Study on the reassembly mechanism of
fluorescent protein
著者
磯貝 純美
内容記述
学位授与大学: Osaka Prefecture University(大阪
府立大学), 学位の種類: 博士(生物科学), 学位記
番号: 論理第94号, 学位授与年月日: 2011-03-31,
指導教員: 多田俊治.
大阪府立大学博士
大阪府立大学博士
大阪府立大学博士
大阪府立大学博士(
(
(理学
(
理学
理学
理学)
)
)
)学位論文
学位論文
学位論文
学位論文
Study on the reassembly mechanism of fluorescent protein
(蛍光
蛍光タンパク
蛍光
蛍光
タンパク
タンパク
タンパク質
質
質の
質
の
の再構成機構
の
再構成機構に
再構成機構
再構成機構
に
に
に関
関
関
関する
する
する
する研究
研究
研究)
研究
Masami Isogai
磯
磯
磯
Abbreviations
BiFC bimolecular fluorescence complementation
CD circular dichroism
DEAE diethylaminoethyl
DLS dynamic light scattering
DSC differential scanning calorimetry
E. coli Escherichia coli
Endo. endothermic
EYFP enhanced yellow fluorescent protein
FPLC fast protein liquid chromatography
FRET fluorescence resonance energy transfer
GFP green fluorescent protein
HEPES 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid
HPLC high performance liquid chromatography
IPTG isopropyl-1-thio-β-D-galactopyranoside
MALDI matrix assisted laser desorption ionization
mRFP monomeric red fluorescent protein
MS mass spectrometry
NaCl sodium chloride
Ni-NTA nickel-nitrilotriacetic acid
OH group hydroxyl group
PAGE poly acrylamide gel electrophoresis
PCR polymerase chain reaction
PDB protein data bank
PEG polyethylene glycol
r.m.s.d root mean square deviation
SDS sodium dodecyl sulfate
S/N signal-to-noise
Tm temperature max
TOF time of flight
Tris tris-(hydroxymethyl)-aminomethane
Contents
Introduction
1
CHAPTER I
Expression and purification of reassembled Venus1. Introduction 6
2. Materials and methods 7
2-1. Expression
2-2. Purification
2-3. Fluorescence spectroscopy
2-4. CD spectroscopy
3. Results and discussion 13
CHAPTER II
Characterization of reassembled Venus1. Introduction 17
2. Materials and methods 19
2-1. MALDI-TOF MS
2-2. DLS measurement
2-3. Crystallization
2-4. Data collection and structure determination
CHAPTER III
Thermal stabilities and reassembly mechanism1. Introduction 30
2. Materials and methods 32
2-1. Preparation of mutants
2-2. DSC measurement
2-3. Preparation of mCherry
2-4. HPLC analysis of Venus and mCherry
3. Results and discussion 35
Conclusion
46
References
48
Introduction
Protein-protein interactions play key roles in mediating signal-transduction
pathways and executing cellular functions. Therefore, defining how each protein
interacts with all possible partners in cells provides new insight into cellular functions
of individual proteins.
Protein-protein interactions have been investigated using many different
approaches. Although many methods have been developed, most methods that enable
direct detection of protein interactions, such as affinity-precipitation or co-purification
require removal of the proteins from their normal environment. The visualization of
protein interactions in living cells provides the potential for direct detection of protein
interactions with minimal perturbation of their normal environment. GFP was
discovered by Shimomura et al. as a companion protein to aequorin, the famous
chemiluminescent protein from jellyfish Aequorea victoria in the early 1990s (Fig. 1).
GFP is a 238 amino acid protein having a β-barrel structure with the chromophore
contained inside the barrel. It has been developed and contributed to visualization of
proteins as a biological marker for tracking gene expression in cells (Heller et al.,
2010). Several assays such as BiFC, FRET, and bioluminescence resonance energy
transfer have been developed and most widely used to visualize protein-protein
interactions in living cells (Shyu et al., 2008). These assays can provide information
regarding when and where protein-protein interactions occur in the cell.
2002; Kerppola, 2006). The basic principle of this technique is the formation of the
fluorescent complex through the association of two non-fluorescent N- and C-terminal
fragments of the fluorescent protein when they are brought together by an interaction
between two proteins fused to the fragments (Figs. 2 & 3). A number of fluorescent
proteins-such as yellow, cyan, green, and red-have been also utilized to study post
translational modifications, protein folding, protein aggregation, protein
conformational change, and protein topology (Hu and Kerppola, 2003; Fan et al.,
2008). In particular, Venus (λex = 515 nm, λem = 528 nm) generated from EYFP has
been recently used in the BiFC assay, because of its fast chromophore maturation rate
and bright yellow fluorescence that is relatively insensitive to changes in pH and ion
concentrations (Nagai et al., 2002; Shyu et al., 2006).
The crystal structure of Venus (whole Venus) was determined by X-ray
crystallography at a resolusion of 2.2 Å (Rekas et al., 2002). The structure of whole
Venus shows an eleven-stranded β-barrel, typical of GFP-derived fluorescent proteins.
Although the Venus-based BiFC assay provides information regarding when and where
protein-protein interactions occur in the cell, background from spontaneous assembly of
the fragments compromises their utility for detecting the interactions. The decrease of
BiFC assay (Kodama and Hu, 2010).
There is no report for the structure and characteristics of reassembled Venus
formed by the complementation between the N- and C-terminal fragments, nevertheless
they should provide important information on the strategies to decrease background
fluorescence in the BiFC assay. Thus, in order to facilitate the application of BiFC to
research of protein-protein interactions, we have attempted to elucidate the structure and
the physicochemical properties of reassembled Venus and its mutants. It was difficult to
obtain the fragment of split fluorescent proteins expressed in E. coli separately due to
intrinsic folding problems and low solubility of these fragments in aqueous solutions.
Recombinant expression of fragment of split fluorescent proteins often results in low
sample yield or most of them express in insolubility form in E. coli cells (Ottmann et al.,
2009). Thus, the author attempted to co-express two fragments of split fluorescent
protein in E. coli and successfully overexpressed reassembled Venus in soluble form.
The author describe the X-ray crystal structure of reassembled Venus at a resolusion of
2.1 Å and its characteristics assessed by chromatography, MALDI-TOF MS, CD
spectra. Thermal stability of reassembled Venus including three mutants, Y143F, Y145F,
H148G, were evaluated by DSC analysis.
