九州大学学術情報リポジトリ
Kyushu University Institutional Repository
体内時計機構におけるcholecystokinin-1受容体の関 与
山川, 裕介
http://hdl.handle.net/2324/2236173
出版情報:九州大学, 2018, 博士(臨床薬学), 課程博士 バージョン:
権利関係:
博士論文
体内時計機構における
cholecystokinin-1 受容体の関与
2019年 3月
九州大学 大学院薬学府 臨床薬学専攻 臨床育薬学分野
山川 裕介
目次
序論 ... 1
第 1 章 光同調における網膜 CCK-1 受容体の関与
緒言 ... 71. 実験方法 ... 10
1-1. 試薬調製法 ... 10
1-2. 実験動物 ... 10
1-3. 行動リズム解析 ... 12
1-4. Quantitative reverse transcriptase-polymerase chain reaction (qRT-PCR) ... 13
1-5. 免疫組織化学染色法 ... 14
1-6. 硝子体内投与……….. 15
1-7. ルシフェラーゼアッセイ………...16
1-8. 統計解析 ... 16
2. 結果 ... 17
2-1. 網膜CCK-1受容体の日内変動 ... 17
2-2. CCK-1受容体欠損マウスにおける網膜時計遺伝子Per1、Per2の 日内変動 ... 18
2-3. SCNにおける光刺激に対するc-Fos発現 ... 19
2-4. SCNおよび網膜における光刺激に対する時計遺伝子Per1、Per2の発現 ... 20
2-5. 網膜における光刺激に対するc-Fos発現………20
2-6. PER2::LUC導入マウスアストロサイトにおけるCCK受容体刺激および 遮断によるルシフェラーゼ反応………22
3. 考察 ... 26
小括 ... 30
第 2 章 網膜以外の CCK-1 受容体の体内時計への関与
緒言 ... 31
第 1節 Calbindin 陽性線維の伸長への関与 ... 35
1. 実験方法 ... 35
1-1. 試薬調製法 ... 35
1-2. 実験動物 ... 35
1-3. 免疫組織化学染色法 ... 35
1-4. 線維伸長の評価法... 35
1-5. 行動リズム解析 ... 36
1-6. 統計解析 ... 36
2. 結果 ... 37
3. 考察 ... 39
第 2節 ラメルテオン経口投与による再同調促進作用 ... 41
1. 実験方法 ... 41
1-1. 試薬調製法 ... 41
1-2. 実験動物 ... 43
1-3. 行動リズム解析 ... 43
1-4. 再同調促進作用の評価 ... 43
1-5. ウエスタンブロッティング………...44
1-6. 統計解析 ... 44
2. 結果 ... 45
3. 考察 ... 48
小括 ... 52
総括 ... 53
引用文献 ... 55
業績 ... 67
謝辞 ... 69
略語表
ARC arcuate nucleus AVP arginine vasopressin CCK cholecystokinin CT circadian time
DLMO dim light melatonin onset DMH dorsal medial hypothalamus GCL ganglion cell layer
GRP gastrin-releasing peptide IEG immediate early gene
IML intermediolateral cell column INL inner nuclear layer
IPL inner plexiform layer
ipRGC intrinsically photosensitive retinal ganglion cell
KO knockout
RPE retinal pigment epithelium
Oc optic chiasm
OLETF Otsuka Long Evans Tokushima Fatty PaPo posterior paraventricular nucleus PB phosphate buffer
PFA paraformaldehyde RAM ramelteon
RHT retinohypothalamic tract
RT-PCR reverse transcriptase-polymerase chain reaction SCN suprachiasmatic nucleus
S.E. standard error
VIP vasoactive intestinal peptide VMH ventral medial hypothalamus
WT wild type
ZT zeitgeber time 3v third ventricle
1
序論
睡眠・覚醒、体温、ホルモン分泌等の生理現象は体内時計により制御され、
外環境の周期的変化に対応した24時間のリズムが認められる。現代社会では 交代勤務やグローバル化、生活様式の多様化によって不規則な生活が常態化し つつある。それに伴う体内時計リズム失調は、睡眠障害や季節性情動障害を誘 発するほか、高血圧、糖尿病などの生活習慣病、がんなどの疾患のリスクファ クターとなる。健康寿命を延ばし、高いQOLを維持するためにも体内時計機 構を解明することは有益であると考えられる。
体内時計には発振機能があり、地球上のほぼ全ての生物は外環境より時間の 情報が遮断された環境であっても、約24時間を周期とする概日リズムを示 す。哺乳類において、体内時計中枢となる発振体が間脳視床下部に位置する視 交叉上核(SCN)に存在していることが知られている。このことは、SCNを破壊 することにより、行動や体温、ホルモン分泌のリズムが消失すること(Gray et
al., 1978)、培養条件下において、SCNを含む摘出脳切片は他の部位からの神
経連絡や液性因子の影響が遮断されても神経発火リズムは保たれること(Green
et al., 1982)、遺伝的変異により短周期性のリズムを持つハムスターのSCNを
Fig. 1. コア時計遺伝子のフィードバックループ
2
野生型ハムスターに移植すると短周期リズムで行動するようになること(Ralph et al., 1990)などの多くの報告により強く支持されている。この発振機能の本体 は時計遺伝子と呼ばれる特定の遺伝子とその産物がネガティブフィードバック 機構を形成し、概日振動体としての発現を制御することによるコアループであ る(Fig. 1)。コア時計遺伝子のClock、Bmal1の産物CLOCKおよびBMAL1 がヘテロ2量体を形成し、E-box配列へ結合することにより時計遺伝子や時計 被制御遺伝子の転写が活性化される。結果としてPer、Cryの産物PERおよび CRYが増加し、複合体を形成して核内に移行しCLOCK/BMAL1のE-box結 合を抑制することで転写の活性化が抑制されていく。この遺伝子レベルでの振 動の結果、他の遺伝子とその産物の発現にリズムが生じる。中枢発振体のSCN で生じたリズムは神経発火やホルモン分泌を通して全身を網羅し、個体レベル でのリズムとして調律されていく。また、体内時計には光、食事、薬物などの
網膜視床下部路(RHT)
中枢
末梢
ホルモン分泌 神経伝達
神経節細胞 アマクリン細胞 双極細胞 錐体細胞 桿体細胞 色素上皮細胞 基底膜 脈絡膜 水平細胞
網膜
Fig. 2. 体内時計のリズム伝達系
3
外的因子による刺激により外環境に生体リズムの位相を同調させる極めて重要 な機能がある。体内時計のリズム伝達系の概略図をFig. 2に示す。特に強い同 調因子として働くのが光である。哺乳類において、光同調は網膜にて光を受容 し、網膜視床下部路(RHT)を通ってSCNへ光情報を伝達する経路が知られて いる(Liou et al., 1986; de Vries et al., 1994)。哺乳類において、網膜は唯一の 光受容器であり、視細胞層の桿体、錐体および神経節細胞層のメラノプシンに より光が受容される。網膜で受容された光は電気信号に変換されて伝達されて いく。すなわち、RHTを経由してSCNに直接投射する神経の多くはメラノプ シンを発現している内因性光感受性網膜神経節細胞(ipRGCs)からの単シナプス 性の経路である。また、一部は外側膝状体(IGL)を経由してSCNに投射する外 側膝状体視床下部路(GHT)を通った伝達が行われる。これら2つの神経はどち らもSCNの腹外側部に投射していることが報告されている(Shinohara et al.,
1993)。この投射領域にはVIPやGRP、calbindinなどの発現が確認されてお
り、光刺激の入力後、速やかに応答が見られる。一方で、背内側部にはAVP などの発現が見られるが、この領域には概日的発振が見られるものの、光刺激 への直接的な反応性に乏しい。しかし、SCN内の強固な細胞間ネットワークに より全体として安定したリズムを刻んでいる。こうして外環境の光情報から体 内時計中枢の同調が行われ、次いで全身へとリズムが出力されていく。まず室 傍核下部領域へとシグナルを送り、ここから多シナプス性に自律神経核群に出 力されると考えられている。迷走神経背側核に至った時間情報は迷走神経など の副交感神経を通して消化管や呼吸器へ出力され、脊髄中間質外側核(IML)に 至った情報は交感神経系を介して血管や全身の臓器および副腎に出力される (Ishida et al., 2005)。時間シグナルを受けた副腎皮質束状層からはコルチゾー ルの分泌が行われる。コルチゾールが全身の各細胞の受容体に結合し、時計遺
