博 士 学 位 論 文
酸化ストレスがもたらす皮膚恒常性機能障害と
高安定型アスコルビン酸誘導体の改善作用
に関する研究
博 士 学 位 論 文
酸化ストレスがもたらす皮膚恒常性機能障害と
高安定型アスコルビン酸誘導体の改善作用
に関する研究
平成 29 年 1 月 10 日
勝山 雄志
目 次 序論・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・1 第 1 章. 細胞内抗酸化作用の亢進における PPAR-γと Nrf2 の関連・・・・・・・・・・・・・7 1-1. 諸言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・7 1-2. 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・7 1-2-1. PPAR-γ agonist による細胞内抗酸化成分産生亢進作用・・・・・・・・・・・・・・7 1-2-2. PPAR-γ および Nrf2 の関連性・・・・・・・・・・・・・・・・・・・・・・・・9 1-3. 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・10 第 2 章. 酸化ストレスによる皮膚バリア, 感覚刺激への影響・・・・・・・・・・・・・・・12 2-1. 諸言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・12 2-2. 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・12 2-2-1. 酸化ストレスによる SPT mRNA 発現への影響・・・・・・・・・・・・・・・・12 2-2-2. 酸化ストレスおよび IL-1α による感覚刺激への影響・・・・・・・・・・・・・・13 2-2-3. IL-1α による感覚刺激への影響・・・・・・・・・・・・・・・・・・・・・・・14 2-2-4. 乾燥による感覚刺激への影響・・・・・・・・・・・・・・・・・・・・・・・・15 2-2-5. NHEKs および乾燥皮膚モデル培養上清による P12 細胞の樹状突起伸長・・・・・16 2-3. 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・17 第 3 章. 新規アスコルビン酸誘導体:3-O-グリセリルアスコルビン酸の安定性評価・・・・・20 3-1. 諸言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・20 3-2. 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・21 3-2-1. 3GA の原体および製剤安定性・・・・・・・・・・・・・・・・・・・・・・・・21 3-2-2. 3GA の化学的な ROS 消去作用・・・・・・・・・・・・・・・・・・・・・・・22 3-2-3. 3GA の細胞内 ROS 消去作用・・・・・・・・・・・・・・・・・・・・・・・・23 3-2-4. 3GA の細胞傷害緩和・・・・・・・・・・・・・・・・・・・・・・・・・・・・24 3-2-5. 再構築表皮モデルを用いた細胞内 ROS 産生抑制・・・・・・・・・・・・・・・25
3-3. 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・26 第 4 章. 3GA 誘導体 (3LGA)の抗酸化機能とそのメカニズムについて・・・・・・・・・・・29 4-1. 諸言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・29 4-2. 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・29 4-2-1. 3GA 誘導体の細胞内抗酸化活性・・・・・・・・・・・・・・・・・・・・・・・29 4-2-2. 3LGA の化学的な ROS 消去作用・・・・・・・・・・・・・・・・・・・・・・ 30 4-2-3. 3LGA の細胞内 ROS 消去作用・・・・・・・・・・・・・・・・・・・・・・・ 31 4-2-4. 3LGA の細胞傷害緩和・・・・・・・・・・・・・・・・・・・・・・・・・・・32 4-2-5. 3LGA の細胞内抗酸化成分産生促進・・・・・・・・・・・・・・・・・・・・・33 4-2-6. 3LGA の PPAR-γ-Nrf2 シグナル活性化・・・・・・・・・・・・・・・・・・・・34 4-3. 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・37 第 5 章. 3LGA の皮膚恒常性機能維持効果・・・・・・・・・・・・・・・・・・・・・・・39 5-1. 諸言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・39 5-2. 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・39 5-2-1. 3LGA の皮膚バリア機能に対する恒常性維持効果・・・・・・・・・・・・・・・39 5-2-2. 3LGA の敏感肌誘導に対する恒常性維持効果・・・・・・・・・・・・・・・・・42 5-3. 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・46 第 6 章. 総括・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・48 実験の部・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・50 参考文献・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・67 謝辞・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・75
略語一覧
ROS: reactive oxygen species
SPT: serine palmitoyltransferase
SCF: stem cell factor
ET-1: endothelin-1
IL-1α: interleukin-1α
PGE2: prostaglandin E2
α-MSH: α-melanocyte-stimulating hormone EGF-R: epidermal growth factor-receptor
MMP-1: matrix metalloprotease-1
MMP-9: matrix metalloprotease-9
NGF: nerve growth factor
Sema3A: semaphorine 3A
TEWL:transepidermal water loss
CPs: carbonylated proteins
Nrf2: nuclear factor E2-related factor 2
Keap1: kelch-like ECH-associated protein 1
ARE: antioxidant response element
GSH: glutathione
HO-1: heme oxygenase-1
GST: glutathione S transferase
NQO1: NAD(P)H quinone oxidoreductase-1
MRP: multidrug resistance-associated proteins
AsA: L-ascorbic acid
PPAR-α: peroxisome proliferator-activated receptor-α
PPAR-γ: peroxisome proliferator-activated receptor-γ
NHEKs: normal human epidermal keratinocytes
HBSS: hanks buffered saline solution
siPPAR-γ: PPAR-γ-small interfering RNA γ-GCS: γ-glutamyl cysteine synthetase
GAPDH: glyceraldehyde 3-phosphate dehydrogenase
RXR: retinoid X receptor
PPRE: PPAR response element
NF-κB: nuclear factor- κB
COX-2: cyclooxygenase-2
RHEEs: reconstructed human epidermal equivalents
HaCaT: immortalized human keratinocytes
3GA: 3-O-glyceryl ascorbate
3LGA: 3-O-laurylglyceryl ascorbate
HPTLC: high performance thin layer chromatography
1 序論 皮膚の構造と機能 皮膚は人体の最外層に存在し, 体内の様々な器官を外部刺激や異物から保護することで, 生体内 機能の恒常性を維持する重要な組織である. また, 皮膚は人の外見を直接的に反映するため, 多く の女性が皮膚を健やかに保つことで, いつもまでも若々しく美しくありたい, 見られたいと願い, それをスキンケアにより実現することができる器官でもある.
皮膚組織は最外部から, 表皮層(Epidermal layer), 真皮層(Dermis layer), 皮下組織(Subcutaneous tissue)に大別される. 表皮は, 角層(Stratum corneum), 顆粒層(Stratum granulosum), 有棘層(Stratum spinosum), 基 底 層 (Stratum basale) に よ り 構 成 さ れ , 真 皮 は , 乳 頭 層 (Stratum papillare), 網 状 層 (Stratum reticulare)に分けられる(Fig. 1). 表皮は皮膚の最外層にあり生体内外の境界膜として生体を 保護し, 角層に適正な水分を保持することにより柔軟性を発揮している1-3) . 表皮を含めた角層の機能として, 外部からの異物の侵入を防ぎ内部からの水分の蒸散を抑制す るバリア機能がある. このバリ ア機能は, 角層細胞間に存在す る角層細胞間脂質のラメラ構造 体と顆粒層に存在するタイトジ ャンクションにより発揮され る. 真皮は, コラーゲン線維やエ ラスチン線維により構築される 細胞外マトリックスにより基本 骨格が形成され, ヒアルロン酸 に代表されるグルコサミノグリ カンがプロテオグリカンとして その間隙を満たしている. また,
2 これら真皮構成成分を合成する線維芽細胞がマトリックス内に散在している. 酸化ストレスが関与する皮膚障害 皮膚構成細胞機能は様々な要因によって低下し, それが原因となって様々な皮膚障害が生じる. 皮膚障害の要因は, 主には内因性因子と外因性因子に大別される. 内因性因子としては主に加齢が 挙げられる. 加齢に伴う皮膚組織の変化を生理的老化と呼ぶが, その変化の速度は緩やかである. 皮膚機能の急激な低下は, 主に外因性因子によって引き起こされる. なかでも, 近年, その主要因 として考えられているのが, 太陽光線に含まれる紫外線である. 紫外線の慢性的な曝露により生じ る皮膚老化を, 光老化と呼ぶ. 光老化は, 皮膚内部において紫外線曝露により過剰生成された活性 酸素種 (Reactive Oxygen Species: ROS)が引き金となって進行することが知られており, 色素斑や深 い直線性のシワの形成など生理的老化とは異なる皮膚の外観変化を示す 4)
. また, ROS により誘発 される生体傷害を酸化ストレスと呼ぶ. ROS がイニシエーターとなる酸化ストレスにより進行する 皮膚老化のメカニズムは, これまでの研究から明らかにされている.
