Title
環境配慮型水路における魚類相の変化( 本文(Fulltext) )
Author(s)
森, 須美子
Report No.(Doctoral
Degree)
博士(農学) 甲第610号
Issue Date
2013-09-10
Type
博士論文
Version
ETD
URL
http://hdl.handle.net/20.500.12099/47821
※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。環境配慮型水路における魚類相の変化
2013 年
岐阜大学大学院連合農学研究科
生物環境科学
(岐阜大学)
森 須美子
目 次 1 はじめに ... 1 2 調査対象地域について ... 3 3 湿地帯の代替環境として整備された環境配慮型水路における調査 ... 5 3.1 地区の概要 ... 5 3.2 調査方法 ... 19 3.3 調査結果 ... 20 3.3.1 水質計測の結果 ... 20 3.3.2 採捕調査結果 ... 26 4 汎用化水田における影響 ... 39 4.1 地区の概要 ... 39 4.2 調査方法 ... 41 4.3 調査結果 ... 43 4.3.1 水路内の環境 ... 43 4.3.2 魚類の生息状況 ... 46 5 ミティゲーションの検討 ... 56 6 まとめ ... 60 7 今後の展望 ... 62 謝辞 ... 64 参考資料 ... 67
1
1 はじめに
生産性向上に向けた圃場整備により,水田の乾田化が進み,用排水路が分離され田面と 水路の連続性が失われるなど我が国の水田生態系は大きく変化してきた(中川,2000;守 山,1997;片野,2002;森,2008).その変化は田面のみならず,水路やため池,小河川 といった水系ネットワーク全体で生じている.とくに低平水田地帯に多く見られた湿地帯 やクリークは,その多くが埋め立て等によって失われている.本調査地においても,魚類 をはじめ,鳥類や哺乳類,両生類など多くの生物が生息する良好な湿地帯が県道整備によ って埋め立てられ,湿地帯と隣接していた水田の汎用化整備も行われた.そこで,湿地帯 と連続する幹線排水路を生態系に配慮した形で整備することとなった. これまで,水路の改修前後における生態系の変化をモニタリングした研究が多く行われ てきた(佐藤・東,2004;堀野ら,2008).しかしながら,代替地を設けての生態系全体の 移設をモニタリングした例は少ない.そこで,本研究では,消失する湿地生態系の代替地 として整備された環境配慮型水路において,どのように生物が移動し,定着していくかに ついて魚類を中心に調査を行い,湿地生態系の代替地として排水路をどのような形で整備 していくべきかについて検討した. また,水田の乾田化が進む要因のひとつとして,水田の汎用化整備があげられる.汎用 化水田では,畑作物の栽培が可能となり,高い生産性が得られるようになるが,そのため には,暗渠を設置するなど圃場内の排水性を高めなければならない.そのため,従来の圃 場整備による排水路(一般に,深さ約60cm)よりもさらに深く掘り下げられ(深さ約 90cm), 田面との落差が大きくなるだけではなく,水田の乾田化が進むことになる.さらに,圃場 整備を実施する際に,客土が行われる場合が多く,表土扱いなどの工法によって,非作付 け期の畦畔や田面を利用して越冬する両生類への影響が懸念される.また,畑作物を作付 けすることにより,圃場及び水路などの水域が時間的にも空間的にも減少することが挙げ られる.しかし,このような汎用化水田は今後も増加していくことが予想される. 水田あるいは農業用水路での生息状況は,種あるいは生活史によって異なるが(斉藤ら, 1988),その環境が変化すれば,そこに生息する生物に大きな影響が及ぶことは容易に推察2 できる.たとえば,学名が稲の周りにいるヒレの広い魚という意味を持つ水田環境を代表 する生物のひとつであるミナミメダカは,平成19 年の環境省の報道によるレッドリストで 絶滅危惧Ⅱ類に指定されるほど減少している.ミナミメダカが減少した原因の一つとして, 水田と水路の間をミナミメダカが自由に移動できなくなった環境があげられる(上月ら, 2000;小澤,1999).前述したように,汎用化水田では田面と水路の落差が増大するため, 物理環境はさらに悪化することが予想されるが,汎用化による水田の高度利用についての 研究は行われているが(若杉ら,2009;齋藤ら,2010)汎用化された水田地帯での物理環境 の変化が,そこに生息する魚類に与える影響について検証した例は少ない. そこで本研究では,汎用化水田に麦など畑作物を導入することによる水管理の変化が, 排水路内の環境と魚類生息の状況にどのような影響があるのかについて検討した.また, その結果をもとに生息環境の保全方法についても検討した.
3
2 調査対象地域について
濃尾平野は愛知県北西部から岐阜県南西部に位置する,木曽川・長良川・揖斐川の 3 河 川による沖積平野である.この西部から南部にかけては低湿であり,水害防止のために町 全体を堤防で囲う輪中地帯となっている(図 1).この輪中は鎌倉時代以降本格的に形成され, 様々な治水工事により 3 河川が分離された.しかし,輪中堤で囲うことによって水害は減 少しても,これらの河川が濃尾平野よりも河床が高い天井河川であったために,地域内に 水が地下水として流れ込み,泥田と化してしまった.その後,土地改良がなされ,湿田の 乾田化が進められ,現在では野菜のハウス栽培も行われている. 調査地はこの輪中地帯の南西部に位置する岐阜県安八郡輪之内町である.平均標高2.5m 前後,総面積2,236ha うち耕地面積 1,150ha の低平地である.輪中内は流れの緩やかなク リーク状の水路が縦横に走り,堀田と呼ばれる特有の環境が残っていたことから(輪之内町, 1996),小型の淡水魚をはじめとして多くの生物が生息していた.一方,排水条件が悪く, 圃場の区画が狭いなど営農には多大な労力を要していたことから,昭和40 年代より圃場整 備が順次行われてきた.現在でも,耕地面積のうち 88.7%を水田が占めるが,その整備状 況は圃場整備以前の用排水路兼用水田地域,近年までの圃場整備である用排水路分離水田 地域,汎用化水田地域が混在している.水管理は各農家が行っており,用排水路分離水田 や汎用化水田の地域でも灌漑期間中は排水路の水位を畦畔と同様の高さまで高く保つ地区 もみられる. 輪中堤に囲まれた閉鎖的水域であり,取排水河川ともに揖斐川を利用している.取水は ポンプ,排水はポンプ及び排水樋門によって行われている.一部の地域で地下水が利用さ れている.また,町内には 5 本の河川が流れており,揖斐川より取水された水は主に農業 用水として利用され,東江川,中江川,中西江川,西江川にそれぞれ排水として注ぎ込み, 大榑川と合流し,最終的に揖斐川に排水される.また,一部で農業用水の反復利用が行わ れている.4 図1 輪之内町全体図 N S E W http://www.tanpopo.ne.jp/~niki/ N W S E
5
3 湿地帯の代替環境として整備された環境配慮型水路における調査
3.1 地区の概要 調査を行った環境配慮型水路は,県道整備およびその周辺の圃場整備により埋め立てら れることとなった湿地帯の代替環境として道下地区に整備された(図 2).環境配慮型水路 として整備されたのは,水路長約 600m,水路面積約 1,100 ㎡,ビオトープ池の面積は約 220 ㎡である.圃場は汎用化整備が行われており,各圃場へはパイプラインにより配水され ており,用水からの生物の供給はほとんどないと考えられる.また,水路内の水位の調整 は,地点4(ビオトープ池)の河川側に設置されている起伏式のゲートと,水路下流端に設置さ れている水門の2 つによって行われている.主にビオトープ池のゲートで水位の調整を行い,中干 し期など排水量が大きい場合には水路下流端の水門を併用している.なお,環境配慮型水路に おける各構造物は,水門操作による水位調節の状況を農家から聞き取り,年間の水位変化を想定 したうえで設計されている.埋め立て前,埋め立て後の湿地帯および各調査地点の状況を写真 1 に示す. 埋め立てられた湿地帯は面積約 22,000 ㎡であり,以前は堀田と呼ばれる水田であった.現在 は,県道及び用排水路が分離された汎用化水田となっている.そのため,素掘りの水路が縦横に 走り,その周辺に圃場整備の行われていない用排兼用水路の水田が広がっていた.水田の畦畔 は低く,植生が繁茂し,灌漑期には水田と水路が連続した水域となる環境であった.また,水路の 両岸や湿地帯周辺にはヨシやマコモ帯が広がっており,魚類だけでなく,多くの鳥類やカヤネズミ などの哺乳類,それにナゴヤダルマガエルをはじめとするカエル類などが生息しており,生物にと って良好な環境であった.また,水際の傾斜も様々で,水位変化が生じても常に水際植生が存在 し,浅瀬や深みなど多様な環境を有していた.この湿地帯が県道建設のため埋め立てられたのは 平成 17 年度の非灌漑期である.6
7 調査は,水域の種類や水路構造から5 つの調査地点を選定し,各地点で調査を行った(図 2).各地点の概要を表 1 に示す.これらの調査地点を含む水路や池の整備は,平成 14 年度 から行われ,平成14 年度に地点 1~地点 3,平成 15 年度に地点 5,平成 16 年度に地点 4, 平成17 年度に湿地帯の埋め立て,平成 18 年度に地点 1 の改修が行われた.いずれも作付 けに影響のない非灌漑期に工事が実施された.なお,湿地帯に生息する魚類が水路へ移行 する期間として約2 年間設けられたが,人為的な生物の移動は行われなかった. 湿地帯の埋め立て前後の状況を写真 1 に示す.改修前は灌漑期になると田面と水路の境 がなくなり,そこに生息する生物が田面と水路を自由に移動していた. 表 1: 調査地点の概要 水域の 種類 護岸 水路床 水面 地点 1 水路 コンクリート 砂泥 地点 2 水路 階段状護岸 砂泥 地点 3 水路 階段状護岸 砂泥 花いかだ 地点 4 池 階段状護岸 砂泥 地点 5 水路 ソダ護岸 砂泥
8
改修される前の湿地帯
改修後の湿地帯
9 地点 1 は,調査年度によってその構造や環境が大きく変わっている.その変化を写真 2 に示す.平成14 年度に,もともと土水路であった水路の法面を整地したが,護岸の整備や コンクリート化は行われなかった.その後,水路内や法面にマコモやスズメノヒエなどの 植物が侵入し,魚類の産卵床や避難場所となる環境が形成された.平成 18 年度の調査は, このような土水路の状態で行ったが,その年の非灌漑期に改修が行われ,コンクリート 2 面張りの水路となった.よって,平成19 年度は,同じ地点ではあるがコンクリート 2 面張 り水路において調査を行った.
