.5 M NaCl、10 mM EDTA) で30分間振盪させた。その後、
1×TBE (44.5 mM Tris、44.5 mM ホウ酸、1 mM EDTA [pH=8.0] ) を入れた容器 中に下からアガロースゲル、ナイロンメンブレン、ろ紙の順番で重ね室温で一 晩 静 置 し 、 毛 細 管 現 象 を 利 用 し て メ ン ブ レ ン へ の 移 行 を 行 っ た 。 さ ら に Hybridization溶液 (50 % ホルムアミド、5×SSC [750 mM NaCl, 75 mM Sodium citrate]、5×デンハルト溶液、500 μg/ml サケ精子、250 μg/ml酵母トランスファー
RNA) を用いてプレハイブリダイゼーションを室温で1時間行った。digoxigenin
(DIG) 標識を行った特異的プローブ (表1) を95 °Cで5分間熱処理して1本鎖 にし、氷冷した。200 ng/mlの特異的プローブを加えた新たなHybridization溶液 にメンブレンを浸漬させ、ハイブリダイゼーションを 65 °C で一晩行った後、
Alkaline Phosphatase (AP) 標識 抗DIG抗体 (Roche, Mannheim, Germany) と反応 さ せ 、 そ の 後 4-nitroblue tetrazolium choloride (NBT) /5-bromo-4-choloro-3-indplyl-phosphate (BCIP) (Roche) と反応させることにより 発色を行った。
In situ ハイブリダイゼーション
In situ ハイブリダイゼーションは通法に基づき行った (Ishimaru et al., 2006)。 PKD1L3プローブはtransmembrane motif 2 (TM2) からC末端までをコードする 領域を含ませ、PKD2L1プローブとTRPM5プローブは全コード領域を含ませて 設計した。そして SP6 もしくは T7 RNA ポリメラーゼ (Roche, Mannheim,
Germany) を用いてDIG標識を行った。マウスを麻酔下で頸椎脱臼させ舌を摘出
した。摘出した舌を4 °Cで30~90分間、4 % パラホルムアルデヒド/PBS (137 mM NaCl、8.10 mM Na2HPO4・12H2O、2.68 mM KCl、1.47 mM KH2PO4、0.9 mM CaCl2・ 2H2O、0.33 mM MgCl2・12H2O[pH=7.5]) で固定し、4 °Cで一連のsucrose溶
液 (10 %で1時間、20 %で1時間、30 %で3時間) に浸漬した。その後、舌を O.C.T.compound (Sakura Finetechnical, Tokyo, Japan) に冷結包埋し、クライオスタ ットを使用して厚さ7 μmの舌凍結切片を作成し、シランコートしたスライドガ ラスを用いて連続凍結切片を作製した。室温で一連のエタノール (100 %、90 %、 80 %、70 %、50 %) に5分間ずつ浸漬し1×PBSで洗浄後、5 μg/ml Proteinase K で30分間処理し、1×PBSで10分間室温にて洗浄した。室温でビーカーに0.1 M
triethanolamine (pH=8.0) を加え、スターラーでよく撹拌し、スライドガラスを浸
して無水酢酸1 mlをシリンジで5分間かけて滴下し、さらに15分間撹拌した。
4×SSC (600 mM NaCl、60 mM Sodium citrate) で 10 分間室温で洗浄後、
Hybridization溶液 (50 %ホルムアミド、5×SSC、5×デンハルト溶液、500 μg/ml サ ケ精子、250 μg/ml酵母トランスファーRNA) でプレハイブリダイゼーションを 室温で1時間行った。スライドガラス上にDIG標識を行った200 ng/mlの特異的 プローブ (表2) を加えた新たなHybridization溶液を滴下し、切片にパラフィル ムをかぶせ、ハイブリダイゼーションを65 °Cで18時間行った。その後、5×SSC で5分、0.2×SSC (30 mM NaCl、3 mM Sodium citrate) で30分の洗浄を65 °C で それぞれ2回行った。続いてスライドガラスをTBS (50 mM Tris/HCl [pH=7.5]、 150 mM NaCl) に室温で5分間静置し、0.5 % blocking reagent (Roche) を含む Blocking溶液/TBSに30分間、AP標識 抗DIG抗体を含むBlocking溶液で60分 間室温にてインキュベートした。TNTバッファー (50 mM Tris/HCl [pH=7.5], 150 mM NaCl, 0.05 % Tween20) で5分間、3回洗浄後、NBT/BCIPと室温で24 時間反応させ、発色を行った。また、ネガティブコントロールとして、センス プローブを用いた (データは示さない)。
免疫組織化学染色
マウスを麻酔下で頸椎脱臼させ舌を摘出した。舌を4 °Cで30~90分間、4 % パラホルムアルデヒド/PBSで固定し、4 °Cで一連のsucrose溶液 (10 %で1時間、
20 %で1時間、30 %で3時間) に浸漬した。その後、舌をO.C.T.compoundに冷 結包埋後、クライオスタットを使用して厚さ9 μmの舌凍結切片を作成しシラン コートしたスライドガラスを用いて連続凍結切片を作製した。連続凍結切片を TNTバッファーで洗浄後、1 % blocking reagentに室温で1時間浸漬し、その後、
1 % blocking reagent を 含 む PKD1L3 抗 体 (rabbit polyclonal IgG; Lifespan Bioscience, WA, USA) も し く は PKD2L1 抗 体 (rabbit polyclonal IgG; Open Biosystems, AL, USA) もしくはGAD67抗体 (goat polyclonal IgG; SantaCruz, CA,
USA) を一次抗体として4 °Cで一晩免疫反応を行った。TNTバッファーで洗浄
後、二次抗体 (Alexa Fluor 555, donkey anti-rabbit IgG for PKD1L3 and PKD2L1;
Invitrogen, OR, USA および Alexa Fluor 546, donkey anti-goat IgG for GAD67;
Invitrogen) を室温で 2 時間反応させた。TNTバッファーで洗浄後、スライドガ
