息環境の変化
著者
上野 綾子, 佐藤 正典, 山本 智子
雑誌名
Nature of Kagoshima
巻
40
ページ
217-223
別言語のタイトル
Changes in macrobenthic fauna and
environmental condition of their habitats of
the Shigetomi-higata tidal flats in Kagoshima
Bay
鹿児島湾の重富干潟における底生生物相及びその生息環境の変化
上野綾子
1・佐藤正典
2・山本智子
3 1〒 890–0056 鹿児島市下荒田 4–50–20 鹿児島大学大学院水産学研究科 2〒 890–0065 鹿児島市郡元 1–21–35 鹿児島大学大学院理工学研究科 3〒 890–0056 鹿児島市荒田 4–50–20 鹿児島大学水産学部 はじめに 干潟とは川から運ばれた砂泥が堆積してできた 地形で,波浪が少なく潮の満ち引きが大きい内湾 などに形成される.そこに棲むさまざまな種の動 物が,有機物の除去や生態系内の物質循環に関し て重要なはたらきを担っており,干潟のエコトー ンとしての機能を支えている(菊池,1993).し かし,1940 年代から 90 年代にかけて干拓や埋め 立てなどで日本の約 40% の干潟が消失し(環境 省自然保護局,1994),干潟生物のもつこのよう な機能の劣化が懸念される. 底生生物相が変化する要因としては,主に干潟 における富栄養化やそれに伴う貧酸素化,餌の減 少等,底質中の環境や水質に関わるものだけでな く,河川や沿岸の流れがもたらす地形や堆積環境 の変化など物理的な要因も考えられる.後者に関 する研究は多くないが,鹿児島湾奥の干潟におい ては,1977 年から 2003 年にかけて,面積が約 200 ha から約 60 ha に減少しており(山本・小玉, 2009),地形の変化に伴う環境変化が底生生物相 に影響を与えている事が予測されている. 調査地である鹿児島県姶良市平松の重富干潟 は,思川の河口,南北 1.5 km にわたって広がる 河口・前浜干潟が組み合わさって形成されており, 鹿児島湾内の干潟では最大のものである.しかし, 重富干潟でも周囲の海岸線の埋め立てが進んでお り,干潟の環境変化が著しいと考えられる.そこ で,松下(1995)および桝屋(2006)と今回の調 査(2012)との比較から,底生生物相と生息環境 の変化を明らかにすることを目的とした. 調査方法 底生生物相の調査及び統計解析 調査は鹿児島湾奥の重富干潟にて,2012 年 7 月 20 日および 21 日の大潮干潮時におこなった (Fig. 1). 山本ほか(2009)を参考にし,干潟上に海岸 線とほぼ垂直に 3 本のラインを引き,1994 年 3, 4 月,2005 年 5 月の調査ラインを再現した.底生 生物の採集地点は,2005 年の調査ステーション の位置をできるだけ再現する事とし,Line 1 では 20 m おきに,Line 2 では 25 m おきに,Line 3 で は 30 m おきに,それぞれ等間隔に設置した.各Ueno, R., M. Sato and T. Yamamoto. 2014. Changes in macrobenthic fauna and environmental condition of their habitats of the Shigetomi-higata tidal flats in Kagoshima Bay. Nature of Kagoshima 40: 217–223.