10
平成18 年度調査時の状況
平成19 年度調査時の状況
11 地点2 は,護岸を階段状にし,各段に植生を繁茂させることにより,多様な水際環境が 創出できるよう整備された地点である(写真 3).水路底から高さ 60cm まではコンクリート 壁となっているが,それより上部は約20cm の段差で階段状になっており,各段に植生が繁 茂することにより,想定された水位変化に対して常に水際植生が存在する構造になってい る.また,非灌漑期の水深確保のため,この地点から下流は上流部と比較して水路底が30cm 低く掘り下げられている.
12
整備直後の地点2
整備後3 年が経過した地点 2
13 地点3 は,護岸と水路床は地点 2 と同様の構造だが,水面に 1m×1m の花いかだを 3 つ 連結させてロープで係留し,水面に浮かべた構造である(写真 4).花いかだは設置が容易で, 水面に浮遊しているため,水位に関わらず常に水路内植生を設けることができる.また, 鳥類など地上の天敵からの避難場所ともなる.なお,花いかだには,主としてマコモを植 栽した.写真4 からもわかるように,設置から 2 年後には当初植栽した植物はなく,外来 種が生育している.3 年後には生育する植物も減少し,浮遊物が漂着していることがわかる.
14
花いかだ設置 2 年後の状況
花いかだ設置3 年後の状況
15 地点 4 は,水路横に設けられたビオトープ池である.排水路とつながっており,排水時 や低水位となった場合の避難場所として,また,止水域を好む生物の生息場所として設け られている.池内部は緩やかな勾配の階段状護岸になっており,灌漑期に水際となる位置 にマコモを植栽した.池底は,常に水深が確保されるよう掘り下げられている.なお,水 路との連結部分には水門等生物の移動を阻害するものは設置されていない.写真 5 に水位 の変化にともなうビオトープ池の様子を示す.
16
低水位時のビオトープ池
高水位時のビオトープ池
17 地点 5 は,水路左岸側がソダ護岸となっている.本調査地のソダ護岸は,主にソメイヨシノの剪定 枝を束ねて木枠で固定したものであり,高さ 100cm,幅 30cm となっている.写真 6 からわかるよう に,ソダは水路底から鉛直方向に積み上げられているため,水路の水位が低下した場合にも,常 にソダによる空隙が水中に存在する.また,水位上昇時には,ソダ護岸上部の水路法面に繁茂す るスズメノヒエなどの植生が一部水没し,水際植生となっている.しかし,植生は水路法面上部にし かないため,水深が 100cm以下になると水際植生は失われてしまう.
18
左岸側に設置されたソダ護岸
高水位になり水路に使っているソダ護岸
19 3.2 調査方法 各地点において,サデ網(目合2mm,開口 95cm)とタモ網(目合 2mm,開口 35cm), カゴ網を用いて魚類を採捕し,種の同定と個体数・体長の計測を行った.採捕した魚類は, 採捕地点に放流した.採捕は3 人 1 組を基本とし,追い込み距離は約 3m,採集面積 6 ㎡と し,採集時間は2 分を目安とした.湿地帯でも同様の方法で調査を実施した. 湿地帯の調査は平成17 年度の 5 月から 1 月にかけて,水路と池の調査は,平成 18 年度 と平成19 年度の 4 月から 1 月にかけて行った.平成 17 年度は 10 回,平成 18 年度は 9 回, 平成19 年度は 5 回実施した.なお,調査期間を,4 月から中干し(7 月から 8 月にかけて) 前までの灌漑前期,中干し期間,中干し後から9 月末の落水(平成 18 年度は 10 月中旬) までの灌漑後期,落水後の非灌漑期の4 期にわけ,各期間に 1 回以上の調査を実施した. 表 2 に調査実施日を示す. 各調査地点において,多項目水質計(TOADKK 製 WQC-24)による水質計測と,水深を 測定した.測定項目は,水温,pH,DO,EC の 4 項目である.なお,流速についても測定 したが,調査中は常に止水状態であった.また,植生の状況など,水路内の環境について も記録した. 表 2: 調査実施日 平成 18 年度 平成 19 年度 灌漑前期 4/17,5/30,6/29 4/21,5/30 中干し 7/27 8/8 灌漑後期 9/11,10/10 9/25 非灌漑期 11/9,12/6,1/18 12/18
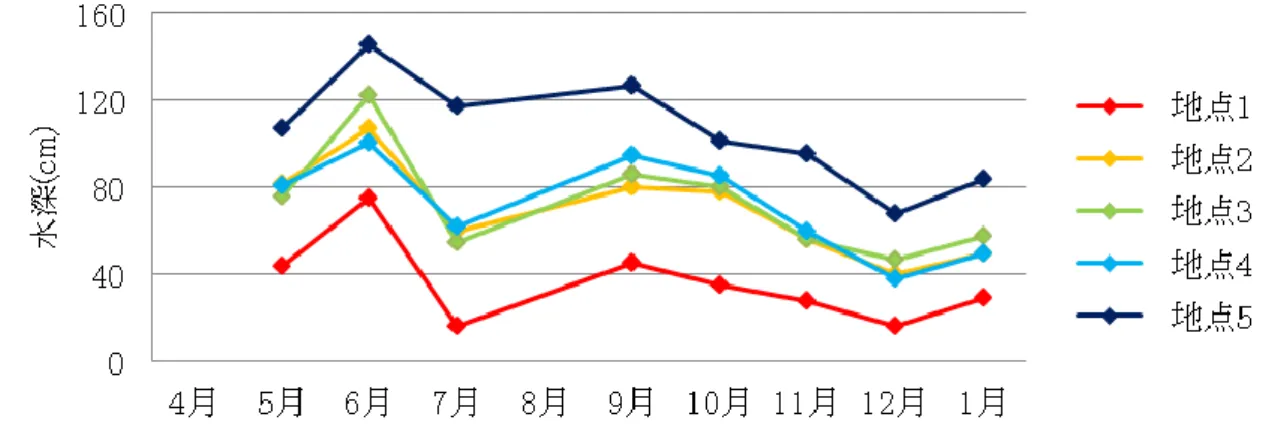
20 3.3 調査結果 3.3.1 水質計測の結果 水質の計測は,水深約 30cm までの深さで,午前中に行った.2 年間の計測結果を比較すると 水深と水温に大きな差がみられた.その他の項目については,地点間における差はみられな かった.年度による差も見られなかった. (1) 水深 水深の変化を図3・図 4 に示す.両年度とも地点により水深は異なるが,取水や落水とい った水田での作付けによる影響が水深の変動に大きく影響している.平成19 年度には,す べての地点で最高水深が平成 18 年度よりも低く,年間を通して低い水深で変動していた. その年の気候や作付け,生育状態により水深は変動してくるが,この環境配慮型水路の集 水域には汎用化水田が整備されているため,平成19 年度のような低い水深で変動すること が多いと推測される.