ラスに作業用バッファー (グリセロールを PBS 溶液にて 6 倍希釈したもの) を 滴下し、カバーガラスを被せ、共焦点レーザー顕微鏡 (FV-1000 及び Fluoview,
Olympus, Tokyo, Japan) を用いて観察した。ネガティブコントロールとして一次
抗体過程を省略した実験も同時に行った。
RT-PCR
マウスを麻酔下で頸椎脱臼させ舌を摘出した。舌上皮下に Tyrode 溶液 (140 mM NaCl、5 mM KCl、 1 mM CaCl2、1 mM MgCl2、10 mM HEPES、10 mM Glucose、10 mM ピルビン酸ナトリウム、pH=7.4) にて溶解した0.2~1 mg/mlエ ラスターゼ (Elastin Products, Owensville, USA) を注入し、室温にて10~15分間放
置した。その後、ピンセットで素早く舌上皮を剥離し、粘膜側を下にしてシリ コンコートした培養皿にピンで止め、Tyrode 溶液で数回洗浄した。この舌上皮 より単離した茸状乳頭ならびに有郭乳頭の味蕾をそれぞれ30個ずつガラスピペ ットにてチューブに回収した。また、味蕾を含まない1 mm×1 mmの舌上皮も チューブに回収した。その後、RNeasy Plus Mini Kit (QIAGEN、Ratingen、Germany) を用いてRNA精製を行った。OneStep RT-PCR Kit (QIAGEN) および遺伝子特 異的プライマー (表2) を用いてRT-PCRを行った (RT:50 °C 30分、PCR:94 °C 30秒、53 °C 60秒、72 °C 90秒を30サイクル)。さらにnested-PCRにより遺伝 子特異的配列を増幅した。サンプルは以下のように増幅した:94 °C 30秒、60 °C 30 秒、72 °C 60 秒を 35 サイクル。各反応混合物は 0.25 Units の Taq DNA polymerase (TaKaRa Ex TaqTM HS: Takara), 1 l の 20 mM Mg2+含有10× PCR buffer, 0.2 mM のdNTP, 0.6 mMの internal primer pair (Table 1) and 0.2 lのfirst
round PCR products。その後、2 %アガロースゲルを用いて30分間電気泳動を行
い、DNAを検出した。
鼓索神経・舌咽神経応答の記録
神経応答の記録実験には、PKD1L3-KOマウス (CT;n=5, GL;n=6)、PKD2L1-KO マウス (CT;n=7, GL;n=6)、PKD1L3/PKD2L1-DKOマウス (CT;n=6, GL;n=6)、WT マウス (CT;n=8, GL;n=8) を用いた。マウスはペントバルビタール麻酔下で手術 し、鼓索または舌咽神経を電気生理学的記録のため剖出した。剖出の手順や記 録は通法に従い行った (Ninomiya et al., 1984, 1991; Ninomiya 1996)。動物をネン ブタール (40-50 mg/kg) 腹腔内投与にて麻酔し、ヘッドホルダーで仰臥位に固定 後、気管カニューレを装着した。内側翼突筋の除去によって舌神経から枝分か れしている右側鼓索神経を露出させた。鼓索神経はその後、鼓室に入る直前で
切断し、周囲の組織から分離、露出させた。顎二腹筋と舌骨の除去によって右 側舌咽神経を露出させた。舌咽神経はその後、頚静脈孔に入る直前で切断し、
周囲の組織から分離、露出させた。これらの神経をそれぞれ、全神経線維束記 録のため銀塩化銀電極に乗せた。不関電極は周囲組織に装着した。舌への味刺 激に対する神経応答は増幅器 (K-1; Iyodenshikogaku, Nagoya, Japan)、オーディオ モニターを介して、オシロスコープにて観察した。全神経線維束応答はインテ グ レ ー タ ー に て 積 分 し 、PowerLab System (PowerLab/sp4; AD Instruments,
Australia) を使用して、後の解析のためにコンピューターに記録した。
鼓索神経単一神経線維応答では、鼓索神経をピンセットにて数本に分割後、
銀-塩化銀電極にのせ、応答を記録した。
鼓索神経・舌咽神経応答の舌への化学刺激
鼓索神経が支配する茸状乳頭を化学刺激するために、舌の前方半分をシリコ ンチューブ製のフローチャンバーで囲んだ (Ninomiya and Funakoshi, 1981)。舌咽 神経が支配する有郭乳頭と葉状乳頭を化学刺激するために、まず口角に切れ込 みを入れ乳頭を露出させ舌の先端を縫合してわずかに引っ張ることで乳頭の溝 を開いた。溶液は重力流でそれぞれの部分に流した。
化学刺激として使った溶液は以下の通りである:100 mM NH4Cl、1-20 mM HCl、 1-50 mM citric acid、1-100 mM acetic acid、10-1000 mM sucrose、10-500 mM NaCl、
0.1-10 mM quinine-HCl (QHCl)、10-1000 mM monosodium glutamate (MSG) (Wako Pure Chemical Industries, Osaka, Japan)、10-1000 mM monopotassium glutamate (MPG) (Sigma, MO, USA)。これらの溶液は蒸留水で解し、室温 (25 °C) で使用し た。
刺激溶液は~0.1 ml/sで30秒間 (鼓索神経)、または60秒間 (舌咽神経) 流し、
その後、舌を60秒間 (鼓索神経) または120秒間 (舌咽神経) 蒸留水で洗浄した。
基準刺激の100 mM NH4Clに対する応答を周期的にモニターし、応答の安定性を 確認した。つまり、各刺激溶液シリーズの始めと終わりに記録した NH4Cl 応答
の大きさが15 %以下の変化のときに安定的と考え、その間に記録した各種刺激 に対する応答をデータ解析に用いた。
鼓索神経・舌咽神経応答のデータ解析
鼓索・舌咽神経全神経束応答の解析は、応答をインテグレーター (時定数=1.0 秒) にて積分し、刺激開始後5秒~25秒 (鼓索神経) または5秒~40秒 (舌咽神 経) までの大きさの平均値を求め、100 mM NH4Cl応答を1.0とした相対値で算 出した。これら相対応答値を統計解析に用いた。基準溶液として、過去の報告 (Damak et al., 2003) と同様に100 mM NH4Clを選んだ。
鼓索神経単一神経線維応答の解析は、10 秒間の味刺激への応答発火数から味 刺激開始前10秒間の自発発火数を差し引いて、正味の応答発火数を求めた。応 答発生の基準は以下の2条件を満たすこととした : (1) 2回の同一味刺激に対し 発生したスパイク数が、共に自発発電のスパイク数の平均+2SD よりも多いこ と、(2) 味刺激により少なくとも3回以上スパイクが生じること、を条件づけた。