The author also attempted to reveal reassembly mechanisms. The folding
mechanism of β-barrel proteins using GFP has been investigated (Kent et al., 2008;
Kent et al., 2009). Proteins composed of β-Strand frequently form oligomers. This
oligomerization is often necessary for protein activity or regulation and its efficiency is
disease, mad cow disease, Parkinson’s disease. This study may provide perspectives
for the folding mechanism of β-strand rich proteins.
Fig. 1 Aequorea victoria
VN155 VC155 Proteins for visualization Reconstitution of fluorescent protein Recovery of fluorescence Interaction of two proteins fused
to each fragments
+
VN155 VC155 Proteins for visualization Reconstitution of fluorescent protein Recovery of fluorescence Interaction of two proteins fusedto each fragments VN155 VC155 Proteins for visualization Reconstitution of fluorescent protein Recovery of fluorescence Interaction of two proteins fused
to each fragments
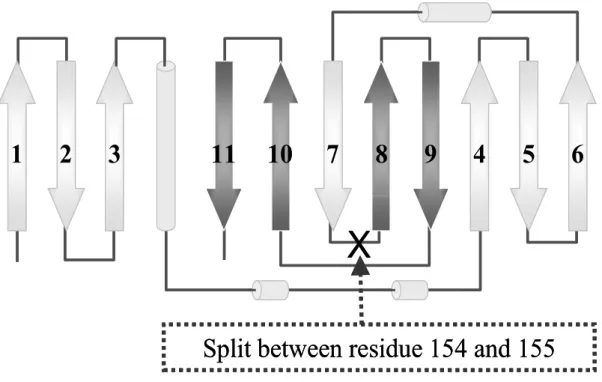
Fig. 3 A topology of fluorescent protein
2
3
11
10
7
8
9
4
5
6
1
X
Split between residue 154 and 155
2
3
11
10
7
8
9
4
5
6
1
X
CHAPTER I
Expression and purification of reassembled Venus
1. Introduction
Monitoring protein–protein interactions and macromolecular complex formation
in living cells is extremely useful for understanding the dynamics and mechanism of
biological processes. In order to facilitate the application of BiFC to research of
protein-protein interactions, the author have attempted to determine the structure of
reassembled Venus complemented between the N-terminal and C-terminal fragments.
Ghosh et al. succeeded to express reassembled GFP in E. coli as a fusion protein
with anti parallel leucine zipper-directed proteins (Ghosh et al., 2000). However, the
fusion protein was expressed in insolubility form in E. coli cells. It was also difficult to
obtain the fragments of split fluorescent proteins expressed in E. coli separately due to
intrinsic folding problems and low solubility of these fragments in aqueous solutions
(Own unpublished results.). Thus, the author developed the coexpression system of
two fragments in E. coli using two plasmids. Fluorescent spectroscopy and CD
analysis were carried out to examine the differences between intact Venus (whole
2. Materials and methods
2-1. Expression
N-terminal fragment (amino acid residues 1-154, VN155) and C-terminal
fragment (amino acid residues 155-238, VC155) of Venus were subcloned into pET-16b
and pET-28a plasmids (Novagen), respectively (Fig. 4). VN155 tagged with (His)10 at
its N-terminus and VC155 tagged with (His)6 at its C-terminus were coexpressed in E.
coli strain BL21(DE3) bacteria (Novagen). Cells were grown using standard culture
medium (LB broth) in a shaker incubator at 37ºC. Expression was induced with 0.5 mM
IPTG at O.D.600nm of 0.5 and cultivation was continued for a further 12 h at 22ºC.
Whole Venus (amino acid residues 1-238) was subcloned into pET-3a plasmid
(Novagen) (Fig. 4) and expressed in E. coli BL21(DE3) bacteria. Cells were cultured
under the same conditions as described above.
2-2. Purification
Cells of reassembled Venus were harvested and resuspended in 50 mM sodium
phosphate buffer, pH 8.0, containing 300 mM NaCl and 10 mM imidazole. After
sonication and centrifugation, the supernatant was loaded onto a Ni-NTA column
(QIAGEN) equilibrated with 20 mM Tris-HCl buffer, pH 8.0, containing 150 mM NaCl
(buffer A) and eluted with 250 mM imidazole. The fractions containing reassembled
Venus were combined and concentrated. The concentrated protein solution was loaded
onto a MonoQ anion-exchange column (GE Healthcare) equilibrated with 20 mM
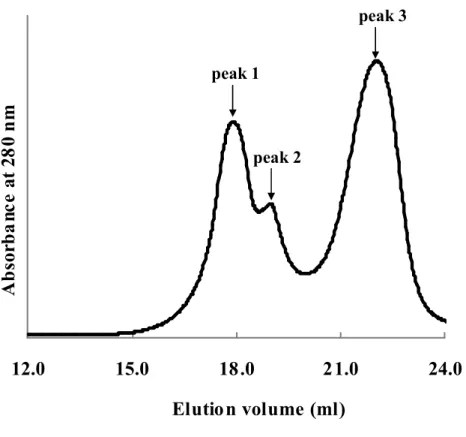
NaCl (20-1000 mM). The anion-exchange chromatography yielded three peaks showing
yellow fluorescence (Fig. 5). The fraction corresponding to each peak was finally
purified by the gel filtration chromatography using a Superdex 200 column (GE
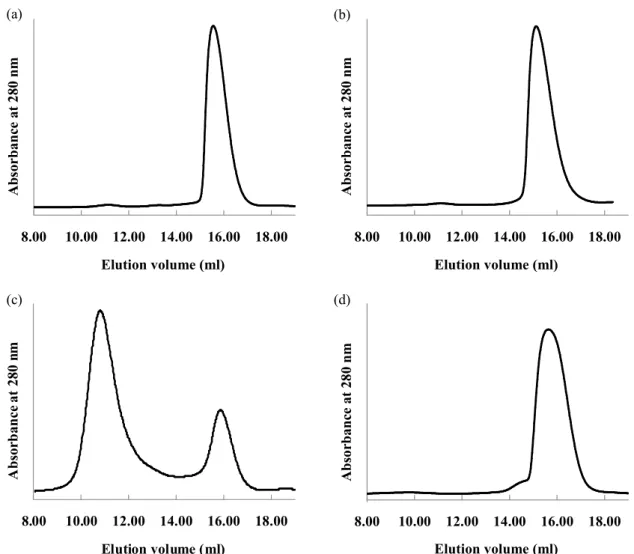
Healthcare) equilibrated with 20 mM sodium phosphate buffer, pH 7.0 (Fig. 6a-c).