TY: Faculty of Fisheries, Kagoshima University, 4–50–20 Shimoarata, Kagoshima 890–0056, Japan (e-mail:
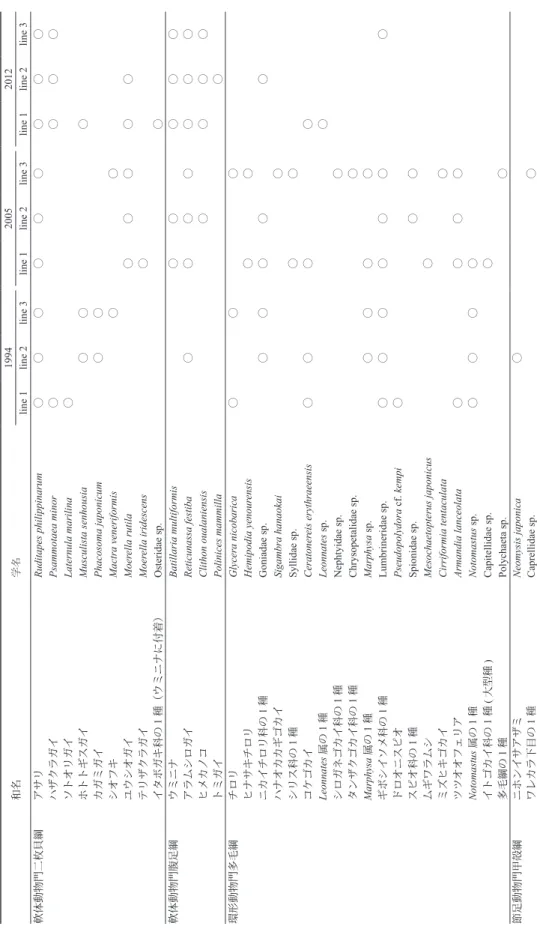
和名 学名 1994 2005 2012 line 1 line 2 line 3 line 1 line 2 line 3 line 1 line 2 line 3 軟体動物門二枚貝綱 アサリ Ruditapes philippinarum 〇 〇 〇 〇 〇 〇 〇 〇 〇 ハザクラガイ Psammotaea minor 〇 〇 〇 〇 ソトオリガイ Laternula marilina 〇 ホトトギスガイ Musculista senhousia 〇 〇 〇 カガミガイ Phacosoma japonicum 〇 〇 シオフキ Mactra veneriformis 〇 〇 ユウシオガイ Moer ella rutila 〇 〇 〇 〇 〇 テリザクラガイ Moer ella iridescens 〇 イタボガキ科の 1種(ウミニナに付着) Osteridae sp. 〇 軟体動物門腹足綱 ウミニナ Batillaria multiformis 〇 〇 〇 〇 〇 アラムシロガイ Reticunassa festiba 〇 〇 〇 〇 〇 〇 〇 ヒメカノコ Clithon oualaniensis 〇 〇 〇 〇 トミガイ Polinices mammilla 〇 環形動物門多毛綱 チロリ Glycera nicobarica 〇 〇 〇 ヒナサキチロリ Hemipodia yenour ensis 〇 〇 ニカイチロリ科の 1種 Goniadae sp. 〇 〇 〇 〇 〇 ハナオカカギゴカイ Sigambra hanaokai 〇 シリス科の 1種 Syllidae sp. 〇 〇 コケゴカイ Ceratoner eis erythraeensis 〇 〇 〇 〇 Leonnates 属の 1種 Leonnates sp. 〇 シロガネゴカイ科の 1種 Nephtyidae sp. 〇 タンザクゴカイ科の 1種 Chrysopetalidae sp. 〇 Marphysa 属の 1種 Marphysa sp. 〇 〇 〇 〇 ギボシイソメ科の 1種 Lumbrineridae sp. 〇 〇 〇 〇 〇 〇 〇 ドロオニスピオ Pseudopolydora cf. kempi 〇 スピオ科の 1種 Spionidae sp. 〇 〇 ムギワラムシ Mesochaetopterus japonicus 〇 ミズヒキゴカイ Cirriformia tentaculata 〇 ツツオオフェリア Armandia lanceolata 〇 〇 〇 〇 Notomastus 属の 1種 Notomastus sp. 〇 〇 〇 〇 イトゴカイ科の 1種 (大型種 ) Capitellidae sp. 〇 多毛綱の 1種 Polychaeta sp. 〇 節足動物門甲殻綱 ニホンイサアザミ Neomysis japonica 〇 ワレカラ下目の 1 種 Caprellidae sp. 〇 Table 1. 採集された底生生物の種リスト.
