九州大学学術情報リポジトリ
Kyushu University Institutional Repository
モルモット腸間膜動脈最終分岐部以降の血管平滑筋 細胞に高密度に存在する新型nifedipine非感受性高 電位活性化型Ca^2+チャネルの電気生理学的および薬 理学的特性の検討
森田, 浩光
九州大学歯学研究科歯学臨床系専攻
https://doi.org/10.11501/3180214
出版情報:Kyushu University, 2000, 博士(歯学), 課程博士 バージョン:
権利関係:
Electrophysiological and pharmacological properties of αnovel type of nifedipine-insensitiveち
high voltage-activated Ca2+ channels,
which predominantly exist in the terminal mesenteric arteηof guinea pig.
A.D.2001
Hiromitsu MoritaThe Second Department ofOral and Maxillofacial Surgery,
Faculty ofDentα:1 Science,
Kyushu University, Fukuokα, 812-8582, Japan
モノレモット腸間膜動脈最終分岐部以降の
血管平滑筋細胞に高密度に存在する 新型nifedipine非感受性
高電位活性化型Ca2+チャネノレの
電気生理学的および薬理学的特性の検討
2001年
森田浩光
九州大学大学院歯学研究科歯学臨床系専攻第二口腔外科学 (主任:白砂兼光教授)
九州大学歯学部附属病院特殊歯科総合治療部 (主任:安部喜八郎助教授)
指導:伊東祐之教授、井上隆司助教授 九州大学大学院医学研究院
分子常態医学部門生体情報薬理学
Contents
1. H!g.………・・・………・・………I
2. 要E………・・・………・・………2
3. 材料と方法…・………・……・………・………・……・……・……4
4. 結果………一…… …一………12
4-1. モノレモット揚間膜動脈最終分岐部平滑筋細胞に高密度に存在するnifedipine 非感受性高電位活性化型Cæ+チャネノレの電気生理学的、および薬理学的特性…12 4・2.L型および新型nifedipine非感受性高電位活性化型Ctl+チャネノレの加齢による 変化………...…・…・・…・・・………ー…・… …・…..…. .一-一…....一一…・…・・・……・…22
4-3. 新型nifedipine非感受性高電位活性化型Ca2+チャネノレに対するP2Y受容体を 介したATPによる制御… - ………・………...….24
5. 考察..……… ………一………30
湯原……... ....・・……・ ・・…・・・…・・・・・・……... ....・・・・・・・・…...……… ・・・・・・・……40
参考文厳…...・・・・・・・・………・・・・・・…・・・・・・・…………・…・・・・・・… -…………・・・・……41
本研究の一部は、Circ. Res汁85, 596-605, 1999似orita H., Cousins H., Onoue H., 1to Y., Inoue R. Predominαnt Distribution of Nifedipine-1fj問nsitive, High Vo/tαge-Activated Cd+ Chαnnel in 刀2e GuineαPig Terminal Mesenteric Artery)に掲載された。
1. 緒言
日甫乳類の血圧調節に深く関わる血管の筋緊張度は、自律神経や血管内皮細胞など から放出される種々の血管作動性物質や、圧変化等の機械刺激で惹起される内因性 筋収縮反応等により、効果的に制御されているl七これらの制御には、平滑筋細胞膜 の過分極および脱分極に伴うカルシウム(Ca2)透過性変化(従って結果的には平滑筋 細胞内 Ca2+濃度変化)が、中心的な役割を果たしている140これまで、このような Ca2+
流入経路として、dihydropyridine(DHP)感受性、高電位活性化型、L型Ca2+チャネル の重要性が強調されてきた。その理由として、このチャネルが全身の血管に分布して いることや、このチャネルに対する選択的阻害薬が血管緊張度や血管外径の変化を 著しく抑制することなどを挙げることができる470しかし、このような研究の 大半は、技術 的な困難から主として外径が比較的大きく(100---1000μmの範囲内)、数層の平滑 筋を有する中枢側の血管平滑筋に限られてきた。
一方、全身の血圧調節には、高密度の交感神経支配を受け一層の平滑筋を有する、
末梢の微小抵抗血管が重要で、あると考えられている830このことは、血圧制御におい て L型Ca2+チャネ ノレのみに着目して論じることの危険性を示しており、それ以外のCa2+
透過性チャネルが微小血管の緊張度の制御に重要な役割を果たしている可能性を暗 示している。
実際、我々は、モルモッ トの小腸粘膜下動脈や腸間膜動脈最終分校に は 、 nifedipine (DHP系Ca2+チャネノレ阻害薬)非感受性、電位依存性 Ca2+チャネノレが予期 せぬほど優勢に発現していることを発見したlooこれまで、様々な血管平滑筋組織で、
DHP に非感受性で速い不活性化を示すCa2+チャネル(T型Ca2+チャネルなど)が報 告されているが、このチャネノレの発現の程度や分布は、L型Ca2+チャネルに比しかな り限られており(参考文献11 ラット大動脈平滑筋培養細胞を除く)、その生理学的意 義を疑問視する向きも多いIl-170
以上の理由から、我々はモノレモット腸間膜動脈の末梢部平滑筋細胞 に存在する nifedipine非感受性電位依存性Ca2+チャネノレに焦点を合わせ、その生物物理学及び 薬理学的性質を解明すべく本研究を遂行した。
2. 要旨
(1 )モルモット腸間膜動脈最終分岐部及び それより末梢の平滑筋細胞に、パッチクラ ンプ法 を適用して、 これまでにないユニークな生物物理学的、薬理学的特性を示す、
nifedipine非感受性、急速不活性化型、電位依存性Ca2+チャネノレ(電流)が存在して いることを発見した。全Ca2+電流に対するnifedipine非感受性Ca2+電流の割合は、 腸 間膜動脈の中枢部から末梢分校部へ向け劇的に増加し、その 最末梢分岐部では殆
ど1000/0を占めていた。この Ca2+電流は、電荷担体として5mmol/Lバリウム(Ba2つを 用いた条件下で、-50mV付近から活性化され-10mV付近で最大活性化を示し、その 500/0活性化値および 不活性化値はそれぞれ-llmV及び -52mVで、あったOまた、その 活性化曲線と不活性化曲線の重なる範囲が-60�-30mVにあることから、静止膜電位 付近で恒常的に活性化されている可能性が示唆され た。
nifedipine非感受性Ca2+電流は減衰(rundown)しにくく、Ca2+ 依存性不活性化を示 さなかったOまたCa2+ より Ba2+ に対し約2倍高 い透過性を示し、ニッケノレ(N?-'-)よりカド ミウム(Cd2+)�こよってより 強い阻害を受ける(50%阻害値(IC50), Cd2+ で6μmol/L、Nj2+
で68μmol/L)など、 高電位活性化型の電位依存性Ca2+チャネノレに特徴的な性質を 示した。
更にN型、 及び P/Q型Ca2+チャネノレの選択的阻害薬で、あるω-conotoxin G VIA 、M VIIC 及びω-agatoxin IV A (各々lμmollL)に対して感受性を示さず、一方
nimodipine (10μmol/Lで約400/0)及びT型Ca2+チャネルの一般的な阻害薬で、ある amiloride (1 mmol/L で約70%、1C50� 107μmollL)では部分的阻害を受けた。これらの
特徴はR型Ca2十チャネルを想起させるもので、あったが、逆転写遺伝子増幅法(RT- PCR法)によってモルモット腸間膜動脈からαlEの messenger RNA (mRNA)を検出
することはできなかった。
以上の結果から、 全く新しい特徴を有するnifedipine非感受性高電位活性化型Ca2+
チャネノレが腸間膜動脈の末梢分岐部に優勢に発現しており 、その血管緊張度の制御 に深く関わっていることが強く示唆された。
(2)腸間膜動脈の中枢及び 末梢側におけるnifedipine感受性及び非感受性高電位 活性化型Ca2+チャネル電流密度の加齢に伴う変化を観察したO中枢及び末梢側の い ずれ の部分においても、nifedipine感受性、即ちL型Ca2+電流の 密度が週齢の 増加 に伴ない約3 倍まで増加するの に対して、nifedipine非感受性Ca2+電流密度は約半 分まで減少した。またこれに対応して、前者の後者に対する比は、 約1 :2から2:1へと 逆転した。この ことから、幼齢期にはnifedipine非感受性高電位活性化型Ca2+チャネ
ルが、 また高齢期にはL型Ca2+チャネルが主に分布していることが明らかになったO (3 ) nifedipine非感受性Ca2+電流に対する種々の血管作動性物質物質の効果につ
いて検討した。このうち交感神経伝達物質であるadenosine 5にtriphosphate (ATP)は、
異なる2つのP2Y受容体を介し、低濃度で増強、高濃度で抑制の二相性効果を示し た。このことから、nifedipine非感受性高電位活性化型Ca2+チャ不ルの活性が、交感 神経の活動によって効果的に制御されていることが示唆された。
3
3. 材料と方法
3-1. 細胞分離
1 00'"'-'2000g ( 生後 2�20 週齢)のモルモット(雌雄混合)を後頭部殴打によって気絶 させ 、断頭致死さ せた。腹部を切開後、小腸こ、と腸間膜動脈及び静脈を摘出し (Figure lA)、0.5mmol/L Ca2+を添加した生理的食塩水(PSS)を満たした解剖用デ、イツ シュのラバー底に、 細いピンを用いて注意深く伸展させた状態で固定した。実体顕微 鏡下に、先端の鋭いヒ。ンセット及び眼科用ハサミを用いて腸間膜動脈最終分校部か ら動脈標本(40'"'-'100μm )を10'"'-'20本作製した。次にこれらの標本を 350C、Ca2+除 去PSS中で約 30分間温め弛緩させ、同温度にて 2mg川Lのコラゲナーゼ(type 1;
sigma, USA)を添加したCa2+除去PSS中で約45 分間、酵素処理をした。最後に、先 端を熱形成したパスツールヒ。ベットを用いて、酵素処理によって消化した血管 標本を Ca2+除去PSS 中で出来るだけ緩やかに振重し、 単一細胞へと分散させた。このように して得られた細胞は、lmmol/Lの Ca2+を加えた生理的食塩水中(氷温:4 '"'-' 1 OOC )に 保存した(FigurelB)。
一方 、粘膜下部の血管については、まず小腸粘膜をヒ。ンセットにて機械的に剥し\だ 後、 はさみを用いて切り出し、0.5mg!mLコラゲナーゼを添加したCa2+除去PSS中に て、350Cで10'"'-'20 分間弱い酵素処理をおこない、 血管部のみを単離した。その後の 酵素処理は、上述した通りである。
細胞は単離した後、6時間以内に実験に供した。以上の方法は九州大学動物実験 倫理委員会の規定に基づくものである。なお第一部の実験(4-1)では、200�250g (生 後4�5週齢)のモノレモットを使用した。
3-2. 電気生理学的研究方法
パッチグランプ。法:増幅器としてAxopatch lD (Ax:on Instruments, USA)を、 そしてデ ータの収集、 及び解析用コンピューターとして Aptiva (ffiM, USA)を使用した。コンヒ。
ューターからの矩形電圧を電位固定した細胞へ加える目的で、また細胞から記録した シグナルをデジタル化しコンピューターに記録させるため、TL-1 ADIDAコンバーター (Axon Instruments, USA)を使用した。電流シグナルはl妊Izの low-passフィルターを かけ、 さらに 2kHzで数値化した後、 コンピューターのハードデ、イスクに保存したOリー ク電流補正には、P/2あるいは P/4法を用いた。すなわち、脱分極ノミノレスを与える直 前に2回、 もしくは4回に分割した過分極ノミルスによって得られた電流波形を加算し、
テストパノレスで、惹起された電流波形に加えることにより補正を行った。また、 数例の実 験ではCd2+存在下で得られた電流波形を引き算することによりリーク補正を行った。
A
B
veln a吋ery
._
Ileum
Figure 1. Microphotographs of the guinea-pig mesenteric vasculature and single arterial myocytes dispersed企om it.