遺伝子 KO の効果を調べるために、two-way repeated measures analysis of
variance (ANOVA) を用いて統計処理を行った。 ANOVAで有意差が得られたと
き、ポストホック Dunnett テストにより各平均値の比較を行った。P 値が 0.05 以下のとき、統計的に有意であると判断した。計算は Microsoft Excel2007 を用 いて行った。
味細胞応答の記録
マウスをジエチルエーテル麻酔下で頸椎脱臼させ舌を摘出した。舌上皮下に
Tyrode 溶液にて溶解した 0.2~1 mg/ml エラスターゼを注入し、室温にて 10~15
分間インキュベートした。その後、ピンセットで素早く舌上皮を剥離し、粘膜 側を下にしてシリコンコートした培養皿にピンで止め、Tyrode 溶液で数回洗浄 した。この舌上皮より茸状乳頭単一味蕾を採取し、記録チャンバーに移した。
残りの茸状乳頭味蕾を含む舌上皮は次のシリーズの実験を行うまで4 °Cにて保
存した。採取された味蕾を含んだ記録チャンバーを共焦点レーザー顕微鏡のス テージに乗せ、顕微鏡観察下で味蕾の粘膜側 (味孔) を刺激ピペットに吸引・保 持した。記録チャンバー内は常にTyrode溶液を灌流させ、刺激ピペット内は味 応答記録時を除き、Tyrode 溶液を灌流させた。共焦点レーザー顕微鏡観察下で 単一味蕾中のGFP発現細胞を同定し、基底膜側より記録電極 (φ1~3 μm、1.5〜
3.5 Ω) を当て、シール抵抗が3~10倍になる程度に吸引し、味応答記録を行った。
味刺激には 1~20 mM HCl、1~30 mM citric acid, 3~50 mM acetic acidを用いた。ま た、味刺激後は、最低でも30秒以上、蒸留水にて刺激ピペット内を灌流、洗浄 した。これらの実験はすべて室温にて行った。
味細胞応答のデータ解析
記録したデータは10秒ごとのスパイク数をカウントし、味刺激前後の蒸留水 を与えた期間の平均スパイク数を自発放電頻度とした。味刺激の投与後10秒間 のスパイク数から自発放電頻度を差し引いた値を刺激に対する応答値とした。
応答発生の基準は以下の2条件を満たすこととした : (1) 2回の同一味刺激に対 し発生したスパイク数が、共に自発発電のスパイク数の平均+2SD よりも多い こと、(2) 味刺激により少なくとも3回以上スパイクが生じること、を条件づけ た。
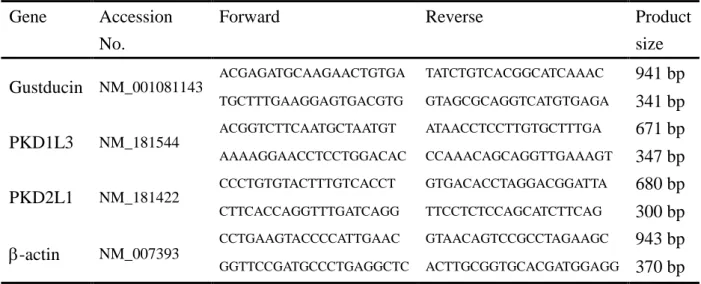
Gene Forward Reverse Product size
PKD2L1
wild-type allele
ttctggtccagtttgctcag catcaagtcccaggagtcaa 6.6kb
mutated allele
ttgatctgcaatgcaatgaacc ccttattccaagcggcttcggccagtaacg 4.4kb
表1 サザンブロッティングに用いたプローブ作成用プライマーの塩基配列 すべての配列は5’→3’で示す。
Gene Accession No.
Forward Reverse Product
size Gustducin NM_001081143
ACGAGATGCAAGAACTGTGA TATCTGTCACGGCATCAAAC 941 bp
TGCTTTGAAGGAGTGACGTG GTAGCGCAGGTCATGTGAGA 341 bp PKD1L3 NM_181544
ACGGTCTTCAATGCTAATGT ATAACCTCCTTGTGCTTTGA 671 bp
AAAAGGAACCTCCTGGACAC CCAAACAGCAGGTTGAAAGT 347 bp PKD2L1 NM_181422
CCCTGTGTACTTTGTCACCT GTGACACCTAGGACGGATTA 680 bp
CTTCACCAGGTTTGATCAGG TTCCTCTCCAGCATCTTCAG 300 bp
-actin NM_007393 CCTGAAGTACCCCATTGAAC GTAACAGTCCGCCTAGAAGC 943 bp
GGTTCCGATGCCCTGAGGCTC ACTTGCGGTGCACGATGGAGG 370 bp
表2 RT-PCRに用いたプライマーの塩基配列
すべての配列は5’→3’で示す。
Upper: outside primers Lower: inside primers
結果
PKD1L3-KOマウス、PKD2L1-KOマウスの作出
PKD1L3-KOマウスは、共同研究者であるDuke大学の松波宏明先生、東京大
学の石丸喜朗先生よりご提供いただいた (Ishimaru et al., 2010)。
PKD2L1-KOマウスは、exon3からexon9を欠損させて作出した (図1.A)。サ
ザンブロットにより、PKD2L1+/+、PKD2L1+/-、PKD2L1-/-を分類した (図1.B)。in situ hybridizationにより、PKD2L1+/+マウスでは有郭乳頭においてPKD1L3、
PKD2L1、甘味・うま味・苦味を受容するII型細胞のマーカーであるTRPM5
(transient receptor potential cation channel subfamily M member 5) のmRNAの発現 が見られた (図1.C)。一方PKD2L1-/-マウス (PKD2L1-KOマウス) ではPKD2L1 のmRNAの発現が見られず、PKD1L3とTRPM5のmRNAの発現は見られた (図 1.C)。
PKD1L3-KOマウスとPKD2L1-KOマウスはそれぞれGFPまたはmCherryがノ ックインされているが (Ishimaru et al., 2010; 図1.A)、これらマウスにおいてGFP
もしくはmCherryの発現は観察されなかった (データは示さない)。
図1 PKD2L1-KOマウスの作出とPKD2L1の発現解析 A. PKD2L1-KOマウスの作出方法。
PKD2L1-KOマウスは、PKD2L1遺伝子のexon3からexon9を欠損させることで作出 した。TM: transmembrane motif, Ex: exon; Cre: Cre recombinase gene; Neo: neomycin resistant gene; loxP: loxP site; DT-A: diphtheria toxin A-chain gene.