21
図3 平成 18 年度の水深
22 (2) 水温 平成 18 年度の水温の変動 7 を図 5 に示す.冬期(12 月~1 月)の地点 1 の平均水温が 6. 6℃,地点 2~5 の平均水温が 10.7℃と地点 1 は他の地点と比較して約 4℃低い.冬期でも 水深 40cm を確保できる他の地点と異なり,地点 1 では冬期の水深が 16.0cm になることも ある.水深の確保は水温の安定維持にも必要であると考えられる. 図 5 水温の年間変動
23 (3) pH pHは灌漑期の平均値が 6.68,非灌漑期のそれが 7.34 と非灌漑期に高くなる傾向が見られた (図 6).1 年度通してpH7 前後で変動しており,魚類の生息に影響を与える数値にはならなかっ た. 図6 pH の年間変動
24 (4) EC(電気伝導度) ECについても灌漑期が 204.5μS/cm,非灌漑期が 236.0μS/cm と非灌漑期が高くなる傾向が みられた(図 7).非灌漑期には水路内の水量が少ないため,希釈効果が低くなり高い値を示すこと になると考えられる. 図 7 EC の年間変動
25 (5) DO(溶存酸素) DO は水温が 25℃以上となった 6~9 月にかけての平均値が 1.3mg/l と低く,水温が 20℃を下 回った 10~1 月の平均値が 4.3mg/l と高くなる傾向を示した(図 8).調査対象とした水路には流れ がほとんどなく,水温の上昇により水中のプランクトンの活動が活発化したために水中の酸素濃度 が低くなったと考えられる.この結果から,水路環境による影響はほとんどないと考えられた. 図 8 DO の年間変動
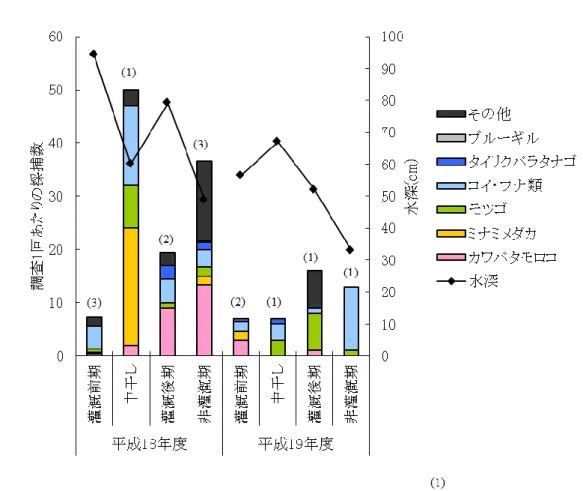
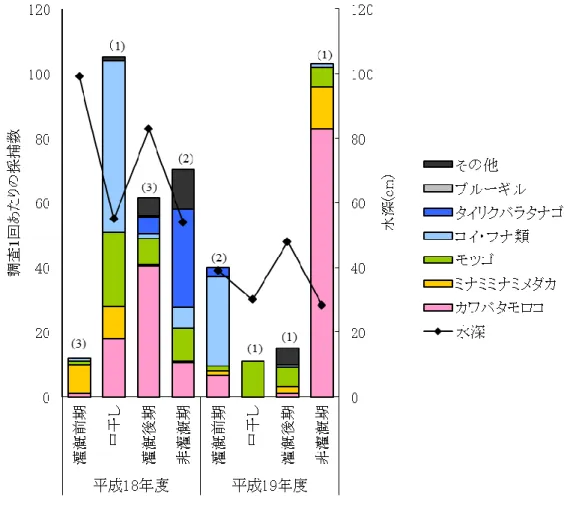
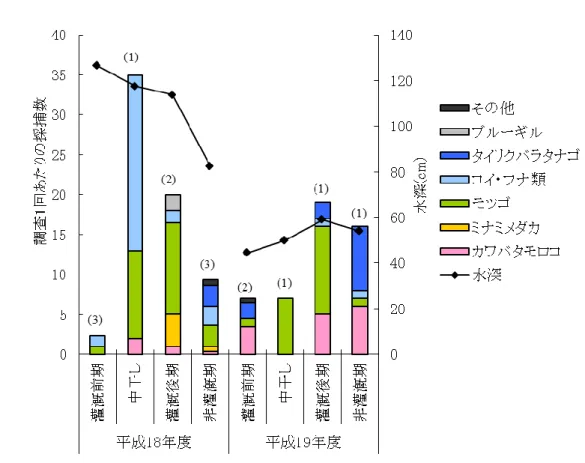
26 3.3.2 採捕調査結果 各地点における調査1回あたりの採捕数を表 3に示す.水田生態系を代表する魚種であるミナミ メダカ、カワバタモロコ,ギンブナ,モツゴといった遊泳魚は多く確認されたが,ドジョウなど底生魚 は少数しか確認できなかった.さらに,外来種であるブルーギルも確認された.平成19年度には, 地点4以外で魚種が減少し,すべての地点で調査1回あたりの採捕数が減少していたが,その理由 は調査地点ごとに異なっていた. なお,各地点での調査結果では,コイとギンブナをコイ・フナ類,ドジョウ・ツチフキ・ヨシノボリ類・ ワタカ・モロコ類をその他の種として表記した.
27 表 3: 調査 1 回あたりの採捕数 湿地帯 地点 1 地点 2 地点 3 地点 4 地点 5 H17 H18 H19 H18 H19 H18 H19 H18 H19 H18 H19 採 捕 数 カワバタモロコ 10.2 11.4 4.2 6.8 1.4 14.9 19.4 9.7 11.8 0.6 3.6 ミナミメダカ 11.9 9.7 1.2 3.1 0.6 4.3 3.6 9.7 0.4 1.1 0 モツゴ 119.4 5.4 1.6 1.9 2.2 9.7 5.2 3.1 11.8 5.0 4.2 ギンブナ 33.3 8.9 1.6 5.1 3.4 8.0 1.4 3.6 1.2 2.1 0.4 コイ 1.0 1.8 0 0.1 0.6 0.7 10.0 1.1 0.8 1.8 0 タイリクバラタナゴ 9.1 3.9 0.2 1.0 0.4 11.2 1.2 1.4 1.6 0.9 2.8 ブルーギル 0 0 0 0.1 0 0.1 0 0.2 0.2 0.4 0 ドジョウ 0.1 0.3 0 0 0 0 0 0 0 0 0 ツチフキ 0.3 0 0 0 0 0 0 0 0 0 0 ヨシノボリ類 0 0 0 0 0 0 0 0.2 0.4 0 0 ワタカ 1.4 0 0 0.1 1.0 3.0 0 0.7 3.4 0 0 モロコ類 3.7 1.3 1.4 6.3 0.4 3.4 1.0 4.3 7.4 0.2 0.2 種数 10 8 6 10 8 9 7 10 10 8 5 調査回数 10 9 5 9 5 9 5 9 5 9 5
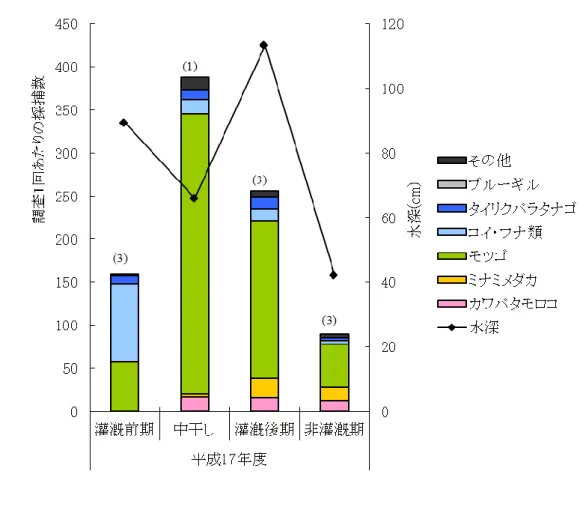
28 (1) 湿地帯 湿地帯での採捕結果を図 9 に示す.ドジョウやツチフキといった底生魚を含む 10 種の魚 類が採捕され,各魚種の採捕個体数も多く,湿地帯が魚類の生息に良好な環境となってい たことがわかる.また,埋め立て時には,以前に放流されたと思われる多くのウナギを目 視にて確認した.湿地帯は平成17 年度の作付け後に埋め立てられたため,平成 17 年度の 調査結果である. 灌漑前期にコイ・フナ類が多く確認されているが,多くは仔稚魚であり灌漑前期に湿地 帯で孵化した個体であると思われる.このような一時的に増加する種ではなく,1 年を通し て多く捕獲されている種がモツゴであり,モツゴはこの湿地帯の優占種であるといえる. 写真1 からもわかるように,湿地帯では灌漑期になると田面と水路の境がなく水路内には ヨシが繁茂していたため,このような環境を好むモツゴが優占種となったと推測される.
29
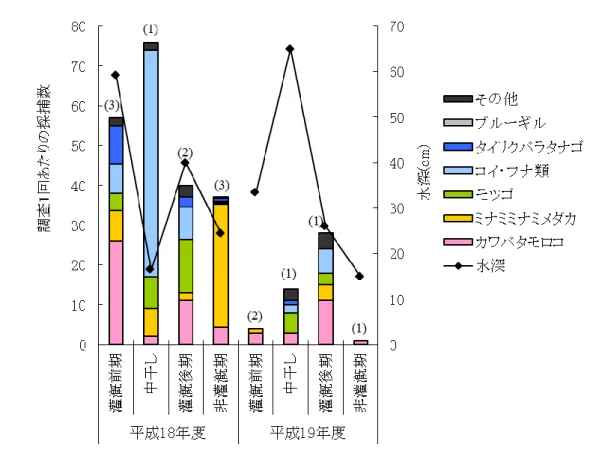
30 (2) 地点1(コンクリート護岸) 地点1における採捕結果を図 10に示す.改修前の平成18年度は,法面だけでなく水路床から 植物が繁茂しており,中干しや非灌漑期の水深が浅い時期でも魚類の産卵床や避難場所となる 環境が整っていた.その結果,ミナミメダカやカワバタモロコなど多くの魚種が採捕され,その仔稚 魚も確認された.コンクリート化された改修後は,種数・個体数ともに減少している.また,中干しや 非灌漑期に確認されていたドジョウが,平成19年度にはどの時期にも確認できなかった. 平成19年度にはミナミメダカやギンブナで仔稚魚が採捕できず,カワバタモロコで1個体採捕さ れただけであったことから,この地点で繁殖が行われていないことが推定される.改修工事により, 魚類の産卵床となる水路法面や水路内の植生が消失したことが,魚類の生息状況に大きな影響を 及ぼしているといえる.