A, bifurcation of mesenteric紅白rial vascula旬re. B, single smooth muscle cells enzymatically dissociated from A (arrow).
5
今回用い た実験条件下におけるシリーズ抵抗値は平均11 +O.4MQ (n=245)であり、
その 50�70%を電気的に補正した。全てのデータ解析用の ソフトウェアーとして pClamp version 6.03 (Axon Instruments, USA)、 およびデータ提示用のソフトウェアー としてKaleida Graph version 3.04 (Hulinks, USA)を使用 した。全ての実験 は室温
(200C -250C )で、11ったO
3-3. 逆転写遺伝子増幅法(RT-PCR法)
腸間膜動脈最終分枝のL型及びR型 Ca2+チャネルの発現を見るため、それぞれ のCa2+チャネノレのαlサブ、ユニットに特異的なフ。ライマーを用い、RT-PCR法によって 増幅され た遺伝子産物の検出を試みた。
モノレモット (200-250g)の小脳及び腸間膜動脈最終分枝 より RNA 抽出キット(ぬlesay Mini-Kit, Qiagen, USA)を用いて 全 RNAを抽出 した。全RNA量lμgを用いて、ラ ンダムヘキサマー、 逆転写酵素(Superscript n, Gibco-BRL, USA)による 逆転写(RT) 反応(全反応容量=20μL )を行い、RNA をcomplementary DNA(cDNA)へ変換さ
せた。 遺伝子増幅法 (PCR)のフ。ロトコーノレ熱変性: 950Cで30秒、アニーリンク、、500C (αlC ) または540C (αlE)で 30 秒、延長反応 720Cで 1分。まず1回目のPCRを逆 転写(RT)反応液 lμL、1st フ。ライマーのフォワード、及びリバースプライマーを用い て10μLスケールで、20サイクル行った後、得られたPCR反応液lμL、nested プライ マーのフォワード 及び、リバースフ。ライマーを用いてさらに30サイクル増幅 したo PCR 法により増幅された遺伝子産物 は 2%のアガロースゲ.ノレで、電気泳動し、エチジウムブ
ロマイド・で、染色した後、紫外線照射下に写真撮影した。
PCRに用い たプライマー は以下の通りである。αlC(L 型)サブユニット, forward:
GGAGTTGGACAAGGCTATGAAGGA ( 1st PCR ( 5306-5329 ) ) CACCGTCCCATGAGAAGCT ( nested PCR ( 5500-5518 ) ) 、 reverse:
GACCT AGAGAGGCAGAGCGAAGGA (1 st and nested PCR (5958・5935) )、αlE(R 型)サブユニット: forward: CTTCCT-GAGGATGACAAGACC(日rst PCR (381-401 ) )、
GAAGTCCATCATGAAGGCCA ( nested PCR ( 680-699 ) ) 、 reverse TCAATGGAAGGCATGTTGG ( 1st PCR ( 2287田2269 )
AGCAAGCATGACTTCCTCTG(nested PCR(1235-1216) ) 。 プライマーの設計には、 既に報告されているcDNA配列(αlCサブユニット, Genba此L 15453,αlEサブユニッ ト; Genba叫c M59786)を参考にした。
3-4. 溶液組成
実験に 用いた溶液組成は以下の通りである。(mmollL ) ナト リウム(Na+) 140, カリ
b
A a
Proximal branch (2nd,3勺
Submucosal branch10pA
200ms
nv ハU門ノι
川
P<O.01 100
80-
20 60
40
問ハU(ポ)(一一同心O一。\一ー一Z)ωogcωU」ω止
B
Submucosal branch T erminal branch
Proximal branch (2nd,3rd
。
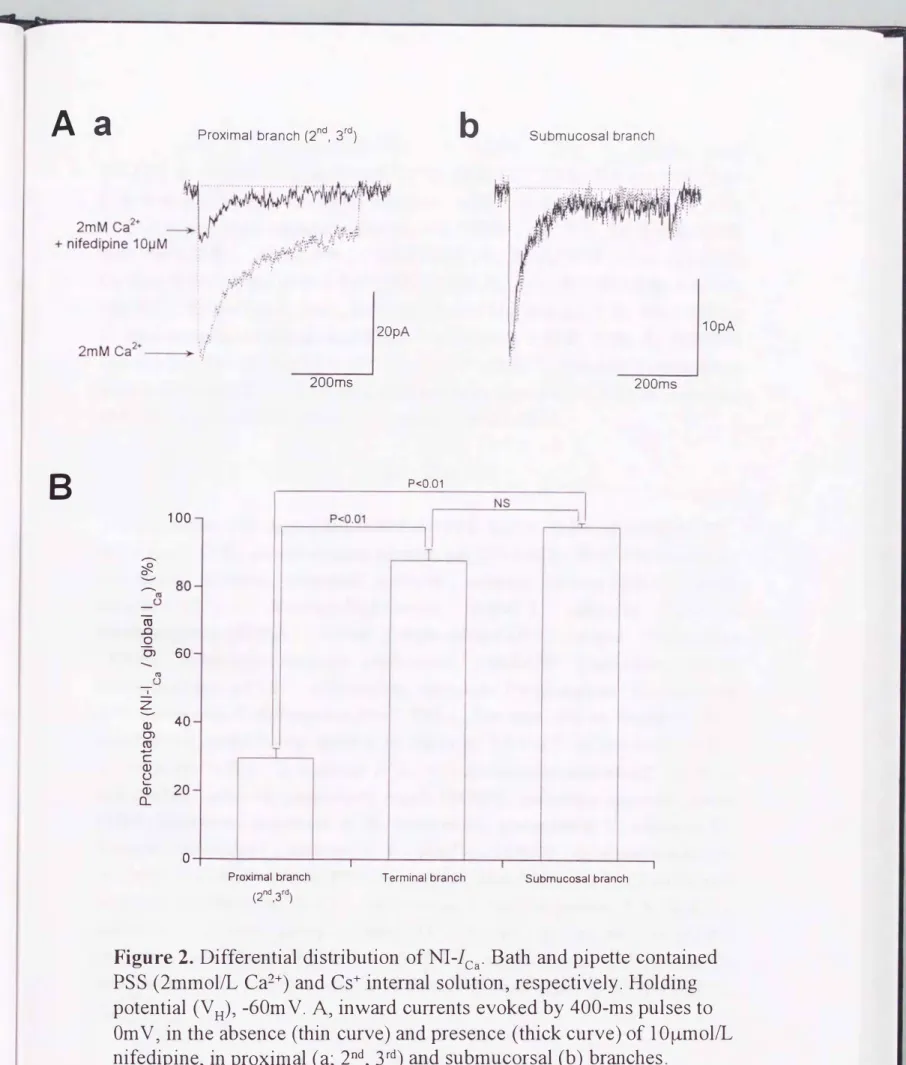
Figure 2. Differential distribution ofNI-lCa. Bath and pipette contained PSS (2mmol/L Ca2+) and Cs+ internal solution, respectively. Holding potential (V H)ヲ-60mV. A, inward currents evoked by 400-ms pulses to OmVヲin the absence (thin curve) and presence (thick curve) of 1 O�mol/L nifedipineヲin proximal (a;
2ベ3rd)
and submucorsal (b) branchesBヲpercentage ofNI-ICa to globallCa. The number of cells tested were 8ヲ 久and
3
for proximalヲterminalヲand submucorsal branchesョrespectively.P values indicate the results of the Tukeyヲs multiple comparison test.