B. WTとPKD2L1欠損アレルを識別するプローブを用いてサザンブロットを行った結
果。
PKD2L1+/+ (WT)、PKD2L1+/-、PKD2L1-/- (PKD2L1-KO) マウスがそれぞれ分類できた。
C. WTマウスとPKD2L1-KOマウスを用いて有郭乳頭でのin situ hybridizationを行った 結果。
WTマウスではPKD1L3、PKD2L1、TRPM5のmRNA発現が確認できた。一方、
PKD2L1 KOマウスではPKD1L3、TRPM5のmRNA発現は見られたが、PKD2L1の mRNA発現は見られなかった。Scale barは20μmを示す。
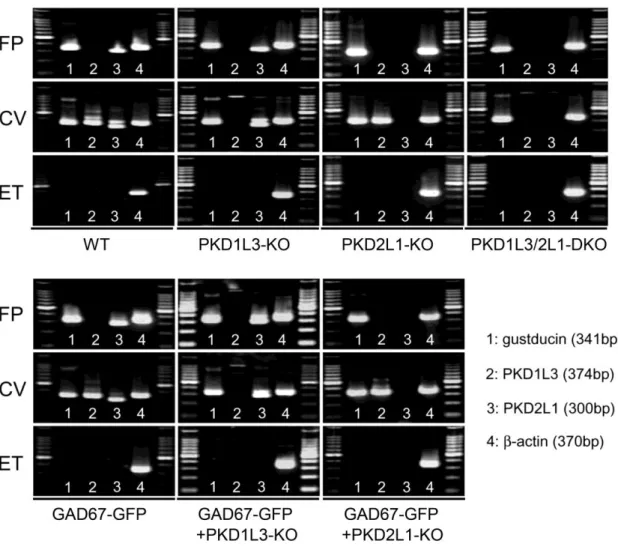
PKD1L3/2L1-DKOマウス、GAD67-GFP+PKD1L3-KOマウス、および GAD67-GFP+PKD2L1-KOマウスの作出と、RT-PCRによる味細胞における PKD1L3、PKD2L1のmRNA発現解析
PKD1L3-KO (PKD1L3-/-) マウスとPKD2L1-KO (PKD2L1-/-) マウスを交配させ、
PKD1L3+/-+PKD2L1+/-マウスを作出した。このマウスどうしをさらに交配させ、
PKD1L3-/-+PKD2L1+/+マウス (PKD1L3-KO)、PKD1L3+/++PKD2L1-/-マウス (PKD2L1-KO)、PKD1L3-/-+PKD2L1-/-マウス (PKD1L3/2L1-DKO)、WTマウスを 作出し、実験に用いた。
in situ hybridizationにより、PKD2L1は有郭乳頭味細胞と茸状乳頭味細胞の両
方で発現が見られるが、PKD1L3は茸状乳頭味細胞では発現が見られず有郭乳頭 味細胞のみで発現が見られることが報告されている (Ishimaru et al., 2006; Huang
et al., 2006)。作出したマウスを用いてRT-PCRでmRNA発現解析を行ったとこ
ろ、WTマウスでは過去の報告と同様に、PKD2L1は有郭乳頭味細胞と茸状乳頭 味細胞の両方で発現が見られたが、PKD1L3は茸状乳頭味細胞では発現が見られ ず有郭乳頭味細胞のみで発現が見られた (図2)。また、PKD1L3-KOマウスでは、
茸状・有郭乳頭味細胞でPKD1L3の発現が見られず、PKD2L1-KOマウスでは茸 状・有郭乳頭味細胞でPKD2L1の発現が見られなかった (図2)。また、
PKD1L3/2L1-DKOマウスでは、茸状・有郭乳頭味細胞でPKD1L3とPKD2L1両
方の発現が見られなかった (図2)。このように、それぞれのKOマウスではKO した遺伝子のmRNA発現が消失した。
また、味細胞ではPKD2L1は5-HTやNCAM、PGP9.5といったIII型味細胞の マーカーと共発現し (Kataoka et al., 2008)、GAD67はIII型味細胞に発現するこ とが知られている (DeFazio et al., 2006; Tomchik et al., 2007)。そこでIII型味細胞 を用いた実験を行うため、GAD67(GAD1)のプロモーター領域制御下に緑色蛍
光タンパク質GFPを発現するGAD67-GFPマウス (Tamamaki et al., 2003) と、
PKD1L3-KO、PKD2L1-KOマウスをそれぞれ交配させて
GAD67-GFP+PKD1L3-KOマウス、 GAD67-GFP+PKD2L1-KOマウスを作出した。
作出したマウスを用いてRT-PCRでmRNA発現解析を行ったところ、GAD-GFP マウスではPKD2L1は有郭乳頭味細胞と茸状乳頭味細胞の両方で発現が見られ
たが、PKD1L3は茸状乳頭味細胞では発現が見られず有郭乳頭味細胞のみで発現
が見られた (図2)。また、GAD67-GFP+PKD1L3-KOマウスでは、茸状・有郭乳 頭味細胞でPKD1L3の発現が見られず、GAD67-GFP+PKD2L1-KOマウスでは茸 状・有郭乳頭味細胞でPKD2L1の発現が見られなかった (図2)。このように、
各KOマウスではKOした遺伝子のmRNA発現が消失した。
図2 RT-PCRによるKOマウスにおけるPKD1L3、PKD2L1のmRNA発現解析
WTマウス、GAD-GFPマウスではPKD2L1は有郭乳頭味細胞と茸状乳頭味細胞の両 方で発現が見られたが、PKD1L3は茸状乳頭味細胞では発現が見られず有郭乳頭味細胞 のみで発現が見られた。また、PKD1L3-KOマウスとGAD67-GFP+PKD1L3-KOマウス では、茸状・有郭乳頭味細胞でPKD1L3の発現が見られず、PKD2L1-KOマウスと
GAD67-GFP+PKD2L1-KOマウスでは茸状・有郭乳頭味細胞でPKD2L1の発現が見られ
なかった。
味細胞特異的に発現が見られるgustducin (McLaughlin et al., 1992) はすべてのマウス において、茸状乳頭と有郭乳頭味細胞で発現が見られたが舌上皮では発現が見られなか った。また、すべてのマウスにおいてハウスキーピング遺伝子のひとつであるβ-actin の発現を確認した。
FP: 茸状乳頭味細胞, CV: 有郭乳頭味細胞, ET: 舌上皮細胞, 100bpマーカーを使用
GAD67-GFPマウス、GAD67-GFP+PKD1L3-KOマウス、
GAD67-GFP+PKD2L1-KOマウスでの免疫組織化学的手法によるPKD1L3、
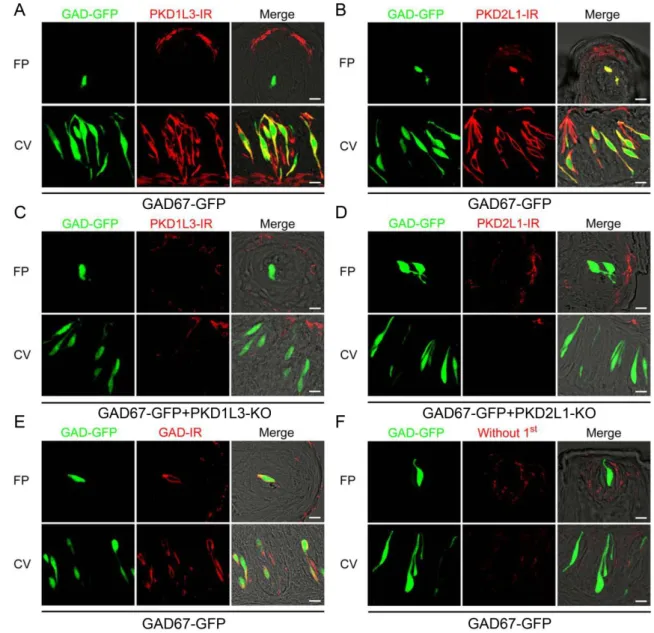
PKD2L1、GAD-IR (immunoreactivity) のタンパク質発現解析
さらに、作出したマウスを用いて免疫組織化学染色でタンパク質発現解析を 行った。PKD2L1はIII型味細胞のマーカーと共発現し (Kataoka et al., 2008)、
GAD67はIII型味細胞に発現することが知られているため (DeFazio et al., 2006;
Tomchik et al., 2007)、GAD67をIII型味細胞のマーカーとして用い、III型細胞に