老化に伴う色素斑としては, 老人性色素斑がある. 老人性色素斑部位では, Stem cell factor (SCF)/c-kit シグナルおよび Endothelin-1 (ET-1) シグナルによりメラニン合成が促進していることが 報告されている5)
. ET-1 の分泌亢進は, Interleukin-1α (IL-1α)により誘導され, UVB 照射により分泌 亢進される IL-1α は, ROS により分泌が促進される6). また, UVB によるメラニン色素合成は, ROS
により産生亢進される Prostaglandin E2 (PGE2) や α-Melanocyte-stimulating hormone (α-MSH)により
制御されている7, 8). さらに, 活性窒素種(Reactive Nirogen Species: RNS)である Nitric oxygen (NO・)
も, メラノサイト内の Tyrosinase の合成をグアニルシクラーゼ系の活性化を介して亢進することが 知られている9) . 以上のことから, ROS や RNS が老人性色素斑形成を含めたメラニン合成に関与し ている. シワの形成は真皮細胞外マトリックスの変化に由来し, 特にコラーゲン線維の減少とエラスチ ン細線維であるオキシタラン線維の消失が要因と考えられている 10, 11) . シワ形成への ROS の関与 についても以下のように明らかにされている. コラーゲン線維の減少は, ROS による Protein tyrosine phosphatase の不活性化に伴う Epidermal growth factor- receptor (EGF-R)の活性化状態の維持
3
と, EGF-R の活性化による c-Jun N-terminal kinases (JNK)シグナルによるコラーゲン分解能の亢進と コラーゲン合成能の低下により説明される. JNK の活性化は, c-Jun および c-Fos の合成を促進し, そ の両者のヘテロダイマーである Activated protein-1 (AP-1) が, Matrix metallo-proteinase-1 (MMP-1) およびコラーゲン合成に対する抑制タンパク質 CCN1/cyr61 の転写活性の亢進を誘導する.
また, ROS は, 脂質過酸化物やカルボニル化タンパク質(CPs)の生成を誘導することによって生体 内タンパク質や酵素活性を変性, 失活させる11)
. さらに, ROS が直接的に DNA に攻撃することによ って, DNA 鎖の切断等が生じ, DNA 障害がもたらされる. このような DNA 障害は, 上述の皮膚老化 シグナルに影響を及ぼすだけでなく, 皮膚癌などの重大疾患の要因とされている. 酸化ストレスと表皮バリア機能の関連性 皮膚バリア機能は, 角層細胞間脂質のラメラ構造体が角質細胞間に充填されることで発揮され, 外部異物の生体内流入や内部からの水の蒸散を防ぐ. 角層細胞間脂質は, 主にセラミド, コレステ ロール, 遊離脂肪酸, コレステロール硫酸から構成され, なかでもセラミドは, 角層細胞間脂質の 大半を占める成分であるため, 皮膚バリア能を顕著に反映する1-3) . これは, 表皮バリア機能が脆弱 な, アトピー性皮膚炎や老人性乾皮症の角層において, セラミド量が顕著に減少していることから も, セラミドの皮膚バリア機能への寄与が推察される 12-14) . セラミドは, スフィンゴシン塩基と脂 肪酸がアミド結合した両親媒性脂質であり, 両者の組み合わせの違いや炭素鎖の異なるものを含 めると, 342 種類ものセラミドが存在するといわれている 15) . セラミドは, セリンとパルミトイル CoA を出発物質として, セラミド合成律速酵素である Serine palmitoyltransferase (SPT)の触媒により 3-ケトスフィンガニンが合成される段階を律速とした de novo 合成経路で合成される16). 合成され たセラミドは糖修飾(グルコシルセラミド)またはリン酸化(スフィンゴミエリン)された後, 顆粒 層内の層板顆粒に貯蔵される. その後, 角層への分化過程または外部刺激によって層板顆粒が放出 され, 層板顆粒内に同様に貯蔵されている β-glucocerebrosidase や sphingomyelinase によって再びセ ラミドに変換されることで, 角層ラメラ構造体を形成する17) . セラミド量は加齢に伴い減少することが知られている. 一方で, 加齢者の経表皮水分蒸散量 (Transepidermal water loss : TEWL)は若者の TEWL と同程度である. しかしながら, 加齢者の表皮バ
4 リア機能はセラミド量の減少に依存して低下しており, この機能低下を補償するために角層総数 を増加させることが知られている12-14, 18) . UVB 照射した正常ヒト表皮角化細胞では, SPT mRNA 発 現が低下することが報告されており 19) , セラミド合成能の低下に由来して表皮バリア機能の低下 をもたらす. このセラミド減少の要因として, 加齢や紫外線曝露によって表皮細胞内で生成される ROS がセラミド合成経路へ影響を及ぼしている可能性が考えられる. 表皮バリア機能の減少と感覚刺激感受性との関係 表皮バリア機能が低下することによって, 様々な皮膚障害が生じる. そのうちの1つに, 感覚刺 激に対する感受性の亢進, 即ち「敏感肌」の誘導が挙げられる. 敏感肌は, 全世界の約 70%の人々 が敏感肌発症の潜在性を有しているため20) , 現代社会の皮膚トラブルの 1 つであるともいわれてい る. このような状態では, サンスクリーン剤を含む化粧品等を肌に塗布した際に掻痒感や痛み, ツ ッパリ感などの自覚症状を日常的に感じるため, Quality of life (QOL) が著しく低下する.
敏感肌の特徴の 1 つとして, 肌の乾燥状態が知られている. 敏感肌は, 健常肌と比較して TEWL が高く, 乾燥状態が進行するほどセラミド量が低下することが知られている21, 22) . セラミド量の低 下による表皮バリア機能の減少が, 外部刺激に対する感受性向上を誘導するものと推察される. ま た, 肌の乾燥が進行する冬期において, 角層中の CPs の存在量が増加することも報告されている23) . CPs は脂質過酸化反応の最終生成物であることから, 冬期の乾燥性皮膚では ROS の過剰亢進によ って酸化ストレスが生じ, 敏感肌形成に何らかの影響を及ぼしている可能性が考えられる. 感覚刺激は感覚神経である c-線維終末の興奮により引き起こされる. 乾燥性皮膚における感覚 刺激の感受性亢進は, 外部刺激を感知する c-線維の終末が表皮上層まで伸長していることが要因 として考えられている. 一方, 健常肌では, c-線維の伸長は真皮層の基底膜直下で留まっている. c- 線 維 の 伸 長 は , 軸 索 ガ イ ダ ン ス 因 子 で あ る , Nerve growth factor (NGF: 神 経 成 長 因 子 ) と Semaphorine 3A (Sema3A: 軸索反発因子)のバランスによって制御されている 24). これら因子は IL-1α との関連性が報告されており, バリア機能を人為的に破壊した皮膚では, IL-1α の産生が惹起 され, NGF の発現亢進と c-線維の表皮内への伸長が確認されている25) . さらに, IL-1α は ROS 生成を 誘導することから, 酸化ストレスが c-線維の表皮内への伸長に関与する可能性が考えられる. この ことから, 酸化ストレスが感覚刺激の感受性亢進を誘導する可能性が推察される.