4
伝子Per1の転写を促進することで時間位相を調節し、同調が行われると考え られている。また、IMLからは上頸神経節を通して松果体へと出力される経路 も存在しており、このシグナルは体内時計の調節において重要な物質であるメ ラトニンの合成リズムに関与することも報告されている。
Cholecystokinin (CCK)は中枢神経系で働く重要な神経伝達ペプチドの一 つであり、大脳皮質、視床下部、脳幹などに存在する(Larsson and Rehfeld, 1979; Rehfeld et al., 1980; Crawley and Kiss, 1985)。げっ歯類においてCCK がSCNに分布していることが報告されている(Silver et al., 1999; LeSauter et al., 2002; Morin and Allen, 2006)。加えて、網膜においてもCCKが存在して おり(Marshak et al., 1990)、ラットにおいてアマクリン細胞上に存在するとい う報告がある(Firth et al., 2002)。CCKの受容体にはCCK-1 (CCK-A)、CCK-
2 (CCK-B)の2種類が知られており、いずれも7回膜貫通型のGタンパク共役
型(Gq)受容体である(Wank, 1995)。Otsuka Long Evans Tokushima Fatty (OLETF)ラットは糖尿病などの代謝異常のモデル動物として見出された動物で あるが、自然発生的に機能的CCK-1受容体の欠損がみられる(Takiguchi et al.,
1997)。CCK-1受容体は満腹感を司る神経伝達への関与が報告されており、成
体のOLETFラットでは摂食量の増加および体重の増加が見られる。また、こ
のOLETFラットには光刺激に対する応答の減弱や位相変化させた環境下への
再同調機能の低下等の光同調機能の低下がみられることを報告している
(Funakoshi et al., 1995; Yamanouchi et al., 1997; Shimazoe et al., 1999)。し かしながら、OLETFラットではCCK-1受容体以外にも変異が起こっていると の報告(Nara et al., 1997; Ogino et al., 2000)があるため、CCK-1受容体遺伝 子特異的欠損型マウスを用いて同様の検討をしたところ、やはり光刺激による 行動リズムの位相変化や時計遺伝子の発現促進の減弱、瞳孔反射の低下が認め
5
られたと報告されている(Shimazoe et al., 2008)。この研究において、機能的
なCCK-1受容体の発現を検討し、SCNよりも網膜における発現が多いことも
明らかとなった。すなわち、光同調への関与という点では、中枢であるSCN ではなく、光を受容する網膜での役割が重要であることが示唆されている。し かし、詳細な関与の経路やメカニズムについては未だ明らかになっていない。
そこで本研究では、第1章においてCCK-1受容体の光同調への関与につい て、特に光受容器である網膜に着目して検討を行った。
また、CCK-1受容体は網膜のみならず、視床下部において後部室傍核や背内 側核、腹内側核および弓状核での発現を確認している(Fig. 3, Shimazoe et al., 2008)。前述のとおり、室傍核は出力系にも関与する領域であり、背内側核は摂 食に関与すると考えられている領域である。SCN には CCK の発現も認められ ることから、近傍の領域への出力や摂食などの非光同調への関与も考えられる。
そこで、第 2 章において網膜以外の CCK-1 受容体の関与について、SCN やメ ラトニンシグナルに着目し、検討を行った。
6
Fig. 3. 視床下部におけるCCK-1受容体の発現
Cck1r 遺伝子と lacZ 遺伝子を組み換えたヘテロ型変異マウスの X-gal
染色。a とbは口吻側、cとd は尾側の切片であり、aとcはそれぞれ bとdの背側の切片である。3v:第3脳室、Me:正中隆起、PaPo:後 部室傍核、DMH:背内側核、VMH:腹内側核、ARCr:吻側弓状核、
ARCc:尾側弓状核
100μm
7
第 1 章 光同調における網膜 CCK-1 受容体の関与
緒言
体内時計の自律的なリズムの周期は正確に24時間ではないため、生物は外環 境との時間差を同調機能によって日々補正している。この同調に関わる因子の 中でも光はほぼ全ての生物における最も強い因子である。光により惹起される 生体リズムの位相変化には時刻依存性があることが知られている(Minors et al., 1991)。この変化は生物の昼行性・夜行性を問わず暗期に顕著であり、主観的夜 の早期における刺激により後退し、主観的夜の終期における刺激により前進す る。このような時刻依存的な反応性は、外環境の明暗環境に対する体内時計のず れを効率よく解消する点において有意義な機能である。しかし、この時刻依存性 を作り出している生理的な機序は未だ明らかでない。
先行研究において、CCK-1 受容体が網膜内層のアマクリン細胞上に発現して い る 一 方 で 、SCN に お い て は 発 現 が 乏 し い こ と を 報 告 し て い る(Fig. 4, Shimazoe et al., 2008)。加えて、内因性アゴニストであるCCK-8sを反応させ た実験により、機能的なCCK-1受容体の発現はSCN上には認められないこと、
網膜上の CCK-1 受容体は Ca2+の放出による電気シグナルを生じることを報告
している(Shimazoe et al., 2008)。CCK-1受容体が発現しているアマクリン細胞 は、30種類以上の形態学的な差異を持つ網膜の神経伝達を抑制性に調節してい るニューロンである(Wong et al., 2007; Masland, 2012)。なかでもCCK-1受容 体はグリシン陽性のアマクリン細胞上に発現しているが、CCKの機能的な役割 については未だ明らかでない。網膜における受光からSCNへの光シグナルの伝 達に重要な役割を担うとされているのが ipRGCs である。桿体・錐体の脱落し たマウスは光同調能を維持するが、加えて ipRGCs を欠落させると光刺激への
8
応答を完全に失う(Hatori et al., 2008)。ipRGCsはSCNに直接投射している他、
視蓋前野オリーブ核へも投射しており、瞳孔反射や光忌避による行動抑制とい った非視覚的光応答にも関与することが知られている(Hattar et al., 2006;
Brown et al., 2010)。ipRGCsにはグルタミン酸やGABA、グリシンといった神 経伝達物質の受容体が発現しており、複雑な神経伝達の制御下にある。また、ア マクリン細胞からの刺激を遮断した場合、ipRGCs の神経発火は不安定となり、
光刺激がなくとも神経発火が起きる(Wong et al., 2007)。加えて、CCK-1受容体 欠損マウスでは光に対する瞳孔収縮が減弱しており(Shimazoe et al., 2008)、
CCK-1受容体はipRGCsの機能を維持するのに重要であると考えられる。
本章では、網膜における CCK-1 受容体の発現の日内変動を検討した。また、
CCK-1受容体欠損の影響を検討するため、CCK-1受容体ノックアウトマウスを
用いて時間生物学的環境の指標となる時計遺伝子Per1、Per2の網膜における発 現の日内変動を調べた。また、欠損による刺激の減弱を評価するため、光刺激へ の応答の指標としてノックアウトマウスに対する光照射後の SCN における c-
FosおよびPer1、Per2の発現を測定した。c-Fosの発現の上昇は非特異的な刺
激の評価であり、SCNにおける時計遺伝子の発現の変化は時間生物学的な応答 の評価である。加えて、硝子体内への CCK-1 受容体アンタゴニストである
lorglumide 投与後の光刺激に対する網膜内の c-Fos 発現を測定し、網膜への直
接的な光刺激に対するCCKシグナル遮断の影響を検討した。
9
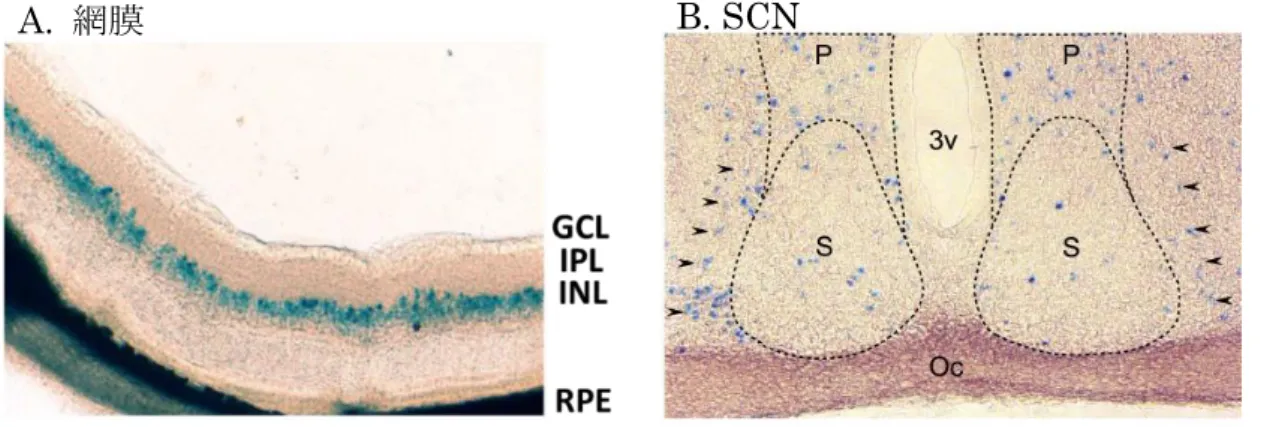
Fig. 4. 網膜およびSCNにおけるCCK-1受容体の発現
Cck1r遺伝子とlacZ遺伝子を組み換えたヘテロ型変異マウスのX-gal染色。
矢印で示すように、SCN よりもその周辺の領域に X-gal 陽性が示された。
GCL:神経節細胞層、IPL:内網状層、INL:内顆粒層、RPE:網膜色素上皮、
3v:第3脳室、S:視交叉上核、P:室傍核、Oc:視神経交叉
A. 網膜 B. SCN
100μm
10
1. 実験方法
1-1. 試薬調製法
・PB (0.1 M, pH7.4)の調製
洗浄液および抗体の希釈溶媒としてPBを用いた。0.2 Mで調製し、使用前に 等量のmili-Q水を加えて0.1 Mとして使用した。0.2 M PBの組成を示す。