おけるPKD1L3、PKD2L1、GAD67-IRの発現を解析した。WTマウスにおいて、
茸状乳頭ではGAD67-GFP発現細胞の95.3 %がGAD67免疫陽性
(immunoreactivity, IR) を示し、有郭乳頭ではGAD67-GFP発現細胞の98.1 %が GAD67-IRを示した (図3.E, 表3)。また、GAD67-GFP+PKD1L3-KOマウスにお いて、茸状乳頭ではGAD67-GFP発現細胞の97.8 %がGAD67-IRを発現し、有郭 乳頭ではGAD67-GFP発現細胞の98.7 %がGAD67-IRを示した (図には示さない,
表3)。GAD67-GFP+PKD2L1-KOマウスにおいては、茸状乳頭ではGAD67-GFP
発現細胞の94.7 %がGAD67-IRを示し、有郭乳頭ではGAD67-GFP発現細胞の
99.3 %がGAD67-IRを示した (図には示さない, 表3)。このようにWTマウスに
よる過去の報告 (Tamamaki et al., 2003) と同様、KOマウスでも茸状・有郭乳頭 ともにほぼすべてのGAD67-GFP発現細胞がGAD67-IRを示した。よって
GAD67-GFPをIII型味細胞のマーカーとして用い、GAD67-GFP発現細胞におけ
るPKD1L3、PKD2L1の発現を解析した。
GAD67-GFPマウスでは、PKD1L3は茸状乳頭ではGAD67-GFP発現細胞で発
現が見られなかったが、有郭乳頭ではGAD67-GFP発現細胞の96.8 %で発現が見 られた (図3.A, 表3)。また、PKD2L1は茸状・有郭両乳頭においてGAD67-GFP 発現細胞の95.7 %および99.4 %で発現が見られた (図3.B, 表3)。
GAD67-GFP+PKD1L3-KOマウスでは、茸状・有郭乳頭ともにGAD67-GFP発現 細胞でPKD1L3の発現が見られず (図3.C, 表3)、GAD67-GFP+PKD2L1-KOマ ウスでは茸状・有郭乳頭味細胞ともにGAD67-GFP発現細胞でPKD2L1の発現 が見られなかった (図3.D, 表3)。以上の結果は、各KOマウスではKOした遺 伝子のタンパク質発現が見られないことを示す。
図3 免疫組織化学的手法によるKOマウスにおけるPKD1L3、PKD2L1、GAD67のタン パク質発現解析
A. GAD67-GFPマウスにおけるPKD1L3の発現解析の結果。
GAD67-GFPマウスにおいて、PKD1L3は茸状乳頭ではGAD67-GFP発現細胞との共 発現が見られず、有郭乳頭ではGAD67-GFP発現細胞との共発現が見られた。
B. GAD67-GFPマウスにおけるPKD2L1の発現解析の結果。
GAD67-GFPマウスにおいて、PKD2L1は茸状・有郭両乳頭でGAD67-GFP発現細胞 との共発現が見られた。
C. GAD67-GFP+PKD1L3-KOマウスにおけるPKD1L3の発現解析の結果。
GAD67-GFP+PKD1L3-KOマウスでは、PKD1L3は茸状・有郭両乳頭でGAD67-GFP 発現細胞と共発現が見られなかった。
D. GAD67-GFP+PKD2L1-KOマウスにおけるPKD2L1の発現解析の結果。
GAD67-GFP+PKD2L1-KOマウスでは、PKD2L1は茸状・有郭両乳頭でGAD67-GFP 発現細胞と共発現が見られなかった。
E. GAD67-GFPマウスにおけるGAD67の発現解析の結果。
GAD67-GFPマウスにおいて、GAD67は茸状・有郭両乳頭でGAD67-GFP発現細胞
との共発現が見られた。
F. GAD67-GFPマウスにおける一次抗体不使用のネガティブコントロール結果。
GAD67-GFPマウスにおいて、一次抗体を使用しなかったところ、茸状・有郭両乳
頭でGAD67-GFP発現細胞と共発現するシグナルは見られなかった。
FP: 茸状乳頭, CV: 有郭乳頭, ET: 舌上皮細胞, GAD-GFP: GAD67-GFP, Scale barは 10μmを示す。
GAD67-GFP GAD67-GFP +PKD2L1-KO
GAD67-GFP +PKD1L3-KO
FP % CV % FP % CV % FP % CV %
PKD2L1-IR /GAD67-GFP
45
/ 47 95.7 164
/ 165 99.4 0
/ 45 0 0
/ 143 0 - - - -
PKD1L3-IR /GAD67-GFP
0
/ 62 0 151
/ 156 96.8 - - - - 0
/ 46 0 0 / 153 0 GAD67-IR
/GAD67-GFP 41
/ 43 95.3 153
/ 156 98.1 44
/ 45 97.8 220
/ 223 98.7 36
/ 38 94.7 143 / 144 99.3
表3免疫組織化学的手法によるKOマウスにおけるPKD1L3、PKD2L1、GAD67のタン パク質発現解析のまとめ
GAD67-GFPマウス、GAD67-GFP+PKD1L3-KOマウス、GAD67-GFP+PKD2L1-KOマ ウスでのGAD67-GFP、 PKD2L1、PKD1L3、GAD67の発現細胞数と、GAD67-GFP発 現細胞におけるPKD2L1、PKD1L3、GAD67の発現割合をそれぞれ示した。それぞれの データは2~3匹のマウスより得られた。-は解析していないことを示す。
FP: 茸状乳頭, CV: 有郭乳頭
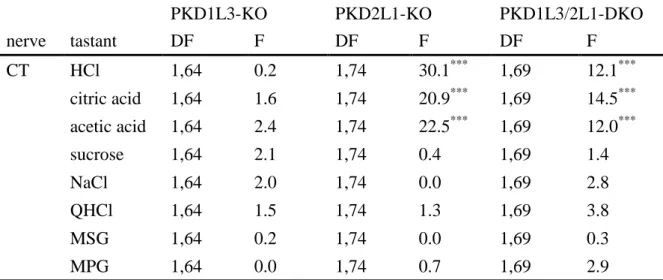
WTマウス、PKD1L3-KOマウス、PKD2L1-KOマウス、およびPKD1L3/2L1-DKO マウスにおける鼓索神経全神経束味応答解析
WTマウス、PKD1L3-KOマウス、PKD2L1-KOマウス、およびPKD1L3/2L1-DKO マウスを用いて、舌前方部を支配する鼓索神経応答解析を行った。いずれのマ ウスにおいても、酸味 (HCl, citric acid, acetic acid)、塩味 (NaCl)、甘味 (sucrose)、