端脚目の 1 種 1 Amphipoda sp. 1 〇 〇 端脚目の 1 種 2 Amphipoda sp. 2 〇 クーマ目の 1 種 Cumacea sp. 〇 〇 〇 モエビ科の 1 種 Hippolytidae sp. 〇 エビジャコ類似種群 * Crangon affinis complex 〇 ニホンスナモグリ ** Callianassa japonica 〇 〇 〇 〇 〇 マメコブシガニ Philyra pisum 〇 〇 〇 ケフサイソガニ Hemigrapsus penicillatus 〇 ヒメアシハラガニ Helice japonica 〇 ホンヤドカリ科の 1 種 Paguridae sp. 〇 〇 〇 〇 〇 テナガツノヤドカリ Diogenes nitidimanus 〇 紐形動物門 ヒモムシ類の 1 種 Nemertinea sp. 〇 〇 刺胞動物門花虫綱 ムシモドキイソギンチャク科の 1 種 Edwardsiidae sp. 〇 〇 ナマコ綱 ヒモイカリナマコ Patinapta ooplax 〇 〇 ステーション上に直径 17 cm の塩ビパイプ(コア) を 3 個ずつ設置し,コアを泥中 10 cm まで挿し込 み,その中の堆積物を 1 mm メッシュの篩で篩っ て残った生物を回収した. 底生生物群集が年ごとにどのように変移して いるかを調べるために,非多次元尺度構成法 (nMDS)を用いた群集解析を行った.今回の解 析には,各ラインで採集された底生生物各種の 1 平方メートルあたりの個体数(個体数/ 1m2)を 使用し,ライン間の非類似度指数を以下の式に よって計算した.また,1994 年,2005 年,2102 年でそれぞれ最干潮位が異なるため,2102 年に 設置した一番沖側のステーションより高いものの み選択した. 非類似度指数の算出方法は以下の式の通りで ある. 上記の式では,S は非類似度指数, は群集 1 の i 番目の個体数, は群集 2 の i 番目の個体 数とする. 環境解析 底質環境については,ステーションごとに干 潟表面 2 cm までの泥を内径 29 mm のコアサンプ ラーで採集し,有機物含有量と粒度組成を解析し た.有機物含有量は,120℃・12 時間の乾燥後と, 電気炉で 650℃・2 時間燃焼後の重量を秤量し, 減量分を重量パーセントで示した.また粒度組成 は,採集した泥を 120℃・12 時間乾燥後,1 mm, 0.063 mm の電動篩を用いて,10 分ずつふるった. その後,0.063 mm 以下の粒子の重量割合である シルトクレイ率と 1 mm 以上粒度の割合を算出し た.ただし、1994 年のサンプルは,水中で水が 濁らなくなるまでふるい,篩に残った泥を乾燥さ せ重量を計測した.分析方法の違いが結果に影響 しないことは確認済みである(山本ほか,2009). なお,泥の採集は原則として生物の調査時に行っ たが,有機物含有量の算出に用いた 1994 年のサ ンプルについては,12 月 14 日に採集した.