NaH2PO4・2H2O (和光) 11.84 g
Na2HPO4・12H2O (和光) 116 g
+H2O 2000 mL
・4% PFA in 0.1 M PBの調製
8 gのPFA粉末(和光)を80 mL程度のmili-Q水に懸濁し、70℃で30 min加 熱した。1 N NaOH水溶液をPFAが溶解するまで加え、mili-Q水を加えて100 mLにメスアップした。使用直前に等量の0.2 M PBを加え、4% PFA in 0.1 M PBとして使用した。
1-2. 実験動物
CCK-1受容体KOマウスは九州がんセンターの瀧口聡一博士より供与を受け、
雌雄の交配による繁殖にて産生し、遺伝子型の判定を行った後に実験に用いた。
このマウスはマウス CCK-1 受容体の exon1、exon2 を neo 耐性遺伝子および lacZ 遺伝子カセットで置換されている。野生型マウスは日本チャールズリバー より雌雄を購入し、KOマウスと同一条件下で繁殖させた。実験には野生型マウ
スおよび CCK-1受容体KOマウス雄性の2-6ヶ月齢のものを用いた。23±2℃
の恒温部屋、自由摂食摂水、明期100 lux、暗期<1 luxの12:12明暗周期条件 下で1~2週間単独で飼育し、周辺環境への同調を完了させた。暗期における作
11
業は赤色光下で行った。本実験は九州大学動物実験規則を遵守して行った。
遺伝子型の判定
3-4週齢のマウスの尾の先端を2-3 mmカットし、1.5 mLチューブに回収し、
下記の組成で調整したBoil Bufferを100 µL加え、55℃で3時間から一晩熱を 加えた。95℃で20 minさらに加熱し、4℃で7000 rpm、5 minの遠心を行い、
上清1 µLをPCRに用いた。
・Boil Bufferの組成
PCR reaction buffer 10× (Boehringer) 5 mL Proteinase K (10 mg/mL, Boehringer) 4 mL Triton X-100 (Sigma-Aldrich) 0.25 mL
H2O
Total 50 mL
・PCR反応液の組成
DNA solution 1 µL
PCR reaction buffer 10× (Boehringer) 2 µL dNTP (10 mM each) 0.5 µL
Primer 1 (20 µM) 0.4 µL
Primer 2 (20 µM) 0.4 µL
H2O 15.7 µL
Taq polymerase (5 U/L, Boehringer) 0.2 µL
Total 20.2 µL
12
・Primer配列
Cck1r -1:5’ – AGTGAGCCATTCACCAGCTCGCCAG – 3’
Cck1r -2:5’ – GTTGCTATATCTCCCTTACCTTGGTAG – 3’
LacZ-1 :5’ – CGCTATTACGCCAGCTGGCGAAAGG – 3’
Wild alleleの検出にはCck1r -1とCck1r -2 (220 bp増幅) Mutant alleleの検出にはCck1r -1とLacZ-1 (250 bp増幅) をそれぞれ用いた。
・サイクリング条件 94℃ 1 min
(94℃ 30 sec, 69℃ 30 sec, 72℃ 1 min) × 35 cycles 72℃ 3 min
(4℃保存)
・電気泳動の条件
2% アガロースゲルを用い、1×TAE buffer中で100 V, 15 min泳動し、エチジ ウムブロマイド染色を行った。Cck1r -/-はWild alleleにおける220 bpバンドの 非検出およびMutant alleleにおける250 bpバンドの検出により同定した。
1-3. 行動リズム解析
自発行動リズムの測定は、20×30×12 cm のケージに一匹ずつ動物を入れ、
真上に設置した放出赤外線検出型の自発運動量センサー(NS-AS01、ニューロサ イエンス)を用いて、1分間隔で行った。データ収録システム(NS-DAS-32、ニュ
13
ーロサイエンス)にてコンピュータ上で運動量を記録し、ClockLab(ニューロサ イエンス)を用いて行動開始時点を算出した。
1-4. Quantitative reverse transcriptase-polymerase chain reaction (qRT-PCR) 頸椎脱臼によりマウスを安楽死させ、速やかに脳および網膜を摘出した。
脳はブレインマトリックス(MBS-S1C、ブレインサイエンス・イデア)を用いて coronalに1 mm厚で薄切り後、解剖用メスにてSCNを含む領域を1 mm3の ブロックで回収した。ホモジナイズを行い、RNeasy Mini Kit (Qiagen)の公式 プロトコルの手順に従い、total RNAの抽出を行った。Total RNAは260/280 nmの吸光度比を1.8-2.0であることにより純度を確認し、Experion (Bio Rad) による電気泳動法にてrRNAのバンド比により品質を確認後、次の工程に進め た。SuperScript® VILO cDNA Synthesis Kit (Invitrogen)を用い、200 ngの
total RNAに対し、公式のプロトコルに従って逆転写を行った。すぐに次の行
程に移らない場合は-20℃で冷凍保存し、使用直前に氷上で融解させた。
Applied Biosystems® 7500リアルタイムPCRを用いてリアルタイムRT-PCR でCck1r及び時計遺伝子Per1/Per2の定量を行った。
プログラムは以下の通りである。
① 95℃ 10 min
② (95℃ 15 sec, 60℃ 1 min)×40サイクル
Ct値は内標準物質としてβ-Actinにて標準化し、ΔΔCt法にて相対定量を行 った。プライマー/プローブセットはApplide Biosystemsより購入した(Table 1)。
14
Table 1 Taqman assays used for RT-PCR
Gene Reporter sequence Assay ID Amplicon length (bp) Cck1r CACCTACTTCATGGGCACTTCCGTG Mm00438060_m1 63
Per1 AGCCCCTGGCTGCCACCATGGCCCC Mm00501813_m1 106 Per2 CCTCCAACATGCAACGAGCCCTCAG Mm00478113_m1 73 β-Actin ACTGAGCTGCGTTTTACACCCTTTC Mm00607939_s1 115
1-5. 免疫組織化学染色法
塩酸メデトミジン(0.3 mg/kg)、ミダゾラム(4 mg/kg)、酒石酸ブトルファノー ル(5 mg/kg)の 3 種混合注射麻酔薬の腹腔内投与にてマウスに麻酔をかけ、4%
PFA (50 mL/匹)にて経心腔的灌流固定を行った後に頭頂部を切開して脳の摘出 を行った。摘出した脳は4% PFAに浸けて一晩置き、次いで20%スクロースへ 浸けて置換した。クライオスタット(Thermo Fisher Scientific)を用いてcoronal に20 µm厚の切片を作成し、24wellプレートに回収した。0.1% Triton X-100 (Sigma-aldrich) in 0.1 M PB (PBT)にて洗浄し、0.5% H2O2による内因性ペル オキシダーゼの除去を行った。1% 正常ヤギ血清(vectastain) in 0.3% PBTにて ブロッキング後、一次抗体としてウサギ由来抗c-Fos IgG(Calbiochem)を暴露さ せ、4℃で 48 時間静置した。二次抗体としてビオチン標識 抗ウサギ IgG
(vectastain)と1時間反応させ、その後アビジンおよびビオチン標識ペルオキシ
ダーゼ(vectastain)と1時間反応させた。Metal Enhanced DAB (Thermo Fisher Scientific)を用いて染色し、スライドガラスに貼り付けた。エタノールおよびキ シレンを用いて脱水し、エンテランニュー(Merck Millipore)を用いて封入した。
15
1-6. 硝子体内投与
LD12:12にて飼育。試験前日のZT12より恒暗環境へ移行し、翌日のZT16
に塩酸メデトミジン(0.3 mg/kg)、ミダゾラム(4 mg/kg)、酒石酸ブトルファノー ル(5 mg/kg)の混合注射麻酔薬の腹腔内投与にてマウスに麻酔をかけ、3 µM Lorglumide or saline 10 µLを硝子体内に投与した。30分後light pulse (70 lux 15 min)を加え、4% PFA (50 mL/匹)にて経心腔的灌流固定を行った後に眼球摘 出を行い、4% PFA に浸けて一晩置き、次いで 20%スクロースへ浸けて置換し た。クライオスタット(Thermo Fisher Scientific)を用いて10 µmの切片を作成 した。0.1% Triton X-100 (Sigma-aldrich) in 0.1 M PB (PBT)にて洗浄し、0.5%