7
ウム(K+) 6, マグネ、ンウム(Mg2+) 1.2, Ca2+ 2, 塩素(Ct) 151.4, グルコース10, 2-[4- (2-hydroxyethyl)ーl-piperazinyl]ethanesulfonic acid ( 1王EPES ) 10 ( pH 7.4,
tris[hydroxymetyl]aminomethane (Tris base)にて調整)0 Ba2+及び、Ca2+を増量する場 合は Na+を減量し、浸透圧が等しくなるよう調整した。低Ca2+溶液(0.5mmolfL)及び
Ca2+除去溶液は Krebs液中のCa2+を減じて作製したOさらに第三部の実験(4-3)にお
ける Ca2+, Mg2+除去液は、Krebs液からCa2+および、Mg2+を除去した上、キレート斉IJとし てethylenediaminetetraacetic acid (EDT A) 200μmoVLを添加し 作成した。電極内液 (mmollL)は: Cs+ 140, Mg2+ 2.0, Cl- 144, クレアチン燐酸5, adenosine 5' -triphosphate sodium salt (Na:!ATP) 1, 0,0にbis(2申aminoethy1 )eth y 1 ene-gl y co 1-N ,N ,N' ,Nヲーtetraacetic acid (EGTA) 10, HEPES 10 (pH 7.2, Tris baseで調整)である。
3-5. 薬品
ATP、guanosine 5にtriphosphate(GTP)、HEPES、EGTA は同仁化学(熊本)より、
tetrodotoxin (TTX)、phosphocreatine disodium salt は和光純薬(東京)より、nifedipine、
nimodipine 、 diltiazem 、 verapamil 、 amiloride 、 cadmium chloride ( CdCl2 ) 、 nickel chloride ( NiC12 ) 、 N-methyl-D-glucamine ( NMDG ) 、 guanosme デー0- ( 2- thiodiphosphate) ( GDP pS )、uridine デーtriphosphate (UTP ) 、 uridine デーdiphospate (UDP)、 2-methylthyoadenosine triphosphate ( 2-MeSATP ) 、 adenosine デー0-(3- thiotriphosphate ( ATPyS ) 、α3・metylene adenosine 5' -triphosphate (α点-methylene ATP)、adenosine 5' -diphosphate (ADP)、EDTA、Tris base、sodium f1uoride (NaF)、
noradrenaline、acetylcholine、suramln は Sigma社(USA)より、ω-conotoxin G VIA、
ω-conotoxin M vn C 、 ω-agatoxin rv A、 4,4'-diisothiocyanostilbene-2,2にdisulfonic acid (DIDS)、calcitonin gene-related peptide (CGRP)、vasoactive intestinal peptide (VIP)、bradykinin、neurokinin 九B、somatostatin、neuropeptide Y、substance P、
histamin、endothelin-1、angiotensin- n、 [Arg8]-vasopressin 、 piridoxalphosphate-6- azophenyl-2' ,4'ーdisulfonic acid (PP ADS)、protein kinase A inhibitor 14-22 amide, cell
permeable, myristoylated (PKA-I)、protein kinase C inhibitor peptide 19-36 (PKC-I)、
adenosine 3' ,デーcyc1ic monophosphate,Nへ02'-dibutylyl-, sodium salt ( db-cAMP ) phorbol-12,13-dibutyrate ( PDBu ) 、(ト[6由((17 ß -3-methoxyestra-1,3,5(10)ーtrien-17- yl)amino )hexyl]-lH-pyrrole-2,5-dione} (U-73122)は Calbiochem社(USA)より購入し た。
3-6. 解析
全てのデータは平均±標準誤差で表した。また有意差の検定には、ステューデ‘ント
_jar釘r巾b
50msb
山仰
OmV
」州
a
5mM Ca2+
一川
+ nifedipine 10ドM
5mM Ba2+-一予
+ nifedipine 1 0ぃM
A
• IC =34nM
50
n=1.0 ー
B
0.95 0.9 0.85
-Cω」」コυω〉一】同一ω区
0.8
105 104
1000
1 10 100
Nifedipine (nM)
0.1 0.01 0.75
0.001
nifedipine 10ぃM 140
マ5
こ4♀0
いM。対�
nifedipine-insensitive プ
current 120
80 60 40 100
(〈己)ω刀コザ一一己E何言ω」」コO
C
20
。
。 1500
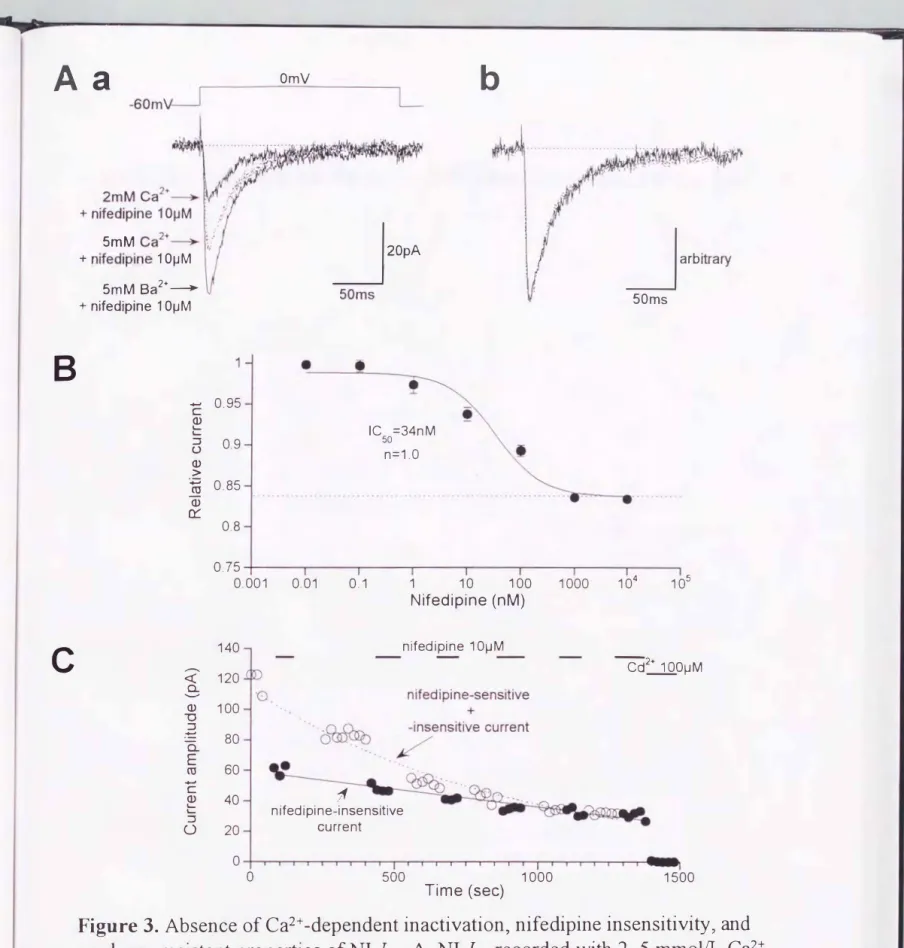
Figure 3. Absence of Ca2+ -dependent inactivation, nifedipine insensitivity, and rundown-resistant properties ofNI-1Ca' AヲNI-九a recorded with 2, 5 mmolfL Ca2+
and 5 mmolfL Ba2+合om the same cell (a).To compare the waveformフthe cu汀ents are scaled to give the same peak amplitude (b). Bヲconcentration-inhibition curve for nifedipine fi口ed to an empirical Hill equation:
(l-r)/ {l +([ drug]/K)n}+r, Ki'
nヲand
r
denote the drug concentrationヲits ICso valueヲcooperativity factorヲand the 企action of unblocked cu汀entヲrespectively. Data were evaluated at Om Vヲwith from 3 to 6 cells. V H=-80m V. Cフrundown time course of global(0)
and NI (lOmmol江)-ICa(・)
during intemal dialysis of nucleotide-f代e Cs+ intemalsolution. Bottom axis denotes time elapsed after onset of whole-cell configulation.
1000
Time (sec)
500
9
のt検定法、Turkey多重比較検定法、一元配置分散分析法(one-wayANOV A)を適 宜使用した。
A
B
�
、』C-L句〆、
てωコ 辛コd
0. E
rn ー
」Cω
L --•
。コ
0.
5
10
。 -10
-20
ー30
-40
ー50
-80 -60 -40 -20 0 20
Membrane potential (mV)
-60mV o→ ...・・-ーーー
さ0.
Ll固5
0...
申d〉、
g -1
てコω 噌圃dc
E-15
仁ココ
-2
-2.