結果と考察 1994 年に出現した生物種数は全体で腹足類 1 種,二枚貝類 6 種,甲殻類 5 種,多毛類 8 種の計 20 種であったが,2005 年には腹足類 3 種,二枚 貝類 4 種,甲殻類 9 種,多毛類 17 種,その他 2 種, 全部で 35 種に増加した.2012 年には,腹足類 4 種, 二枚貝類 5 種,甲殻類 3 種,多毛類 4 種,その他 1 種で計 17 種と再び減少した(Table 1). 1994 年,2005 年,2012 年の各ステーションに おける底生生物の個体数を,Fig. 2–4 に示す.ス テーションを設置した場所は年によって異なる が,縦にほぼ同じ場所で採集したデータが並ぶよ うに配置した.ライン 1 では,2005 年に腹足類 が増加することで全体の個体数も増加し,その後 2012 年にはさらに増加した.ライン 2,3 では, 2005 年に多毛類が増加して全体の個体数が増加 したが,2012 年には再び減少している. 1 m2あ た り の 平 均 個 体 数 は,1994 年 に は 231.17(個体数/ m2)であったが,2005 年には 433.05(個体数/ m2)と増加した.しかし,種 組成別にみると,二枚貝が減少し,多毛類と腹足 類が増加していた(Fig. 5).2012 年には 108.36(個 体数/ m2)と大幅に減少し,平均個体数密度は 種数と同じ変動パターンを示した.特に二枚貝類 は 1994 年から 2005 年の変化よりさらに減少して いた. 1994 年 の 優 占 種 は ア サ リ(Ruditapes philippinarum)であり,2005 年では多毛類のツツ オフェリア(Armandia lanceolata)であった.また, 2012 年は腹足綱ウミニナ(Batillaria multiformis) であり,これは 2005 年では全体の約 20% であっ たのに対し 2012 年には 75% で,7 年の間に急激 に増加していた(Fig. 5). 1994 年には出現していたが 2005 年には見られ なかった種として,ホトトギスガイ(Musculista senhousia)やハザクラガイ(Psammotaea minor), Fig. 2. Line 1 における生物群集の変化.縦軸は 1 m2あたり の個体数を,横軸は基点からの距離をそれぞれ表す.また, 年によってステーション位置(m)が異なるため,棒グ ラフが表示されていない地点は,その年ステーションを 設置していないことを示す. Fig. 3. Line 2 における生物構造の変化.縦軸は 1 m2あたり の個体数を,横軸は基点からの距離をそれぞれ表す.また, 年によってステーション位置(m)が異なるため,棒グ ラフが表示されていない地点は,その年ステーションを 設置していないことを示す.
カガミガイ(Phacosoma japonicum)などの二枚 貝が挙げられる.その一方で,1994 年に出現し なかったが 2005 年に出現した種として,ウミニ ナ,ヒメカノコ(Clithon oualaniensis)などの腹 足類であった(Table 1).また,山本ほか(2009) によると,アサリの 1 個体あたりの重量が減少し, 多毛類に限っては大型種が減り小型種の増加して いる事が解っている.2005 年には出現し 2012 年 には出現しなかった種は,ツツオオフェリアや Marphysa 属の 1 種(Marphysa sp.)ムギワラムシ (Mesochaetopterus japonicus)など多くの多毛類で あった.一方で,1994 年と 2005 年には出現しな かったが 2012 年に出現した種として,肉食性の トミガイ(Polinices mammilla)が挙げられる(Table 1). 各ラインの生物群集を nMDS により解析した ところ,二次平面上では 1994 年,2005 年,2012 年, それぞれの年代ごとでプロットが分かれた(Fig. 6).重富干潟の底生生物群集構造は,1994 年か ら 2005 年,2012 年と継続してして変化している と考えられる.具体的には,1994 年から 2005 年 にかけて種数・個体数は増加していたが,埋在性 である二枚貝類の減少や多毛類の小型種の増加が 進む一方で,表在性の腹足類ウミニナは増加して いた.また,2005 年から 2012 年にかけては種数・ 個体数も大幅に減少,特に埋在性である二枚貝, 多毛類が多く減少し,腹足類ウミニナが優占種と なるほど増加していた(Fig. 5). 生物群集における変移の要因として,有機物 含有量,高度,粒度の変化から考察を行った. 強熱減量から算出した有機物含有量は,Line 1 Fig. 4. Line 3 における生物群集の変化.縦軸は 1 m2あたり の個体数を,横軸は基点からの距離をそれぞれ表す.また, 年によってステーション位置(m)が異なるため,棒グ ラフが表示されていない地点は,その年ステーションを 設置していないことを示す. Fig. 6. ラインごとの生物群集の非多次元尺度法(nMDS)の 結果. Fig. 5. 各ラインの生物相の年変化 .