Cells of whole Venus were harvested, resuspended in buffer A, and sonicated. The
lysate was centrifuged to obtain a crude sample. The supernatant was loaded onto a
DEAE sepharose ion-exchange column (GE Healthcare) equilibrated with buffer A and
eluted with a linear gradient of NaCl (100-700 mM). Whole Venus samples were further
purified by anion exchange chromatography using MonoQ column followed by a size
exclusion chromatography using Superdex 200 (Fig. 6d).
All purification steps were carried out at 4ºC and an anion exchange
chromatography using MonoQ and size exclusion chromatography using Superdex 200
were performed on the ÄKTAFPLC system (GE Healthcare).
2-3. Fluorescence spectroscopy
Whole Venus, reassembled Venus were diluted to 0.1 mg/ml with 20 mM sodium
phosphate buffer, pH 7.0 for fluorescence spectral analysis. Fluorescence spectra were
2-4. CD spectroscopy
Samples were diluted to 0.1 mg/mL with 20 mM sodium phosphate buffer, pH
7.0 for CD spectral analysis. CD experiments were performed on a J-820
spectropolarimeter (JASCO) with a Peltier PTC-423L thermo-unit (JASCO). The
far-UV CD spectra (260-190 nm) were recorded using a 0.1 cm path length cell under
constant nitrogen flush with a step size of 0.2 nm, bandwidth of 1 nm, and an
Fig. 4 Constructions of VN155, VC155 and whole Venus.
pET-16b(+)
His-tag
VN155
A
m
p
Nde I
BamH I
(Mw 20 kDa)
pET-28a(+)
His-tag
VC155
K
an
Nco I
Xho I
(Mw 10 kDa)
pET-3a(+)
Whole Venus
A
m
p
Nde I
BamH I
(Mw 27 kDa)
Fig. 5 Anion exchange chromatogram of reassembled Venus.
The elution points of three components of reassembled Venus are shown
along the top.
12.0
15.0
18.0
21.0
24.0
Elutio n volume (ml)
A
b
so
rb
a
n
c
e
a
t
2
8
0
n
m
peak 1 peak 2 peak 3Fig. 6 Size exclusion chromatograms.
(a) Peak 1 of reassembled Venus. (b) Peak 2 of reassembled Venus.
(c) Peak 3 of reassembled Venus. (d) Whole Venus.
8.00 10.00 12.00 14.00 16.00 18.00 Elution volume (ml) A b so rb a n ce a t 2 8 0 n m 8.00 10.00 12.00 14.00 16.00 18.00 Elution volume (ml) A b so r b a n ce a t 2 8 0 n m 8.00 10.00 12.00 14.00 16.00 18.00 Elution volume (ml) A b so r b a n ce a t 2 8 0 n m 8.00 10.00 12.00 14.00 16.00 18.00 Elution volume (ml) A b so rb a n c e a t 2 8 0 n m (a) (b) (d) (c)
3. Results and discussion
Reassembled Venus was expressed in E. coli as a polyhistidine fusion protein.
The fluorescent fraction isolated by a Ni affinity column chromatography was
separated into three peaks on the anion exchange chromatography (Fig. 5). The profiles
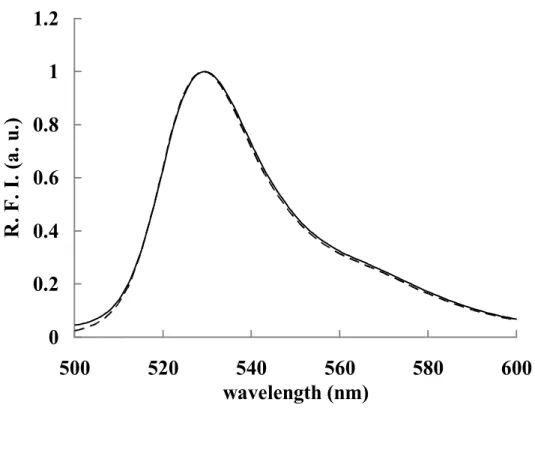
of fluorescent spectra of these three peaks were identical to that of whole Venus (Fig.
7). The N- and C-terminal fragments of split fluorescent protein do not exhibit the
absorption or fluorescence characteristics unless two fragments of split fluorescent
protein completely associate to form a fluorescent complex having a β-barrel
conformation (Sakamoto and Kudo, 2008; Kent et al., 2008). Thus, three peaks should
contain the fluorescent complex formed through the association of two non-fluorescent
fragments at least as a parent compound.
Each peak was further purified by a size exclusion chromatography. The size
exclusion chromatograms of peaks 1 and 2, respectively, showed a single peak (Figs.
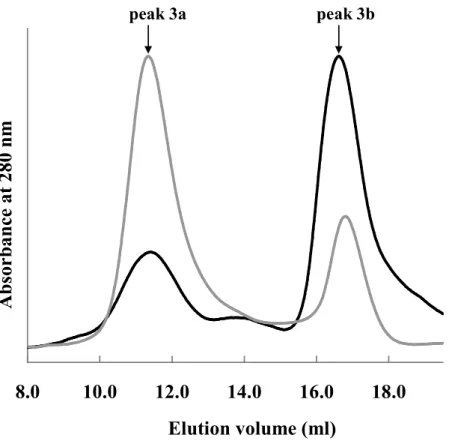
6a & 6b). The size exclusion chromatogram of peak 3 yielded two peaks after
incubation for 1 day at 4ºC (Fig. 6c). Both peaks were fluorescent. MALDI-TOF MS
analysis showed that the main components of peak 3 were VN155 and VC155
fragments of Venus as described below. Thus, peak 3a emerged earlier and peak 3b
emerged later were estimated to be an oligomer (octamer to decamer) and a monomer
of reassembled Venus based on their retention volumes, respectively. This view was
supported by CD spectrum of peak 3b which was identical with that of peak 1 (Fig. 8).
After incubation for 3 days at 37ºC, the area of peak 3a further decreased and that of
peak 1, after incubation for 3 days at 37ºC, did not give any other peaks except original
peak. These findings suggest that the oligomer is converted to the monomer in an
irreversible manner or the rate of conversion to oligomer is extremely slow.
Fig. 7 Fluorescence spectra
Fluorescent spectra of whole and reassembled Venus (peak 1) are
indicated solid line and dashed line, respectively.
0
0.2
0.4
0.6
0.8
1
1.2
500
520
540
560
580
600
wavelength (nm)
R
.
F
.
I.
(
a
.
u
.)
Fig. 8 CD spectra
Peak 1 of reassembled Venus and peak 3b of reassembled Venus are
indicated solid line and dashed line, respectively.