5
-80 -60 -40 -20 0 20 40 60
Membrane potential (mV)
40 60
control
+nifedipine 1 O�M
20pA 200ms
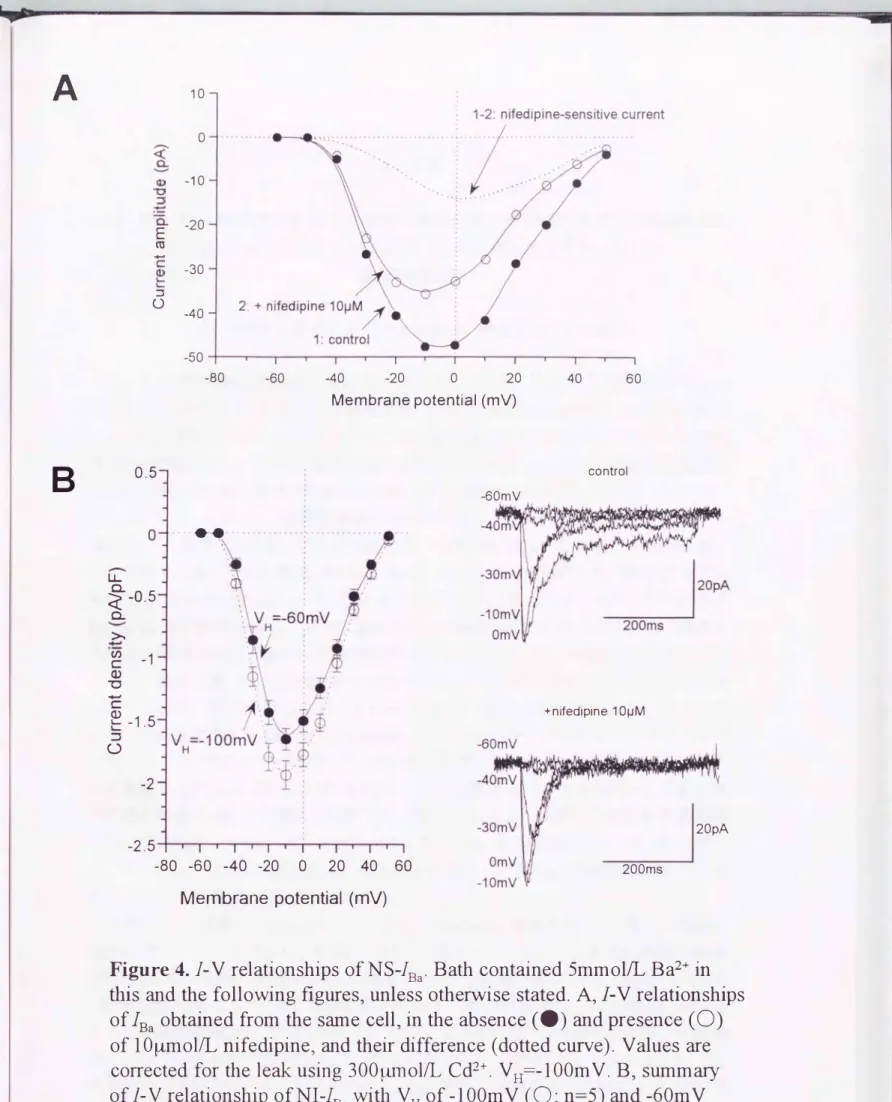
Figure 4.
1-
V relationships ofNS-1Ba.
Bath contained 5mmolIL Ba2+ in 白is and the following figuresヲunless otherwise stated. AヲιV relationships oflBa
obtained from the same cellヲin the absence (・) and presence(0)
of 10�mollL nifedipineヲand their difference (dotted curve). Values are corrected for the leak using 300�ol江Cd2+. VH=-100mV. Bヲsumm征y ofιV relationship of
NIーらa
with VH of -100mV(0;
n=5) and -60mV (・;nニ8). Data are mean + SEM. Illustration on the right side indicate ac加al traces at -60ヲー羽田30. -10. and OmV.4. 結果
4-1. モルモット腸間膜動脈最終分岐部平滑筋細胞に高密度に存在するnifedipine 非感受性高電位活性化型Ca2+チャネノレの電気生理学的、および
薬理学的特性
腸間膜微小動脈におけるnifedipine非感受性Cd+電流
モルモット腸間膜動脈最終分岐部 (Figure 1 A)より 単離した単一平滑筋細胞(Figure
1B)は、長方形または長楕円形の様相を呈しており、長径が50-70μm、短径が約5
μm、入力電気容量が13.6+ 0.2pF ( n=205,範囲; 8�23pF)で、あったOこれらの筋細胞 は、パッチ電極を近づ、けるよ電極先端から漏出したATPに 反応して急激な収縮を起
こした。これにより非筋細胞との識別を容易におこなうことができた。
Figure 2は、モルモット腸間膜動脈の中枢側(第2,3 分岐部)、最終分岐部 及び 小 腸粘膜下分枝部より 単離した それぞれの単一筋細胞において、OmVへの脱分極パ ノレスで活性化される Ca2+電流 を、10μmoνL nifedipineの存在下及び非存在下 で比 較したものであ る。nifedipineは、このCa2+電流に対し、部位によって異なる抑制効果 を示した。すなわち中枢側の分枝においては、nifedipine の前投与により Ca2+電流の
大部 分が消失したが、最終分枝及び粘膜下分枝部 では、ほんの僅かな抑制しか見ら れず、一過性で速い不活性化を示 す成分が残存した。このnifedipine 非感受性 電流 成分の振幅は、電位依存性Na+ チャネノレの阻害薬で、あるTTX 10μmoVLやCl- チ ャネノレの阻害薬で、あるDIDS 100μmollLの前投与によって抑制されず、またNa+や
Cl-をそれぞれN孔⑪Gやベンゼンスルホン酸で、置換しでも全く影響を受けなかった(デ ータ未提示)0 Figure 2B にまとめたように、全Ca2+電流に対する nifedipine非感受性 Ca2+電流の割合は、 中枢側より粘膜下及び最終分岐部より単離した血管平滑筋細胞 において、有意に大きかった(粘膜下分枝, 98 + 10/0, 最終分枝, 87+3%, 中枢側 分 枝; 30+3%)0これらの結果は、nifedipine非感受性Ca2+電流が腸間膜動脈系の末梢
部に向け豊富に発現していることを強く示唆している。
生理的 Ca2+濃度( 1-2mmol!L) で記録した nifedipine非感受性Ca2+電流の電流 振 幅は非常に 小さくら�20pA)、 電流とノイズ、との差が小さいため、定量的な評価が困難 で、あった。この問題を回避し、さらに 細胞内へのCa2+流入によって二次的に流れる Ca2+依存性 電流によって生じる誤差や、大量のCa2+の反復流入によ る細胞収縮の結 果生じる膜容量減少の影響を少なくするため、細胞外Ca2+(2mmoVL)を僅かに高濃
度のBa2+(5mmoIlL) で置換した。L型Ca2+チャネルにおいては、この方法によって、
電位依存性をほとんど変化させることなしCa2+電流の振幅が数倍に増加することが報 告されている210実際5mmolILBa2+を用いることによって 、nifedipine非感受性Ca2+
A
ぽ〉ー・会。明V
au c
氾 V 一山mV 百 5
m
日 5 4 6 4 8 陪=5-一-wo k -PV Au e rTE nH 0.8
0\の 2V円伺 0.6
0.4 0.2
60
-40 -20 0 20 40
Membrane potential (mV)
-60 ー80
。
•
•
?、
• 168 6
』守
(ωE)(さωU」ωaootOF)ωεzco一石〉一ぢ〈
12 10
B
4 2
。
40 60 20
Membrane potential (mV)
ー20 。 -40
Figure 5. Activation characteristics of N1-1Ba. A, activation curves evaluated by tail current analysis (inset)・ヲsolid curveラNI-らa (n=7)フ 0, dotted curveヲ
NSーらa
(ieラ global minus NI -lBa; 白t of data points with the Boltzmann equationヲas follows: l=lma/[l+exp(Vm-VO.S)/k]ヲ
where 1ふax' V mラVo.sヲandk denote出e tail current amplitude conditioned to
V
m' its maximumラmembrane potentialヲ500/0 activation potentialヲand slope factorフrespectively. Bヲrelationship between the membrane potential and the time constant for 100/0 to 900/0 activation. Data are mean +SEM
金Oill..4___cells._
13
電流の電流 振幅は約2倍に増加したが (Figure3Aa )、その波形に殆ど変化はなか っ た(Figure3Ab)。このことは、この成分に Ca2+依存性の不活性化 がないことを示唆して
いる。また、2mmolIL Ca2+及び 5mmoVL Ba2+で、得られたnifedipine非感受性電流の 電流一電圧関係 を、各々のピーク値 で正準化 し重ね合わせるとほとんど一致したこと から (データ未提示)、これら 2つの条件下で、得られ る nifedipine非感受性Ca2+電流 の電位依存性は、ほぼ同等で、あることが示唆されたO
Figure 3Bは、5mmolν/江L Ba2+濯流下で
度抑制曲線を示す050%抑制を引きおこ す値 (IC5o)は 34nmoVL であり、lμmoVL以 上の高濃度では、それ以上の抑制が観察されなか った。この34nmolILとし\う値は、過 去に報告された他の組織における L 型Ca2+チャネノレの値と同等で、あったOこのことか ら、本研究で用いた腸間膜動脈のnifedipine感受性成分は、典型的な L型Ca2-rチャ ネノレ活性を反映してい ると考えられた。実際、パッチ電極内からヌクレオチド、燐酸を除 去 したり5mmol/L フッ素(F-) を添加した時に、nifedipine感受性成分 が速かな減衰を 示す(Figure3 C, 0)ことは、この解釈をさらに支持する ものである。11 これに対して nifedipine 非感受性成分は、これらの処置に対し比較的抵抗性を示 した(Figure3C,
・)。そ してnifedipine感受性成分が完全に 減衰した後、1 または10μmol/Lの nifedipineは、残存したnifedipine 非感受性 Ca2+電流に全 く効果を示 さなか った(対
照の101 + 10/0; n=5 )。
これらの結果は、nifedipine非感受性成分が、部分 的に し か 抑制されずに残存した L型Ca2+電流を反映してい るので はないことを強く示唆している。また同時に、
nifedipine がnifedipine非感受性Ca2+チャネルとL型Ca2+チャネルとを選択的に分離 する道具として有用で、あることを示唆している。
nifedipine非感受性ccÝ守竜流は高電位活性化型ccÝふチャネノレである
Figure 4 A は、 同一細胞から記録したnifedipine感受性及び非感受性 Ba2+電流の 典型的な電流-電圧関係を示す(保持電位ニー100mV)。またFigure4B は、保持電 位 を-60mV又はー100mVにしたときのnifedipine非感受性Ba2+電流の電流一電圧曲 線を複数の異なる細胞からの平均として(n=5�8)表したものである。これらの結果から、
nifedipine 非感受性 Ba2+電流は、保持電位に 関係なく- 50mV よりも脱分極側で 活性 化 され、-10mV付近で最大振幅を示すことが明ら かになった。
一方、nifedipine感受性Ba2+電流 活性化 の関値および最 大活性化 電 位は 、 nifedipine非感受性Ba2+電流 (Figure4A, 点線 )に比べて僅か に脱分極側にあったが、
それらの差は10mVに満たなか ったo nifedipine感受性(Figure5A, 0)および nifedipine非感受性(Figure5A, ・) Ba2+電流の電位と活性化の関係は、S状曲線に 従い、Boltzm ann回帰によって得られた50%活性化 値と傾斜係数はそれぞれ、
A
+20mV -50--110mV耕:為
一
泌総線側き
�_::o.戸y、よ士宇・,:唱・ 、明司、 外\,�晶、τ?:,",.'T
' ー ・ ー .