5
生体内抗酸化システムの発現制御機構
ROS による酸化傷害に対し, 生体内では独自の ROS に対する防御機構「生体内抗酸化システム」 を備えることで皮膚機能の恒常性を維持している. 細胞内には ROS 消去作用を発揮する複数の抗 酸化ペプチド, タンパクが存在し, これらは酸化ストレスに応答して発現が制御されている. この 生体内抗酸化システムは, Nuclear factor E2-related factor-2(Nrf2) - Kelch-like ECH-associated protein 1 (Keap1)シグナルによって制御されていることが良く知られている26). 定常状態において, Nrf2 は細 胞質に局在し, アクチン結合性因子である Keap1 と相互作用することで不活性化され, プロテアソ ーム系によって恒常的に分解を受けている. 細胞は, 酸化ストレスに曝されることにより Keap1 の レドックスセンサーである SH 基が ROS や RNS 等の作用によって酸化され, Nrf2 は Keap1 から脱 離し, 核内に移行する. Nrf2 は, 複数の抗酸化系因子の遺伝子の上流に存在する Antioxidant response element (ARE) に結合することで, これら因子の発現を亢進する27). Nrf2 によって制御され ている抗酸化系因子としては, グルタチオン(GSH)や Heme oxygenase-1(HO-1) などの抗酸化タン パク質, Glutathione S transferase (GST) や NAD(P)H quinone oxidoreductase-1(NQO1) などの第Ⅱ相解 毒酵素, Multidrug resistance-associated proteins (MRP) などのトランスポーターが知られている27-30)
. このように Nrf2-keap1 シグナルは, 皮膚機能恒常性維持のためには非常に重要なシステムであるが, 近年 Nrf2-keap1 シグナルをその上流で制御している因子も存在しているとの報告がなされており 31) , 本シグナル経路の全容解明には至っていない. また, 生体内抗酸化物質である Catalase や GSH は加齢や紫外線曝露により減少していくことが報 告されている 32, 33) . この事実は, 生体内の抗酸化システムを維持, 向上させることが, 皮膚老化の 予防において極めて重要であることを示唆している. 本研究が解決しようとする課題 本研究は, 酸化ストレスに対する生体内の防御機構を詳細に検証するとともに, 酸化ストレスと 皮膚障害の関連性を明らかにすること, さらに酸化ストレスにより惹起される皮膚障害の予防, 改 善に対する対策を提案することを目的として実施した.
6
ペルオキシソーム増殖活性化受容体である Peroxisome proliferator-activated receptor-γ (PPAR-γ) に ついては, 近年, 生体内抗酸化システムに寄与する可能性が報告されている 31) . そこで, 第 1 章で は, 生体内抗酸化システムの詳細なメカニズムの解明を行うことを目的とし, PPAR-γ に着目して 生体内抗酸化システムである Nrf2-keap1 シグナルとの関連を明らかにするための検証を行った34) . 第 2 章では, 酸化ストレスがおよぼす皮膚障害への影響について, 特に皮膚機能の恒常性維持に 最も重要な皮膚バリア機能に着目して, その酸化ストレスと皮膚バリア機能の関連性を検証した 35) . また, 現代社会の皮膚トラブルの 1 つである敏感肌について, 感覚刺激に対する感受性亢進と 酸化ストレスとの関連性について検証することで, その誘導メカニズムを考察した. 上記検証を行った結果, ROS による皮膚障害の予防, 改善には抗酸化剤の使用が有効であるとい う結論に至った. そこで我々は, 種々ある抗酸化剤のなかから, アスコルビン酸(AsA)に着目し, 皮膚恒常性機能 維持効果について検証を行った. AsA は, ROS 消去作用やメラニン産生抑制作用をはじめ, 多彩な 生理活性効果を有しており36, 37) , 安全性も高いことから, 健康サプリメントや化粧品として多くの 人に親しまれている成分である. しかし, AsA は化粧品製剤に配合した場合の安定性の低さが課題 とされている. そこで, 第 3 章では, AsA の最大の欠点である安定性の低さを改善するため, 化粧品 の汎用素材であるグリセリンを修飾した AsA 誘導体, 3-O-グリセリルアスコルビン酸(3GA)の安定 性について, 化合物および製剤安定性の観点から評価し, さらに AsA 由来の生理活性を確認するた め, AsA の基本的な効果である抗酸化作用について評価を行った38) . 第 4 章では, 第 3 章の結果をも とに, グリセリルアスコルビン酸の生理活性をさらに向上させるために疎水基を導入した誘導体 について抗酸化評価を行い, ラウリル基を導入した 3-O-ラウリルグリセリルアスコルビン酸 (3LGA)に最も高い抗酸化活性を有することを見出し, その抗酸化の作用メカニズムについて検証 した34) . さらに第 5 章では, 安定性および生理活性機能を向上させた 3LGA の酸化ストレスによっ て生じる皮膚障害に対する改善効果について検証を行った35) . 以上の結果を総合的に考察することにより, 酸化ストレスにより惹起される皮膚恒常性機能低 下は, 新規 AsA 誘導体であるグリセリルアスコルビン酸により改善されることが明らかとなった.
7
また, グリセリルアスコルビン酸誘導体による生体内抗酸化システムの活性化作用は新たな知見 であり, 本研究の成果が人類の QOL の向上に寄与することを期待する.
8 本 論 第 1 章. 細胞内抗酸化作用の亢進における PPAR-γ と Nrf2 の関連 1-1. 諸言 生体内では外的ストレスや異物から身を守るために, 様々な防御機構が存在している. なかでも, Nrf2-keap1 シグナルを中心とする生体内抗酸化システムは, 生体内の酸化還元状態を制御すること で皮膚機能恒常性を維持する, 重要な防御機構である. PPAR-γ は, 主に脂質合成や表皮分化に関与する因子として知られているが39, 40), 近年, PPAR-γ が 細胞内抗酸化物質である GSH や Catalase の発現を亢進させるとの報告があり41) , 生体内抗酸化シ ステムへの関与が示唆されている. そこで, 本章では生体内抗酸化システム活性化メカニズムを詳細に検証するため, PPAR-γ と Nrf2 との関係性について検証した. 1-2. 結果 1-2-1. PPAR-γ agonist による細胞内抗酸化成分産生促進作用 PPAR-γ の生体内抗酸化システムへの関与を 検証するため, PPAR-γ agonist である GW192942) を用いて細胞内 ROS 消去に関する検証を行っ た. 本試験では, 主に H2O2 をはじめとする 種々の ROS と反応することにより蛍光を発す る DCFHDA を用いた 43) . まず, GW1929 を NHEKs に処理すると細胞内 ROS が有意に減少 することが確認された(Fig. 1-1) 44) .
Figure 1-1. Intracellular ROS suppression of NHEKs treated with or without GW1929.
Bars indicate means ± S.D. (n = 4). *p<0.05 indicates a significant difference against control.
9
先行研究から, PPAR-γ が Catalase や GSH の合成律速酵素である γ-Glutamyl cysteine synthetase (γ-GCS) の発現を亢進することが報告されており 41), 本結果と合わせて考察すると, GW1929 が細 胞内抗酸化物質の産生を促している可能性が推察された. そこで, NHEKs に GW1929 を処理し, 24 時間後の細胞内抗酸化成分の mRNA 発現を測定した. その結果, GW1929 処理によって, Catalase を はじめ, γ-GCS や Heme oxygenase-1 (HO-1), NAD(P)H quinone oxidoreductase-1 (NQO1) の mRNA 量 が有意に増加した(Fig. 1-2) 34), 44)
. GW1929 が複数の抗酸化成分の mRNA 産生量を増加させたことか ら, これら成分の転写制御因子である Nrf2 の mRNA 発現について確認した. 結果として, GW1929 処理によって, Nrf2 mRNA 量が有意に発現亢進することが確認された(Fig. 1-3) 44)
.
Figure 1-2. Stimulating effects of catalase, γ-GCS, HO-1and NQO1 mRNA expression by GW1929 in NHEKs. Bars express means ± S.D. (n = 4). **p<0.01 indicates a significant difference against
control.
Figure 1-3. Stimulating effects of Nrf2 mRNA expression by GW1929 in NHEKs.
10 1-2-2. PPAR-γ および Nrf2 の関連性
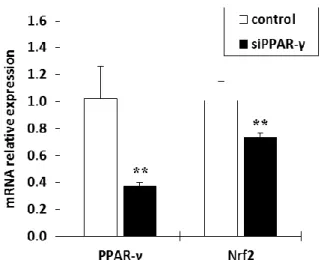
上記結果から, PPAR-γ が Nrf2 発現を制御している可能性が考えられたため, RNA 干渉法を用い て両因子の関連性を確認した. siPPAR-γ を処理した NHEKs では, PPAR-γ mRNA 発現が低下すると ともに, Nrf2 mRNA 発現も有意に低下することが確認された(Fig. 1-4) 34)
.
Figure 1-4. Effects of siPPAR-γ on the mRNA expression level of PPAR-γ and Nrf2 in NHEKs.