H2O2 による内因性ペルオキシダーゼの除去を行った。1% 正常ヤギ血清 (vectastain) in 0.3% PBTにてブロッキング後、一次抗体として1:2500ウサギ 由来抗c-Fos IgG (Calbiochem)を暴露させ、4℃で48時間静置した。二次抗体 として1:200ビオチン標識抗ウサギIgG (vectastain)と1時間反応させ、その後 アビジンおよびビオチン標識ペルオキシダーゼ(vectastain)と1時間反応させた。
Metal Enhanced DAB (Thermo Fisher Scientific)を用いて染色し、スライドガ ラスに貼り付けた。エタノールおよびキシレンを用いて脱水し、エンテランニュ ー(Merck Millipore)を用いて封入した。切片ごとに200 µm 幅の領域をランダ ムに選択し、領域中の神経節細胞層(GCL)、内顆粒層(INL)に発現するFos陽性 細胞数を計測した。
16
1-7. ルシフェラーゼアッセイ
Per2 遺伝子の下流にルシフェラーゼ遺伝子を導入したマウスから単離し株化 した PER2::LUC アストロサイトを用いた。細胞を 35 mm dish に 1×106 cells/ml、ルシフェリン(100 nM)入りphenol red 不含DMEM培地中に播種 し、24時間後に、CCK受容体アナログであるCCK-8s (300 nM)およびCCK-1 受容体遮断薬であるlorglumide (300 nM)を投与し、1時間ごとにルミサイクル によりルシフェラーゼ活性を測定した。
また、上記同条件でPER2::LUCマウスアストロサイトを播種し、播種した細 胞に対してデキサメタゾン (100 nM)を処置し、その後のルミサイクルにてルシ フェラーゼ活性リズムを 10 分毎に測定した。デキサメタゾン処置後36 時間ま
たは48時間にlorglumideを投与し、リズムの変化を観測した。
1-8. 統計解析
結果は平均値±標準誤差にて表記する。
また、日内変動の解析のため、コサイナー法解析を cosinor program (by Refinetti, http://www.circadian.org/softwar.html)を用いて行い、P < 0.05にて 有意な日内変動があるとした。各値は野生型マウスの ZT2の値を基準(1.0)とし て示した。
光照射による変動は同一遺伝子群において、照射vs非照射の対応のないt検 定にて行い、P < 0.05 にて有意な差があるとみなした。解析には JMP pro 11
(SAS Institute)を用いた。各値は野生型マウスの非照射群の値を基準(1.0)と して相対的に示した。
17
2. 結果
2-1. 網膜CCK-1受容体の日内変動
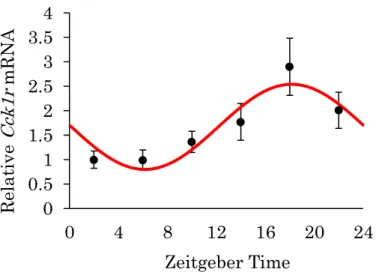
網膜におけるCCK-1受容体の発現の変動を検討するため、12:12明暗条件 下における4時間ごとのCCK-1受容体mRNAの発現量を定量した(Fig. 5)。
Zeitgeber Time(ZT) 0は明期の開始時間、ZT12が暗期開始時間である。
コサイナー法により、WTマウスの網膜においてCCK-1受容体の発現に ZT18付近をピークとする有意な日内変動が認められ(P < 0.01、平均値=
1.67、頂点位相=17.84、 振幅=0.87)、ピーク時には約3倍まで発現の増加を
示した。すなわち、明期に発現が低下し、暗期に発現が増加するという日内変 動が存在することが示された。
Fig. 5. Daily rhythm of Cck1r mRNA expression in the mouse retina.
The expression levels of Cck1r mRNA every 4 hours under the LD cycle in WT mouse retina were assessed by PCR. The data represent the mean ± S.E. (n = 6). The value are calculated relative to the mean of ZT 2. The expression show significant daily rhythm on cosinor analysis (P < 0.01).
0 0.5 1 1.5 2 2.5 3 3.5 4
0 4 8 12 16 20 24
Relative Cck1r mRNA
Zeitgeber Time
18
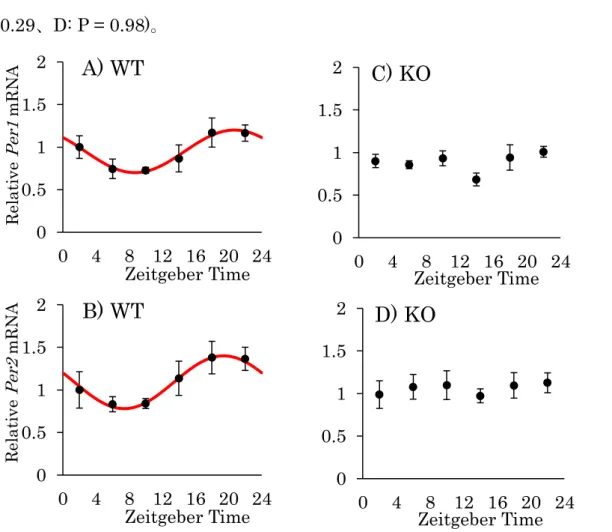
2-2. CCK-1受容体欠損マウスにおける網膜時計遺伝子Per1、Per2の日内変動
CCK-1受容体の欠損による時間生物学的な影響を調べるべく、WTおよびKO
マウスの網膜における時計遺伝子Per1、Per2の発現の日内変動を比較検討した
(Fig. 6)。WTマウスにおいて、Per1、Per2ともに有意な日内変動を示し、夜間
の発現が上昇していたのに対し、KOマウスにおいてはどちらも統計学的に有意 な日内変動を示さなかった (A: 平均値=0.95、頂点位相=20.70、振幅=0.25、
P < 0.01、B: 平均値=1.09、頂点位相=19.40、振幅=0.31、P < 0.01、C: P = 0.29、D: P = 0.98)。
Fig. 6. Daily rhythm of Per1 and Per2 mRNA expression in the mouse retina.
The expression levels of Per1 (A: WT, C: KO) and Per2 (B: WT, D: KO) mRNA every 4 hours under the LD cycle in mouse retina were assessed by PCR. The data represent the mean ± S.E. (n = 6). The value are calculated relative to the mean of ZT 2. The genes expression levels of WT mice show daily rhythm on cosinor analysis (P < 0.01).
In contrast, mRNA levels of KO mice represent no significant variation.
0 0.5 1 1.5 2
0 4 8 12 16 20 24 Zeitgeber Time A) WT
0 0.5 1 1.5 2
0 4 8 12 16 20 24 Zeitgeber Time C) KO
0 0.5 1 1.5 2
0 4 8 12 16 20 24 Zeitgeber Time B) WT
0 0.5 1 1.5 2
0 4 8 12 16 20 24 Zeitgeber Time D) KO
Relative Per1 mRNARelative Per2 mRNA
19
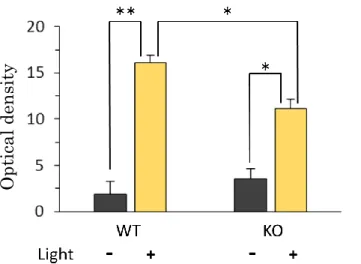
2-3. SCNにおける光刺激に対するc-Fos発現
光刺激への分子生物学的な応答に CCK-1 受容体欠損の影響が見られるかを SCNにおける c-Fosの発現にて検討した。光照射は70 luxの照度で15分間行 い、照射開始後1時間でサンプリングを開始した。c-Fosは免疫組織化学染色法 にて染色し、SCN領域(Fig. 7A)の平均グレースケールから非発現領域(Fig. 7B) のグレースケールを引いたOptical Densityを計測した。
ImageJを用いて計測したoptical densityを野生型およびCCK-1受容体KO マウスとで比較した(Fig. 8)。両群とも光照射により有意な発現の上昇が認めら れた。しかし、その上昇量はKOマウス群で有意に抑制された。
Fig. 7. WTマウスにおけるc-Fos免疫染色像 ABC-DAB法を用いたc-Fos免疫染色像。
A: SCN領域、B: c-Fos非発現領域
A B
20
2-4. SCNおよび網膜における光刺激に対する時計遺伝子Per1、Per2の発現
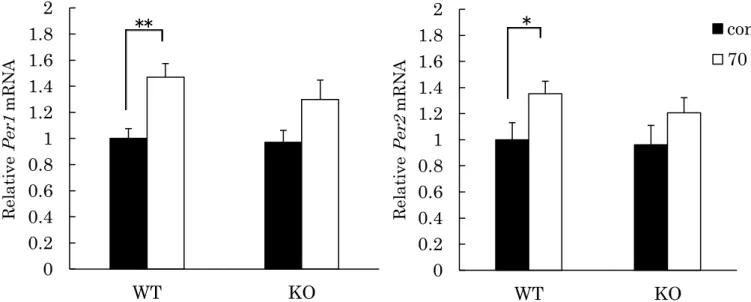
a)SCNにおける光刺激による時計遺伝子Per1、Per2の誘導
光刺激による時間生物学的な応答にCCK-1受容体の欠損が影響するかを検討 するため、SCN領域における光照射後の時計遺伝子Per1、Per2 mRNAの発現 量をリアルタイム PCRにて測定した。光照射は 70 luxの照度で15 分間行い、
照射開始後 1 時間で臓器の摘出を開始した。例数は各群 8 匹ずつで行った。結
果をFig. 9に示す。Per1、Per2共にWTマウスでは有意な発現量の上昇が見ら
れた(Per1; 1.00±0.07 vs 1.50±0.09, P < 0.01, Per2; 1.00±0.11 vs 1.35±0.08, P < 0.05)。一方、KOマウスではPer1、Per2共に発現量が上昇する傾向が見ら れたものの、有意な差ではなかった(Per1; 0.93±0.08 vs 1.22±0.13, P = 0.073,
Fig. 8. Light-induced c-Fos expression in the mouse SCN.
