東京大学医学部附属病院薬剤部(〒1138655 東京都文 京区本郷 731)
e-mail: yoshihide-tky@umin.ac.jp
本総説は,2019 年度日本薬学会奨励賞の受賞を記念し て記述したものである.
2019 The Pharmaceutical Society of Japan ―Review for award―
脂溶性物質の体内動態制御機構の解明とその理解に基づくトランスレーショナルリサーチ
山 梨 義 英
Translational Research Based on Understanding the Regulatory
Mechanisms of in Vivo Behaviors of Fat-soluble Compounds
Yoshihide Yamanashi
Department of Pharmacy, The University of Tokyo Hospital; 731 Hongo, Bunkyo-ku, Tokyo 1138655, Japan. (Received May 26, 2019)
Several fat-soluble compounds such as cholesterol and fat-soluble vitamins have important physiological activities in the body, and their excess and/or deˆciency have been reported to be closely associated with the onset and progression of several conditions such as lifestyle-related diseases. It is important to clarify not only the physiological activities but alsoin vivo kinetics of fat-soluble compounds to understand their in vivo activity (toxicity). This review introduces our recent(reverse) translational research in a combination of basic and clinical studies to reveal the regulatory mechanisms ofin vivo behaviors of fat-soluble compounds and eŠects of their disruption in humans.
Key words―transporter; cholesterol; fat-soluble vitamin; low-density lipoprotein; Niemann-Pick C1-like 1; ATP-binding cassette transporter G5 and G8
1. はじめに 戦後,日本の食生活は欧米化の流れにより大きく 変容し,日本人一人当たりの脂質(脂溶性物質)の 摂取量は大きく増加するとともに,摂取する脂質の 質(種類)も変化した.この変化は日本人の栄養状 態の改善に貢献した一方で,脂質異常症や肥満,糖 尿病といった生活習慣病の罹患率の増加につながっ たとされている.したがって,われわれが摂取する 脂質の量及び質の変化が,生体にどのような影響を 及ぼすのか明らかにすることは,生活習慣病の発症 (進行)メカニズムを理解するうえで,また,生活 習慣病の予防や治療を行ううえで極めて重要である. 脂質が生体内でどのような働きを担っているのか を明らかにするためには,各脂質の活性を明らかに するとともに,摂取した脂質がどのように体内を移 動し,各組織にどの程度分布する(取り込まれる) のかを明らかにする必要がある.脂質の活性につい ては古くから研究が盛んに行われ,脂質の有する様 々な機能が明らかとなってきた.1)一方,脂質の体 内動態については未解明な点が非常に多く,その制 御機構も含めてほとんど明らかとなっていない.こ のような背景の下,筆者は,生活習慣病と密接に係 わる脂質(脂溶性物質)の体内動態制御機構に関す る研究を基礎と臨床の両面から進めている.本総説 では 代表 的 な食 事 中脂 質で あ るコ レス テ ロー ル [Fig. 1(A)]の消化管吸収と胆汁排泄の制御機構に ついて筆者の研究成果を中心に概説するともに,コ レステロールと同様の輸送分子が係わることが明ら かとなってきた脂溶性ビタミンや脂溶性薬物の体内 動態制御機構についても,これまでに得られた研究 成果を紹介したい. 2. コレステロールの消化管吸収と胆汁排泄の制 御機構 コレステロールの恒常性は生体内での合成,胆汁 酸などへの異化,胆汁中への排泄,そして消化管か らの吸収のバランスによって保たれ,各プロセスの 破綻は生活習慣病の発症・進行と密接に係わること が報告されている.24)上記プロセスの中で,消化 管吸収と胆汁排泄については,近年の遺伝子改変マ ウスを用いた解析やヒトの遺伝子変異解析から,コ レス テロ ー ル取 り 込み トラ ン スポ ータ ー であ る
山梨義英 東京大学医学部附属病院薬剤部助教. 薬学(博士).2005 年東京大学薬学部 卒業,2010 年同大学院薬学系研究科博 士後期課程修了後,2010 年 4 月より東 京大学医学部附属病院 22 世紀医療セン ター特任助教,2011 年 8 月よりニュー ヨーク大学ランゴーン医療センター博 士研究員を経て,2013 年 8 月より現職.