11
1-3. 考察
PPAR は, ペルオキシソーム増殖活性化の核内受容体として発見され, 3 つのサブタイプ(-α, -β, -γ) が存在する. PPAR は, そのリガンドが特異的ドメインに結合すると, 核内受容体の一種であるレチ ノイド X 受容体 (RXR) とヘテロ二量体を形成し, 標的遺伝子のプロモーター領域に存在する PPAR response element (PPRE) 配列に結合することで, 標的遺伝子の発現を制御する45). なかでも PPAR-γ は, 様々な組織に存在し, 抗動脈硬化作用や抗糖尿病作用, 抗ガン作用などの生理作用が知 られている45)
. 一方, 皮膚組織への PPAR-γ の作用についても報告されている. PPAR-γ のアクティ ベーターである ciglitazone を keratinocyte に処理すると, 表皮分化マーカーである Loricrin, Involucrin, Filaggrin の発現が亢進した. また, テープストリッピングまたはアセトン処理を行った ヘアレスマウスに ciglitazone を適用することで, 皮膚バリア機能の回復が促進され, 炎症反応が抑 制された40)
. さらに, 抗炎症作用については, PPAR-γ agonist が UVB 照射による Cyclooxygenase-2 (COX-2)や Prostaglandin E2 (PGE2)の発現亢進を抑制することも報告されている
46)
. PPAR-γ の抗炎症 メカニズムとして Nuclear factor-kappa B (NF-κB)の炎症性サイトカインへの DNA 結合を阻害するこ とが明らかにされている46)
. このように PPAR-γ は皮膚の恒常性維持に重要な役割を担っている. 近年の先行研究において, PPAR-γ が生体内抗酸化物質である Catalase や γ-GCS の発現を亢進する ことが報告されている 41)
. そのため, PPAR-γ が抗酸化作用にも寄与している可能性が示唆された. そこで, PPAR-γ agonist である GW1929 を用いて検証を行った. NHEKs に GW1929 を処理すると, 細 胞内 ROS 量が減少し(Fig. 1-1), 複数の抗酸化成分の mRNA の発現亢進が確認された(Fig. 1-2).
この結果から, PPAR-γ は細胞内抗酸化成分の産生亢進を介して抗酸化作用を発揮することが示 唆された. また, 複数の抗酸化成分の発現を亢進させたことから, その上流に存在する転写制御因 子である Nrf2 への関与も考えられた. そこで, Nrf2 mRNA 発現レベルを測定したこところ, GW1929 処理によって有意な亢進が認められた(Fig. 1-3). しかしながら, Nrf2 の発現亢進レベルは, 細胞内抗酸化成分の亢進レベルの程度と比較すると劣っていた. この理由としては, Catalase や γ-GCS のプロモーター領域に PPRE 領域が存在することが報告されており41) , 細胞内抗酸化成分は PPAR-γ による直接的な制御と Nrf2 による間接的な制御を受けている可能性が示唆された.
12
次に, PPAR-γ が Nrf2 を制御しているか否かを, RNA 干渉法を用いて検証した. siPPAR-γ を処理す ることによって, PPAR-γ mRNA とともに Nrf2 mRNA 発現も有意に低下した(Fig. 1-4). これまでの 先行研究から, PPAR-γ が Nrf2 と相互作用することや Nrf2 遺伝子のプロモーター領域に PPRE 領域 が存在することが報告されている31, 47) . そのため, PPAR-γ は Nrf2 の PPRE 領域に結合することによ り Nrf2-keap1 シグナルを制御していることが示唆された. 本研究の結果と先行研究から推察すると, PPAR-γ は, Nrf2-keap1 シグナルと細胞内抗酸化成分の 両者の発現を亢進することにより皮膚機能恒常性維持に寄与している可能性が示唆された(Fig. 1-5).
Figure 1-5. Proposed mechanism of intracellular antioxidation system by PPAR-γ contribution. Electrophiles/Nrf2 inducers: tBHQ, tert-butylhydroquinone; NEPPs, neurite
outgrowth-promoting prostaglandins.heme oxygenase-1 (HO-1), NAD(P)H-quinone oxidoreductase-1 (NQO1), glutamate-cysteine ligase (GCL), glutathione reductase (GR), sequestosome-1 (SQSTM1) and the cystine/glutamate anionic amino acid transporter (xCT).
13 第 2 章. 皮膚バリア機能, 感覚刺激に対する感受性向上への酸化ストレスの影響 2-1. 諸言 セラミドは, 角層細胞間脂質の主成分として皮膚バリア機能形成に寄与している. しかしながら, セラミドは, 加齢や慢性的な紫外線曝露によって減少し, 表皮バリア機能が低下することが知られ ている12-14) . このセラミド量の減少は, 高い TEWL を示し, 過度な水分蒸散により乾燥性皮膚の形 成を誘導する. 一方, 冬期に生じる乾燥性皮膚では皮膚の掻痒感を伴うケースが多く, 乾燥性皮膚 では感覚感受性の亢進が示唆されている. また, 感覚刺激感受性が亢進している, いわゆる「敏感 肌」では, 高い TEWL を示すことが報告されている21) . これらの事実から, 表皮バリア機能の低下 と敏感肌の形成の関連性が示唆される. これまでの先行研究から, セラミド量減少の要因である加齢や紫外線曝露によって, 皮膚内では ROS の産生量が増加することが明らかにされている 4). また, 肌の乾燥が進行する冬期において, 角層中の Interleukin-1 receptor antagonist (IL-1RA)/IL-1α 比の上昇や, 脂質過酸化反応の最終生成物 であるカルボニル化タンパク質(CPs)が増加することも報告されている 23) . さらに, 人工的に作製 した乾燥皮膚モデルにおいても, IL-1α や CPs の産生量が亢進することも報告された48) . これら事実 から考察すると, ROS がセラミド合成経路や乾燥性敏感肌の誘導に影響を及ぼしている可能性が考 えられた. そこで本章では, 過去の知見を基に, ROS と皮膚バリア機能および乾燥性敏感肌との関連性につ いて検証することとした. 2-2. 結果 2-2-1. 酸化ストレスによる SPT mRNA 発現への影響 酸化ストレスが及ぼす皮膚バリア機能への影響を, セラミド合成律速酵素である SPT を指標に 検証した. NHEKs に H2O2を 1 時間処理し, 6 時間培養後の SPT mRNA レベルを測定すると, H2O2濃 度依存的に SPT mRNA 発現が有意に低下した(Fig. 2-1) 35) .
14
Figure 2-1. Influence of H2O2 on the mRNA expression level of SPT in NHEKs.
Bars express means ± S.D. (n = 4). *p<0.05 and **p<0.01 indicate a significant difference against control.
2-2-2. 酸化ストレスによる感覚刺激への影響
酸化ストレスによる感覚刺激への影響を検証するため, NHEKs に H2O2または IL-1α 処理を行い,
敏感肌関連因子の発現レベルを確認した. まず, H2O2 処理を行った NHEKs では, NGF および
MMP-9 mRNA レベルが有意に亢進した(Fig. 2-2a). 加えて, H2O2処理よって IL-1α タンパク質につ
いても有意な増加が確認された(Fig. 2-2b) 44)
.
Figure 2-2. Influence of H2O2 on the (a) mRNA expression levels of NGF and MMP-9 and (b) IL-1α protein in NHEKs. Bars express means ± S.D. (n = 4). *p<0.05 and **p<0.01 indicate a significant
15 2-2-3. IL-1α による感覚刺激への影響
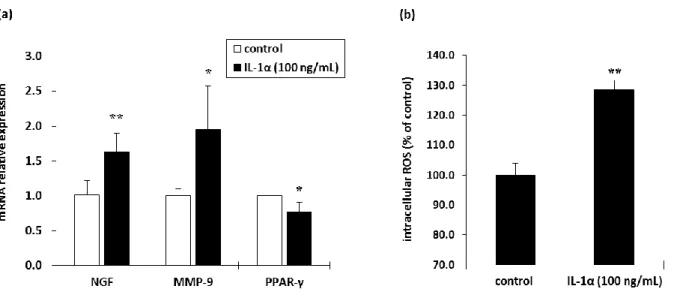
次に, IL-1α を NHEKs に処理した際の挙動を確認した. IL-1α 処理によって H2O2処理と同様に
NGF および MMP-9 mRNA の有意な発現亢進も確認された(Fig. 2-3a). また、細胞内 ROS レベルも 有意に亢進した(Fig. 2-3b). 一方で, IL-1α 処理によって, PPAR-γ mRNA 発現が有意に低下した(Fig. 2-3a). そこで, 次に PPAR-γ antagonist である BADGE49) を用いて, PPAR-γ と敏感肌との関連性を検 証した. BADGE を処理した NHEKs では, IL-1α タンパク質が有意に増加するとともに, NGF mRNA レベルも有意に亢進していることが確認された(Fig. 2-4) 44)
. この結果から, PPAR-γ が感覚刺激の感 受性亢進に影響を及ぼしている可能性が示唆された.