Fos expression was assessed by immunohistocshemically. The mice were exposed to 70 lux white lights for 15 min at circadian time of 16. Values are shown as the means ± S.E. (n = 5-6). There were significant differences of optical density in the SCN between WT and CCK-1R KO mice by Tukey- Kramer HSD test (*P < 0.05, **P < 0.01).
Optical density
21
Per2; 0.96±0.13 vs 1.21±0.10, P = 0.154)。
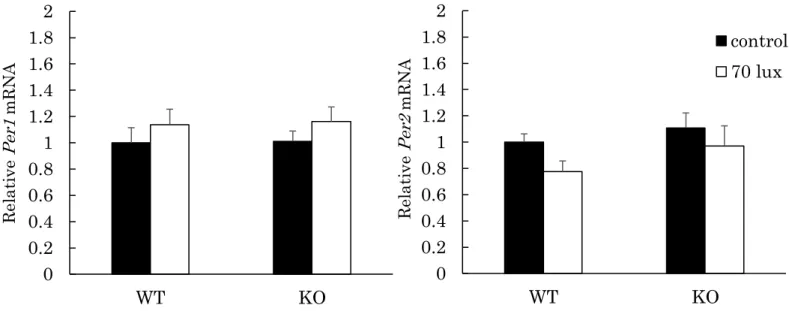
b)網膜における光刺激による時計遺伝子Per1、Per2の誘導
光刺激による網膜の時間生物学的な応答にCCK-1受容体の欠損が影響するか を検討するため、光照射後の時計遺伝子Per1、Per2 mRNAの発現量をリアル タイムPCRにて測定した。例数は各群6匹ずつで行った。結果をFig. 10に示 す。Per1、Per2共にWTマウス、KOマウスのいずれにおいても有意な発現量 の変化はなかった。WT (Per1; 1.00±0.11 vs 1.14±0.12, P = 0.19, Per2; 1.00±
0.06 vs 0.78±0.08, P > 0.5)、KO (Per1; 1.01±0.08 vs 1.16±0.11, P = 0.29, Per2; 1.11±0.11 vs 0.97±0.15, P = 0.48)。
Fig. 9. Light-induced Per1, Per2 mRNA expression in the mouse SCN.
The expression levels of Per1 and Per2 mRNA in mouse SCN were assessed by PCR. The mice were exposed to 70 lux lights for 15 min at Zeitgeber time of 16. The data represent the mean ± S.E. (n = 8). The values are calculated relative to the mean of WT control. *P < 0.05, **P < 0.01
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 2
WT KO
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 2
WT KO
control 70 lux
** *
Relative Per2 mRNA
Relative Per1 mRNA
22
2-5. 網膜における光刺激に対するc-Fos発現
CCK-1 受容体拮抗薬 lorgrumide 投与下で、光刺激に対する応答がどのよう
に変化するかを検討した。網膜においてc-Fos陽性細胞数を内顆粒層(INL)およ び神経節細胞層(GCL)それぞれで調べた(Fig. 11A-C)。
GCL において、saline 群および lorglumide 投与群のどちらにおいても光照 射による有意な c-Fos 陽性細胞数の増加が認められた(Saline; 2.15±0.20 vs 6.53±0.05, Lorglumide; 2.31±0.20 vs 4.85±0.13, P < 0.01)。 し か し 、
lorglumide投与群における陽性細胞の増加はsaline群に比べ有意に抑制された
(P < 0.05、Fig. 11B)。一方、内顆粒層においてはどちらの群においても光照射 による陽性細胞数の増加は乏しく、有意な差ではなかった(Fig. 11C)。
Fig. 10. Light-induced Per1, Per2 mRNA expression in the mouse retina.
The expression levels of Per1 and Per2 mRNA in mouse retina were assessed by PCR. The mice were exposed to 70 lux lights for 15 min at Zeitgeber time of 16. The data represent the mean ± S.E. (n = 6). The values are calculated relative to the mean of WT control.
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 2
WT KO
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 2
WT KO
control 70 lux
Relative Per1 mRNA Relative Per2 mRNA
23
Fig. 11 Light-induced c-fos positive cells expression in the mouse retina.
Mean ± S.E. (n = 6) *P < 0.05, **P < 0.01, Tukey-Kramer’s HSD test. Sal: Saline, Lor:
Lorglumide
0 1 2 3 4 5 6 7
Sal Lor
C) INL Control
Light pulse
0 1 2 3 4 5 6 7
Sal Lor
c-Fos positive cells
B) GCL
** **
*
200 µm
GCL INL
A) 光刺激後の網膜における c-Fos陽性細胞
c-Fos陽性細胞を矢印で示す。
24
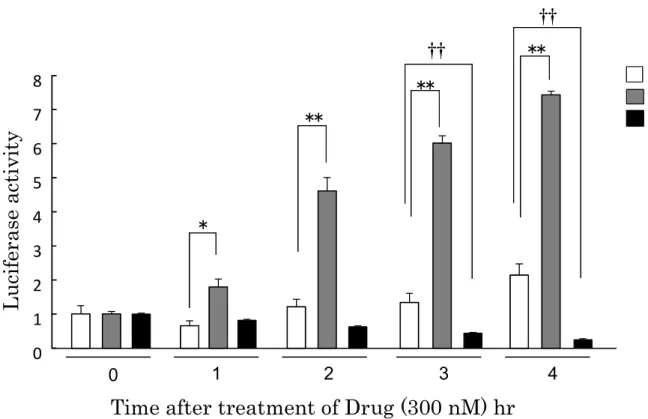
2-6. PER2::LUC 導入マウスアストロサイトにおける CCK 受容体刺激および遮
断によるルシフェラーゼ反応
PER2::LUC導入マウスのアストロサイトに対し、CCK受容体アナログであ
るCCK-8sおよびCCK-1受容体遮断薬であるlorglumide を投与し、経時的 にルシフェラーゼ発光を測定した(Fig. 12)。CCK-8sを投与した群では経時的 なルシフェラーゼ活性の上昇が検出された。一方、lorglumide群においては経 時的なルシフェラーゼ活性の低下が検出され、反応後3、4時間においては DMSO群に比べて有意な活性の低下が認められた(P < 0.01)。
0 1 2 3 4 5 6 7 8
Luciferase activity
DMSO CCK-8 Lorglumide
0 1 2 3 4
Time after treatment of Drug (300 nM) hr
*
**
††
††
**
**
Fig. 12 Luciferase activity by CCK receptor stimulation or blockade in PER2::LUC mouse astrocytes.
Mean ± S.E. (n = 4-6) *P < 0.05, **P < 0.01, †P < 0.05, ††P < 0.01, Tukey- Kramer’s HSD test.
25
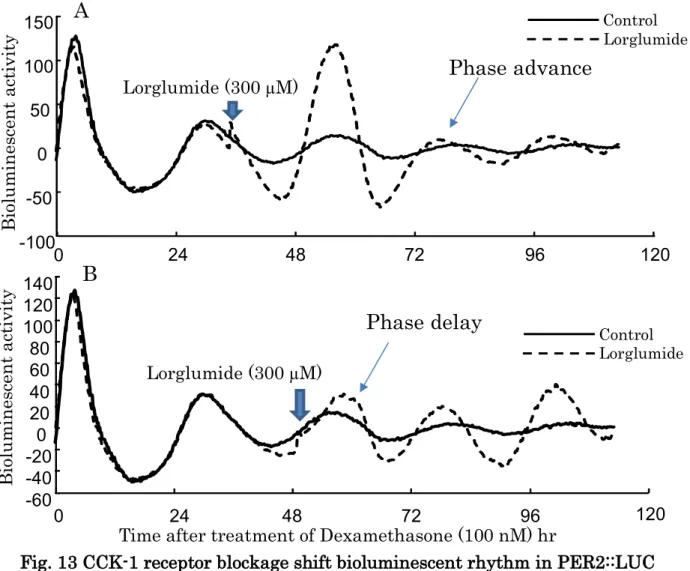
また、PER2::LUC導入マウスのアストロサイトに対し、デキサメタゾン
100 nM投与後の生物発光を経時的に測定した(Fig. 13)。36時間後に
lorglumideを投与すると生物発光活性のリズムが変位し、位相の前進が見られ
た(Fig. 13A)。一方、48時間後にlorgrumideを投与した場合は位相の後退が 見られた(Fig. 13B)。
-100 -50
0 50 100 150
0 24 48 72 96 120
Bioluminescent activity
Phase advance Lorglumide (300 µM)
A
-60 -40 -20 0 20 40 60 80 100 120 140
0 24 48 72 96 120
Lorglumide (300 µM)
Time after treatment of Dexamethasone (100 nM) hr
Bioluminescent activity
Phase delay B
Fig. 13 CCK-1 receptor blockage shift bioluminescent rhythm in PER2::LUC mouse astrocytes.