31
32 (3) 地点2(階段状護岸) 平均水深は平成18年度が69cm,平成19年度が57cmであった.平成18年度は非灌漑期でも水 深が60cm以上となることがあり(調査9回中4回),階段状護岸に移植された植生帯を水生生物が 利用できる環境であった.一方,平成19年度は5回行った調査のうち,水位がコンクリート壁天端を 越えたことが1度しかなく,植生のないコンクリート2面張りの水路と同様の単純化された環境となる 期間が長く続いた.これは,事前の聞き取り調査から想定していた排水路水位よりもさらに低い水 位で管理されていたことによるものである. 採捕調査の結果,平成18年度には10種,19年度には8種と他の調査地点と比較して比較的多く の種が確認された(図 11).しかし,平成19年度には,種数は多いものの仔稚魚を中心に採捕数 が著しく減少した.このように,水位差にして僅か10cm程度であるが,水際植生の有無によって繁 殖状況および生息魚数が大きく変化するため,水位変化に対して余裕を持った施設設計を行う必 要がある.
33
34 (4) 地点3(花いかだ) 地点2と同様平成19年度には,水深60cm以上となることがほとんどなく,護岸に水際植生帯が形 成されることがほとんどなかったが,花いかだは水面に浮いているため,水位低下の影響はある程 度回避できると予測された. 採捕調査の結果(図 12),水位の低下した平成19年度の非灌漑期に多くの個体が採捕された. 花いかだが投入されたのは平成16年度である.1年目には当初植栽されたマコモが繁茂し,2年目 にはマコモが消失し代わってアメリカセンダングサなどの外来種が繁茂していた.3年目には2年目 に侵入した外来種も多くが消失してまばらになり,4年目となる平成19年度には植物が枯死してしま った.そのため,生息魚類が減少したと考えられる.一般に,植物の水中根は生息空間として利用 されるだけでなく,餌資源として採食されてしまうため,草勢は次第に衰えてくる.一方,それに置き 換わる形で外来種を中心とした植生が繁茂するが,同様に採食されて枯死し,最後には植生が消 失する. 非灌漑期におけるカワバタモロコの体長分布を図 13に示す.平成19年度に仔稚魚を含む80個 体以上が採捕されていることから,餌資源としての水中根が消失しても水面を被覆する機能が鳥類 など地上の天敵からの避難場所として,水中に生息する魚類にとって重要な環境条件となってい るといえる.花いかだは護岸植栽と比較して素早く植生帯を設けることができるという長所があるが, 環境配慮の面からは,一時的ではあるものの外来種の侵入は喜ばしいことではなく,花いかだの 維持には定期的な管理が必須である.また,花いかだを係留するロープにゴミがたまりやすく,植 生の維持と同様,維持管理作業が欠かせない. 平成19年年度の調査で,灌漑前期にコイ・フナ類が多数捕獲されている.これは,5月調査時に 10mm~15mmのコイの稚魚が50個体捕獲されたためである.5月調査の2週間前に,同地点で二 枚貝の捕獲調査を行った際に花いかだに多数の魚卵が付着していることが確認されており,この 卵が孵化したものと思われる.卵確認時,花いかだには植生は繁茂しておらず,花いかだのネット および漂流物に付着していた.このことから,花いかだに植生が繁茂していなくても,魚類の産卵 場,生育場となることが可能であることが推測される.
35
図 12 地点3における採捕結果(図内のカッコ書きは調査回数)
36 (5) 地点4(ビオトープ池) 当初,池にはマコモ帯が形成される予定であったが,実際には植生のない状態となり,水中に緑 藻類が繁茂していた.これは,階段状護岸の場合と同様,計画水位と実際の水位が異なっていた ためである.高水位時に水際となると考えられた位置にマコモ帯を植栽したが,実際には計画より も高い位置まで水位が上昇したため(写真5参照),マコモが活着する前に腐敗してしまった. 採捕調査の結果,植生帯はなかったものの,両年とも10種の生物が確認され,多くの魚類が利 用していることがわかる(図 14).採捕された魚種のほとんどで仔稚魚も確認できた.これは,先に 述べた緑藻類が繁茂したことによる影響が大きいと考えられるが,マコモなど湿性環境に生育する 植物を移植することにより,さらに魚類の産卵床や避難場所などを提供するものと考えられる.また, 個体数は少ないが,唯一ヨシノボリ類が確認された地点でもある. 図 14: 地点4における採捕結果(図内のカッコ書きは調査回数)
37 (6) 地点5(ソダ護岸) 平成18年度の灌漑期には水深が100cmを超えることもあり,ソダ護岸上部の水路法面の植生により 水際植生帯が形成され,魚類の産卵床や避難場所を有する環境となっていた. 採捕調査の結果,環境配慮型水路の他地点とは異なり,モツゴが優占種となっていた(図 15). もともと湿地帯においてモツゴは優占種であったため,最も近い位置にあるこの地点がその影響を 強く受けた可能性がある.平成18年度には,他の地点と比較してモツゴの仔稚魚が多く確認され, この地点での繁殖の可能性が示唆されたが,翌19年度には仔稚魚が1個体も採捕できなかった. 他地点よりモツゴの採捕率は高いため,モツゴが選択的にソダ護岸に生息している可能性は大き いが,産卵床として適しているかは不明である. この水路では同時にブルーギルの仔稚魚が確認されている.ブルーギルの生育環境はモツゴ のそれと類似している.ブルーギルの混入により優占種が交代した例(遊磨ら,1997)も報告されて おり,外来種対策が必要な環境である.また,木枠やソダは腐敗するため,定期的な補修が必要 である.
38
39
4 汎用化水田における影響
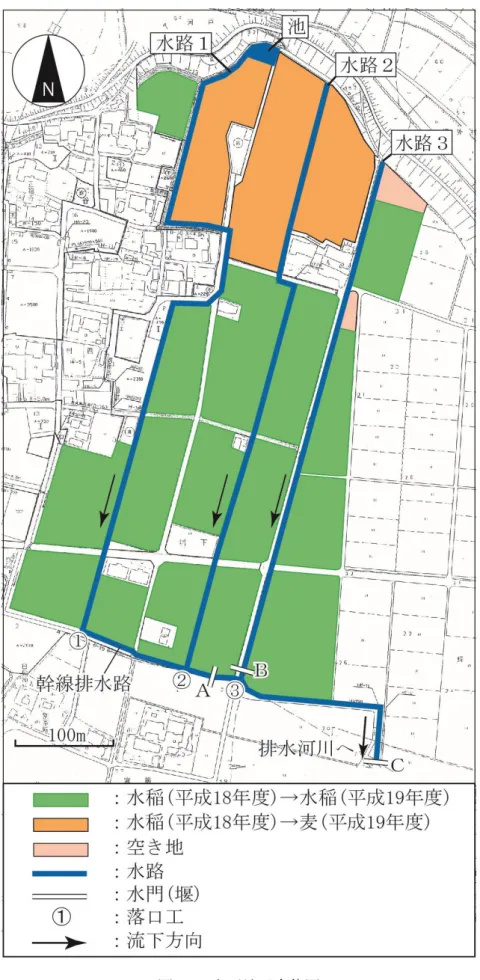
4.1 地区の概要 水田の汎用化を目的に,平成16 年度から 21 年度までの工期で経営体育成基盤事業「輪 之内本戸地区」が実施された.調査対象としたのは,このような圃場整備が進行しつつあ った本戸地区(図16 参照)を南北に走る 3 本の排水路(水路 1~3)と,各排水路が地区 南部を東西に走る幹線排水路と合流する3 地点(①~③),さらに水路 1 の上流部にビオト ープの機能を期待して人工的に設置した「池」(以下,「ビオトープ池」と称す)である. 水路及び水路周辺の概要をそれぞれ表 4 に示す.なお,調査対象地区の上流側(北側)は 輪中堤によって遮断されているため,水路内に生息する淡水魚は,水路内で繁殖したか, あるいは,下流の排水河川から遡上してきた個体に限定される. 平成17 年度の冬期に水田の汎用化を目的として,水路 1 及び 2 とそれに隣接する圃場が 整備された.平成18 年度から畑地転換が行われ,麦が作付けされた. 水路3は,30年以上前に整備された水路で,水路内には砂泥が堆積し,ヨシなどの植生が 繁茂している.また,水田内の小水路(用排兼用水路)と接続しており,平成18,19年度 ともに周辺圃場は水稲が作付けされていた.調査対象とした3本の排水路は幹線排水路に合 流し,さらに水門Cを通して,排水河川に流下している. 表 4: 各水路の概要 水路 1 水路 2 水路 3 合流点① 合流点② 合流点③ ビオトープ池 水路整備状況 用排分離 用排分離 用排兼用 - - - - 水路延長(m) 780 620 550 - - - - 水路幅(m) 0.6~0.9 0.6~0.9 0.6 1.0 1.0 1.0 - 水路の深さ(m) 0.9 0.9 0.6 1.3 1.3 1.5 - 側面 Co※ Co※ Co※ Co※ Co※ Co※ ソダ 石積み底面 砂泥/Co※ 砂泥/Co※ 砂泥/Co※ 礫/Co※ 礫/Co※ 砂泥/Co※ 石積み
砂泥堆積(cm) 0~5 0~10 0~25 - - 0~1 -
40
41 4.2 調査方法 水路内(水路 1~3)の調査地点は,約 100m 間隔に 1 地点程度,とくに水路湾曲部や道路下の日 陰部分を中心に選定した.合流部(①~③)は,写真 7 に示すような落口工の直下流の深み(幅 1. 0m×長さ 1.0m×深さ 0.2m)になっている地点で調査を行った.各調査地点では,サデ網(開口 95cm,深さ 70cm,目合い 2mm)とタモ網(開口 45cm,深さ 45cm,目合い 2mm)を使用して魚類を 採捕し,種の同定,個体数,体長を計測した.採捕の際に追い込む距離は水路フルームの 2 アー ム分(約 1.7m)とし,2 人 1 組で実施した.ビオトープ池の捕獲調査は,カゴ網を使用し,3 人 1 組 になって採集面積 4m2(幅 2m×追い込み距離 2m)を約 1 分間の採集時間で,2 回実施した.採捕 した個体は,測定後元の箇所に放流した.採捕した地点で,水質計(TOADKK 製 WQC-24)によ る水質計測と,水深を測定した.水質の測定項目は,水温,pH,DO,EC の 4 項目である.なお, 無降雨時の排水路内には水の流れがほとんどないことから流速の測定は省略した.また,水路内 の植生や堰の有無など水路内の環境についても記録した. 調査は,平成 18 年と平成 19 年の 4 月から 12 月までの各月 1 回(平成 18 年 8 月は欠測),計 17 回実施した.