-4000
-2000
0
2000
4000
6000
8000
10000
190
210
230
250
Wavelength (nm)
[θ
](
d
eg
.c
m
2/d
m
o
l)
Fig. 9 Size exclusion chromatogram of peak 3 of reassembled Venus.
The chromatogram of peak 3 incubated for 1 day at 4ºC is present in gray,
and that of peak3 incubated for 3 days at 37ºC is present in
black. For
comparison, the absorbance is normalized by the maximum absorbance.
The elution points of peak 3a and peak 3b are shown along the top.
8.0
10.0
12.0
14.0
16.0
18.0
Elution volume (ml)
A
b
so
r
b
a
n
ce
a
t
2
8
0
n
m
peak 3a peak 3bCHAPTER II
Characterization of reassembled Venus
1. Introduction
GFP has been the subject of continued interest since its gene was first cloned in
1992. Crystal structures of both the monomeric and dimeric forms of GFP have been
solved previously (Ormo et al., 1996; Yang et al., 1996). Chromophore is situated in the
middle of a distorted α-helix that runs through the center of a β-barrel in the folded GFP
structure. Maturation of GFP proceeds in three major steps, beginning as the
238-residue single GFP polypeptide folds into its nearly native conformation. Residues
65–67 of the folded protein then undergo several chemical reactions necessary for
chromophore formation, including cyclization and dehydration. The final rate-limiting
step in the maturation process involves the oxidation of the Cα–Cβ bond of Tyr66 by
aerial oxygen. It has been shown that the protein structure itself plays an essential role
in creating and maintaining a semi-rigid environment around the chromophore where
bulk solvent molecules are excluded and the conformational flexibility of the
chromophore is low. It leads to a state with a high quantum yield.
Maturation process takes ~3 h at room temperature and its efficiency further
decreases at 37°C. Venus was developed to improve maturation rate at 37ºC. The
crystal structure of Venus (PDB: 1MYW) (whole Venus) was determined by X-ray
crystallography at a resolusion of 2.2 Å (Rekas et al., 2002). The structure of whole
Background from spontaneous assembly of the fragments compromises their
utility for detecting protein-protein interactions. Fluorescent protein fragments have a
finite ability to associate with each other independent of an interaction between
proteins fused to the fragments. To facilitate the application of BiFC, the author have
attempted to determine the X-ray crystal structure of reassembled Venus and its
2. Materials and Methods
2-1. MALDI-TOF MS
The molecular mass of each component was measured on an autoflex II
MALDI-TOF analyzer (Bruker Daltonics) using saturated α-cyano-4-hydroxycinnamic
acid in 50%(v/v) acetonitrile and 0.1%(v/v) trifluoroacetic acid as a matrix.
Reassembled Venus was performed in the 20 mM phosphate buffer (pH 7.0) at
concentrations of 100 µg/mL.
2-2. DLS measurement
Measurements of the hydrodynamic radii of whole and reassembled Venus were
performed at 4ºC in the buffer containing 20 mM phosphate buffer (pH 7.0), 100 mM
NaCl at concentrations of 0.5 mg/mL using a Wyatt DynaPro Nanostar dynamic light
scattering instrument (Wyatt Technology). The data were analyzed using Dynamics 6.0
software.
2-3. Crystallization
Prior to crystallization, the purified monomer of reassembled Venus was dialyzed
against buffer A (20 mM Tris-HCl buffer, pH 8.0, containing 150 mM NaCl) and
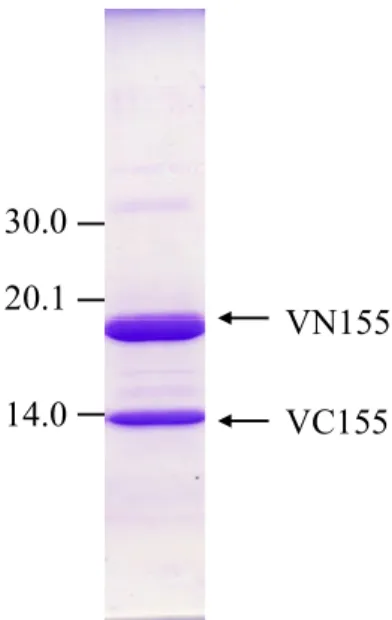
concentrated using an Ultafree (Millpore) filter. Protein purity was evaluated by
SDS-PAGE (Fig. 10). Crystallization trials were carried out at 20ºC using the
sitting-drop vapor-diffusion technique by mixing 0.5 µL of 35 mg/mL protein sample
the commercially available sparse-matrix screening kit Index (Hampton Research).
Small but good shaped crystals were obtained (Fig. 11) from the reservoir solution
consisting of 25% (w/v) PEG 3350, 0.2 M Lithium sulfate and 0.1 M HEPES buffer pH
7.5.
2-4. Data collection and structure determination
A diffraction data set was collected to a resolution of 2.1 Å at 100 K on an ADSC
CCD detector using synchrotron radiation of wavelength 1.0 Å at the BL5A station of
Photon Factory, Japan. The data set was processed and scaled with the HKL2000
software package (Otwinowski and Minor, 1997). The crystals belong to the space
group P212121, with unit-cell parameters a = 59.05, b = 116.05, c = 156.85 Å. The
crystal structure was solved by the molecular replacement using the program MOLREP
(Vagin and Teplyakov, 1997) with the structure of Venus (PDB: 1MYW) as a search
model. The solution revealed that the crystal contains a tetramer as the asymmetric unit.
The obtained model was subjected to iterative rounds of model building and refinement
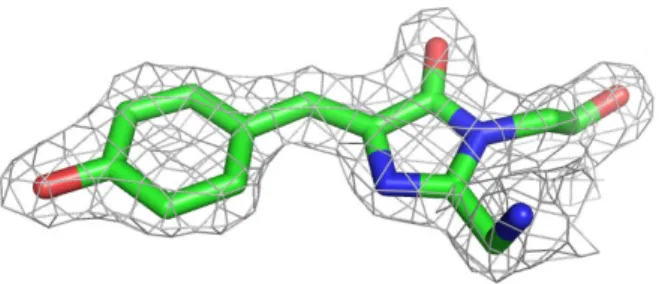
using the programs COOT (Emsley and Cowtan, 2004) and REFMAC5 (Winn et al.,
2003) (Fig. 12). The 2Fo-Fc difference map showed clear electron density
residues were in their most favored regions, 8.6% of the residues were in additionally
allowed regions, 0.3% of the residues were in generously allowed regions and no
residues were in disallowed regions (Fig. 13). Data collection and structure refinement
statistics are summarized in Table 1. All structure figures were prepared with the
program PyMOL (DeLano, 2002, http://www.pymol.org.). Coordinates have been
deposited with the Protein Data Bank with the following accession codes: 3AKO.