Lorglumide administered 36 hours later, the rhythm of the bioluminescence activity shifted and phase advancement was observed (Fig. 13A). On the other hand, when lorgrumide was administered 48 hours later, phase delay was observed (Fig. 13B).
Control Lorglumide
Control Lorglumide
26
3. 考察
本章では、光同調におけるCCK-1受容体の関与を明らかにするために、網
膜CCK-1受容体の日内変動、CCK-1受容体欠損マウスにおける網膜時計遺伝
子Per1、Per2の日内変動、SCNにおける光刺激に対するc-Fos発現および時
計遺伝子Per1、Per2の発現、網膜における光刺激に対する時計遺伝子Per1、
Per2の発現について検討した。
その結果、WTマウスにおいて、網膜のCCK-1受容体の発現には日内変動 が存在することが明らかとなった。一方、CCK-1受容体欠損マウスでは、網膜 上の時計遺伝子の発現にリズムが認められなくなっており、網膜における時間 生物学的調節にCCK-1受容体の発現が関与している可能性が示された。
今回の実験ではCCK-1受容体は暗期にその発現が上昇していた。当研究室 において、CCK-1受容体は網膜上ではグリシン陽性アマクリン細胞に限局して 発現していることを報告している(Shimazoe et al., 2008)。アマクリン細胞の 主な働きは双極細胞よりグルタミン酸系入力を受け、神経節細胞へと抑制系の 神経伝達を行うことである(Jacoby et al., 2015)。網膜を構成する細胞の中でも 最も形態学的に多様な細胞であり、GABAやグリシン、ドパミンといった様々 な伝達物質により抑制系の制御をすることが知られている(Vuong et al.,
2015)。アマクリン細胞からのシグナルを遮断したipRGCsは光刺激を与えな
い状況でも神経発火を示すなど、アマクリン細胞は網膜におけるシグナル伝達 の安定化に重要な役割を示すことが報告されている(Wong et al., 2007)。先行 研究により、マウスの網膜アマクリン細胞はCCK-8sにより神経発火が引き起 こされることが分かっており(Shimazoe et al., 2008)、生体内でも受容体発現 の変動によるCCKシグナルのリズムが存在する可能性がある。このシグナル 強度の変動が時間生物学的にどのような役割をしているかについてはさらなる
27
検討が必要であるが、暗期のCCK-1受容体の発現増加によって抑制系伝達を 強め、ipRGCsの自発的神経発火を抑えるという働きを持つとも考えられる。
また、CCK-1受容体KOマウスは網膜の時計遺伝子Per1/Per2のmRNAの 発現に日内変動が認められなくなっていた。瞳孔反射の光感受性には日内変動 が存在し、さらに時計遺伝子を変異させたマウスでは、光感受性が低下すると いう報告がある(Owens et al., 2012)。本研究において、時計遺伝子の発現が完 全に消失したわけではなく、リズムの消失も網膜でのみの確認であったが、
我々が以前報告した(Shimazoe et al., 2008)ようにCCK-1受容体KOマウスに おいて瞳孔反射の光応答が減弱したのはこの網膜の概日リズムが弱まったため ではないかと考えられる。
次に、SCNにおけるc-Fos発現について検討した。c-Fosは細胞への刺激に 応答して鋭敏に発現が上昇することから神経刺激のマーカーとして広く用いら れており、当研究室では、CCK-1受容体が欠損しているOLETFラットでは、
正常ラットに比べて光誘発c-Fos発現が有意に低下することを報告している (shimazoe et al., 1999)。今回、野生型およびKOマウスで光照射によりc-Fos の発現が認められたが、KOマウス群の発現は野生型と比べると有意に抑制さ れていた。網膜におけるCCK-1受容体の発現部位は内層のアマクリン細胞上 であり、光伝達のより下層に光受容能を持つipRGCsが存在する。網膜内の光 伝達において、桿体や錐体で受容された光刺激は双極細胞に伝達され、その後 神経節細胞へと伝達されてRHTを通ってSCNに至る。アマクリン細胞は双極 細胞および神経節細胞を抑制的に制御することで神経発火を調節する役割を担 っている。CCK-1受容体の欠損により、アマクリン細胞が関与する経路の伝達 が阻害され、SCNへの刺激の伝達が減弱したと考えられ、光刺激伝達への
CCK-1受容体の関与の可能性が示された。
28
ただし、前述の通り下流にipRGCsが存在するため、アマクリン細胞を介さな い直接的なipRGCsへの刺激が伝達し、KOマウスにおいても刺激がSCNに 届き、完全な遮断とはならなかったと考えられる。
WTマウスでは、光刺激によりSCNにおいて時計遺伝子であるPer1および Per2の有意な発現増加が認められた。しかし、CCK-1受容体欠損マウスでは 光刺激によって、有意な時計遺伝子の増加は認められなかった。この結果は、
Shimazoe et al (2008)の報告と一致している。一方、網膜においては、WTお
よびCCK-1受容体欠損マウスのいずれも、光刺激によって時計遺伝子発現の
増加は認められなかった。光刺激を行うことにより、網膜の時計遺伝子の発現 にも影響を及ぼすという報告がある(Namihira et al., 2001; Lahouaoui et al., 2014)。種差や照射光などの条件の違いもあるが、これらの報告は大きく発現 量が増加するものではなく、差が有意ではないものもある。今回我々の結果も 若干ではあるがPer1の増加がある。網膜への光刺激による影響はKOでも変 わらなかった。げっ歯類において網膜でのPer1、Per2の発現は層特異的であ ることが報告されており、mRNA発現は内顆粒層で促進される。内顆粒層に限 った発現の検討など、より詳細な条件では影響が見られる可能性もある。
硝子体内にCCK-1受容体アンタゴニストであるlorgulmideを投与し、光刺 激を加えた場合の網膜内での応答を検討した。内顆粒層におけるc-Fosの発現
はVEH群およびlorglumide群どちらにおいても乏しく、照射の有無による有
意な差は認められなかった。一方、神経節細胞層における発現はより詳細に発 現部位を調べた研究では、ドパミン作動性アマクリン細胞への時計遺伝子の発 現が報告されている(Dorenbos et al., 2007; Pozdeyev et al., 2008)。アマクリ ン細胞は形態学的に多様なニューロンであり、双極細胞や神経節細胞のみなら ず、アマクリン細胞同士でも抑制系の制御をすることが知られている。ドパミ
29
ン作動性ニューロンにもグルタミン酸やGABA、グリシンの受容体が存在して いることが報告されている(Newkirk et al., 2013)。グリシン作動性アマクリン 細胞を介する経路の刺激は桿体からのon型双極細胞からの信号であり、主に 弱い光により生じる。今回の直接的な光刺激による伝達でCCK-1受容体欠損 の影響が出なかったのは、錐体からの信号によりグリシン作動性アマクリン細 胞を介さない経路が存在するためである可能性がある。一方、KOマウスでは
CCK-1受容体の発現の変動が及ぼすCCKシグナルの日内変動が失われること
により、ドパミン作動性ニューロンの時計遺伝子の発現リズムが乱れた可能性 がある。今後、アマクリン細胞間でのより詳細なネットワークを検討すること で、網膜上の概日リズム維持のメカニズムを明らかにすることが期待される。
また、PER2::LUC導入マウスのアストロサイトを用いて、CCK-1受容体シ グナルがPER2発現に及ぼす影響を検討した。CCK-1受容体アナログである
CCK-8sを投与すると投与後4時間まで経時的にルシフェラーゼ活性が上昇し
ており、PER2発現を促す因子であるということが示唆された。これは結果2-
1、2-2のCck1rおよびPer2の日内変動で得られた結果とも合致しており、
CCK-1受容体の発現が増加して刺激が増えるのに続いてPer2の発現が増加す
るという関係が生体内でも存在していることを支持するものである。一方で、
CCK-1受容体遮断薬であるlorglumideを投与するとルシフェラーゼ活性が抑
制されたり、生物発光のリズムの位相が投与時間依存的に変位することが示さ れ、時間生物学的な環境への関与が示唆される。
以上の結果より、網膜のCCK-1受容体は網膜概日リズムの維持に重要な役 割を担っていることが示唆された。
30
小括
本章では、光同調へのCCK-1受容体の関与について特に網膜上に発現した ものに着目し、検討を行った。その結果、以下の点が明らかになった。
1)網膜CCK-1受容体は発現に日内リズムが存在し、明期に発現が低下し、暗
期に発現が増加する。
2)CCK-1受容体KOマウスでは網膜における時計遺伝子Per1、Per2の発現リ
ズムが消失することから、CCK-1受容体は網膜概日リズムの維持に重要であ る。
3)CCK-1受容体KOマウスにおける、光刺激に対するSCN内の時計遺伝子