42 水路 1,水路 2
水路3
43 4.3 調査結果 4.3.1 水路内の環境 水路内環境の調査結果を表5,各水路の状況を写真 8 に示す.水路内の環境で最も大きく 変化したのは水路1及び水路2 の水深であった.水稲のみが栽培された平成 18 年は,幹線 排水路中に設置された堰A(高さ 0.9m)における堰上げによって灌漑期の水深が 90cm を上回り,排水路内の水深が側壁を超えて畦畔まで上昇していた.その結果,水路内には 植生は無かったものの,畦畔植生が水没して水際に植生が生じていた.平成19 年度は,水 路下流側の圃場で麦の転作が導入されたため,圃場内の地下水位低下を目的として排水路 水位が下げられたものの,魚類保全に配慮して高さ約14cm の堰板が排水路内に約 30m 間 隔で設置され水深の維持が図られた.しかし,排水路内の水深は,暗渠管出口を塞がない ように最大でも30cm を越えないように管理されており,前年度のように畦畔の植生が水没 することは無く,水際植生は確認されなかった.一方,水路 3 では,水門B(高さ 0.65 m)によって灌漑期の水深は60cm 前後で維持され,水面が畦畔植生まで達するだけでなく, 水路内にも植生が繁茂していた. 水質については,転作時における暗渠排水の影響が予想されたが,大きな変化は認められなか った.合流部及びビオトープ池でDO の最小値が水路内よりも高い値を示している.合流部は水路 との落差によって常に水が撹拌される状態であったこと,さらに,ビオトープ池では水面をホテイア オイが繁茂し光合成によって酸素が供給され,DO の最小値を高い値で維持したと考えられる.ま た,水路内の DO 値は無降雨時の水の流れが少ない時期に低くなる傾向が見られた.EC は,中 干し後の8 月に低く,落水後の 11~12 月にかけて高い値を示す傾向が見られた.中干し後の再 堪水時には,水量が増加するために一旦低い値を示すが,収穫に伴う落水後は水路内の水量も 少なくなるために高い値を示したと考えられる. 落口工①~③の 3 地点では水路改修により写真 2 に示すような落差が生じている.それぞれの 落差(下流側水面から上流側水路底までの標高差)の年間平均値は,落口工①で 0.27m,②で 0.33m,③で 0.55m となっており,幹線水路から各水路への遡上は水路内の水位が上昇する豪 雨時を除いて困難であると考えられる.
44 以上のことから,汎用化水田に麦作を導入したことによる影響は,排水路水深の低下及びそれ に伴う水路内植生の消失であり,水質にはほとんど変化がないことが示めされた.また,改修によっ て生じた水路間の落差は,現状では魚類の移動に支障をきたすと考えられるため,簡易的な魚道 を設置するなど何らかの対策が必要である. 表 5: 各地点の環境と水質結果 調査地点 調査年 水深 (m) pH EC (μS/m) DO (mg/l) 水温 (℃) 植生 水路 1 H18 0.04~1.00 6.0~7.8 117~457 0.29~5.95 9.2~28.3 畦畔 H19 0.03~0.29 6.0~7.6 192~310 0.53~4.99 7.9~27.7 なし 水路 2 H18 0.02~0.92 6.0~7.3 94~357 0.20~5.83 7.9~31.3 畦畔 H19 0.01~0.28 6.7~7.5 197~340 0.82~6.05 7.5~27.4 なし 水路 3 H18 0.03~0.66 6.4~7.4 76~296 0.56~6.50 8.6~31.8 畦畔,水路内 H19 0.01~0.67 6.8~7.9 154~255 0.68~5.89 7.1~31.9 畦畔,水路内 合流部① H19 0.21~0.34 6.8~7.7 307~317 2.69~5.83 8.3~27.0 なし 合流部② H19 0.15~0.31 6.7~7.5 271~300 3.57~5.25 11.5~26.5 なし 合流部③ H19 0.12~0.29 6.8~7.9 302~304 3.38~5.38 8.7~27.9 なし ビオトープ池 H19 0.15~0.83 7.0~7.9 190~277 4.25~6.86 12.0~28.0 池岸
45
水路1(平成 18 年度) 水路1(平成 19 年度)
水路2(平成 18 年度) 水路2(平成 19 年度)
水路3(平成 18 年度) 水路3(平成 19 年度)
46 4.3.2 魚類の生息状況 各調査地点における魚類の調査結果を表6に示す.表中の捕獲数とは,平成18年度は8回,平 成19年度は9回の調査における総捕獲数である.なお,ビオトープ池及び落口工①~③は,平成 18年度の水稲収穫後に改修が行われたため,平成19年度から調査を開始した.また,調査結果か ら(1)式で表わされるSimpsonの多様度指数D (宮下・野田,2003)を表6に示した.
S i iP
D
1 21
(1) ここに,S は群集に含まれる種の数,Piは種 i の個体数が群集の全個体数に占める割合である. 各水路等における魚類の多様性について Simpson の多様度指数からのその大まかな傾向を検 討する.まず,用排兼用水路と連続し灌漑期の水深が大きい未整備の水路 3 では,平成 18 年度 および平成 19 年度ともに 0.56 以上の大きな値を示した.これに対して,汎用農地として圃場整備 が完了した整備済みの水路 1 と 2 については,いずれも水路 3 より低い値を示した.水路 1 は種数 と捕獲数が減少し,多様度指数も 0.46 から 0.19 にまで低下している.水路 2 については,種数や 捕獲数は減少しているが,多様度指数は平成 19 年度の方が高い値を示している.これは,ミナミメ ダカの捕獲数が大幅に減少したために,多様度指数の算定時にミナミメダカの捕獲数による影響 が小さくなり多様度指数が大きくなったといえる.落口工①~③は水面積 1m2と生息空間が限定さ れていることもあり,水路 1~3 の多様度指数に比べてかなり低い数値を示しているが,水路 3 の下 流端に位置する落口工③の多様度指数は,水路 1 および 2 の下流端にある落口工①と②の値に 比べて大きく,合流する排水路の影響を強く受けている.次に,この地域で捕獲された各魚種の傾 向について述べる.水路 1 及び水路 2 では,平成 18 年度から平成 19 年度にかけてカワバタモロ コが増加したが,その他の多くの魚種の捕獲数は著しく減少している.とくにミナミメダカやギンブ ナでその傾向が顕著である.一方,水路 3 ではドジョウの捕獲数が減少していたが,その他の魚種 は,増減の変化が少ないか,あるいは増加していることから,水路 3 における魚類の生息環境は安 定していたと考えられる.47 表 6: 採捕結果 種名 ビオトー プ池 水路 落口工 水路 1 水路 2 水路 3 落口工① 落口工② 落口工③ H19 H18 H19 H18 H19 H18 H19 H19 H19 H19 捕 獲 数 ギンブナ 12 315 1 255 9 182 220 1 1 3 ゲンゴロウブナ 0 0 0 0 0 0 1 0 0 0 オイカワ 0 0 0 12 0 26 2 0 0 0 カワバタモロコ 742 2 20 32 49 443 415 0 6 15 ワタカ 0 5 0 0 0 0 44 0 3 1 タモロコ 0 1 1 0 0 1 0 0 0 0 デメモロコ 0 2 0 3 0 0 11 0 0 0 モツゴ 2 0 0 0 0 1 0 0 0 0 コイ 0 3 0 13 2 2 5 0 0 0 タイリクバラタナゴ 2 1 0 0 2 25 22 0 0 0 ドジョウ 0 2 0 48 1 63 45 10 8 8 ナマズ 0 0 0 0 0 0 1 1 0 0 ミナミメダカ 201 629 185 2,226 100 1,204 964 761 268 281 カムルチー 0 0 0 0 0 1 0 0 0 0 ヨシノボリ類 0 0 0 0 0 2 30 19 3 16 総個体数 969 960 207 2,589 163 1,950 1,760 792 289 324 種数 5 9 4 7 6 11 12 5 6 6 多様度指数 0.37 0.46 0.19 0.25 0.53 0.56 0.63 0.08 0.14 0.24