Fig. 10 SDS-PAGE. Fig. 11 A crystal of reassembled Venus.
VN155 VC155 30.0 20.1 14.0
Fig. 12 The 2Fo-Fc electron density map of the chromophore
The 2Fo-Fc electron density map contoured at 1.5 σ showing the refined
structure of the chromophore of reassembled Venus.
Table 1 Summary of data collection and refinement statistics.
aData Set reassembled Venus Data collection
X-ray source BL5A, PF
Wavelength (Å) 1.00 Space group P212121 Unit cell (Å) a = 59.05 b = 116.05 c = 156.85 Resolution (Å) 50.0 – 2.10 (2.18-2.10) Unique reflections 63893 (6263) Completeness (%) 99.9 (99.9) Rmerge (%)b 10.2 (32.9) I/
σ
23.9 (5.8) Redundancy 11.0 Refinement statistics Resolution (Å) 50.0 – 2.10 Reflections 60532 R-factor (%)c 16.7 Rfree (%) 22.9 R.m.s.deviations Bond length (Å) 0.016 Bond angle (˚) 1.8 aValues in parentheses are for the highest resolution shell.
b
Rmerge=ΣhΣj|Ihj-<Ih>|/ΣhΣj|Ihj|, where h represents a unique reflection and j represents symmetry-equivalent indices. I is the observed intensity and <I> is the mean value of I.
c
3. Results and discussion
Based on its retention volume in size exclusion chromatography (Fig. 6) and
MALDI-TOF MS data (Table 2), peak 1 was elucidated to be a monomer of
reassembled Venus. The structure of the species corresponding to peak 1 was
confirmed by X-ray structure analysis. The retention volume indicated that peak 2 was
also a monomer of reassembled Venus. However, the molecular weight of VN155
found in MS is approximately 180 Da larger than the calculated value (Table 2). These
results suggested that a part of reassembled Venus might be modified during incubating
in E. coli.
DLS data (Table 3) indicated that peak 3 was a molecular aggregate which have
a molecular weight corresponding to an octamer of reassembled Venus. However, as
previously indicated in CHAPTER I, this octamer is converted to a monomer in an
irreversible manner or the rate of conversion to oligomer is extremely slow.
The crystal structure of monomeric reassembled Venus was determined at 2.1 Å
resolution. There are four reassembled Venus molecules formed through the
association of two VN155 and VC155 fragments in an asymmetric unit (Fig. 14). Each
molecule has an eleven-stranded β-barrel fold including a chromophore in the middle,
concentration of protein and in high-salt concentration. The dimer structure may be
stabilized in crystals. The locations of the residues surrounding the chromophore of
reassembled Venus are quite similar to those surrounding the chromophore of whole
Venus (Fig. 16). Fluorescent spectrum of reassembled Venus was identical to whole
Venus (Fig. 7). There is no difference between the environment surrounding
chromophore of reassembled Venus and that of whole Venus.
In the β-barrel fold between reassembled Venus and whole Venus, a slight but
significant difference was found. R.m.s.d. values of the mainchain calculated using
LSQKAB with monomer of reassemble and whole Venus are 0.59 Å for N-terminal
fragment (amino acid residues 1-154) and 1.66 Å for C-terminal fragment (amino acid
residues 155-230), respectively. As compared with whole Venus, the seventh β-strand
(β7) of reassembled Venus is shortened (Figs. 17 & 18). Unlike whole Venus, two
amino acid residues, Asn146 and Ser147, didn’t contribute to form β7. The loop
(amino acid residues from 138 to 147) of reassembled Venus was expanded and more
flexible than that of whole Venus by lacking of hydrogen bonds between β7 and β10.
This means that the formation of the β-barrel structure of reassembled Venus is
Table 2 TOF-MS of reassembled Venus.
Comparison between the found and calculated masses of proteins.
Table 3 Hydrodynamic Radii, polydispersity values (Pd%) and
molecular weight of reassembled Venus, as determined by DLS
analysis.
peak1
Found
Calculated
VN155
VC155
VN155
VC155
19925
19920
10558
10552
20104
10553
19917
10552
peak2
peak3
peak1Radius (nm) %Mass Mw (kDa)
2.5 6.3 30 245 100.0 99.8 peak3 %Pd 13.2 19.1 Polymerization monomer octamer
Fig. 14 Crystal structure of reassembled Venus.
Ribbon diagram of a tetramer of reassembled Venus, which consists of
VN155 (Chain A, C, E, G) and VC155 (Chain B, D, F, H).
Chain A
Chain B
Chain E
Chain F
Chain C
Chain D
Chain G
Chain H
Chain A
Chain B
Chain E
Chain F
Chain C
Chain D
Chain G
Chain H
Chain A
Chain B
Chain E
Chain F
Chain C
Chain D
Chain G
Chain H
Fig. 15 Superposition of the main chain of the dimer structure of
reassembled (yellow) and whole Venus (cyan).
chromophore
Tyr203
His148
Asn146
Ser205
Glu222
Thr62
Arg96
Gln94
Fig. 17 Comparison of reassembled Venus and whole Venus.
Whole Venus is represented in cyan. VN155 and VC155 of reassembled
Venus are represented in orange and yellow, respectively.
Fig. 18 β
β
β
β7 of reassembled Venus is shorter than that of whole Venus.
Hydrogen bond (dashed line) between β7, β10 and β11of reassembled
(yellow) and whole Venus (cyan).
β ββ β7 β ββ β8 β β β β9 β ββ β10 β β β β11 β ββ β7 β β β β8 β β β β9 β ββ β10 β ββ β11 β β β β4 ββββ4 C C N N C N β β β β7 β β β β10 β ββ β11 β ββ β7 β β β β10 β ββ β11
CHAPTER III
Thermal stabilities and reassembly mechanism
1. Introduction
Since the identification of the GFP from jellyfish Aequorea victoria in the early
1990s, a large number of fluorescent proteins have been isolated from natural sources,
primarily from marine animals and corals (Hsu et al., 2010). mFruits are
second-generation mRFPs that improved brightness and photostability compared to the
first-generation mRFP1 (Shu et al., 2006). mCherry (λex = 587 nm, λem = 610 nm)
derived from Discosoma sp. (Figs. 19 & 20), has been developed for its fast
chromophore maturation rate as same as yellow fluorescent protein, Venus (Shaner et
al., 2004). The brilliant redness, short maturation time, and the long excitation and
emission wavelengths of mCherry make the new BiFC system for analyzing protein–
protein interactions in living cells and for studying multiple protein – protein
interactions when coupled with other BiFC systems. This new red BiFC system was
developed by splitting mCherry, into two fragments between amino acids 159-160 as
well as splitting Venus.