"ム
品、 、・� ,;....:,降、‘・--
A nv nu にJv
m
B
10 cñ、、
E 8
E 咽C 8
6
Q) E 4回d
。+Jー由亡c〉に田s 3 O 4
2
。
-120 -100 -80 ー60 -40 -20
Membrane potential (mV)
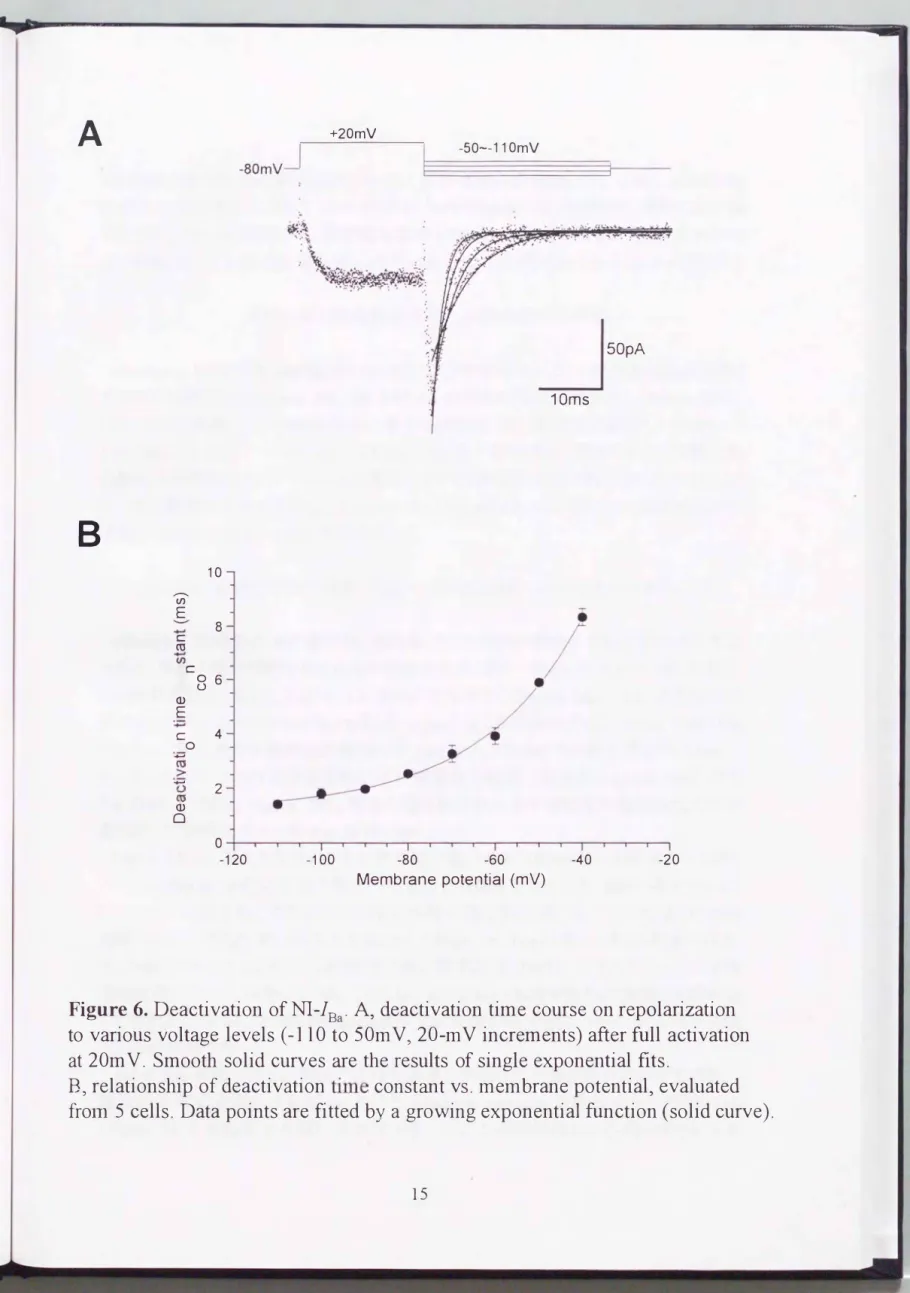
Figure 6. Deactivation ofNI-lBa. Aヲdeactivation time course on repolarization to various voltage levels (-110 to 50m Vヲ20-mV increments) after full activation at 20m V. Smooth solid curves訂e the results of single exponential fits.
Bヲrelationship of deactivation time constant vs. membrane potentialラevaluated frolll 5 cells. Data points are fitted by a growing exponential function (solid curve).
15
nifedipine感受性Ba2+電流に対してー15,5及び-8.4mV CFigure 5A, 点線)、nifedipine 非感受性Ba2+電流に対してー11.0及び・11,3mVCFigure 5人実線)で、 極めて近い値 で、あった。これらの結果から、腸間膜動脈のnifedipine非感受性Ba2+電流(チャネノレ) は、高電位活性化型電位依存性Ca2+チャネルとして分類可能で、あることが判明した。
活性化及び脱活性化キネティクスの電位依存性
nifedipine非感受性Ba2+電流の活性化はミリ秒単位で、起こり、 脱分極の程度を強く するほど加速されたCFigure 3 A、4 B 及び6A の実際の電流記録参照)0 Figure 6B に 示すように(実線)、10�90%活性化に要する時間は、 強く膜電位に依存し( -30mV で 13 +2ms 対 20mVで4+1ms)、48,2mV当たりe倍の減少が認められた。同様に脱 活性化の時間経過も、テール(tail )電流の単一指数関数回帰の結果から明らかなよう に、強し\膜電位依存性を示した(Figure 6A) 0 Figure 6B に示すように、 脱活性化の時 定数は22,9mV当たりe倍の増加を示した。
nifedipine非感受性Bdキ電流の速い不活性化と遅い不活性化のキネティクス
nifedipine 非感受性 Ba2+電流は、脱分極パルスの持続時間を20ms よりも長く設定 すると、単一の指数関数で表される不活性化を示した(一例として Figure 6B参照)。
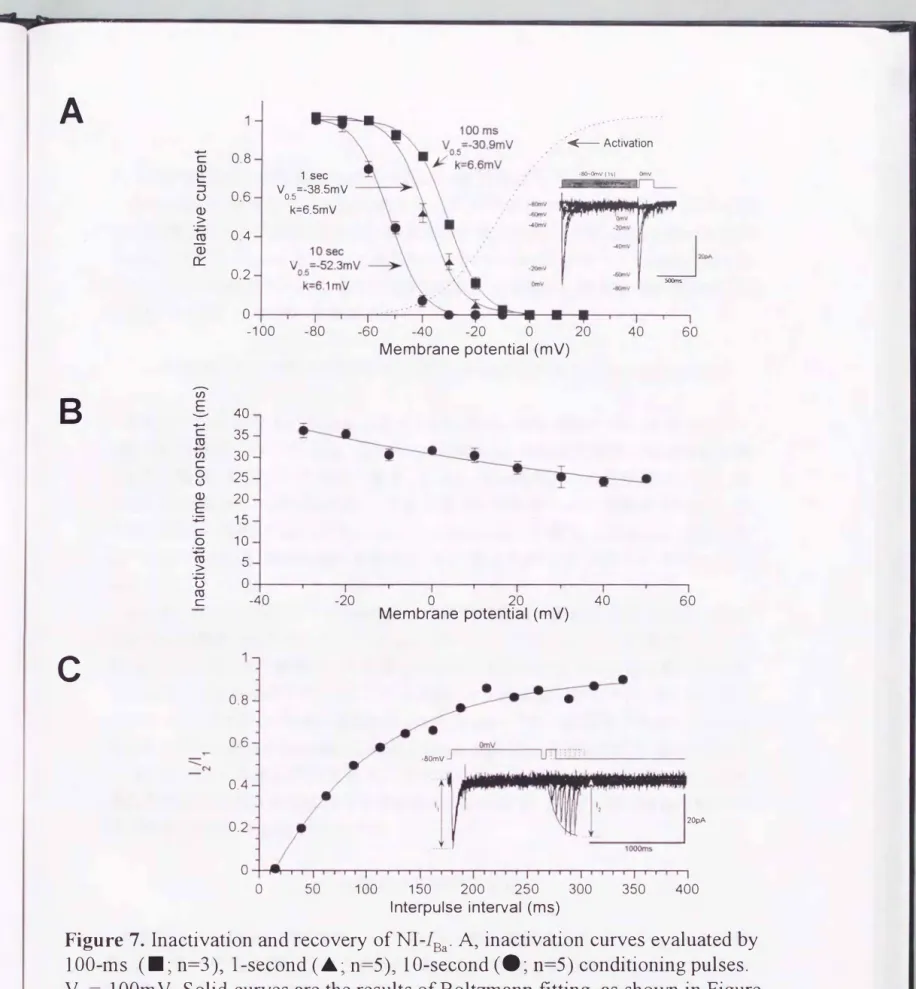
この時間経過は、前述したようにCa2+とBa2+で、差がなく(Figure3 Ab)、 Ca2+依存性の不 活性化によるものではないと考えられた。-10mV より脱分極側では、100ms の脱分極 パルスによる完全な不活性化が見られ(Figure 7 A)、+20mVでの時定数は約30ms で あったOさらに、 この時定数は比較的広い膜電位の範囲で変化がなく(102 ,lmV 当た りe 分のーの滅少� Figure 7B)、顕著な電位依存性を示す活性化や脱活性化の時定 数と比べて対照的で、あった(Figure 5B、6B)。
Figure 7A は、異なる3つの長さの条件脱分極パルスCI00ms、ls、10s)によって得ら れたnifedipine非感受性 Ba2+電流の不活性 化曲線を示している。図から明らかなよう に 、パノレス時間を長くするほど不活性化曲線が過分極方向へシフトした。Boltzm ann 回帰によって求めた500/0不活性化電位は100ms、ls、10sに対して各々-30,9 、 -38,5、
-52,3mVで、あった。このことは秒単位の長い時間経過で進行して行くもう一つの不活 性化状態の存在を示唆している。このようなnifedipine非感受性Ba2+電流の比較的速 くて完全な不活性化は、nifedipine感受性Ba2+電流の不完全で、ゆっくりとした不活性
化とは好対照で、あった。
nifedipine非感受性Ba2+電流の活性化曲線と不活性化曲線を重ね合わせてみると、
電流が恒常的に流れる領域(し\わゆるwin d ow current )が存在すること がわか る (Figure 7 A の実線及び点線)。注目すべきことに 、 この範囲はおよそる0--30mVにあ
J m
4←- Activation 命nV ー叙〉ーQmV (1',
至宝E主要望�
1 sec V =-38.5mV
0.5 k=6.5mV 0.8
0.6
0.4
0.2
日cω」」コυω〉一戸町一
ω区
A
60 40
-60 -40 -20 0 20
Membrane potential (mV)
-80 O
-100
40 35 30 25 20 15 10
(ωE)MCEωcoυωEZCO一芯〉一ぢ何「」
B
40 60
o 20
Membrane potential (mV)
ー20 5
O -40
-80mV
Lf 1.1臥弘且品世...】L"II...uJ..I...J....,l,...u..・.u...J...lo.I.J企.IJ.L�
I I \f iIi I 20pA
0.8
C
. /
戸/ /
0.6
0.4
一\
0.2
400 350
150 200 250 300
Interpulse interval (ms)
50 100 G
O