苦味 (QHCl)、うま味 (MSGおよびMPG) に対する濃度依存的応答が見られた
(図4, 5)。PKD2L1-KOマウスとPKD1L3/2L1-DKOマウスでは、すべての酸味物
質 (HCl, citric acid, acetic acid) に対する応答がWTマウスと比べて有意に減少し た (図5, 表4)。その減少量をそれぞれの応答の平均値を用いて計算したところ、
PKD2L1-KOマウス、PKD1L3/2L1-DKOマウスともに平均値レベルで25-45%程
度だった(表5)。塩味・苦味・甘味・うま味応答には変化が無かった (図5, 表 4)。
図4 WTマウス、PKD1L3-KOマウス、PKD2L1-KOマウス、PKD1L3/2L1-DKOマウス における鼓索神経応答記録例
いずれのマウスにおいても酸味 (HCl, citric acid, acetic acid)、塩味 (NaCl)、甘味 (sucrose)、苦味 (QHCl)、うま味 (MSGおよびMPG, MPGは図に示さない) に対する応 答が見られた。NH4Clは基準溶液として用いた。
NH4Cl: 100 mM, HCl: 10 mM, citric acid: 10 mM, acetic acid: 30 mM, sucrose: 500 mM, NaCl: 100 mM, QHCl: 10 mM, MSG: 100 mM
記録例中のbarは30秒を示す。
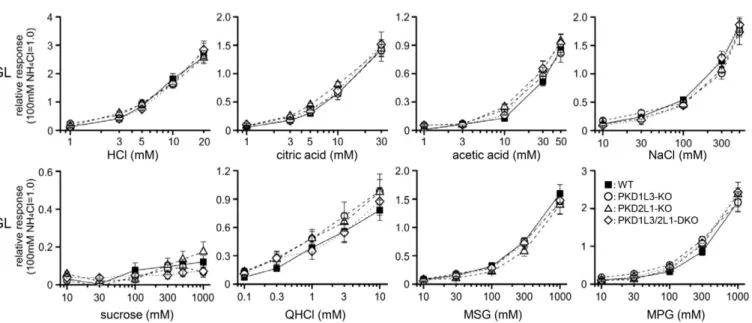
図5WTマウス (n=8)、PKD1L3-KOマウス (n=5)、PKD2L1-KOマウス (n=7)、
PKD1L3/2L1-DKOマウス (n=6) における鼓索神経応答の濃度応答曲線
PKD2L1-KOマウスとPKD1L3/2L1-DKOマウスでは、WTマウスと比べ、HCl、citric acid、acetic acidのすべての酸味応答が有意に減少した (two-way ANOVA; P<0.001: 表3、
post hoc Dunnett’s tests; *: P<0.05, **: P<0.001 for PKD2L1-KOおよび+: P<0.05, ++: P<0.01 for PKD1L3/2L1-DKO)。塩味・苦味・甘味・うま味応答は変化が無かった (two-way ANOVA; P>0.05: 表3)。データは平均値±Standard Error of the Mean (S.E.M.) で示す。
CT: 鼓索神経
PKD1L3-KO PKD2L1-KO PKD1L3/2L1-DKO
nerve tastant DF F DF F DF F
CT HCl 1,64 0.2 1,74 30.1*** 1,69 12.1***
citric acid 1,64 1.6 1,74 20.9*** 1,69 14.5***
acetic acid 1,64 2.4 1,74 22.5*** 1,69 12.0***
sucrose 1,64 2.1 1,74 0.4 1,69 1.4
NaCl 1,64 2.0 1,74 0.0 1,69 2.8
QHCl 1,64 1.5 1,74 1.3 1,69 3.8
MSG 1,64 0.2 1,74 0.0 1,69 0.3
MPG 1,64 0.0 1,74 0.7 1,69 2.9
表4 WTマウス、PKD1L3-KOマウス、PKD2L1-KOマウス、PKD1L3/2L1-DKOマウス の各味刺激に対する鼓索神経相対応答の分散分析結果
鼓索神経相対応答におけるtwo-way ANOVA解析での自由度とF値を示した。
PKD2L1-KOマウスとPKD1L3/2L1-DKOマウスでは、WTマウスと比べ、HCl、citric acid、
acetic acidのすべての酸味応答が有意に減少した (two-way ANOVA; ***P<0.001)。塩 味・苦味・甘味・うま味応答は変化が無かった (two-way ANOVA; P>0.05)。図5に示 したデータを使用した。
CT: 鼓索神経, DF: 自由度, F: F値
PKD2L1-KO PKD1L3/2L1-DKO
nerve tastant KO/WT KO/WT
CT HCl 0.72 0.77
citric acid 0.55 0.65
acetic acid 0.56 0.56
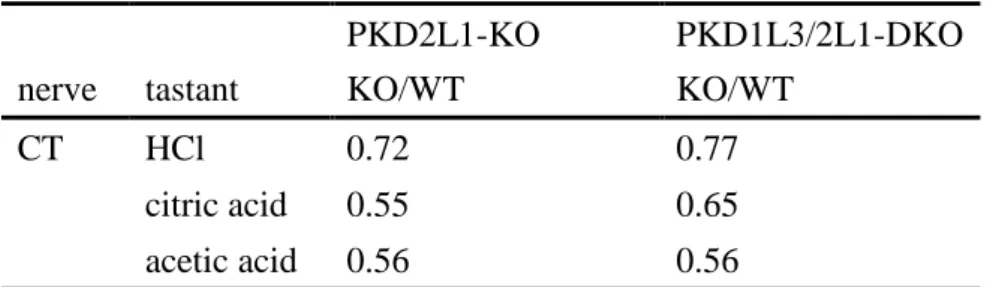
表5 PKD2L1-KOマウス、PKD1L3/2L1-DKOマウスにおける各種酸味刺激に対する鼓索 神経相対応答の平均値レベルでの変化量
PKD2L1-KOマウスとPKD1L3/2L1-DKOマウスでは、WTマウスと比べ、HCl、citric acid、acetic acidのすべての酸味応答が有意に減少した (two-way ANOVA; ***P<0.01)。
その減少量はPKD2L1-KOマウスとPKD1L3/2L1-DKOマウスともに平均値レベルで 25-45%程度だった。
HCl (10 mM), citric acid (10 mM), acetic acid (30 mM) のデータの平均値を使用。
CT: 鼓索神経
WTマウス、PKD1L3-KOマウス、PKD2L1-KOマウス、およびPKD1L3/2L1-DKO マウスにおける舌咽神経全神経束味応答解析
WTマウス、PKD1L3-KOマウス、PKD2L1-KOマウス、およびPKD1L3/2L1-DKO マウスを用いて、舌後方部を支配する舌咽神経応答解析を行った。いずれのマ ウスにおいても、酸味 (HCl, citric acid, acetic acid)、塩味 (NaCl)、甘味 (sucrose)、
苦味 (QHCl)、うま味 (MSGおよびMPG)に対する濃度依存的応答が見られた
(図6, 7)。酸味応答はすべてのKOマウスでWTマウスとの差が見られなかった
(図7, 表6)。塩味・苦味・甘味・うま味応答も変化が無かった (図7, 表6)。