Per1、Per2、およびc-Fosの発現は減少しており、CCK-1受容体は光刺激
伝達に関与している。
4) PER2::LUC導入マウスのアストロサイトを用いた検討において、CCK-8s
の投与はルシフェラーゼ活性の上昇を示し、Lorglumideの投与はルシフェ ラーゼ活性を抑制したことから、CCK-1受容体シグナルがPER2発現に関 与している。
以上より、CCK-1受容体は時計遺伝子PER2発現に関与し、網膜の時間生 物学的な環境の維持に重要な役割を担っていることが示唆され、欠損により光 同調能が減弱する可能性が示唆された。
31
第 2 章 網膜外の CCK-1 受容体の体内時計機構への関与
緒言
CCKは腸管で働くホルモンとして発見された(Ivy and Oldberg, 1928)。CCK- 1 受容体は末梢では腸管や膵臓などに発現しており、時計遺伝子 Clock 変異マ ウスでは膵 CCK-1 受容体の発現が低下するという報告もある(Oishi et al.,
2006)。CCKは中枢では大脳皮質や視床下部、脳幹などに存在しており、代謝制
御や記憶、不安などの情動にも関与するという報告がなされている(Hökfelt et al., 2002; Lo et al., 2008)。このようにCCKは末梢・中枢どちらにおいても生 理機能を調節する重要なペプチドであり、網膜以外のCCK-1受容体も体内時計 機構へ関与している可能性がある。
Ca2+結合タンパク質の一つである Calbindin (CalB)は、主に腎臓や中枢神経 に存在しており Ca2+の輸送、濃度調節、センサーとして働いている。CalB は SCN 内にも存在しており、欠損により SCN の時計遺伝子の日内変動や光刺激 による最初期遺伝子fosの発現に影響が出るなど、体内時計機構に関与すること が報告されている(Kriegsfeld et al., 2008)。ハムスターSCNのCalB陽性細胞 において、核内と細胞質のCalB存在比に日内変動が存在していることが報告さ れており(Hamada et al., 2003)、CalBは主観的夜の初期においてコア領域から 線維に沿って SCN 内の他の領域へと広がっていく。CalB 陽性細胞は SCN の コア領域への発現が多いが(Fig. 14)、背内側部への発現は乏しい。CalBは線維 に沿った移動により、背内側部の VP 細胞との共発現にも日内変動を示すこと が報告されており(LeSauter et al., 2009)、SCN内の細胞間ネットワークを構築 する重要な役割を持つと考えられる。
32
一方、概日リズムの周辺環境への同調には、光刺激を伝達する網膜視床下部路 を介した光同調および温度や食餌、運動による刺激を介した非光同調があるこ とが知られている(Tahara et al., 2017)。非光同調の強力な因子のひとつとして メラトニンの存在が挙げられる。メラトニンは生体内でトリプトファンから合 成され、脳の松果体から主として分泌されるホルモンである。血清中のメラトニ ン値は幅広い生物種で昼に低値、夜間に高値となる概日リズムを示すことが知 られており(Reiter et al., 1993, Falcón et al., 2010)、睡眠・覚醒や自発運動、体 温変化などの多くの生理現象の制御に関わっていることが明らかになっている (Pévet et al., 2006)。また、メラトニンはSCNにおいてcalmodulin-dependent protein kinase II (CaMKII)による神経の長期増強を阻害することも報告されて おり(Fukunaga et al., 2002)、これにより非光同調を生じさせていると考えられ
Fig. 14. ハムスターSCNにおけるCalB陽性細胞および陽性線維 DABを用いたハムスターSCNを含むSaggital切片のCalB免疫染色 像。SCNのコア領域に集積が見られる。
Silver Rより提供。
33
る。メラトニン受容体は MT1、MT2、MT3に分類され、それぞれの受容体が全 身の様々な臓器に発現し、異なった役割を担っている(Fig. 15)。中でも睡眠に関 与するMT1やSCNにおける神経発火リズムの位相を変化させるMT2受容体が 体内時計の調節に関わっているとされている。
Ramelteon(RAM)はメラトニンMT1、MT2受容 体の特異的アゴニストであり、不眠症における入眠 困難の改善に用いられている。臨床において RAMは 就寝前に服用されているが、RAMにも体内時計の位 相調節作用があり、かつ服用時点によってその効果
が違いを示すことが分かっている(Oliver et al., 2011)。これは、体内時計の位相 調節作用を持つ SCN 中のメラトニン受容体の発現に日内変動があることに起
Fig. 16. RAMの構造
Fig. 15. メラトニンとその受容体が関わる生理的機能
CNS:中枢神経系、PGE:プロスタグランジン、KLH:キ ーホールリンペットヘモシアニン。
Dubocovich and Markowska. 2005より一部改変。
34
因すると考えられている(Amanda et al., 2001)。メラトニンシグナルとCCK-1 受容体の関与は未だ明らかではない。
そこで本章においては、網膜以外に発現するCCK-1受容体の体内時計機構へ の関与を評価するため、第1節にてSCNにおけるCalB陽性線維の伸長にCCK- 1受容体の欠損が影響するか否かについて検討した。
また、第 2 節において RAM の投与時点を新規明暗環境下の暗記開始時点に 定め、再同調機能に与える影響の評価を通じ、メラトニンシグナルとCCK-1受 容体シグナルとの関係を再同調促進作用の観点から検討を行った。
35
第1節 Calbindin 陽性線維の伸長への関与
1. 実験方法
1-1. 試薬調製法
第一章に準じる方法にてPB (0.1 M, pH 7.4)および4% PFA in 0.1 M PBを調 整した。
1-2. 実験動物
第 1 章 1-2. 実験動物に準ずる方法にて飼育・繁殖させたマウスを用いた。実
験には2-6か月齢の雄性マウスを用いた。各実験前に12 hr:12 hrの明暗環境下 で1-2週間単独飼育を行い、行動リズム解析にて同調を確認後試験に移った。本 実験は九州大学動物実験規則を遵守して行った。
1-3. 免疫組織化学染色法
第一章に準じる方法にてマウス脳切片標本を作製した。なお、一次抗体として ウサギ由来抗Calbindin-D28k IgG (CHEMICON)を用いた。
1-4. 線維伸長の評価法
蛍光顕微鏡(BZ9000、KEYENCE)を用いて明視野で SCN 領域を撮影し、50 µm グリッド線を染色像の上に重ね、免疫染色陽性線維との交差数を計測した。
各動物につき10枚の切片について計測を行い、その合計数を伸長の指標として 評価した。
36
1-5. 行動リズム解析
第一章に準じる方法にて行動量を測定し、明暗環境への行動リズムの同調の 確認を行った。
1-6. 統計解析
結果は平均値±標準誤差にて表記する。
また、日内変動の解析のため、コサイナー法解析を cosinor program (by Refinetti, http://www.circadian.org/softwar.html)を用いて行い、P < 0.05にて 有意な日内変動があるとした。
37
2. 結果
視交叉上核におけるCalB陽性線維の伸長の日内変動を検討するため、4時間 毎の6点でサンプリングを行った。ZT2とZT14の染色像の代表例をFig. 17に 示す。
CalB 陽性線維伸長の指標として、染色切片を撮影した画像に 50 µm の格子 を重ねた時の線維との交差数を用いた。各個体につき10枚ずつの切片を作成し、
その交差数の合計をWTとKOとで比較検討した。結果をFig. 18に示す。WT マウスにおいて、CalB陽性線維とグリッド線との交差数に有意な日内変動が見
Fig. 17. SCN 領域のCalB染色像
ABC-DAB 法を用いた SCN 領域の CalB 染色像。青矢印にて
CalB陽性線維の例を示す。
38
られ(平均値=283.5、頂点位相=12.41、振幅=59.89、P < 0.001)、暗期初期に 陽性線維の伸長にピークが見られた。一方、KOマウスでは有意な日内変動は認 められなかった(P = 0.38)。
Fig. 18. The extension of the CalB immunoreactive fibers in mouse SCN.
CalB positive fibers were determined by immunohistochemically. The extension of CalB-ir fibers was assessed by number of crossing to 50 µm grid. Data represent the means ± S.E. (n = 4). There were significant daily rhythm in the WT mice SCN (P < 0.001, by cosinor analysis), but there were not shown rhythmically variation in the CCK-1R KO mice SCN (P = 0.67).