Figure 2-3. Influence of IL-1α on (a) the mRNA expression levels of NGF, MMP-9 and PPAR-γ and (b) the intracellular ROS level in NHEKs.
16
Figure 2-4. Influence of (a) IL-1α protein and (b) NGF mRNA expression by PPAR-γ antagonist (BADGE) in NHEKs. Bars express means ± S.D. (n = 4). *p<0.05 and **p<0.01 indicate a significant
difference against control.
2-2-4. 乾燥による感覚刺激への影響 先行研究において, 3 次元皮膚モデルから人工的に作製した乾燥皮膚では, 敏感肌誘導に関与す る IL-1α や CPs レベルの産生亢進が確認されており, 本モデルが乾燥肌研究に有用であることが 示唆されている 48) . そこで, 本研究においても 3 次元皮膚モデルを用いて乾燥皮膚を作成し, 敏 感肌に関与する因子の挙動を確認した. 乾燥処理を行った後 24 時間培養した皮膚モデルでは, 先
Figure 2-5. Influence of skin dryness on the (a) mRNA expression levels of NGF, MMP-9 and PPAR-γ, (b) IL-1α protein in reconstructed human epidermis equivalents.
17
行研究と同様に IL-1α の発現亢進が確認され(Fig. 2-5a, b), 本研究モデルが乾燥に伴う敏感肌誘導 メカニズムの検証に有用であることが改めて示唆された. また, 乾燥処理によって NGF や MMP-9 mRNA レベルの有意な亢進が確認された(Fig. 2-5a). 一方で, PPAR-γ mRNA 発現量は有意 に低下していた (Fig. 2-5a) 44) . 2-2-5. NHEKs および乾燥皮膚モデル培養上清による PC12 細胞の樹状突起伸長 酸化ストレス処理した NHEKs および乾燥処理した皮膚モデルを培養後, 培養上清を回収し(CM), ラット副腎髄質由来褐色細胞腫である PC12 細胞に CM を処理した. PC12 細胞は, 定常状態では球 状の様相を示すが, NGF によって分化シグナルが活性化されると樹状突起が伸長することから, 神 経モデル細胞として広く利用されている50) . 本試験においても, 定常状態において, PC12 細胞は球 状の様相を示していたのに対し, CM を 48 時間処理することで神経樹状突起の伸長が確認された (Fig. 2-6) 44).
Figure 2-6. Influence on the elongation of dendrite in PC12 cells induced by oxidative stress (a, b) or skin dryness (c, d). Detected by microscopy. (Bar scales: 25 μm)
18 2-3. 考察 セラミドは皮膚バリア機能形成に強く寄与している物質である. 皮膚バリア機能が脆弱なアト ピー性皮膚炎患者や, 過度な乾燥状態にある老人性乾皮症の角層では, セラミド量が減少している ことが報告されている 12-14) . また, 若者の角層と比較して, 加齢皮膚ではセラミド量が減少してい ることも確認されており18) , 皮膚バリア機能を維持するには, セラミド産生量の低下を抑制する必 要がある. 生体内でセラミドが合成されるまでには複数の酵素が関与している. なかでも, SPT は セラミド合成律速酵素であるため51) , SPT の発現がセラミド産生量に大きな影響を与える. そこで, ROS による皮膚バリア機能への影響を, SPT mRNA 発現を指標に検証した. NHEKs に H2O2を処理
することによって, SPT mRNA 発現の有意な低下が確認された(Fig. 2-1). この結果から, 酸化スト レスが表皮バリア機能の低下を惹起し, 乾燥性皮膚の形成を誘導することが明らかとなった. 先行 研究では, UVB 照射による SPT mRNA の発現低下が報告されている19) . UVB 照射は生体内におい て複数種の ROS 産生を亢進する. 本研究の結果と合わせて考察すると, UVB 照射によるセラミド 合成能の低下に ROS が関与していることが示唆された. 敏感肌は, 皮膚バリア機能や炎症の程度によって複数のタイプに区分される20) . そのなかでも共 通する特徴が, NGF の産生亢進とそれに伴う c-線維終末の表皮内への伸長である. アトピー性皮膚 炎や乾癬の表皮内では, 健常皮膚と比較して NGF の発現が亢進している24, 52) . また, アセトン処理 により表皮バリアが破壊された皮膚では, IL-1α 産生に伴う NGF 発現の亢進が確認されていること から23) , 皮膚の乾燥と NGF 産生には関連性があることが示唆される. また, CPs 頻度の高い角層を 持つ皮膚は, 皮膚表面水分量の減少と高い TEWL が報告されていることから23) , ROS が皮膚の乾燥 に関与している可能性が示唆される. そこで, 本研究では ROS と NGF 産生の関連性に着目して検証を行った. H2O2処理した NHEKs
において, NGF mRNA 発現の亢進が確認され(Fig. 2-2a), ROS による NGF 発現の誘導が示唆された. また, MMP-9 の発現亢進も確認された(Fig. 2-2a). MMP-9 は基底膜構成成分である collagenⅣや laminin332 等を分解する酵素である53). そのため, ROS による MMP-9 発現亢進によって基底膜構成 タンパク質の分解に伴う基底膜構造の破壊が, c-線維の表皮内への伸長を助長している可能性が示 唆された. 次に, ROS と IL-1α の関連性について確認すると, H2O2処理によって IL-1α 量が増加する
19
とともに(Fig. 2-2b), IL-1α 処理によっても細胞内 ROS 量が増加した(Fig. 2-3b). さらに, IL-1α 処理に よっても, NGF および MMP-9 mRNA 発現が亢進した(Fig. 2-3a). これら結果から, ROS と IL-1α は互 いに刺激しあいながら, NGF 発現を誘導しているものと考察される. また, 興味深い結果として, IL-1α 処理によって PPAR-γ mRNA 発現が有意に低下した(Fig. 2-3a). そこで, PPAR-γ と敏感肌との 関連性を検証すべく, PPAR-γ antagonist である BADGE を NHEKs に処理した. その結果, BADGE 濃 度依存的に IL-1α の産生量が増加し(Fig. 2-4a), NGF mRNA 発現の亢進が確認された(Fig. 2-4b). 第 1 章の結果や先行研究結果41) から, PPAR-γ は生体内抗酸化システムを制御する可能性が推察される.
それゆえ, IL-1α 発現亢進を介した生体内抗酸化システムの機能低下は, 更なる細胞内 ROS 量の増 大と NGF, MMP-9 発現亢進による敏感肌形成を誘導することが示唆された.
続いて, 肌の乾燥によっても敏感肌形成が誘導されるか否かについて乾燥皮膚モデルを用いて 検証したところ, 乾燥刺激によって, NGF および MMP-9 の mRNA, IL-1α の発現が有意に亢進し, 一 方で PPAR-γ mRNA 発現は有意に低下した(Fig. 2-5). 過去の先行研究においても, 人為的に作製し た乾燥皮膚では IL-1α の発現亢進が確認されており48, 54)
, 肌が乾燥状態に陥ることで IL-1α が惹起 され, それに伴って敏感肌が誘導される可能性が示唆された. また, 乾燥皮膚モデルの培養上清は 神経細胞の樹状突起の伸長を促進した(Fig. 2-6). 以上の結果から, ROS は皮膚バリア機能や敏感肌 形成の重要なトリガーであることが示唆された(Fig. 2-7).
20
Figure 2-7. Proposed mechanism of the skin barrier destruction and the sensitive skin development by ROS generation.