Fig. 1. Chemical Structures of NPC1L1-related Compounds
Fig. 2. Regulation of Intestinal Cholesterol Absorption by NPC1L1 and ABCG5/G8 Niemann-Pick C1-Like1 ( NPC1L1 )5,6)と , コ レ ス テロール排出トランスポーターである ATP-binding cassette transporter G5 と G8 のヘテロダイマー (ABCG5/G8)7,8)が重要な役割を担うことが明らか となっている(Figs. 2 and 3). 2-1. NPC1L1 によるコレステロールの消化管吸 収制御 2-1-1. NPC1L1 と エ ゼ チ ミ ブ エ ゼ チ ミ ブ [Fig. 1(B)]は消化管からのコレステロール吸収を 阻害することで脂質異常症を改善する薬物であり, 現在,日本を含む世界 90 ヵ国以上で広く臨床使用 されている.エゼチミブの標的タンパク質は長らく 明らかとなっていなかったが,2004 年に Altmann らのグループから NPC1L1 遺伝子欠損(KO)マウ スの消化管コレステロール吸収がエゼチミブを投与 した野生型(WT)マウスと同程度(約 30%)にま で低下し,NPC1L1 KO マウスではエゼチミブの効 果が認められないことが報告された.5)これらの結 果 は , NPC1L1 が エ ゼ チ ミ ブ 感 受 的 な コ レ ス テ ロールの消化管吸収において重要な役割を担うこと を示唆するものであった.その後,エゼチミブの活 性代謝物であるグルクロン酸抱合型のエゼチミブと NPC1L1との直接的な結合が示され,9)現在では, NPC1L1 がエゼチミブ(及びその代謝物)の標的 タンパク質であると考えられている. 2-1-2. NPC1L1に よ る 消 化 管 ス テ ロ ー ル 吸 収 Altmannらによる NPC1L1 KO マウスの表現型 が報告されてからしばらくの間,in vitro において NPC1L1の機能を再現することが困難だったこと もあり,NPC1L1 の詳細な機能解析は進まなかっ た.そのような背景の下,筆者はそれまで報告例の なかった培養細胞で NPC1L1 の機能を再現する in vitro 機能評価系の構築を試みた.10)まず,消化管
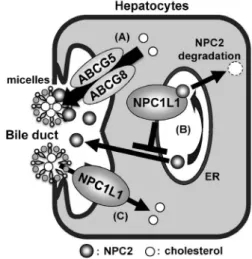
Fig. 3. Regulation of Biliary Cholesterol Excretion by
Func-tional Associations among NPC2, ABCG5/G8, and
NPC1L12,16)
(A) ABCG5/G8-mediated cholesterol eŒux from hepatocytes to bile is enhanced by biliary NPC2. (B) NPC1L1 interacts with NPC2 at the en-doplasmic reticulum (ER) and down-regulates NPC2 expression and secre-tion by accelerating NPC2 degradasecre-tion. (C) In addisecre-tion to its well-known function as a cholesterol importer at the bile canalicular membrane, NPC1L1 indirectly suppresses ABCG5/G8-mediated cholesterol eŒux by negatively regulating biliary NPC2 secretion.
上 皮 が ん 由 来 の 培 養 細 胞 で あ る Caco-2 細 胞 に NPC1L1 cDNA を安定導入した NPC1L1 高発現細 胞を構築 した.構 築した NPC1L1 高 発現細胞 で は,生理的局在と一致して,NPC1L1 は頂端側膜 (アピカル側膜)に高発現していた.一般に,消化 管管腔内で脂溶性の高い化合物は胆汁酸ミセルに よって可溶化されて存在している(Fig. 2).そこ で,胆汁酸(タウロコール酸)・リン脂質(ホス ファチジルコリン)・コレステロールからなる人工 胆汁酸ミセルで可溶化したコレステロールを用いて 細胞内取り込み実験を行った.その結果,NPC1L1 高発現細胞ではコントロール細胞と比較して,コレ ステロールの有意な取り込み量の上昇が確認され, この上昇はエゼチミブにより濃度依存的に阻害され た.すなわち,in vivo で観察されていたエゼチミ ブ感受的かつ NPC1L1 依存的なコレステロール取 り込みを in vitro で再現することに成功した. 続いて,コレステロールと同様に NPC1L1 KO マウスで消化管吸収が低下する b-シトステロール (食事中に含まれる主要な植物ステロールの 1 つ) [Fig. 1(A)]についても in vitro 機能評価系を用い て解析したところ,NPC1L1 依存的な細胞内取り 込み量の上昇が認められた.10)興味深いことに, b-シトステロールに対する NPC1L1 の取り込み活 性はコレステロールに対する取り込み活性の約 3 分 の 1 であり,in vivo で観 察されていた植 物ステ ロールの低吸収性が NPC1L1 の低い取り込み活性 で説明される可能性が示唆された.