Figure 7. Inactivation and recovery ofNI古a. A, inactivation curves evaluated by 100-ms
(・;
n=3)ヲl-second (..; n=5)ョ10-second(・;
n=5) conditioning pulses.VHニー100mV. Solid curves are the results ofBoltzmann fittingヲas shown in Figure 41egend. Bフinactivation time constant obtained by single exponential fitting is plo抗ed against the membrane potential. Data are mean + SEM企om 3 to 5 different cells,
and the curve is fitted by a single exponential白nction (solid curve). Cヲrecoveη time course evaluated at -80m V. 11 and 12 denote出e amplitude ofNI-1Ba at Om V at the frrst pulse (10 seconds) and those during recovery with interpulses of various durationヲrespectively. The time course of recovery is fi抗ed by a single exponentia1 (function)
17
り、生理的膜電位の変動範囲とほぼ一致した(考察参照) 。
nifedipine非感受性Ba2+電流不活性化の大部分(約900/0)は、数百ms以内で完 全に回復した。その時間経過 は単一指数関数で表わされ、その時定数は四80mVで約 100msで、あった(Figure 7C)。 一方残りの約10%の回復は、秒オーダーの時定数を持
つもう一つの回復相を示した。最初の速い回復は、nifedipine感受性Ba2+電流と比較 するとかなり短かった(データ未提示)。
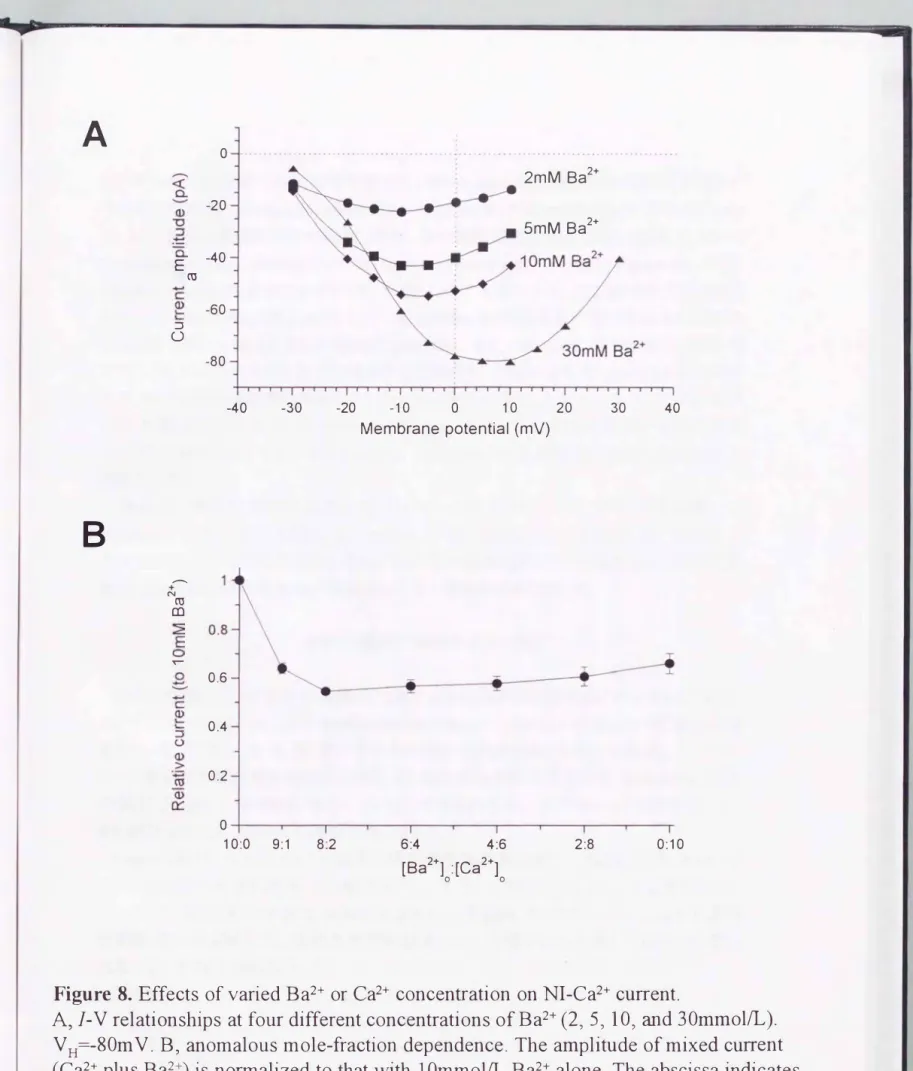
二価無機イオンの濃度増加による効果とanomalous mole-fr•αction dependence
潅流液中のBa2+やCa2+濃度を上昇させると、 電流一電圧関係の活性化関値やピー ク値の脱分極側へのシフトが起こると共に、nifedipine非感受性電流の電流振幅が増
大した(Figure 8 )。例えばBa2+の場合、2 から 30mmolILへの濃度変化により、 約 20mV脱分極側へシフトがみられた。Ca2+の場合も同程度のシフトが観察された が、 最
大電流振幅の増加はBa2+よりも小さかった(30mmollLの場合、nifedipine非感受性 Ca2+電流の振幅は、nifedipine非感受性Ba2+電流の約112で、あった(0.5 6+0.1 1;
n=5))。
Ba2+及びCa2+の混在下で、のnifedipine非感受性電流の振幅は、Ba2+とCa2+の濃度 比に対して複雑な依存性を示した(Fi忠lre 8B)。 すなわちBa2+と Ca2+濃度の合計を 10mmolILに固定 して濃度比のみ を変化させると、Ba2+:Ca2+が8:2の時に電流値は最 小となった。 にもかかわらず、これらのこ価陽イオンが単独で存在するときには、Ba2+
の方が Ca2+よりも約50%高い透過性を示した(Figure 8B の右端 及び左端)。 このよう な一見矛盾した現象はanomalousmole-合action depen dence と呼ばれ22 、後述するよう に、 他のタイプの高電位活性化型Ca2+チャネルに共通した性質で、あり、 チャネノレの透 過孔内の複数の結合部位に対する透過性イオンCBa2+及び Ca2+)問の複雑な相互作 用の結果であると考えられている2山。
無機及び有機的阻害薬
本実験のnifedipine非感受性Ca2+チャネルと他の電位依存性Ca2+チャネルとの類 似点、相違点 をさらに明らかにするため、幾つかの既知の電位依存性 Ca2+チャネル
阻害薬の効果について検討した。 Figure 9Aは、 5mmolILBa2+ を電荷担体とした 時の nifedipine非感受性Ba2+電流に対するCd2+及びNi2+の濃度抑制曲線を示す。Hill解
析により求めたCd2+及びNi2+の500/0 抑制値はそれぞれ5.8 及び68μmoVLで、あったO このことはCd2+の方が約10倍、nifedipine非感受性Ca2+チャネノレに対する阻害活性
が高いこと を示している。
また別の一連の実験 においては、nifedipine以外の汎用されている幾つかのL型
A
〈CL
てー@ E
ココ L E 悶 喝
LLc ω 回--d 仁ココ
B
N + ∞ ( 国
2 CE 〉
て-
、+。
、::::.- +
qLLc d
--J ιコJ
E
】申 正s aJ ,
。
会\
2mM Ba2+-20
、\、
_ 5mM Ba2+-40
-60
-80
i
i i I i i i I i i i i i I i i i i I i i i i I -40 -30 -20��1\
0.6
0.4
0.2
。
10:0 9:1 8:2
-10 。 10
Membrane potential (mV)
6:4 4:6
[Ba2+] 。: [Ca2+] 。
20 30 40
γ
2:8 0:10
Figure 8. Effects of varied Ba2+ or Ca2+ concenむation on N1-Ca2+ current.