21 第 3 章. 新規アスコルビン酸誘導体:3-O-グリセリルアスコルビン酸の安定性評価 3-1. 諸言 AsA は熱や酸化, 光に対して非常に不安定であるため, 水溶液中では経時的に分解して褐変化が 生じる. さらに, 化粧品製剤に配合した際には, AsA 由来の強力なイオン性が原因となって, 製剤 中の乳化破壊による層分離や粘性製剤の減粘, 製剤の変色を生じさせるなど, 化粧品製剤の安定性 をも著しく低下させる. そこで上記課題を改善するため, 新規 AsA 誘導体の開発を行った55) . 具体的には, ①AsA の安定 性を高めること, ②AsA の水溶性を維持し, さらに製剤安定性に影響を及ぼさないこと, を課題と した. 誘導体化にあたって, AsA への導入基に化粧品の汎用素材で非イオン性の水溶性化合物であ るグリセリンに着目した. 高いプロトン解離性を示す AsA の 3 位水酸基56) にグリセリンを導入す ることで, 安定性を高め, 且つ水溶性を維持する分子設計を行った. 上記分子設計のもとに検討を 重ねた結果, 我々は新規な AsA 誘導体として 3-O-グリセリルアスコルビン酸 (3-O-Glyceryl Ascorbate:3GA) を合成した(Fig. 3-1) 55).
本章では, 3GA の安定性と化粧品製剤に配合した際の製剤安定性について評価することとした. 加えて, 3GA が AsA 由来の生理活性を有しているか否かについて, 抗酸化効果を指標に検証した. AsA は他の抗酸化物質と比較しても優れた抗酸化効果を有しており, AsA が有する生理活性のなか でも特に特徴的な機能である.
22 3-2. 結果 3-2-1. 3GA の原体および製剤安定性 3GA の原体安定性について検証するため, 3GA 水溶液を長期保管した際の残存率を測定した. そ の結果, AsA は経時的に残存率が著しく低下したのに対し, 3GA は水溶液中において残存率の低下 は認められなかった (Fig. 3-2) 38) .
Figure 3-2. Stabilities of 3GA in aqueous solutions.
Each aqueous solution containing AsA or 3GA was stored at 25˚C for 12 weeks. Remaining ratios of test samples in the aqueous solutions were measured at 0, 2, 4, 8 and 12 weeks after preparation by HPLC. Changes in the contents of test samples in aqueous solutions were expressed as remaining ratios (%) calculated with contents at 0 weeks as percentages.
次に, 3GA の製剤安定性を, ジェル製剤における粘度変化を指標に評価した. 被験物質未配合の ジェルと比較して, AsA はジェルに配合することによって粘度が著しく低下した. また, 最も汎用 性の高い AsA 誘導体である Magnesium ascorbyl phosphate (MAP) については急激な減粘が生じ, 本 試験での測定条件において粘度の測定には至らなかった. 一方, 3GA はジェルに配合しても粘度低 下が起こらなかった (Fig. 3-3) 38)
23
Figure 3-3. Stabilities of gel formulations.
Gel samples containing test samples were prepared, and the viscosities of the gel samples were measured at 24 h after preparation. A gel sample without any test material was also prepared as a control. N. D ; not detect.
3-2-2. 3GA の化学的な ROS 消去作用
AsA の生理活性機能のうち, 特に特徴的な機能である抗酸化作用について検証した. まず, ESR スピントラップ法を用いて, 3GA の化学的な ROS 消去能を確認した. なお, 本試験では, 紫外線曝 露によって皮膚内に産生することが知られている・O2
-
, ・OH, NO・を評価対象とした. 3GA の各種 ROS 消去活性は AsA の消去活性には劣るものの, ・O2
, NO・に対しては, 1 mM, ・OH に対しては, 0. 1 mM の濃度にて ROS, RNS 消去活性が認められ, その効果は濃度依存的な増加が確認された(Table 3-1) 38).
Table 3-1. Radical scavenging abilities of 3GA.
Scavenging abilities against ・O2-, ・OH and NO・ of 3GA were measured using ESR spin-trapping methods. Values indicated means ± S. D. (n = 3). **p<0.01 indicated a significant difference against each control.
24 3-2-3. 3GA の細胞内 ROS 消去作用
培養細胞を用いて 3GA の細胞内での ROS 消去作用について評価を行った. 定常状態での細胞内 ROS レベルに対する 0. 1 mM から 10 mM の 3GA による作用を Fig. 3-4 に示した. 3GA 添加による 有意な細胞内 ROS レベルの減少が濃度依存的に確認された. 次に, 酸化ストレス曝露条件下にお ける細胞内 ROS レベルの評価を行った. HaCaT 細胞に UVB 照射または H2O2処理を行うと, 細胞内
の蛍光強度の有意な上昇が認められ, 細胞内 ROS の産生亢進が確認された. これに対し, 3GA 前処 理により, 細胞内 ROS レベルの上昇が濃度依存的に抑制されることが確認された(Fig. 3-5) 38)
.
F
igure 3-4. Intracellular ROSsuppression of HaCaT keratinocytes treated without/with 3GA at a steady state.
Bars express means ± S. D. (n = 4). **p<0.01 indicated a significant difference against the control.
Figure 3-5. Intracellular ROS suppression of HaCaT keratinocytes treated without/with 3GA after exposure to oxidative stress.
Bars express means ± S. D. (n = 4). **p<0.01 indicated a significant difference against each groups.
25 3-2-4. 3GA の細胞傷害緩和
外因性 ROS は過剰に細胞に曝露されることにより細胞を死に至らしめることはよく知られてい る. そこで, 3GA の外因性 ROS による細胞致死作用の緩和について, 細胞生存率を指標として評価 した. HaCaT 細胞に UVB 照射, H2O2処理, SIN-1 処理を行うことにより有意な細胞生存率の低下が
確認され, 各 ROS 曝露による細胞傷害性が認められた. 10 mM の 3GA 前処理は, 検討した全条件に おいて細胞生存率の低下を有意に抑制し, 細胞傷害に対する緩和作用を有することが確認された (Fig. 3-6) 38).
Figure 3-6. Protective effects against oxidative stress induced cell damage by 3GA in HaCaT keratinocytes. Bars express means ± S. D. (n = 4). *p<0.05 and **p<0.01 indicate a significant
26 3-2-5. 再構築表皮モデルを用いた細胞内 ROS 産生抑制
UVB 照射再構築表皮モデルの細胞内 ROS 産生に対する 3GA の抑制作用について評価を行った. 本検討では, 1 O2, ・O2 -と反応することにより化学発光を示す MCLA を用いた57) . UVB 照射した再生 表皮モデルから強い化学発光が確認され, 皮膚モデル内における顕著な ROS の産生亢進が確認さ れた. これに対し, 5 mg/mL の 3GA を皮膚モデルの表面側から前処理した再生表皮モデルでは, 化 学発光が減弱した(Fig. 3-7) 38) .
Figure 3-7. Suppressive effects of UVB-induced ROS by 3GA in RHEEs.
Detection and visualization of chemiluminescent signals due to ROS generation in RHEEs using the NightOWL. These are reperesentative images. High levels of ROS generation in RHEEs are indicated by the red color and low levels are indicated by the blue color.