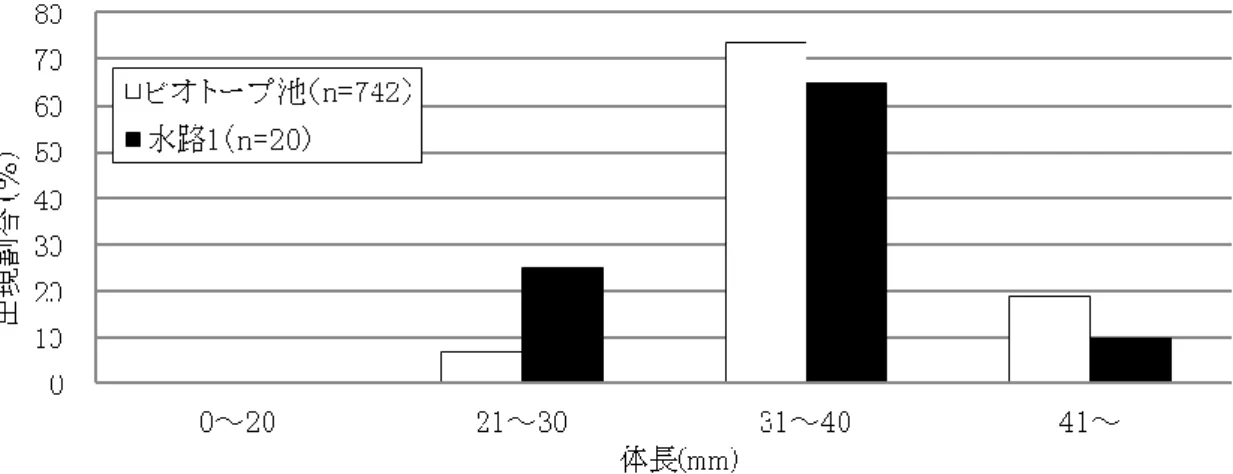
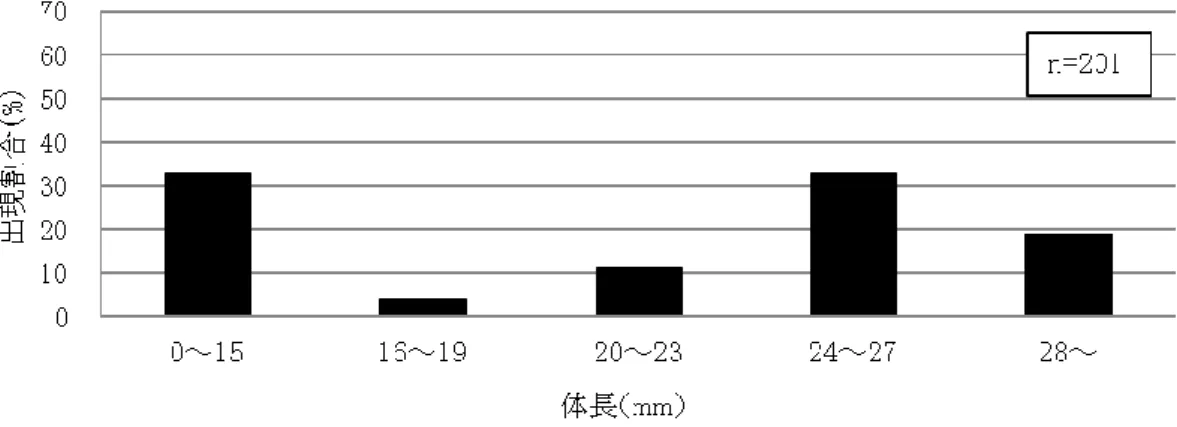
48 (1) カワバタモロコの生息状況 平成 18 年度の冬期に水路1の上流端に新設されたビオトープ池(写真 9)は,捕獲された種数は 少ないものの,ミナミメダカとカワバタモロコが多数生息していた.とくに,平成 19 年度に捕獲したカ ワバタモロコの総数は 742 個体と極めて多く,また,図 17 の体長分布図に示すように, 3 個体(総 捕獲数の 0.4%)の仔稚魚(体長 20mm 以下を仔稚魚とする)を確認し,ビオトープ池内で繁殖して いる可能性が示唆された.ビオトープ池と連続している水路 1 におけるカワバタモロコの捕獲数は, 平成 18 年度の 2 個体から平成 19 年度の 20 個体に増加したものの,図 17 に示すように,仔稚魚 が全く確認されなかった.これより,ビオトープ池で繁殖した個体が流下したことによって水路 1 内 の成魚の捕獲数は増加したが,仔稚魚は捕獲されなかったことから,水路 1 内で繁殖できなかった と考えられる.また,水路 2 では,図 18 に示すように,平成 18 年度に捕獲したカワバタモロコは 32 個体でそのうち仔稚魚 3 個体(9.4%)を確認し,水路 2 内で繁殖した可能性が示唆された.しかし, 排水路の水深が低下した平成 19 年度に捕獲数が 49 個体と若干増加したものの仔稚魚が全く確 認されず,繁殖できない環境に変化したと考えられる.一方,常時水際に植生が存在した水路 3 で はカワバタモロコの捕獲数が多い.図 19 に示すように,平成 18 年度は総捕獲数 443 個体のうち仔 稚魚が 133 個体(29.8%),平成 19 年度は総捕獲数 415 個体のうち仔稚魚が 54 個体(12.8%)で, 両年度とも水路内での繁殖を確認した.すなわち,ビオトープ池および水路 3 がこれに連結する水 路系のカワバタモロコの保全に大きな役割を果たしていることが示された.コンクリート 3 面張りの水 路においてもカワバタモロコが生息している例があるが(金井ら,1998),水路 3 と同様に水路底に 泥が堆積し,水生植物が繁茂している環境である.このことから,水路内の植生がカワバタモロコの 生息に必要な環境要素であることがわかる.
49
50
図 17 ビオトープ池と水路 1 におけるカワバタモロコの体長分布(平成 19 年度)
図 18 水路 2 におけるカワバタモロコの体長分布
51 (2) ミナミメダカの生息状況 ビオトープ池におけるミナミメダカの体長分布(図 20)を見ると,体長 20mm(孵化後 3 カ月程度) 未満の仔稚魚は総捕獲数の約 37%を占め,ビオトープ池内で繁殖していることが示された.水路 2 における平成 18 年度から 19 年度にかけてのミナミメダカの体長分布の変化(図 22)より,平成 18 年度は体長 20mm 未満の仔稚魚が総個体数の 38%を占め繁殖が確認できたが,平成 19 年度に は体長分布のピークが右側に移行したものの 20mm 未満の仔稚魚がほとんど確認できなかった. すなわち,平成 19 年度のミナミメダカは,平成 18 年度に孵化した個体が成長したもので,平成 19 年度は繁殖しなかったと考えられる.これは,平成 18 年度は整備直後のため田面からの漏水を 危惧して高い水位で管理されたことに対して,平成 19 年度は暗渠からの排水が阻害されない 水位で管理されたことにより水位が下がり,水路内の植生として機能していた畦畔植生まで水面が 上昇せず,繁殖環境がなくなったことが仔稚魚をほとんど確認できない原因と考えられる.一方, 水路 1 は,水路 2 と同様平成 19 年度に麦作が導入され排水路水位が低下し,ミナミメダカの繁殖 環境が消失されたことが予想された.しかし,図 21 の体長分布図から平成 19 年度も体長 20mm 未 満の仔稚魚が相当数確認された.これは,水路 1 の上流のビオトープ池で繁殖した仔稚魚が流下 したことによるものと思われる.なお,図 23 からも分かるように,平成 18 年度と 19 年度とも灌漑期の 排水路水位が高く管理された水路 3 では,両年度の体長分布に大きなお傾向変化はなく,この水 路内でミナミメダカの繁殖と生育が安定的に繰り返されていることが示されたといえる.
52
図 20 ビオトープ池におけるミナミメダカの体長分布(平成 19 年度)
図 21 水路 1 におけるミナミメダカの体長分布
53
54 (3) ギンブナの生息状況 平成 18 年 5 月には,日雨量 100mm を超える降雨が観測されている.この降雨時に水路 1 及び 2 において,ギンブナが遡上するのを目視で確認している.その後,多数のギンブナの仔稚魚が捕 獲されるようになった.図 24,25 から中干しまでの灌漑前期は仔稚魚が多く,中干し後は成魚が多く なる傾向がわかる.しかし,平成 19 年度には,遡上個体も繁殖個体も確認することはできなかっ た.この原因として,通常の降雨時には排水路水位が上昇しても水路内の落差が解消されず遡上 できなかったこと,水路内に産卵床となる植生がなかったことなどが考えられる. 図 24 水路 1 におけるギンブナの体長分布(平成 18 年度,n=315) 図 25 水路 2 におけるギンブナの体長分布(平成 18 年度,n=255)
55 水路1と水路2 における魚類の調査結果から,平成 18 年度に繁殖した仔稚魚は,降雨時に 落口工より下流へ流下し,水路内で越冬しなかったと推定される.一方,水路3 の捕獲数はほとん ど変化しておらず,ギンブナは水路内で定着していた.これより,水路内で繁殖や生息できる環境 が必要となるだけではなく,水路内の落差部に水路魚道を設けること,さらに,水路の一部に遊泳 力の乏しい魚種や仔稚魚が生息できる区間を設け,そこが越冬地として機能するよう水深の確保 に配慮できれば,ギンブナの繁殖も可能になると考えられる.これは,類似した生活史を有するオ イカワやコイについても同様のことが言える. 以上のことから,水路内の魚類が減少した要因は,水深が低下したことによって畦畔植生が生 息場所として利用できなくなったこと,下流部に遡上が困難な落差の発生,および定着・越冬の拠 点となるような場所の欠如にあるといえる.とくに水路内の植生は,産卵床となるだけでなく,植生 の内部は流速が低下し遊泳能力の低い仔稚魚の隠れ場となるため,上記の問題点の解決には非 常に重要である.また,植生に付着した藻類などが餌となるなど多面的な役割を果たしている.