図6 WTマウス、PKD1L3-KOマウス、PKD2L1-KOマウス、PKD1L3/2L1-DKOマウス における舌咽神経応答記録例
いずれのマウスにおいても酸味 (HCl, citric acid, acetic acid)、塩味 (NaCl)、甘味 (sucrose)、苦味 (QHCl)、うま味 (MSGおよびMPG, MPGは図に示さない) に対する応 答が見られた。NH4Clは基準溶液として用いた。
NH4Cl: 100 mM, HCl: 10 mM, citric acid: 10 mM, acetic acid: 30 mM, sucrose: 500 mM, NaCl: 100 mM, QHCl: 10 mM, MSG: 100 mM
記録例中のbarは30秒を示す。
図7 WTマウス (n=8)、PKD1L3-KOマウス (n=6)、PKD2L1-KOマウス (n=6)、
PKD1L3/2L1-DKOマウス (n=6) における舌咽神経応答の濃度応答曲線
酸味応答はすべてのKOマウスでWTマウスとの差が見られなかった (two-way ANOVA; P>0.05: 表6)。塩味・苦味・甘味・うま味応答も変化が無かった (two-way ANOVA; P>0.05: 表6)。データは平均値±S.E.M.で示す。
GL: 舌咽神経
PKD1L3-KO PKD2L1-KO PKD1L3/2L1-DKO
nerve tastant DF F DF F DF F
GL HCl 1,69 0.7 1,69 0.0 1,69 0.0
citric acid 1,69 0.2 1,69 3.8 1,69 0.6
acetic acid 1,69 0.9 1,69 3.8 1,69 2.9
Sucrose 1,69 1.7 1,69 0.6 1,69 2.3
NaCl 1,69 0.6 1,69 2.2 1,69 0.2
QHCl 1,69 3.8 1,69 3.4 1,69 0.4
MSG 1,69 0.3 1,69 3.4 1,69 0.7
MPG 1,69 2.6 1,69 0.7 1,69 2.7
表6 WTマウス、PKD1L3-KOマウス、PKD2L1-KOマウス、PKD1L3/2L1-DKOマウス の各味刺激に対する舌咽神経相対応答の分散分析結果
舌咽神経相対応答におけるtwo-way ANOVA解析での自由度とF値を示した。酸味応 答はすべてのKOマウスでWTマウスとの差が見られなかった (two-way ANOVA;
P>0.05)。塩味・苦味・甘味・うま味応答も変化が無かった (two-way ANOVA; P>0.05)。
図7に示したデータを使用した。
GL: 舌咽神経, DF: 自由度, F: F値
WTマウス、PKD1L3-KOマウス、PKD2L1-KOマウス、およびPKD1L3/2L1-DKO マウスにおける舌咽神経全神経束オフ応答解析
PKD1L3とPKD2L1の両分子を強制発現させたHEK細胞では、酸を取り除い
た後に起こるオフ応答が確認されている (Inada et al., 2008)。両分子は、舌咽神 経支配領域である舌後方部有郭乳頭味細胞では共発現するが、鼓索神経支配の 舌前方部茸状乳頭味細胞ではPKD2L1のみ発現する。そこで、WTマウス、
PKD1L3-KOマウス、PKD2L1-KOマウス、およびPKD1L3/2L1 DKOマウスを用
いて、舌後方部を支配する舌咽神経における酸味オフ応答解析を行った。舌咽 神経全神経束応答をインテグレーター (時定数=1.0秒) にて積分し、酸味刺激除 去後10秒間の応答曲線下面積 (area under the curve: AUC) を求め、同様に求め
た100 mM NH4Cl後のAUCを1.0とした相対値で算出し、この値をオフ応答の
大きさと定義した (図8. A)。その結果、いずれのマウスにおいても、酸味 (HCl, citric acid, acetic acid) に対するオフ応答は、WTマウスと比較して有意に減少し た (図8. B~D, 表7)。
図8 WTマウス、PKD1L3-KOマウス、PKD2L1-KOマウス、およびPKD1L3/2L1-DKO マウスにおける舌咽神経酸味オフ応答解析記録例と濃度応答曲線
A. WTマウスとPKD1L3/2L1-DKOマウスにおける舌咽神経オフ応答解析記録例。
点線はベースラインを示す。灰色線は刺激開始時 (図中s: stimuli)と洗浄時(図中w:
wash out)を示す。刺激は60秒間行った。
B-D. WTマウス (n=5-8)、PKD1L3-KOマウス (n=5-6)、PKD2L1-KOマウス (n=5-7)、
PKD1L3/2L1-DKOマウス (n=5-8)におけるHCl (B), citric acid (C), acetic acid (D)に対 する舌咽神経オフ応答の濃度応答曲線。
PKD1L3-KOマウス、PKD2L1-KOマウス、PKD1L3/2L1-DKOマウスのいずれのKO マウスでも、WTマウスと比べ、HCl (B)、citric acid (C)、acetic acid (D)のすべての 酸味オフ応答が有意に減少した (two-way ANOVA; P<0.001: 表7、post hoc Dunnett’s tests; #:P<0.05, ##: P<0.01 for PKD1L3-KO; *: P<0.05, **: P<0.01 for PKD2L1-KO; +: P<0.05, ++: P<0.01 for PKD1L3/2L1-DKO)。データは平均値±S.E.M.で示す。
PKD1L3-KO PKD2L1-KO PKD1L3/2L1-DKO
nerve tastant DF F DF F DF F
GL HCl 1,62 26.9*** 1,67 15.4*** 1,64 27.2***
citric acid 1,72 9.2** 1,76 20.7*** 1,82 21.3***
acetic acid 1,68 27.2*** 1,69 24.7*** 1,77 28.9***