27 3-3. 考察 AsA は強力な還元作用によって多彩な生理活性を発揮する一方で, その反応性の高さにより安 定性の低さが指摘されている. AsA は熱や酸素, 光に対して非常に不安定であるため, 水溶液中に おいて経時的な分解が生じる. 分解された AsA はデヒドロアスコルビン酸に変換されることで, 生 理活性を失う. また, この変換過程に発生する AsA ラジカルが要因となって, 生体内の有害作用が 引き起こされる. 一方で, AsA の安定性の低さは, 以下の観点から製剤安定性にも影響を及ぼす. AsA は, 乳化製剤に配合することでそのイオン性の性質から乳化破壊を引き起こし, 粘性製剤にお ける粘度の低下をもたらす. また, AsA は分解されることで褐変化が生じるため, 製剤の色味も変 化させてしまう. さらに, 製剤中での AsA 濃度低下によって, 化粧品製剤を皮膚へ塗布時に期待さ れる生理活性も低下する. 上記課題を解決すべく, AsA の安定性を向上させた誘導体の開発がこれ まで数多くなされてきた. 現在開発されている AsA 誘導体のほとんどは, エンジオール基の一方ないし双方に置換基を導 入した化合物である. 特に代表的な誘導体として, 2 位の水酸基にリン酸マグネシウム塩を付加し た MAP やグルコースを導入したアスコルビン酸グルコシド(AG)が知られている58, 59) . 両化合物は, 置換基の導入によって安定性が長期間維持されるだけでなく, 皮膚内に移行すると, それぞれ生体 内に存在するフォスファターゼおよびα-グルコシダーゼによって置換基が脱離し, AsA が遊離する ことで生理活性が発揮される. その他にもテトライソパルミチン酸を導入した油溶性のテトラヘ キシルデカン酸アスコルビルなども開発され, AsA 由来の皮膚老化に対する予防, 改善効果が確認 されている 6) . これらプロビタミン C は, 化粧品市場を中心に広く流通しているものの, 製剤安定 性の向上には幾分改善の余地があるのが現状である. ゆえに, より安定かつ安全な AsA 誘導体の開 発は, 一般消費者の更なる QOL 向上のためには必要不可欠と考える. そこで, 我々は上記課題を解決すべく新たな AsA 誘導体の開発に着手した. 安定性向上に重要な 置換基の選定について種々の検討を行ったところ, グリセリンを結合させた 3-O-グリセリルアス コルビン酸(3GA)の安定性が高く, 化粧品原料として優れていることを見出した. グリセリンは非 イオン性の低分子化合物で, 最も認知度の高い保湿剤のひとつである. 本質的に人の健康や環境に 対して無毒で, 不快な臭気もなく, 安定性の高い化合物であるため, AsA の安定性を向上させる置
28
換基としては最適と考えられる. 実際に 3GA は, グリセリル基の導入によって AsA 由来のイオン 性が低減することによって, 高い原体安定性が確認された (Fig. 3-2). さらに, AsA や従来の AsA 誘 導体である MAP 配合時において観察されたジェル製剤の粘度低下が 3GA 配合時には見られず, 粘 性製剤に容易に配合できることが示唆された(Fig. 3-3). ゆえに, このような優れた製剤安定性を示 した 3GA には, 製剤中での継続的な生理活性の発現が期待される. 本研究では, AsA の有する生理活性機能のうち, 抗酸化作用に注目して検討を行った. 紫外線曝 露によって皮膚内で産生される ROS は, 皮膚老化に密接に関与している. UVB 照射により皮膚細 胞内で産生される ROS や RNS としては・O2 - , NO・等が報告されている60). 実際に ESR スピントラ ップ法を用いた ROS 消去試験において, 3GA は, ・O2
, ・OH, NO・に対する消去活性を示し(Table 3-1), 特に・OH に対して高い消去活性を示した. ・OH は, ROS のなかでも特に酸化力が強く, 生体分 子の酸化は主に・OH によるものと考えられており61)
, 3GA による ROS 消去活性は有効な抗老化作 用として機能することが期待される.
HaCaT 細胞を用いた評価において, 3GA は定常状態の細胞内 ROS レベルを低減させ(Fig. 3-4), 外 因性の酸化ストレスに対する ROS 産生亢進に対しても抑制作用を示した(Fig. 3-5). さらに, 3GA に は, UVB 照射および外因性 ROS より生じる細胞傷害を緩和する作用が確認された(Fig. 3-6). なかで も, SIN-1 添加によって産生される ONOO による細胞傷害性緩和作用を示したことは興味深い. ONOO- は, 生体内では NO・と・O2 との反応によって生成され, タンパク質におけるチロシン残基 のニトロ化, ミトコンドリアの電子伝達系障害, エネルギー代謝変異によるアポトーシスを誘導す るなど, 特徴的かつ強力な細胞障害性を有する62) . そのため, ONOO- に対する細胞障害緩和作用は, 生体内における基本的機能の恒常性を維持するための重要な機能であり, 3GA で確認された効果は, 抗酸化作用のなかでも効果的な機能のひとつといえる. 単層培養細胞系において効果が確認されたので, 次に角層浸透性を含めた効果を確認するため 再構築表皮モデルを用いて 3GA の抗酸化作用を評価した. 実際の皮膚への塗布を想定して, 3GA を 表皮モデルの表面側 から投与したにもかかわらず, ROS 産生亢進に対する顕著な抑制作用を示し た(Fig. 3-7). この結果から, 3GA が角層から浸透して細胞膜を通過し, 細胞内へ取り込まれること により ROS 消去作用を発揮することが示唆された.
29
以上結果より, 3GA は従来の AsA 誘導体よりも高い製剤安定性を有し, AsA 由来の生理活性を発 揮する高安定型の AsA 誘導体であることが示された.
30 第 4 章. 3GA 誘導体 (3LGA)の抗酸化機能とそのメカニズムについて 4-1. 諸言 AsA は, 抗酸化効果をはじめとする様々な生理活性を発揮することで皮膚老化を予防, 改善する 優れた化合物である. 一方, その水溶性の性質から皮膚への経皮吸収性が低く, イオントフォレー シスなどの施術が効果を最大限発揮する方法となっている63) . 3GA についても水溶性化合物である ため, 皮膚への経皮吸収性に優れているとは言えず, 生理活性機能の向上に関してさらなる改善の 可能性が考えられた. そこで, 3GA の安定性を維持しつつ, 生体内での抗酸化作用を増強させる目 的として, 3GA のグリセリンをアンカーとして疎水基を導入した複数の誘導体を合成した55) . 本章では, 3GA 誘導体の抗酸化機能について評価するとともに, そのメカニズムを検証したとこ ろ, 他の AsA 誘導体には見られない新規な抗酸化メカニズムが確認されたため, その作用について 詳細に検証を行った. 4-2. 結果 4-2-1. 3GA 誘導体の細胞内抗酸化活性 疎水基の導入による生理活性機能への影響を, NHEKs を用いた細胞内 ROS 消去能を指標に検証 した. 活性強度は, 細胞内 ROS の 50%阻害濃度(IC50)をもとに評価した. 結果を Fig. 4-1 に示す. 3GA
の細胞内抗酸化活性は, 既存の AsA 誘導体である MAP とほぼ同等の活性強度であった. 一方で, 疎水基を付加した誘導体では, 鎖長に依存して細胞内 ROS 消去能が増加し, C12 を付加した 3-O-ラ ウリルグリセリルアスコルビン酸(3LGA)において, IC50値が 3.5 μM と最も高値を示した(Fig. 4-1) 44).
31
Figure 4-1. Suppressive effects against H2O2-induced ROS by 3GA derivatives in NHEKs.
4-2-2. 3LGA の化学的な ROS 消去活性
3LGA の化学的な ROS 消去作用を DPPH ラジカルおよび H2O2に対する消去能を指標に評価した.
3LGA は, AsA には劣るものの, 濃度依存的に DPPH ラジカルおよび H2O2に対する有意な消去活性
を示した(Fig. 4-2) 34)
.
Figure 4-2. Radical scavenging abilities of 3LGA. Scavenging abilities of 3LGA and AsA against (a) DPPH radicals and (b) H2O2 were measured using colorimetric analyses.
32 4-2-3. 3LGA の細胞内 ROS 消去作用
NHEKs を用いて, 酸化ストレスに対する 3LGA の細胞内 ROS 消去能を評価した. NHEKs に H2O2
処理および UVB 曝露を行うと, 細胞内 ROS が有意に亢進した. これに対し, 3LGA 処理した細胞で は細胞内 ROS の産生亢進が有意に抑制された(Fig. 4-3a, 4-4a). 一方で, 3LGA と同濃度域の MAP を 処理した細胞には, 有意な ROS 消去作用は認められなかった(Fig. 4-3b, 4-4b) 34)
.
Figure 4-3. Intracellular ROS suppression of NHEKs treated with or without (a) 3LGA or (b) MAP after H2O2 treatment. Bars express means ± S. D. (n = 4). **p<0.01 indicates a significant difference
between groups.
Figure 4-4. Intracellular ROS suppression of NHEKs treated with or without (a) 3LGA or (b) MAP after UVB irradiation. Bars express means ± S. D. (n = 4). **p<0.01 indicates a significant difference
33 4-2-4. 3LGA の細胞傷害緩和 酸化ストレスによって生じる細胞傷害に対する 3LGA の緩和作用に関して, 細胞生存率を指標に 検証した. H2O2処理した NHEKs では, 細胞生存率が有意に低下した. 一方, 3LGA を前処理するこ とによって細胞生存率の低下が有意に抑制された. また, 同濃度域の MAP を処理した NHEKs には, 有意な細胞生存率の改善が認められなかった(Fig. 4-5) 34) .
Figure 4-5. Protective effects against oxidative stress induced cell damage by 3LGA or MAP in NHEKs. Bars express means ± S. D. (n = 4). **p<0.01 indicates a significant difference between groups.