A,1圃V relationships at four different concentrations of Ba2+ (2ヲ5ヲ10ヲ組d 30mmollL). VH=-80mV. Bヲanomalous mole-fraction dependence. The amplitude of mixed current (Ca2+ plus Ba2+) is normalized to伽t with 10mmolIL Ba2+ alone. The abscissa indicates the molar ratio of Ba2+ to Ca2+ヲkeeping the sum of Ca2+ and Ba2+ concentrations at
10mmol/L. The number of cells tested was 5.
19
Ca2+チャネノレ阻害薬の抑制効果を調べたo dihydropyridine と構造的に異なるL型Ca2+
チャネノレ阻害薬 、phenylalkylamine系のverapamil 及びbenzothiazepine系のdiltiazem は、Ba2+電流を濃度依存的に抑制したが、 最大抑制濃度である100μmol/Lにおいて もnifedipine以上の抑制効果を示さなかった(Turkeytest でP>0.05; Figure 9 )。一方、
dihydropyridine化合物でありながら T型Ca2+チャネノレに対しても 抑制作用が報告さ れているnimodipine (10 μmoVL)は25、nifedipine非感受性Ba2+電流を約60%抑制し た(独立2群のt 検定によりP<O.Ol; Figure 9B )。また、 5mmol/LのFを含む内液を用 いてL型Ca2+チャネル を完全 に減衰させた場合も、10μmol/Lのnimodipine は残存 する Ba2+電流を同程度に 抑制した(データ未提示)。また、もう一つのT型Ca2+電流 の阻害薬として知られている amiloride も、この nifedipine 非感受性Ba2+電流を濃度 依存的に抑制したが(IC50=107μmol/L) 、2mmol/Lの高濃度でも完全な 抑制は見ら れなかったO
最後に、種々の高電位活性化型Ca2+チャネルに対するベフ。チド性 阻害薬(ω- conotoxin GVIA及びMVIIC 、ω-agato幻n IV A)の効果について検討した。Fig.9Bに 示すように、これらの阻害薬は、 それぞれの最大阻害活性を示す濃度でも、腸間膜動 脈のnifedipine非感受性Ba2+電流対して 全く効果を示さなかったO
逆転写遺伝子増幅法(RT-PCR)
前述の結果から、本研究の対象と してきたnifedipine非感受性Ca2.,.チャネルと“R型"
Ca2+チャネルには、幾つかの重要な類似点があるこ とが明らかになった(詳細は考察 参照)。そこで我々は、R 型Ca2+チャネルの分子実体と考えられているαlEサブユニ ットが腸間膜動脈最終分|岐部に発現しているか否か を検討するため、nifedipine感受 性成分(Pigure2参照)に 対応していると考えられるαlCサフcユニット を 対照として 、 RT-PCR法 によるmRNAの検索 をおこなったO
Figure 10 は、モルモットの小脳及び腸間膜動脈 最終分枝から抽出した全RNA に 対して おこなったRT-PCR法の結果を示している。小脳からは、 αlC、αlEサブ、ユニッ トし1ずれに対してもそのPCR産物が検出された( Figure 10の Cレーン)。しかし腸間 膜動脈最終分枝からは、電気生理学的検索によって僅かしか発現していない と思わ れるαlC しか検出されなかったO
A
M
1 M -一 o 同 hu v Rd v'
e川山「C〆。 / k - o
m日n/aC !〉へγ@、/ \ ふ け り 、
社八
、\
弘 、
-muエ
/
•
Cd2+
IC50=5.8J..lM n=1.2
•
0.8
0.6 0.4
】Cω」」コUω〉一日同一ω広
0.2
0.1
Figure 9. Pharmacology ofN1-1Ba. All data were evaluated at Olll V. Aラ concentration-inhibition curves for Cd2+ヲNi2+フand amilorideヲwhich are fitted by the Hill equation shown in Figure 2B. For Cd2+ and Ni2+ヲ出e value of
r
is set to O. V H=-60m V. Bヲsummary of inhibition by various blockers (labeled). V H=・80m V.Data are means+
SEM 企om 3 to 13different cells. CTx indicates ω-conotoxin; Agaヲω-agatoxin.
21
0.01
三ュFO=>28Xトue 三ュF〈一〉O,xト0・
0.001
EュOOF
10・5 0.0001
Concentration (M)
2ュFEωNE--一℃
三ュOOF
EュOF
10・6
三ュ←-FC伺己巴
ω〉
三ユOFωC一色一刀OE-
c
10・7
三ュo 三ユFωc一色一万ω』=」
0 10・8
0.6 0.4
0 .
2。
0.8
戸C ω」」コUω〉一戸町一ω区
B
4-2. L型および新型nifedipine非感受性高電位活性化型Ca2+チャネノレ(/) 加齢による変化
全Bcl+電流に対するnifedipine非感受性Bd+電流の割合の加齢による変化
Figure llAは、腸間膜動脈最終分校部の平滑筋細胞から記録した全Ba2+電流に対 するnifedipine非感受性Ba2+電流の割合を週令別にフ。ロットしたもので、あるOこの図よ り、週令が増すにつれ 全Ba2+電流に対するnifedipine非感受性Ba2+電流の割合が減 少することが示唆された。一方、中枢側の分校では、 加齢による nifedipine 非感受性 Ba2+電流の割合変化は僅かで、あった。
nifedipine感受性(L 型)、 および非感受性Bcr+電流密度の加齢による変化
Figure llBは、nifedipine感受性Ba2+電流( L型Ba2+電流)及びnifedipine非感受 性 Ba2+電流を細胞膜容量により電流密度に換算し、週令ごとにその平均値を まとめた ものである。この結果から、L型Ca2+チャネノレの電流密度は2 週令から20週令にか け、急激に増加することが分かる(約3倍)。一方、nifedipine非感受性Ba2+電流は減 少傾向を示したが、L 型Ca2+チャネノレ増加の程度と比較すると軽度で、あった(約半 分)。またこれに対応して、前者の後者に対する比は、約1:2 から2:1へと逆転した。
以上のことから、FigurellAにおける全 Ba2+電流に対する nifedipine非感受性成分 の割合の週令増加に伴う減少は主にL型Ca2+チャネル密度の増加によるもので、ある ことが示唆された。
nifedipine非感受性Cclキチャネノレの生理学的および薬理学的特性の 加齢による変化
次に、nifedipine 非感受性Ca2+チャネルの生理及び薬理学的性質が加齢と共に変 化しているか否かを、生後3�5 週、 及び18-20週齢の2 群のモルモットの腸間膜動 脈最終分枝を用いて比較検討した。それぞれの群において、500/0活性化電位は- 11.0mV 及び-11.3mV、50%不活性化電位は申52.3mV及び-50.7mVとほとんど差がな く、 また薬理学的特性(Cd2+、Ni2+及び a milori deの 50%阻害値)に有意な差が認めら れなかった(Table2)。このことから、nifedipine非感受性Ca2+チャネルの特性は加齢 によってほとんど、影響を受けないことが示唆された。
α1C α1E
1k bp 500 bp
C MA C MA
Figure 10. RT圃PCR detection ofVDCCαlC組dαlE subunit mRNA in guinea pig cerebellum and terminal branches of guinea pig mesenteric arteries. Shown訂e the results of nested PCR using
lμg total RNA. Le釦nost lane in each panel shows a 100国bp ladder. C indicates cerebellum; 1.ιl\, mesenteric紅白ry.
23
4・3. 新型nifedipine非感受性高電位活性化型Cti+チャネノレに対する P2Y受容体を介したATPによる制御
新型nifedipine非感受性Cdキチャネルに及ぼすATPの効果
濯流液に 10μmol/Lnifedipineを添加した5mmolILBa2+pSSを用い、この組織の静 止膜電位で、ある-60mVからOmVへの脱分極パルスを与えると、Figure12A に示す Ba2+電流が観察されたOこの電流に対し 100μmollLおよび10mmolILATPを投与す ると、電流振幅の増加と 減少がそれぞれ観察された。 この増強 および抑制効果は、
ATP濃度のlμmol/Lから 10mmol/L の範囲に亘って観察されたCFigure12 A, B )。
細胞外液中に 5mmolILBa2+を添加した条件下で、は、nifedipine非感受性Ca2+電流 の振幅は数10pA程度であり、最大約200/0程度の ATP による増強効果を評価する には、電流とノイズ、の差が少なく定量性に問題があると考えられた。そこで以下の実験 では、二価イオン濃度をマイクロモーラー(μmoνL)以下まで減少させ、Na+ が Ca2+
チャネルを透過で、きる条件を設定して実験をおこなった。また、二価イオン濃度を減少 させることによって、ATP の二価イオンに対する直接捕捉効果を減じ、遊離型 ATPの 効果を観察することも期待できた。
新型nifedipine非感受性Cdキチャネノレ由来のNa+電流
濯流液のCa2TおよびMg2+を除去し、10μmol/Lのnifedipineを添加した 1 40mmol/L Na+の条件下で、 Na+電流を観察すると、2mmolILCa2+潅流時の最大振幅時の電流値 に比して約80倍の電流が観察された。このNa+電流は TTX 10μmol/L に耐性を示 し、10μmol/LのCd勺こより完全に抑制された(ICso= 0.33μmoVL)。また、このNa+電 流はnimodipine 10μmol/Lで部分的抑制を受けた(約45% )。さらに、細胞外のCa2+
濃度を0.015μmol/L、1.75μmol/Lと上昇させるとNa+電流の約10%、40%の抑制が 観察され、100μmol/L で完全に抑制された。その後ミリモーラー(mmollL)の濃度ま で上昇させるとCa2+電流が観察された(データ未提示 )。以上の結果からこのNa+電流 は新型nifedipine非感受性Ca2+チャネル由来であることが強く示唆された。
Na+電流に及ぼすATPの効果
この新型 nifedipine非感受性 Ca2+チャネル由来の Na+電流 に対し、細胞外より 1 nmollL-1 mmollLのATPを作用させると、Figure 14Aに 示す様に増強、および抑制 の二相性効果が観察された。増強効果は 1nmollL-10μmol/Lの範囲で、また抑制効 果は 100μmol/L -10mmollLの範囲で観察された。さらにマイクロモーラー(μmollL)
ト、3 1ノ、
Table 1. Summary ofDHP-insensitiveラrapidly-inactivating Ca2+ current
U1E
R-type rbE-II doe-1
actJvatlOn VU1 -50--40mV -50mV -10mV
[-40mV] 1・40mV]
VO.5 -2mV -29.1mV . [-16mV]
τ叫(10・90%) 0.98ms .. 2ms 12ms [5ms]
mactlvahon VO.5 -62,・68rnV -78.1mV -46mV
[-59mV]
30ms .. 99.6ms 't:inact
[152rns]
deactivation τdeact 0.21ms #
permeability Ca2+くBa 2+ .