Fig. 19 Discosoma sp.
2. Materials and Methods
2-1. Preparation of mutants
Three mutants of VN155 (Y143F, Y145F, H148G) were constructed by
site-directed mutagenesis of plasmids carrying VN155 (pET-16b) by PCR using Pfu
Turbo (Stratagene) and primers (Table 4). The sequence of mutants were verified by
DNA sequencing with a dye terminator cycle sequencing kit (Beckman Coulter) and a
CEQ2000 fragment analysis system (Beckman Coulter). Reassembled Venus
consisting of VN155 mutants and VC155 (denoted as rV-Y143F, rV-Y145F,
rV-H148G) were coexpressed and purified by the same procedures as those used for
reassembled Venus.
2-2. DSC measurement
Calorimetric experiments were carried out with a nanoDSC (TA instruments).
Samples were prepared in concentrations of 0.5 and 1.0 mg/mL. The buffer used for the
sample was 20 mM sodium phosphate, pH 7.0. Experiments were performed over a
temperature range of 25-95ºC at a scan rate of 1 ºC/min and excess pressure of 2.8 atm.
in E. coli strain BL21(DE3) bacteria (Novagen). Whole mCherry (amino acid residues
1-238) was subcloned into pRSET-B plasmid and expressed in E. coli strain
BL21(DE3)pLysS bacteria. Cells were grown using LB broth in a shaker incubator at
37ºC. Expression was induced with 0.5 mM IPTG at O.D.600nm of 0.5 and cultivation
was continued for a further 12 h at 22ºC.
Cells of reassembled mCherry were harvested and resuspended in 50 mM
sodium phosphate buffer,pH 8.0, containing 300 mM NaCl and 10 mM imidazole.
After sonication and centrifugation, the supernatant was loaded onto a Ni-NTA column
equilibrated with 20 mM Tris-HCl buffer, pH 8.0, containing 150 mM NaCl (buffer A)
and eluted with 250 mM imidazole. The fractions containing reassembled Venus were
combined and concentrated. Reassembled mCherry was finally purified by the gel
filtration chromatography using a Superdex 200 column (GE Healthcare) equilibrated
with 20 mM sodium phosphate buffer, pH 7.0.
Cells of whole mCherry were harvested, resuspended in buffer A, and sonicated.
The lysate was centrifuged to obtain a crude sample. The supernatant was loaded onto a
DEAE sepharose ion-exchange column (GE Healthcare) equilibrated with buffer A and
eluted with a linear gradient of NaCl (100-700 mM). Whole mCherry samples were
eluted in the wash buffer. Whole mCherry samples were further purified by a size
2-4. HPLC analysis of Venus and mCherry
Whole Venus, reassembled Venus, whole mCherry and reassembled Venus were examined by HPLC analysis at room temperature. HPLC analysis was performed using L-6200 Intelligent Pump (HITACHI), ELITE LaChrom L-2400 UV Detector (HITACHI), D-2500 Chromato-Integrator (HITACHI). Columns were TSKgel SuperSW2000 4.5*300 (TOSOH) and TSK guardcolumn SuperSW 4.6*3.5 (TOSOH). mCherry and Venus were detected at 587 nm and 515 nm, respectively.
Table 4 List of primers.
Y143F forward 5’- CACAAGCTGGAGTTCAACTACAACAGC -3’
Y143F reverse 5’- GCTGTTGTAGTTGAACTCCAGCTTGTG -3’
Y145F forward 5’- GGAGTACAACTTCAACAGCCACAAC -3’
Y145F reverse 5’- GTTGTGGCTGTTGAAGTTGTACTCC -3’
H148G forward 5’- CAACTACAACAGCGGCAACGTCTATATC -3’
H148G reverse 5’- GATATAGACGTTGCCGCTGTTGTAGTTG -3’
3. Results and discussion
Based on this structural features, mutations in β7 of VN155, Y143F, Y145F and
H148G (Figs. 21 & 22), were introduced to see if any change in the thermal stability of
reassembled fluorescent complex is observed.
DSC studies were carried out on whole Venus, reassembled Venus and mutants
(Fig. 23, Table 5). The profiles were analyzed mainly in terms of the peak temperatures,
because all of the samples showed irreversible transition. The thermogram of whole
Venus shows a sharp single peak at 89.0ºC (Tm). On the other hand, the thermogram of
monomeric reassembled Venus shows a sharp single peak at a lower temperature of
77.5ºC. The Tm value of the monomeric reassembled Venus did not fall at a lower
concentration of 0.1 mg/mL. It can therefore be presumed that the dissociation and the
thermal denaturation of monomeric reassembled Venus occur simultaneously at around
Tm value.
The thermograms of three mutants of reassembled Venus, rV-Y143F, rV-Y145F
and rV-H148G, showed Tm values of 77.1ºC, 83.8ºC and 72.1ºC, respectively (Table 5,
Fig. 23). The OH group of Tyr143 forms a hydrogen bond with the carbonyl O atom of
Ser208 (Fig. 22a). Despite the hydrogen bond is lacking, Tm value of the rV-Y143F
mutant was the almost same as that of reassembled Venus. Surprisingly, the
substitution of Tyr145 with phenylalanine enhanced the thermal stability compared
with monomeric reassembled Venus. Tyr145 and His169 are linked by the hydrogen
bond network through two water molecules (Fig. 22b). The substitution seems to cause
hydrophobic amino acids, Val61 and Ile167, or cause the rearrangement of those amino
acids into alternative dense-packing structure. In contrast, the significant decrease in
thermal stability was observed when His148 was substituted with glycine. This mutant
seems to be useful for BiFC assay, because the lacking of the hydrogen bonds (Fig.
22c) may weaken the binding force between N- and C-fragments of Venus.
DSC measurement for the oligomer of reassembled Venus showed two peaks;
one peak is identical to that for the monomer of reassembled Venus and has a
maximum at 77.6ºC, and the other peak spreads through the range from 40ºC to 65ºC
(Fig. 24, cyan). The size exclusion chromatography indicated that the oligomer was
readily converted to the monomer. Thus, it can be considered that the conversion from
oligomer to monomer occurs while temperature rises to ~65ºC.