34 4-2-5. 3LGA の細胞内抗酸化成分産生促進
上記結果より, 3LGA は化学的な ROS 消去作用は AsA 比較すると低いが, 細胞内 ROS 消去作用 は優れた作用を発揮したことから, 細胞内の抗酸化システムを活性化している可能性が推察され た. そこで, 3LGA を処理した NHEKs の細胞内抗酸化成分の mRNA レベルを測定した. 3LGA を 24 時間処理した NHEKs では, Catalase や γ-GCS をはじめとする複数の細胞内抗酸化成分の mRNA レ ベルが有意に亢進していた(Fig. 4-6a). 3LGA 処理によって細胞内 GSH 量も濃度依存的に増加して いた (Fig. 4-7a). 一方で, MAP を 3LGA の 10 倍量まで NHEKs に適用したものの, これら因子に有 意な変化がみられなかった(Fig. 4-6b, 4-7b) 34)
.
Figure 4-6. Stimulating effects of catalase, γ-GCS, HO-1and NQO1 mRNA expression by (a) 3LGA or (b) MAP in NHEKs. Bars express means ± S. D. (n = 4). *p<0.05 and **p<0.01 indicate a significant
difference against 0 μM.
Figure 4-7. Stimulating effects of intracellular GSH levels by (a) 3LGA or (b) MAP in NHEKs.
35 4-2-6. 3LGA の PPAR-γ-Nrf2 シグナル活性化
3LGA が複数の細胞内抗酸化成分の産生を亢進したことから, PPAR-γ-Nrf2 シグナルを活性化し ている可能性が推察された(第 1 章参照). そこで, 3LGA 処理による PPAR-γ および Nrf2 mRNA 発現 について確認した. NHEKs に 3LGA を 6 時間処理すると, PPAR-γ および Nrf2 mRNA レベルが有意 に増加した(Fig. 4-8a, 4-9a). さらに, 細胞中の Nrf2 タンパク質を免疫染色すると, 3LGA 処理よって Nrf2 タンパク質レベルも顕著に増加していることが確認された(Fig. 4-10) 34).
Figure 4-8. Effects on the expression levels of PPAR-γ mRNA by (a) 3LGA or (b) MAP.
Bars express means ± S. D. (n = 4). **p<0.01 indicates a significant difference against 0 μM.
F
Figure 4-9. Effects on the expression level of Nrf2 mRNA by (a) 3LGA or (b) MAP.
36
Figure 4-10. Nrf2 expression in NHEKs treated with or without 3LGA detected by immunofluorescent staining and confocal microscopy. (Magnification: 100×)
次に, 3LGA の細胞内抗酸化成分の産生亢進が, PPAR-γ の産生亢進に起因しているか否かを検証 するため, PPAR-γ antagonist を用いて検証した. Fig. 4-11 には, Catalase および γ-GCS mRNA レベル を示している. 3LGA を処理することで, Catalase および γ-GCS mRNA 発現は有意に亢進するのに対 し, PPAR-γ antagonist である BADGE を同時に処理すると, その効果が消失することが確認された. さらに, 3LGA の PPAR-γ および Nrf2 に対する影響を検証するため, RNA 干渉法によって PPAR-γ を ノックダウンした. すると, 3LGA による両 mRNA の産生亢進は有意に低下することが確認された (Fig. 4-12) 34).
37
Figure 4-11. Effect of a PPAR-γ agonist (GW1929) or antagonist (BADGE) on the expression of (a) γ-GCS or (b) catalase mRNAs. Bars express means ± S. D. (n = 4). *p<0.05 and **p<0.01 indicate a
significant difference between groups.
Figure 4-12. Effects of 3LGA on the mRNA expression level of (a) PPAR-γ and (b) Nrf2 in siPPAR-γ treated NHEKs. Bars express means ± S. D. (n = 4). *p<0.05 and **p<0.01 indicate a
38 4-3. 考察
AsA の生理活性を経皮的に生体内で発揮させるためには, 疎水性の性質を有する角層を効率的 に透過しなければならない. 化合物の角層浸透性を向上させる手段として, アルキル鎖を導入する ことによって, 化合物の疎水性を高めることを試みた. また, この疎水的な物性は細胞膜透過性の 向上も期待された. 従来の AsA 誘導体である MAP と同程度の抗酸化活性を有していた 3GA にア ルキル鎖を導入すると, 鎖長の長さに準じて細胞内 ROS 消去能が向上し, 3GA にラウリル基を導入 した 3LGA は, MAP と比較して 1000 分の 1 以下の濃度で顕著な抗酸化活性を示した(Fig. 4-1). 一般 的に経皮吸収されやすい物理的性質として, 分子量 500 以下でオクタノール/水分配係数(logP) が 1 ~3 程度と言われている64, 65)
. 3LGA は分子量が 418 で, 尚且つ logP 値が 2.27 (計算値)であること から, 経皮吸収されやすい物性であることが推察される. 実際に 3LGA の抗酸化作用を詳細に検証 すると, 化学的な ROS 消去能は AsA には劣る作用を示したが(Fig. 4-2), NHEKs を用いた培養細胞 系においては, プロ AsA である MAP と比較して優れた細胞内 ROS 消去能を示した(Fig. 4-3, 4-4, 4-5). そのため, 3LGA が生体内において優れた抗酸化作用を発揮する理由の 1 つとして, 疎水的な 性質による細胞膜透過性が向上している可能性が示唆された.
細胞内の酸化ストレス除去の方法として, 外部から抗酸化物質の適用による ROS 消去以外に細 胞内の生体内抗酸化システムを活性化する方法が考えられる. 3LGA は化学的な ROS 消去能は AsA に劣るものの, 高い細胞内 ROS 消去能を示したため, 生体内抗酸化システムを活性化している可 能性も考えられた. 本機能は上述の通り, 生体内機能の恒常性を維持するためには重要な機構であ る. そこで, 3LGA が生体内抗酸化システムを活性化しているか否かについて検証を行った. NHEKs に 3LGA を処理することによって, Catalase をはじめ複数の細胞内抗酸化成分の mRNA 発現亢進 (Fig. 4-6a)と, GSH の産生量の有意な増加が確認された(Fig. 4-7a). 一方で, MAP 処理ではこれら因 子の発現に変動は見られなかったことから(Fig. 4-6b, 4-7b), 本作用は AsA に由来する作用ではなく, 3LGA に特異な作用であることが示唆された.
3LGA が複数の細胞内抗酸化因子の発現亢進に寄与していたことから, PPAR-γ-Nrf2 シグナルを 活性化している可能性が考えられた(第 1 章参照). そこで, 3LGA 処理後の PPAR-γ および Nrf2 発現 の挙動を確認したところ, 両因子の発現の有意な亢進を示した(Fig. 4-8, 4-9, 4-10). 次に, 3LGA の
39
細胞内抗酸化因子の発現亢進が PPAR-γ 産生に起因しているかを確認するため, PPAR-γ antagonist である BADGE を処理したところ, 3LGA による γ-GCS および Catalase 発現の亢進がキャンセルさ れた(Fig. 4-11). さらには, siPPAR-γ 処理によって, 3LGA による PPAR-γ および Nrf2 発現亢進もキャ ンセルされた(Fig. 4-12). これら結果から, 3LGA は PPAR-γ の発現亢進を介して生体内抗酸化シス テムを活性化することが示唆された. 生体内抗酸化システムを活性化する化合物として, α-トコフェロールが知られている. α-トコフ ェロールはクロマン環とイソプレン鎖から構成される化合物で, HaCaT に処理することで濃度依存 的に細胞内の GSH 量が亢進する. 一方で, 還元性 OH 基を置換したリン酸トコフェロールやイソプ レン鎖を脱離させた Trolox には, 細胞内 GSH 量の亢進作用は認められなかった 66) . また, PPAR-γ の生理的リガンドのひとつにリノール酸が知られている67) . 3LGA が PPAR-γ の産生を促すメカニズ ムについては現時点では不明だが, 長鎖アルキル鎖長の存在や疎水基と還元性 OH 基を同一分子内 にもつ両親媒性構造が生体内抗酸化システムの活性化に何らかの影響を及ぼしている可能性が考 えられる.
以上の結果から, 3GA 誘導体である 3LGA は, 従来の AsA 誘導体にはみられない, 生体内抗酸化 システムを活性化するメカニズムを通じて高い抗酸化作用を発揮することを見出した.