Ni2+ IC50=66μM 28μM 33μM
Cd 2+ IC 50= 1 μM >80%(10ドM) -0%(1ドM)
ω-CTx-GVIA (1-5μM) NE NE slight reduction
(5μM)
ω-Aga IVA (100nM) NE 33% (200nM) NE (200nM)
[IC 50=51 n問
ω-CTx MVIIC (1-5ドM) NE NE
nimodipine (10μM) NE
amiloride . 16.4%(lmM)
ionic conditions (mM) 5 or 10Ba 4Ba 40Ba
[10Ba] [5Ba]
references 6,26,41 28,30 6,41
.
BII human はIE
-20mV -40mV
. +3.5mV
1.3ms -33.9mV -71mV
82ms* 25ms
0.294m�
Ca2+くB 2+
140μM 27.4μM 0.86μ恥f 0.8μM
NE 13%
NE -10%
NE 18%
NE .
.
40Ba 15Ba
32 38
T-type -70-・60mV
-27mV [-40mV]
2ms ..
-53, -63mV [-80m V]
-20ms ..
[-20ms“]
1.61 ms再 Ca2+=Ba 2+
90-100%
(く100ドM) 2か50μM NE NE NE
63%
IC 50 =30-1.55μM
5 or 10Ba [20Ca]
1,8,24,26
αlG
(CavT.l) -60mV
-21mV [-38mV]
-50mV
.
-
40Ba [10Ba]
25
αlH
(CavT.2 this study -70mV -50mV -44mV -11.0mV -2ms泳 4.0ms -75.3m丸 -52.3mV -20msホ 27.6ms ..
-4rns # 1.8ms # Ca 2+<Ba 2+
68μM 5.81lM NE(lドM) NE (100nM) NE(1μM)
. 36% (10μM)
75% (lmM) (ICq)=107μM)
10Ba 5Ba
5
濃度の ATP はこのNa+電流の電流-電圧曲線を偏位させたが、その電流の最大振 幅量には影響を与えなかった(Figure13A, B)。この時テール電流解析により活性化 曲線を描くと、50%活性化電位が約10mV負電位側にシフトしていることが分かった (control及び ATP1μmolJL投与; )1頃に Vos=-3 3.1mV, -4 2.8mV)。一方、不活性化 曲線には何ら変化がみられなかった(データ未提示 )。これらの結果からマイクロモー ラー(μmol/L)濃度のATPによるNa+電流の増強効果は主に Ca2+チャネルの活性化 を促進することによるもので、あることが示唆された。
purine、pyrimidineおよびP2受容体詰抗薬の効果
ATP 以外のヌクレオチドの効果に ついても検討した。UDP、 お よ び UTPは、
1nmolIL寸00μmollLの範囲では全くこのNa+電流に影響を与えなかった(データ未 提示)。しかしADPは ATPと同等の増強および抑制作用を示し、ATPyS はATPより も低い濃度範囲(1nmol/L-l OOnmol/Lで増強、100nmoù包-10μmollLで抑制 )で効 果を示した。また 2 -MeSATPは1nmollL-10μmoνLの範囲で抑制効果のみを示した (Figure 14A)。一方、P2受容体括抗薬、 suramin (100μmol江)、 PPADS ( 3 0μmollL) の前投与によって、ATP による増強効果、抑制効果はともにほぼ完全に消失したが、
P 2X受容体の選択的な搭抗薬α,ß-methyleneATPは無効で、あった。
以上の結果よりATPの増強及び抑制効果は P2Y受容体を介したものであることが示 唆された。
G蛋白質の関与
P2Y受容体は、G蛋白質共役型の受容体であることが知られている。そこで以下の実 験では、P2Y受容体と共役している G蛋白質やその下流の情報伝達系を明らかにす るため、G蛋白のキネティクスに影響を与える薬物や種々の蛋白リン酸化酵素阻害薬
/活性化薬の効果について検討した。
まず、電極内にGDPßS 500μmolJLを添加し、G蛋白質の代謝回転を抑制すると、
ATP (1 nmol/L-1 mmol/L )による nifedipine非感受性Na+電流の増強・抑制効果はほ ぼ完全に消失した。次に、 phospholipase C (PLC)阻害薬の U-73122、 proteinkinase C ( PKC)阻害薬の PKC inhibitor peptide (PKC-I)を用いて、この電流に対する影響を観
察した。
U-73122は以前から種々のチャネノレに対する非特異的な作用が指摘されてきたが、
我々の結果でも 1μmol江の 投与により、上記の nifedipine非感受性Na+電流を約 65010抑制した(データ未提示 )。従って、U-73122はこの実験においては適正なツール として使用できないことが示唆された。
A
B
100寸
,注、,-r.、、、
J
、
--ー、、∞問 80
4E。1コ3 60
、、 、ロ3 ∞ 何
Z 、、_...
ー」田E ロ山
20 LJ_
。
2.5 イ
lム
〈EζL L 4U
〉- d3 、
1.5
C ω てコ
場 cω LL 剛
-
- d
。コ
。
P<0.01 Pく0.01
② /
2-5
. 岨』 ・ .
10-13
Age (w)
nifedipine-insensitive Ba2+ current ーーーー
I
nifedipine-sensitive Ba2+ current
2-5 10-13
Age (w)
e
16-20
*, # P<0.05 16-20
Figure 11. Changes ofNS-lBa and N1-1Ba by ageing. All data were
evaluated by a depolarizing pulse from
-60
to Om V. Aヲthe fraction ofNIlBa wi白respect to globallBa is plotted against the age. Closed circles represent the data from terminal branches
(5th, 6th)釦d
open circles山 proximal branches(2ベ3
rd)
of mesenteric arterial tree. Bヲcu汀ent densities ofNSーらa and N1-1Ba with民spect to the postnatal age27
一方 PKC -I(150μmol/L)を細胞内投与すると、ATPによる抑制効果は有意に減少 したが、増強効果はほとんど、影響を受けなかった(Figure14B )。さらにPKC の活性化 薬である phorbol-12,13・estel ( PDBu ) 250nmol/L を投与するとそれ自体によって抑制 効果があらわれ、その後に ATP1mmolILを追加投与しても更なる抑制効果は観察さ れなかった(データ未提示)。以上のことより、ATPによるnifedipine非感受性Na+電 流の抑制効果は、PKCに関与するG蛋白質を介した反応であることが示 唆された。
一方、細胞膜透過型のcyclic AMP (cAMP )のアナログ、で、あるdi butylylcyclic AMP ( db-c AMP)や細胞膜透過型のprotein kin ase A阻害ペプチド( PKA心を用い、その 効果を観察した。細胞外よりdb-cAMP (1 mmollL)を投与すると、Na"-電流は増強され、
その後のlμMATP投与によっても更なる増強効果は観察されなかった(データ未提 示)。また、PKA-I(41.3nmollL)の投与後、ATP(10μmollL� lmmollL)を累積投与し た場合、抑制効果のみが観察された(Figure 14B)。これらの結果は、ATPによる増強 効果が、主にPKAに関連したG蛋白質を介したものであることを示唆している。
最後に PKA-Iと PKC-Iを同時に 投与すると ATP(lμmol/L-1mmollL)による増強 および抑制効果は完全に消失した (データ未提示)。以上の結果を単純に解釈すると、
腸間膜動脈のnifedipine非感受性Ca2+チャネノレに対する ATPの増強効果には Gs 蛋白質/PKA系が、また抑制効果にはGq/11蛋白質/PLC/PKC 系が重要な役割を 果たしていると考えることができる。
A
B
Activation Inactivation Age(w)
I
VO.5 k VO.5 k
3-5
I
-11.0mV -11.3mV 国52.3mV 6.1mV18-20 -9.4mV
I
-11.1 mVI
-50.7mV 4.9mVAge(w) Cd2+(IC50) Ni2+(IC50) lamiloride(IC50)
3-5 5.8μM 68μM 107μM
18-20 5.6μM 63μM 124μM
Table 2. Biophysical (A) and pharmacological (B)阿ameters of NI-IBa at 1\νo different ages
Data indicate mean values obtained from 5 to 10 different cells.
29