56
5 ミティゲーションの検討
以上の結果から,水域の確保や植生の重要性を明らかにすることができた。水田地域での水管 理は農家に任せられており,地域によって異なってくるため,具体的な数値目標を掲げることは難 しいが,魚道の設計では体高の 2 倍程度の水深が必要とされており,最浅水深がその値となるよう に設計しておく必要がある. 魚類の生育ステージにより生息場所は異なるが(中村,2007),植生帯はどの生育ステージでも 隠れ場ともなり,餌場や産卵場を提供する重要な環境要素であることが裏付けられた.汎用化水田 での調査を行った地域でも水路 1 と水路 2 にも植生帯を設けることが望ましいと判断し,平成 19 年4 月に U 字工をトンネル状になるよう水路内に伏せて置き,その上にアヤメなどを植栽したプラ ンターを設置した.水路底面からプランター上部まで約 0.4m の高低差がある.この対策は,水路 内に隠れ場(U 字溝下)や植生(プランター内)を提供するために工夫したものである.しかし,環 境調査の結果から,麦作が導入された平成19 年度の場合,排水路の水深が 0.3m 以下になるよう に管理されたことから,プランターまで水面が達せずプランター内の植生が魚類の繁殖や生息環 境としての役割を果たさなかった.また,このプランターを 10~20m 程度の間隔で配置したが,ど の程度の間隔が適当であるかの基準も明らかでなかった.植生が連続していることが望ましいが, 植生帯は通水阻害の一因でもあり,農家から嫌煙される可能性がある. そこで,水路内に植生が繁茂し,魚類の生息に適した環境であると思われる水路 3 を含む水域 において,調査地点の魚類の捕獲数と,上下流に関係なく最も近い植生までの距離との関係を明 らかにすることによって,植生を配置する際の間隔について検討した(図 26).魚類の捕獲数は平 成 19 年度のものを,植生の位置は水路 3 とそれに接続している用排兼用水路のものを使用した. 調査対象としたのは汎用化水路でも多く確認されたミナミメダカ,カワバタモロコ,ギンブナの 3 種 である. 写真8 からもわかるように,水路 3 とそれに接続した用排兼用水路の水路底には砂泥が 厚く堆積し,水路内の各所に植生が分布していた.水路内に繁茂している植生は,ギョウ ギシバやヨシ・メヒシバといったイネ科の植物やイヌタデやミゾソバといったタデ科の植 物,ヒメクグやタマガヤツリといったカヤツリグサ科の植物が多く確認された.57
魚類の生息は植生以外の要因によっても影響されるので,個体数は植生からの距離にしたがっ
て単純に減少するような関係は見られなかった.しかし,図 27 からわかるように 3 種の魚類とも植生 からの距離が 40m以内に 50%以上が生息し,100m を超えると捕獲数が減少する傾向がみられ る.
58
59
60
6 まとめ
本研究では,環境配慮型水路として整備された排水路において魚類の移動・定着の視点 から,排水路をどのように整備していくべきかについて検討した.湿地と異なり,護岸勾 配の大きい農業用排水路は,水位の上下にともなう環境の変化が大きいことに留意する必 要がある.農業用排水路では,水位変化を予測した施設設計が必須であるが,農家の水管 理は多様である.そのため,水路内の環境を目的通りに維持するためには,水位変化に対 して,余裕を持った計画を行うことが必要である. 魚類の採捕結果から,ミナミメダカやカワバタモロコなどの遊泳魚は,整備された排水 路においても成魚・仔稚魚ともに確認できたが,ドジョウやツチフキなどの底生魚や,泥 の中にもぐっていたウナギはほとんど確認できず,平成19 年度には 1 個体も採捕できなか った.水路が整備されてから湿地帯を埋め立てるまでの期間を 2 年間設けたこともあり, 本調査地では人為的な魚類の移動は行わなかった.しかし,底生魚は新たな環境が整備さ れて 2 年が経過しても移動していないことになる.水田魚道へのドジョウの遡上行動の誘 発要因として水流の発生が必要であることがわかっているが(鈴木ら,2001;楠田ら,2009), 整備された環境配慮型水路は流れがほとんどなく,これがドジョウの遡上に影響していた ことも考えられる.整備を行う地域と代替地が離れている場合には人為的な魚類の移植が 行われている.生物の移植には,餌や底質,地形など魚類の生息に関して考慮すべき環境 条件は多いが,本調査の結果から,湿地帯を埋め立てるためには,消失地と代替地が連続 している地形条件であっても,とくに底生魚に関しては人為的な移動を行う必要があるこ とが示唆された. 調査結果から,水際や水中の植生が魚類の生息に重要で不可欠な環境要素であることが 改めて確認できた.湿地帯ではヨシやマコモ,それにガマといった抽水植物以外にも,水 中の藻類や浮葉植物など多様な植生が存在していた.環境配慮型水路内においても,マコ モや緑藻類,花いかだに多くの魚が定着していたが,水位変化に対しては湿地帯の植生と 比較して不安定であった.これらの結果から,水際植生の多様性を確保し,さらに水面を 被覆する植生帯を設ける必要性が示唆された.61 今後の課題として,植生の管理やゴミの除去といった維持管理体制を確立すること,定 期的なモニタリングを長期にわたり継続することで生物相の変化,とくに外来種の侵入状 況を把握していくことが重要である.また,面的な広がりを持つ湿地帯に対し,直線的な 排水路では水際に生息するカヤネズミやダルマガエルは全く確認されなかった.そのため, 魚類以外の生物についての調査を実施し,それらの結果を総合して水路の効果を判定すべ きである. 整備された汎用化水田に麦などの畑作物が栽培されることによって排水路の水深が低く管理さ れることが明らかになった.このことによって,水路内の植生環境が大きく変わり魚類の生息に大き な影響を与えていることがわかった.よって,水深が変化しても対応できるような環境を水路内に創 出する必要がある. 植生帯の有無が魚類の生息に大きな影響を与えており,この植生配置の際の基準となる適正な 間隔を示すことができた.今後,他の環境下でも多種の魚種について同様の検討を行うことが望ま れる.また,植生帯を設ける際に移植する植物種や工法についても検討することが必要であろう. 調査の結果,魚類の移動障害となっている落口工の落差も明らかになった.その落差の 直下流の深みは中干しなど急な落水があった際の魚類の避難場所の役割を果たしている可 能性が示唆された.この落差に水路魚道を設ける場合には,深みをなくすのではなく,深 みを有効に活用できる魚道の設置が望ましいと考えられた. 本研究を行った地域では,周辺住民の環境への関心が高く,堰板の設置など住民の理解が大き い.地域の環境を保全していくためには,地域住民の理解と協力が不可欠である.今後は,地域 生態系の保全方法だけでなく地域住民の協力が得られるような手法を模索していかなければなら ない.
62
7 今後の展望
本研究では,水域の変化に生息を大きく影響される魚類相の変化から,圃場整備を行う に当たっての留意点を明らかにしてきた. 地域内に魚類が退避できる場所を設けることが望ましいが,調査結果から落水に伴って 流下する個体が多く見られたこと,上流部にあるビオトープ池から流下した個体が水路内 で生息していたことから,保全しようとする地域内において中流部~下流部に設けること が望ましい. そのような環境は一時的に孤立しても,灌漑期に地域の水域ネットワークと連続するこ とができれば,その地点から拡散し地域全体で繁殖することが可能になると考えられる. しかし,孤立している間に水域が消失する,水温が生息可能な範囲を超える等にならない ように対策を講じておく必要がある. 水田環境に生息するのは魚類だけではない.消失した湿地帯で確認されていた小動物や 両生類の中にも整備後は生息が確認できない種もある. 圃場整備事業を実施する地域に生息するすべての生物を保全することが望ましいが,種 によって保全方法は異なってくるため,その地域で生息する生物を把握しどの種に重点を 置くかを事前に検討しておかなければならない. 環境は常に一定ではない.その年の気候や作付状況により異なってくるものである.水 田生態系は,人為的な環境であり,人為的な撹乱を必要とする.このため,整備した環境 配慮型水路も維持管理は欠かせない.例えば,花いかだは定期的な植物の植え替え,いか だ部分の修繕が必要である.また,水路に対してどの大きさの花いかだが適しているのか, どの植物が適しているのか等の具体的な指標は確立できていない.階段護岸では計画水深 を実際の水深が下回り,水深の変化に対応した護岸の植生が生かせていなかった.整備後 も実際の作付けに伴う水深変動に合わせて,手を加えていくことが必要となってくる. 本調査地域では植生帯から40m以内に 50%を超える魚類が生息していることが確認でき, この地域での目安となる値を明らかにすることができた.しかし,魚種によってこの値は 異なってくると考えられるため,今後他地域や他種での調査検討が必要であろう.63 圃場整備事業を実施するにあたり,環境に配慮することが原則となってから10 年以上が 経過しており,様々な工法で施工されている.しかし,本研究でも明らかになったように, 整備後に現場の状況に応じて変更しなければならないことも多い. 本調査地域では,環境配慮型水路を整備するにあたり近隣小学校でどのようなビオトー プにしたいのか模型作りを行う,地域の子ども達と実際に生物を捕獲し,その結果を考え, その地域にまつわる民話を聞く,地域の農家の方の意見を聞くなどの機会が設けられた. このような地域住民との交流は,維持管理も含め保全活動を行っていく上で非常に重要な ことであり,積極的に実施していく必要があるといえる.