0 50 100 150 200 250 300 350 400
2 6 10 14 18 22
Fiber Crossing
ZeitgeberTime
A) WT
0 50 100 150 200 250 300 350 400
2 6 10 14 18 22
ZeitgeberTime
B) KO
39
3. 考察
CalBは、カルシウム結合タンパク質であり、SCNの腹外側部に存在している ことが知られている(Silver et al., 1996)。SCNにおいて、このCalBが存在し ている部位のみを破壊することで行動などの日周リズムが消失することが知ら れており、体内時計機構に重要な役割を果たしていることが明らかになってい る(Kriegsfeld et al., 2008; Butler et al., 2011)。そこで、WTマウスおよびCCK- 1 受容体欠損マウスのSCNにおける線維上のCalB発現の日内変動について検 討した。その結果、WTマウスにおいては、ZT12付近をピークとする日内リズ ムが観察されたのに対し、CCK-1受容体欠損マウスではリズムが消失していた。
CCK-1受容体欠損マウスは、行動リズムなどの表現系には大きな違いはないが、
光刺激に対する同調機能が低下していることが明らかになっている(Shimazoe et al., 2008)。
CalB 陽性細胞数はほとんど日内の変動を示さないが、CalB の核内と細胞質 の存在比に変動が見られる(Hamada et al., 2003; Cayetanot et al., 2007)ことか ら、腹外側部から背内側部へ神経線維を移動することにより伝達を行なってい ると推測されている。SCN におけるCalB の分子生物学的な役割は未だ不明な 点も多いが、神経細胞におけるCalBは細胞内Ca2+濃度の調節やCa2+の輸送と いった機能をもっている(German et al., 1997)。また、高いCa2+親和性により、
Ca2+の緩衝剤として働き、生体環境を安定化させていると考えられている。SCN における刺激の伝達および発振には Ca2+の存在が不可欠であることが報告され ており(van den Pol et al., 1992)、CalBの線維内移動もSCNの機能の維持に重 要であると考えられる。したがって、この発現リズムの消失が、CCK-1 受容体 欠損マウスの同調機能低下に関わっている可能性がある。
また、縫線核においてCCKはセロトニン神経系と共存している(Boden et al.,
40
1991)。セロトニン神経系は縫線核から直接 SCN に投射しており、神経活動を
抑制的に調節している(Boulaich et al., 1994; Glass et al., 2000)。自然発症的に
CCK-1 受容体を欠損しているOLETFラットにおいて、セロトニンの機能亢進
が起こっており、セロトニン受容体を遮断することで光刺激への応答が回復す ることを報告しており(Shimazoe et al., 2004)、CCK-1受容体の欠損により、セ ロトニン神経系との相互作用が失われた影響が表れている可能性も否定できな い。いずれにしても、このリズムの消失が具体的にどのようなメカニズムに起因 するものかを明らかにするためには、より詳細な相互作用の機序解明が必要で ある。
本研究により、CCK-1 受容体KO マウスでは SCN 内の線維上の CalB発現 の増減に日内変動が認められなくなっており、SCN内の細胞間ネットワークに 影響が出ている可能性が示唆された。
41
第2節 ラメルテオン経口投与による再同調促進作用
1. 実験方法
1-1. 試薬調製法
経口投与の溶媒として0.5% CMC水溶液を用いた。
Carboxy methyl cellulose 50 mg
H2O 10 mL
Total 10 mL
Ramelteon (AdooQ BioScience) 1 mgをCMC水溶液1 mLに懸濁し、均一にし た後に-20℃で保存し、使用直前にCMC水溶液で希釈して用いた。
Ramelteon (1 mg/mL) 10 µL
CMC水溶液 990 µL
Total 1000 µL
ウエスタンブロッティングに以下の試薬を調製して用いた。
6x SDS Sample buffer
4x Tris-HCl-SDS (pH 6.8) 7 mL
Glycerol 3.6 mL
SDS 1.2 g
β-mercaptoethanol 0.6 mL
Bromophenol Blue 適量
42
Running buffer
Glycine 14.4 g
Tris 3.0 g
SDS 1.0 g
H2O 1000 mL
Total 1000 mL
Membrane transfer buffer
Glycine 14.4 g
Tris 3.0 g
SDS 0.2 g
Methanol 200 mL
H2O 800 mL
Total 1000 mL
PBS-T
10x PBS 100 mL
25% Tween 20 4 mL
H2O 896 mL
Total 1000 mL
Blocking buffer
PBS-T 40 mL
BSA 2 g
43
1-2. 実験動物
第1章 1-2. 実験動物に準ずる方法にて飼育・繁殖させた C57BL/6Jマウスお
よびCCK-1受容体欠損マウスを用いた。実験には2-3ヶ月齢の雄性マウスを用
いた。各実験前に12 hr:12 hrの明暗環境下で1-2週間単独飼育を行い、行動リ ズム解析にて同調を確認後試験に移った。本実験は九州大学動物実験規則を遵 守して行った。
1-3. 行動リズム解析
自発行動リズムの測定は、20×30×12 cm のケージに一匹ずつ動物を入れ、
真上に設置した放出赤外線検出型の自発運動量センサー(NS-AS01、ニューロサ イエンス)を用いて、1分間隔で行った。データ収録システム(NS-DAS-32、ニュ ーロサイエンス)にてコンピュータ上で運動量を記録し、ClockLab(ニューロサ イエンス)を用いて行動開始時点を算出した。
1-4. 再同調促進作用の評価
個別に飼育し明暗環境に同調させたCCK-1受容体KOおよびWTマウスを、
実験開始当日(day1)に新規明暗環境として明暗周期を 6 時間前進させた。一日 後(day2)、新規環境の暗期開始直後(ZT12)にWTマウスに対してはsalineまた はlorglumideを体重あたり0.2 mg/kgで、KOマウスに対してはsalineを腹腔 内投与した。30分後、0.5% CMC水溶液(VEH)またはRAMを体重あたり0.1
mg/kgで経口投与した。前日と比較して行動開始時点の時間のずれが30分以内
である状態が三日以上続けて認められたとき、行動位相の同調が完了したもの と定義して明暗周期の変更から同調完了までの日数を測定した。
44
1-5. ウエスタンブロッティング
明暗環境で飼育したマウスをZT14において頸椎脱臼し、素早く脳を取り出し た。その後、SCN領域を含む1 mm3の切片を切り出し、100 µLのLysis Buffer を加え、氷冷中で30分間おき、14,000 rpm(16,000×g)10分間4℃で遠心し た。上清を回収し、5 µLの上清に対して25 µLの6x sample bufferを加え混和 した。5分間ボイル後に室温で5分間冷やし1.5 mm厚の10% acryl amideゲ ルに30 µLずつロードした。Running bufferを注ぎ、80 V 20分間泳動し、次 いで120 Vで90分間泳動した。泳動したゲルをmembrane transfer bufferに 移し、80 Vで60分間泳動し、メンブレンにタンパク質を転写した。ブロットし たメンブレンを回収し、blocking bufferに移して振盪しながら室温で1時間イ ンキュベートした。1 次抗体として抗 MT1R (CUSABIO)、抗 TATA binding protein TBP抗体(Abcam)、抗β-Actin (Santa Cruz)、抗p-CREB-1 (Santa Cruz) をblocking bufferで1:1000に希釈し、メンブレンを浸して4℃で一晩インキュ ベートした。PBS-Tで5分間3回洗浄し、blocking bufferで1:1000に希釈し たペルオキシダーゼ標識2次抗体でメンブレンを室温60分間インキュベートし た。PBS-Tで10分間3回洗浄し10 mL ECL(enhanced chemiluminescence
substrate)で1分間インキュベートした。ブロット膜をX線フィルムに感光し
て検出した。
1-6. 統計解析
Day0-10の各日において、VEH群とRAM群それぞれでWTマウスへのsaline、
lorglumide投与およびsalineを投与したKOマウスで行動開始時点の前進に違
いがあるかを比較した。また、再同調完了までに要する日数を、それぞれの前投 薬の条件下におけるcontrol群とRAM群とでstudent’s t-testにて比較した。
45
2. 結果
RAMによる再同調促進作用を検討するために、明暗=12 hr:12 hr の環境に 同調後、6時間明暗環境を前進させることで、時差ぼけ状態を作り出し、解消さ れるまでの行動を観測した。行動記録の代表例をFig. 19に示す。
Fig. 20 に明暗環境変化後 10 日目までの行動開始時点の前進を示す。また、
Fig. 21に再同調完了までの日数を示す。
WTのsaline前投薬群においては、VEH群に比べ、RAM群で投与後2日後
まで行動開始時点の前進が大きい傾向にあった(各群 n = 10, Day3, RAM vs VEH = 3.06±0.44 vs 2.14±0.38, P=0.138; Day4, RAM vs VEH = 4.55±0.39
vs 3.22±0.43, P=0.072)。また、再同調完了日までの日数はRAM群が有意に短
かった(n = 10, RAM vs control = 6.10±0.128 vs 7.40±0.15, P=0.028)。一方、
WT の lorglumide 前投与群における投与では、RAM 群での行動開始時点の前
進が抑えられ、再同調までの日数に有意な差はなかった(n = 10, RAM vs control
= 8.0±0.18 vs 9.0±0.12, P=0.080)。また、KOマウスでもlorglumide投与と Control RAM
4 0 3 2 1
5 6 7 8 9 Day
Fig. 19. 再同調完了までのアクトグラムの代表例
矢印の時点で投与を行った。グレーの背景の部分は暗期、白の背景 の部分が明期である。