表7 WTマウス、PKD1L3-KOマウス、PKD2L1-KOマウス、およびPKD1L3/2L1-DKO マウスにおける舌咽神経酸味オフ応答解析の分散分析結果
舌咽神経オフ応答におけるtwo-way ANOVA解析での自由度とF値を示した。
PKD1L3-KOマウス、PKD2L1-KOマウス、PKD1L3/2L1-DKOマウスのいずれのKOマ ウスでも、WTマウスと比べ、HCl、citric acid、acetic acidのすべての酸味オフ応答が 有意に減少した (two-way ANOVA; **P<0.01, ***P<0.001)。図8に示したデータを使用 した。
GL: 舌咽神経, DF: 自由度, F: F値
WTマウス、PKD2L1-KOマウスにおける鼓索神経単一神経線維応答解析 舌前方部を支配する鼓索神経全神経束応答解析より、PKD2L1-KOマウスと
PKD1L3/2L1-DKOマウスにおいて、酸味応答の有意な減少が見られた (図5)。
このような減少が起きた理由として、酸味応答神経線維の数の減少、もしくは 酸味応答神経線維における酸味応答の減少が考えられる。そこでWTマウスと
PKD2L1 KOマウスを用いて、鼓索神経単一神経線維応答解析を行った。酸味に
応答する鼓索神経単一神経線維の割合は、WTマウスとPKD2L1-KOマウスで有 意な差がなかった (表8, χ2検定; P>0.1)。しかし、酸味に応答する鼓索神経単 一線維のすべての酸味刺激 (HCl、citric acid、acetic acid) に対する応答は、
PKD2L1-KOマウスでWTマウスと比較して有意に減少していた (図9, 表9)。
WT PKD2L1-KO
CT % CT %
応答数 /記録数
47 / 93
50.5 40 / 84
47.6
表8 WTマウスとPKD2L1-KOマウスにおける鼓索神経単一神経線維酸味応答線維数
酸味 (10mM HCl) に応答する鼓索神経単一神経線維の割合は、WTマウスと PKD2L1-KOマウスで変化が無かった (χ2検定; P>0.1)。
CT: 鼓索神経
図9 WTマウスとPKD2L1-KOマウスにおける鼓索神経単一神経線維の酸味応答記録例 と濃度応答曲線
A. WTマウスとPKD2L1-KOマウスにおける鼓索神経単一神経線維酸味応答記録例。
酸味に応答する鼓索神経単一線維の酸味応答は、PKD2L1-KOマウスでWTマウス より減少していた。
B-D. WTマウス、PKD2L1-KOマウスにおけるHCl (B, WT: n=10~24; PKD2L1-KOマウ ス: n=14~21), citric acid (C, WT: n=12~14; PKD2L1-KOマウス: n=12~16), acetic acid (D, WT: n=12~17; PKD2L1-KOマウス: n=14~21) に対する鼓索神経単一神経線維応 答の濃度応答曲線。
酸味に応答する鼓索神経単一神経線維での解析を行ったところ、PKD2L1-KOマウ スで、WTマウスと比べ、HCl (B)、citric acid (C)、acetic acid (D)のすべての酸味に 対する鼓索神経単一神経線維応答が有意に減少した (two-way ANOVA; P<0.05:
表9、post hoc t-test; *: P<0.05, **: P<0.01)。データは平均値±S.E.M.で示す。
PKD2L1-KO
nerve tastant DF F
CT HCl 1,129 11.9***
citric acid 1,108 19.0***
acetic acid 1,142 6.3*
表9 WTマウスとPKD2L1-KOマウスにおける鼓索神経単一神経線維酸味応答の分散分
析結果
鼓索神経単一神経線維応答におけるtwo-way ANOVA解析での自由度とF値を示した。
PKD2L1-KOマウスで、WTマウスと比べ、HCl、citric acid、acetic acidのすべての鼓索 神経単一神経線維酸味応答が有意に減少した (two-way ANOVA; *P<0.05, ***P<0.001)。
図9に示したデータを使用した。
CT: 鼓索神経DF: 自由度, F: F値
GAD67-GFPマウス、GAD67-GFP+PKD2L1-KOマウスの茸状乳頭GAD67-GFP 発現味細胞における味応答解析
WTマウスにおいて、PKD2L1はGAD67-GFPと共発現する (図3 B.)。舌前方 部の茸状乳頭GAD67-GFP発現味細胞は、酸味に応答することが報告されている (Yoshida et al., 2009)。そこで、PKD2L1が茸状乳頭GAD67-GFP発現味細胞にお いて酸味伝達に関与しているかを解明するため、GAD67-GFPマウス、
GAD67-GFP+PKD2L1-KOマウスの茸状乳頭GAD67-GFP発現味細胞における味
応答解析を行った。酸味に応答する茸状乳頭GAD67-GFP発現味細胞の割合は、
GAD67-GFPマウスとGAD67-GFP+PKD2L1-KOマウスで有意な差が無かった
(表10, χ2検定; P>0.1)。GAD67-GFPマウス、GAD67-GFP+PKD2L1-KOマウス
ともにHCl、citric acid、acetic acidのすべての酸味に対して応答が見られたが、
酸味に応答する茸状乳頭GAD67-GFP発現味細胞の酸味応答は、
GAD67-GFP+PKD2L1-KOマウスでGAD67-GFPマウスと比較して有意に減少し
ていた (図10, 表11)。
GAD67-GFP GAD67-GFP+
PKD2L1-KO
FP % FP %
応答数 /記録数
37 / 112
33.0 26 / 93
28.0
表10 GAD67-GFPマウスとGAD67-GFP+PKD2L1-KOマウスにおける茸状乳頭 GAD67-GFP発現酸味応答細胞数
酸味 (10mM HCl) に応答する茸状乳頭GAD67-GFP発現味細胞の割合は、
GAD67-GFPマウスとGAD67-GFP+PKD2L1-KOマウスで変化が無かった (χ2検定;
P>0.1)。
FP: 茸状乳頭