HPLC analysis was carried out on whole Venus, reassembled Venus, whole
mCherry and reassembled mCherry to confirm whether reassembled mCherry forms an
oligomer as well as reassembled Venus. Reassembled mCherry is a split red fluorescent
protein at amino acid residue 159/160. The size exclusion chromatography of whole
mCherry yielded one peak as same as whole Venus (Fig. 25). On the other hand, the
chromatography of reassembled mCherry yielded two peaks as same as reassembled
of mCherry has a great thermal stability compared to those of Venus.
BiFC assay utilizes the formation of the fluorescent complex through the
association of two non-fluorescent N- and C-terminal fragments of the fluorescent
protein when they are brought together by an interaction between two target proteins
fused to the fragments. Thus, oligomerization will not be preferable for BiFC assay,
because it will prevent the interaction between two target proteins and increase
background. In the β-strand swapping, it has been also reported that proline residues
play an important role (Bergdoll et al., 1997). The cis-trans proline isomerization, a
very low energy barrier, is a relatively slow process that can affect the protein folding
pathway. Interestingly, reassembled Venus has three proline residues, Pro187, Pro192
and Pro196, and mCherry has two proline residues, Pro186, Pro190, on the loop region
between β9 and β10 (Fig. 27) and they seem to play a critical role in the domain,
including β10- and β11-strands or β8- and β9-strands, swapping. It is suggested that
replacing a proline with another amino acid should depress the formation of oligomers.
Here, the author proposes the model of reassembly mechanism of fluorescent
protein (Fig. 28). After coexpression in E. coli, most of VN155 and VC155 form a
metastable oligomer by β-strand swapping as described above. Then, this oligomer is
converted irreversibly to a more stable monomer. However, ∆H of oligomer (16.3
KJ/mol) was lower than that of monomer (22.9 KJ/mol). It is suggested that a part of
Fig. 21 Three mutants of VN155 of reassembled Venus.
Y143F Y145F H148G Y143F Y145F H148GFig. 22 Close-up views of three amino acids around β
β
β7.
β
Hydrogen bonds are indicated by dashed lines. Water molecules are drawn
with pink spheres. (a) Tyr143. (b) Tyr145. (c) His148.
Tyr143 Ser208 His169 Tyr145 His148 chromophore Arg168 (a) (b) (c)
Fig. 23 DSC of whole and reassembled Venus and its mutants.
Heat capacity curves for whole Venus (black), peak 1 of reassembled
Venus (red), Y143F (green), Y145F (pink), H148G (blue).
Table 5 Comparison of T
mvalue (ºC).
20 30 40 50 60 70 80 90 100 E x ce ss H ea t C ap ac it y Temperature / oC 20 kJ K-1 mol-1
reassembled Venus
77.5
Mutant rV-Y143F
77.1
rV-Y145F
83.8
Fig. 24 DSC of reassembled Venus.
Heat capacity curves for peak 1 of reassembled Venus (red) and peak 3 of
reassembled Venus (cyan).
20 30 40 50 60 70 80 90 100 E x ce ss H ea t C ap ac it y Temperature / oC 20 kJ K-1 mol-1 peak 1 peak 3 ∆H = 22.9 KJ/mol ∆H = 16.3 KJ/mol
Fig. 25 Chromatogram of whole (top) and reassembled mCherry
whole mCherry
reassembled mCherry
0
20 min
Fig. 26 DSC of Venus and mCherry.
Heat capacity curves for peak 1 of reassembled Venus (green), whole
mCherry (pink), reassembled mCherry (blue), respectively.
30 40 50 60 70 80 90 100 110 60 80 100 120 140 160 k J K -1 m o l -1 Temperature / oC VNVC1 1.0 mg/ml cherry 1.0 mg ml cherryR 1.0 mg/ml
30 40 50 60 70 80 90 100
110
Temperature (ºC)
160
140
120
100
80
60
E
n
d
o
.
E
x
ce
ss
H
ea
t
C
ap
ac
it
y
(k
J
K
-1m
o
l
-1)
Fig. 27 Position of prolines in reassembled Venus and mCherry.
Prolines are depicted as spheres.
reassembled Venus
mCherry
2
3
11
10
7
8
9
4
5
6
1
Pro Pro Pro
2
3
11
10
7
8
9
4
5
6
1
2
3
11
10
7
8
9
4
5
6
1
Pro Pro Pro
Pro187
Pro192
Pro196
Pro186
Fig. 28 Model of reassembly mechanism of fluorescent protein.
oligomer
metastable conformation VN155 VC155 stable conformationmisfold
disaggregation coexpression in E. coliConclusion
Bimolecular fluorescence complementation (BiFC) assay has been used
widely to visualize protein-protein interactions in cells. However, there is a problem
that fluorescent protein fragments have an ability to associate with each other
independent of an interaction between proteins fused to the fragments. To facilitate the
BiFC assay, the author attempted to determine the structure and characteristics of
reassembled fluorescent protein, Venus.
The author succeeded to coexpress of N- and C- terminal fragments of
fluorescent protein, Venus. In the coexpression using E. coli, there were two distinct
thermodynamically stable and metastable forms, monomer and oligomer of
reassembled Venus. The author also succeeded to identify these distinct components by
use of MALDI-TOF MS, DLS measurements, CD spectroscopy, DSC analysis and
X-ray crystallography. Crystal structure of reassembled Venus was determined at 2.1 Å
resolution. It had an eleven-stranded β-barrel fold, typical of GFP-derived fluorescent
proteins.
Despite the overall structure of monomeric reassembled Venus was quite
aggregation including oligomerization is said to occur by specific intramolecular
associations involving the recognition of a sequence partner in another molecule rather
than in the same molecule during the folding process. The author proposed that
cis-trans proline isomerization caused β-strand swapping during folding process. Thus,
it can be expected that replacing a proline with another amino acid depresses the
formation of oligomers. The interface plays an important role in the complementary
association of two non-fluorescent N- and C-terminal fragments. Based on the
structural features, the author mutated amino acids adjacent β7 and measured Tm
values. The results have clearly showed that the mutation was susceptible to thermal
stability of reassembled fluorescent complex. Thus, the substitution of amino acids in
four β-strands (β8, β9, β10 and β11) of the C-fragment and three β-strands (β3, β4 and
β7) of N-fragment is also expected to facilitate the application of BiFC to research of
protein-protein interactions. The crystal structure of the monomer of reassembled
Venus including water molecules provides new insights into decreasing background
References
M. Bergdoll, M.H. Remy, C. Cagnon, J.M. Masson, P. Dumas,
Proline-dependent oligomerization with arm exchange. Structure. 5 (1997) 391-401.