64
謝辞
本学位論文の作成にあたり多大なご尽力とご指導を賜りました水利環境研究室の千家正 照教授,伊藤健吾准教授,静岡大学の土屋智教授,ならびに調査に協力してくださいまし た同研究室の学生に深く感謝申し上げます. データを提供していただき,時には貴重な経験となる機会を与えてくださいました東海 農政局計画部資源課の皆様,調査に同行させていただきました(財)日本生態系協会の皆様, 様々な資料を提供してくださいました輪之内町役場産業振興課の皆様,調査を温かく見守 り時にはお話を聞かせていただきました輪之内町の皆様に心より感謝申し上げます. 平成23 年 9 月 森 須美子65
参考文献
小澤祥司(1999):生きもの豊かな水田を再び,水情報,19(4),pp.8-11. 片野 修 (2002): 魚類保全の視点からみた水田生態系,琵琶湖研究所所報 第19号,滋賀県 琵琶湖研究所,pp.72-76. 金井直幸・板井隆彦 (1998):第2章 希少淡水魚:保護の実例,森 誠一,魚から見た水環境-復 元生態学に向けて/河川編-,第2版,pp.61-79,(株)信山社サイテック 上月康則・佐藤陽一・村上仁士・西岡健太郎・倉田健悟・佐藤家康・福田守(2000):都市近郊用 水路網におけるメダカの生息環境要因に関する研究,環境システム研究,28,pp.313-320. 楠田 聡・笠原 昇(2009) :水田魚道を用いたドジョウの遡上,魚と水,45-3,pp.19-22. 斉藤賢治・片野 修・小泉顕雄 (1988): 淡水魚の水田周辺における一時的水域への侵入と産卵, 日本生態学会誌,38,pp.35-47. 齋藤はるみ・印藤久喜・石島光男(2010): 食糧自給力強化の観点から見た農地整備の効果,水 土の地,78(12),pp.1011-1014 佐藤太郎・東 淳樹(2004): 農業用小河川における生態系に配慮した排水路改修が魚類相と生 息環境に及ぼす影響,野生生物保護,9,pp.63-76. 鈴木正貴・水谷正一・後藤 章(2001):水田水域における淡水魚の双方向移動を保証する小規模 魚道の試作と実験,応用生態工学,4(2),pp.163-177. 中川昭一郎 (2000): ほ場整備における生態系への配慮「農村と環境 第16号」(農村環境整備 センター編),農村環境整備センター,pp.48-53. 中村智幸(2007):水路と水田の生態系,水谷正一,第1版,pp.40-50,社団法人 農村漁村文化 協会,東京都 堀野治彦・中桐貴生・荻野芳彦 (2008): 環境配慮型用水路の魚介類生息および通水機能への 影響評価,農業農村工学会論文集,254,pp.161-167. 丸山幸太郎・片野知二・内堀伸勇・大橋清・片野雅夫 (1996): 開けゆく輪之内,第1版,江戸時 代の輪之内,pp.111-114,輪之内町企画課66 宮下 直・野田隆史(2003):「種多様性」『群集生態学』,東京大学出版会,pp.73-105. 森 淳 (2008): 水田生態系の特徴と変質-水田生態工学の視点から-,農業農村工学会論文 集,254,pp.127-137. 守山 弘 (1997): 水田を守るとはどういうことか-生物相の視点から-,農山漁村文化協会. 遊磨正秀・田中哲夫・竹門康弘・中井克樹・渕側祐一・小原明人・今泉眞知子・佐藤 浩・土井田 幸郎 (1997): 瀬田月輪大池における魚類群集の変遷-12年間の生物学実習の結果より-,滋 賀医科大学基礎学研究,8,滋賀医科大学,pp.19-36. 若杉晃介・藤森新作 (2009): 水田の高度利用を可能とする地下水位システムFOEAS,水土の知, 77(9),pp.705-708
67
67 表 道下地区月別捕獲個体数 平成18 年度 ギンブナ カワバタ モロコ ミナミ メダカ モツゴ ワタカ コイ モロコ類 タイリク バラタナゴ ブルーギル その他 計 地点1 4月 3 24 17 4 18 66 5月 2 54 6 3 6 11 82 6月 17 6 23 7月 43 2 7 8 14 2 76 8月 9月 10 18 4 14 2 1 49 10月 4 4 13 6 4 31 11月 1 1 45 1 1 49 12月 8 1 9 地点2 4月 1 1 5月 5 5 6月 13 1 1 1 16 7月 15 2 22 8 3 50 8月 9月 6 13 1 2 3 25 10月 2 5 2 3 2 14 11月 1 40 5 1 42 3 1 1 94 12月 6 1 2 1 10 地点3 4月 2 25 1 28 5月 1 2 3 6月 3 2 5 7月 48 18 10 23 5 1 105 8月 9月 1 13 9 1 24 10月 2 68 1 7 2 9 9 1 99 11月 8 3 20 10 28 69 12月 8 2 9 1 28 48 地点4 4月 18 2 20 5月 2 15 2 1 20 6月 5 3 2 2 3 15 7月 24 2 1 20 10 1 2 60 8月 9月 1 1 9 1 7 19 10月 1 3 6 2 3 1 1 17 11月 1 3 5 5 2 9 1 26 12月 44 56 5 2 12 11 130 地点5 4月 5月 6月 4 3 7 7月 7 2 11 15 35 8月 9月 3 16 3 22 10月 2 8 7 1 18 11月 1 1 1 2 5 12月 3 1 1 7 2 14
68 表 道下地区月別捕獲個体数 平成19 年度 ギンブナ カワバタ モロコ ミナミ メダカ モツゴ ワタカ コイ モロコ類 タイリク バラタナゴ ブルーギル その他 計 地点1 4月 5月 6 2 8 6月 7月 8月 2 3 5 3 1 14 9月 10月 6 11 4 3 4 28 11月 12月 1 1 地点2 4月 1 1 1 3 5月 5 2 3 1 11 6月 7月 8月 3 3 1 7 9月 10月 1 1 7 5 2 16 11月 12月 12 1 13 地点3 4月 5 9 1 1 5 21 5月 4 2 2 50 1 59 6月 7月 8月 11 11 9月 10月 1 1 2 6 5 15 11月 12月 1 83 13 6 103 地点4 4月 3 33 1 4 17 1 5 1 65 5月 4 1 3 2 10 6月 7月 8月 4 1 5 9月 10月 1 2 22 5 32 2 1 1 66 11月 12月 2 20 29 3 54 地点5 4月 6 2 1 4 13 5月 1 1 6月 7月 8月 7 7 9月 10月 11月 1 5 11 2 19 12月 1 6 1 8
69 表 体長別捕獲個体数 平成18 年度 水路 1(18) 5月 6月 7月 ギンブナ コイ ギンブナ オイカワ ミナミメダカ ギンブナ タモロコ ドジョウ ミナミメダカ 7 8 9 10 1 11 5 12 12 13 12 3 3 14 11 2 7 15 9 8 2 16 6 8 4 17 9 12 7 18 7 2 14 6 5 19 5 7 4 2 20 1 2 11 1 3 21 2 7 1 22 9 2 3 23 12 3 4 24 4 2 25 8 1 26 12 1 27 11 1 1 1 28 11 1 29 8 2 30 4 1 2 1 31 7 4 32 14 1 1 33 7 1 34 9 4 35 6 4 1 36 3 6 37 5 7 38 2 2 39 2 40 3 3 1 41 3 3 1 42 1 5 43 4 1 44 45 46 47 1 48 2 49 50 体長 (mm)
70 表 体長別捕獲個体数 平成18 年度 水路 1(2/6) 5月 6月 7月 ギンブナ コイ ギンブナ オイカワ ミナミメダカ ギンブナ タモロコ ドジョウ ミナミメダカ 51 52 53 54 55 1 56 1 57 58 59 60 61 62 63 64 65 66 計 77 2 149 1 79 64 1 2 47 体長 (mm)
71 表 体長別捕獲個体数 平成18 年度 水路 1(3/6) 9月 ギンブナ タイリクバラタナゴ ワタカ カワバタモロコ デメモロコ ミナミメダカ 7 8 9 8 10 10 11 10 12 6 13 2 14 4 15 1 16 1 17 1 18 19 20 2 21 1 1 22 1 2 23 1 24 1 25 1 1 26 27 1 1 28 29 30 1 31 32 33 34 35 36 37 38 39 40 1 41 42 43 44 45 46 47 1 48 49 50 体長 (mm)
72 表 体長別捕獲個体数 平成18 年度 水路 1(4/6) 9月 ギンブナ タイリクバラタナゴ ワタカ カワバタモロコ デメモロコ ミナミメダカ 51 1 52 53 54 1 55 1 56 1 57 3 58 1 59 60 61 1 62 63 64 65 66 計 9 1 2 2 2 52 体長 (mm)
73 表 体長別捕獲個体数 平成18 年度 水路 1(5/6) 10月 11月 12月 ギンブナ ミナミメダカ オイカワ ミナミメダカ ギンブナ ミナミメダカ 7 2 8 5 9 2 10 7 11 6 1 12 5 1 4 13 9 2 2 14 16 3 5 15 12 1 5 16 24 4 10 17 25 7 13 18 22 1 9 22 19 22 7 13 20 18 12 16 21 4 1 7 8 22 7 5 11 23 1 2 3 24 1 2 2 25 1 2 3 26 2 27 1 28 1 29 1 30 1 1 31 32 1 33 34 1 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 1 体長 (mm)