で 2004 年から 2012 年の間に干潟上部で 2% から 10% に変化していたことを除けば,1–2% の範囲 で変動しており,大きな変化は見られなかった (Fig. 7).この事から,生物群集変化の要因として, 富栄養化や貧酸素は考えにくい. しかし,シルトクレイ率では大きな変化がみ られた(Fig. 8).Line 1 において,1994 年にほと ん ど の ス テ ー シ ョ ン で 30% 以 上 で あ っ た が, 2005 年には 10% 以下となり,2012 年にもほぼ同 じ値を示した.Line 2 では,1994 年から 2005 年 にかけて全体で 10% ほど低下していたが,2012 年には上昇している地点もみられた.Line 3 は中 部で 1994 年から 2005 年に低下していたが,2012 年にかけてはあまり変化がみられなかった. 一方で,1 mm 以上の粒子の割合については, 1994 年のデータが得られなかったため,2005 年 と 2012 年の比較を行った(Fig. 9).Line 1 にお いては,上部から中部にかけて 10–15% ほど上昇 していた.また,Line 2 と Line 3 では 2012 年の 上部から下部にかけて変動があるものの,全体的 Fig. 7. ラインごとの強熱減量.縦軸は強熱で減少した割合, 横軸は基点からの距離を表す. Fig. 8. ラインごとのシルトクレイ率変化.縦軸はシルトク レイ率,横軸は基点からの距離を表す. Fig. 9. ラインごとの 1 mm 以上の粒子率.縦軸はサンプル全 体量における直径 1 mm 以上の粒子の割合,横軸は基点 からの距離を表す.
に上昇していることが分かった. これらのことから,シルトクレイ率の変動と 粒度の大型化は著しく,この環境変化が生物群集 構造に大きな影響を及ぼしていると考えられる. 粒度の大型化は,何らかの原因で波や河川などの 流れが変化し,細かい砂粒が流された結果とも考 えられる.流れの変化は干潟全体の地形変化もも たらしている可能性があるため,今後は,生物相 の調査を継続すると共に,リモートセンシングな ど新たな手法を応用して,干潟の地形変化を詳し く調査する必要がある. 謝辞 甲殻類の種同定にあたりご指導・ご助言下さっ た, 鈴 木 廣 志 教 授( 鹿 児 島 大 学 水 産 学 部 ), ArcGIS 解析に関してご協力頂いた鶴成悦久氏(鹿 児島大学水産学研究科),ならびに調査の際にご 協力下さった NPO 法人くすの木自然館の皆様に この場を借りて厚く御礼申し上げる.最後に,野 外調査にご協力下さった 2012 年度鹿児島大学海 洋センター生物多様性研究室の先輩・同輩の皆様 に深く感謝する. 引用文献 環境省自然保護局.1994.海域生物環境調査報告書(干潟, 藻場,サンゴ礁調査).財団法人海中公園センター,東 京.291 pp. 菊池泰二.1993.干潟生態系の特性とその環境保全の意義. 日本生態学会誌,43: 223–235. 松下耕治.1995.鹿児島湾の重富海岸および喜入海岸にお ける干潟底生動物の分布.平成 6 年度鹿児島大学理学 部卒業論文.119 pp. 桝屋 藍.2006.重富海岸における干潟底生動物相の変化 (1994–2005).平成 17 年度鹿児島大学水産学部卒業論 文.11 pp. 山本智子・小玉敬興.2009.過去 60 年間における鹿児島湾 奥の海岸線の変.Nature of Kagoshima, 35: 55–57. 山本智子・桝屋 藍・松下耕治・佐藤正典.2009.鹿児島 湾の重富干潟における底生動物相の変化 — 1994 年と 2005 年の比較 —.ベントス学会,64: 32–44.