W. DeLano, The PyMOL Molecular Graphics System. DeLano Scientific, San
Carlos, CA. http://www.pymol.org., (2002).
P. Emsley, K. Cowtan, Coot: model-buildings tools for molecular graohics.
Acta Cryst. D. 60 (2004) 2126-2132.
J.Y. Fan, Z.Q. Cui, H.P. Wei, Z.P. Zhang, Y.F. Zhou, Y.P. Wang, X.E. Zhang,
Split mCherry as a new red bimolecular fluorescence complementation system for
visualizing protein-protein interactions in living cells. Biochem. Biophys. Res.
Commun. 367 (2008) 47-53.
I. Ghosh, A.D. Hamilton, L. Regan, Antiparallel leucine zipper-directed
protein reassembly: Application to the green fluorescent protein. J. Am. Chem. Soc.
122 (2000) 5658-5659.
W.T. Heller, H.M. O’Neill, Q. Zhang, G.A. Baker, Characterization of the
influence of the ionic liquid 1-butyl-3-methylimidazolium chloride on the structure and
C.D. Hu, Y. Chinenov, T.K. Kerppola, Visualization of interactions among
bZIP and Rel family proteins in living cells using bimolecular fluorescence
complementation. Mol. Cell. 9 (2002) 789-798.
C.D. Hu, T.K. Kerppola, Simultaneous visualization of multiple protein
interactions in living cells using multicolor fluorescence complementation analysis.
Nat. Biotechnol. 21 (2003) 539-545.
K.P. Kent, W. Childs, S.G. Boxer, Deconstructing green fluorescent protein. J.
Am. Chen. Soc. 130 (2008) 9664-9665.
T.K. Kerppola, Design and implementation of bimolecular fluorescence
complementation (BiFC) assays for the visualization of protein interactions in living
cells. Nat. Protoc. 1 (2006) 1278-1286.
Y. Kodama, C.D. Hu, An improved bimolecular fluorescence
complementation assay with a high signal-to-noise ratio. Biotechniques. 49 (2010)
793-805.
T. Nagai, K. Ibata, E.S. Park, M. Kubota, K. Mikoshiba, A. Miyawaki, A
variant of yellow fluorescent protein with fast and efficient maturation for
cell-biological applications. Nat. Biotechnol. 20 (2002) 87-90.
M. Ormö, A.B. Cubitt, K. Kallio, L.A. Gross, R.Y. Tsien, S.J. Remington,
Crystal structure of the Aequorea victoria green fluorescent protein. Science. 273
C. Ottmann, M. Weyand, A. Wolf, J. Kuhlmann, C. Ottmann, Applicability of
superfolder YFP bimolecular fluorescence complementation in vitro. Biol. Chem. 390
(2009) 81-90.
Z. Otwinowski, W. Minor, Processing of X-ray diffraction data collected in
oscillation mode. Methods Enzymol. 276 (1997) 307-326.
A. Rekas, J.R. Alattia, T. Nagai, A. Miyawaki, M. Ikura, Crystal structure of
Venus, a yellow fluorescent protein with improved maturation and reduced
environmental sensitivity. J. Biol. Chem. 277 (2002) 50573-50578
S. Sakamoto, K. Kudo, Supramolecular control of split-GFP reassembly by
conjugation of β-cyclodextrin and coumarin units. J. Am. Chem. Soc. 130 (2008)
9574-9582.
N.C. Shaner, R.E. Campbell, P.A. Steinbach, B.N.G. Giepmans, A.E. Palmer,
R.Y. Tsien, Improved monomeric red, orange and yellow fluorescent proteins derived
from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 22 (2004) 1567-1572.
X. Shu, N.C. Shaner, C. A. Yarbrough, R.Y. Tsien, S.J. Remington, Novel
chromophores and buried charges control color in mFruits. Biochemistry. 45 (2006)
9639-9647.
Y.J. Shyu, C.D. Suarez, C.D. Hu, Visualization of AP-1-NF-κB ternary
complexes in living cells by using a BiFC-based FRET. Proc. Natl. Acad. Sci. U S A.
105 (2008) 151-156.
E. Takács, O. Barabás, M.V. Petoukhov, D.I. Svergun, B.G. Vértessy,
Molecular shape and prominent role of β-strand swapping in organization of dUTPase
oligomers. FEBS Lett. 583 (2009) 865-871.
A. Vagin, A. Teplyakov, MOLREP: an automated program for molecular
replacement. J. Appl. Cryst. 30 (1997) 1022-1025.
M. Winn, G.N. Murshudov, M.Z. Papiz, Macromolecular TLS refinement in
REFMAC at moderate resolutions. Methods Enzymol. 374 (2003) 300-321.
F. Yang, L.G. Moss, G.N. Phillips, The molecular structure of green
List of publication
Masami Isogai, Yoshihiro Kawamoto, Kazuto Inahata, Harumi Fukada, Kenji
Sugimoto and Toshiji Tada, Structure and characteristics of reassembled fluorescent
protein, a new insight into the reassembly mechanism, Bioorg. Med. Chem. Lett., In
Acknowledgements
The author expresses gratitude to Dr. Toshiji Tada, Professor of Graduate school
of Science, Osaka Prefecture University, for his kind guidance, invaluable advice and
stimulating discussion on this work.
The author wishes to thank Dr. Satoru Tokutomi and Dr. Ikuo Fujii, Professors of
Graduate school of Science, Osaka Prefecture University, for their helpful advice and
critical reading of this thesis.
The author also wishes to thank Dr. Kenji Sugimoto, Professor of Graduate
school of Life and Environmental Sciences, Osaka Prefecture University, for providing
materials and helpful suggestions.
Thanks are due to Dr. Takayoshi Kinoshita, Associate professor of Graduate
school of Science, Osaka Prefecture University, and Dr. Maki Onda, Assistant
professor of Graduate school of Science, Osaka Prefecture University, for their helpful
suggestions and observations.
The author is grateful to Dr. Harumi Fukada, Associate professor of Graduate
school of Life and Environmental Sciences, Osaka Prefecture University, Osaka
Prefecture University, for her kind cooperation.
Thanks are also due to Dr. Shigenori Nishimura, Assistant professor of Graduate
school of Life and Environmental Sciences, Osaka Prefecture University, for his kind
The author would like to thank the beamline staff at KEK-PF BL5A for
assistance in the synchrotron experiments.
Finally, special thanks are due to all members of Laboratory of Structural
Biology, Department of Biological Science, Graduate school of Science, Osaka