また,NPC1L1 による b-シトステロールの細胞内取り込みがエゼ チミブ感受的であることも示され,この結果は,シ トステロール血症(植物ステロールの体内異常蓄積 を特徴とする遺伝性疾患で,ステロール排出トラン スポーターである ABCG5/G8 が原因遺伝子として 報告されている)7)の治療におけるエゼチミブの使 用に分子的根拠を与えるものであった. 2-1-3. NPC1L1の遺伝子変異と消化管コレステ ロール吸収 近年の遺伝子解析技術の発展ととも に,ヒトにおける NPC1L1 の遺伝子多型や遺伝子 変異についても情報が集まりつつある.実際,ヒト で見い出された NPC1L1 の遺伝子変異とコレステ ロールの消化管吸収効率,コレステロールの血液中 濃度,心血管イベントの発生率との関連が報告され ている.11,12)しかし,それらの変異が NPC1L1 の発 現や機能に及ぼす影響についてはほとんど明らかと なっていなかった.そこで,コレステロールの吸収 効率が低いヒトから見つかった NPC1L1 の遺伝子 変異11)のうち,細胞外領域に集中している 6 つの変 異(A395V,D398G,G402S,T413M,R417W, G434R)に着目し,上述の in vitro 評価系を用いて 解析を行った.13)その結果,4 つの変異(D398G, T413M,R417W,G434R)はタンパク質発現量の 低下並びに,細胞膜への移行不全を引き起こすこと が見い出され,細胞膜上の発現量が野生型と比較し て顕著に低下することが明らかとなった.一方, A395V,G402S の変異体については,野生型と同 程度の細胞膜上での発現が認められたが,コレステ ロールの取り込み活性は,野生型と比べて有意に低 下していた.また,わずかながらに細胞膜に発現す る R417W,G434R 変異体についても NPC1L1 の 膜発現量あたりのコレステロール取り込み活性が低 下することが明らかとなり,NPC1L1 の細胞外に 位置する A395G434 の領域がコレステロールの輸 送活性又は基質認識に重要であることが示唆され た.これらの結果は,NPC1L1 の遺伝子変異が, コレステロール低吸収性の原因となり得ることを示 唆するものであると同時に,将来,NPC1L1 によ るコレステロール輸送機構や基質認識機構の詳細を
解析する上で重要な知見を提供するものであった. 2-2. NPC2・ABCG5/G8・NPC1L1 の相互機能 連関によるコレステロールの胆汁排泄制御 コレ ステロールの胆汁排泄は,生体内からコレステロー ルを排出する主要な経路の 1 つであり,その促進は 血液中(又は肝臓中)コレステロールの濃度低下の 一要因となり得る一方で,その破綻は,脂質異常症 や脂肪肝,コレステロール胆石など,様々な生活習 慣病の発症につながることが知られている.2)胆汁 中へのコレステロール排泄は,肝細胞の毛細胆管側 膜に発現する ABCG5/G8 を介した肝細胞から胆汁 中へのコレステロール排出14)と,同じく毛細胆管側 膜に発現する NPC1L1 による胆汁中から肝細胞へ のコレステロール再吸収3,4,6)のバランスにより制御 されていることが知られている.ただし,胆汁中に おいてコレステロールは胆汁酸ミセル中に存在して いることから,細胞膜におけるコレステロール輸送 活性には,トランスポーターとミセル間でコレステ ロール引き抜き及び受け渡しに関与する活性制御因 子が想定された.筆者は,そのような活性制御因子 の候 補 とし て, 胆 汁中 タン パ ク質 の 1 つ であ る Niemann-Pick C2(NPC2)に着目し,検討を行っ た.1518) 2-2-1. NPC2 に よ る ABCG5 / G8 の コ レ ス テ ロール排出活性の促進 NPC2は強いコレステ ロール結合能を持ち,細胞内においては,NPC1L1 の類縁分子である Niemann-Pick C1(NPC1)と協 調して,リソソームから他のオルガネラや細胞膜へ の細胞内コレステロール輸送を担うことが知られて いる.19)2006年に Klein らにより,肝臓に発現する NPC2 が 胆 汁 中 に 分 泌 さ れ る こ と が 報 告 さ れ た が,20)胆汁中における NPC2 の生理機能は不明で あった.そこで,細胞外に分泌された NPC2(分泌 型 NPC2)が NPC1L1 や ABCG5/G8 のコレステ ロール輸送活性に及ぼす影響を各トランスポーター の in vitro 機能評価系を用いて解析した.15)その結 果,分泌型 NPC2 は,NPC1L1 のコレステロール 取り込み活性にはほとんど影響を及ぼさない一方で, ABCG5/G8 のコレステロール排出活性を濃度依存 的に上昇させることが見い出され,ABCG5/G8 の 活性促進因子として機能し得ることが明らかとなっ た. そこで,生体内における胆汁中 NPC2 の生理機 能を検討するために,NPC2 に対する short hairpin RNA(shRNA)発現アデノウィルスをマウスに感 染させて肝臓特異的 NPC2 ノックダウンマウスを 作出し,その胆汁中脂質濃度を解析した.その結 果,胆汁酸やリン脂質の濃度には変化が認められな かった一方で,コレステロール濃度は胆汁中 NPC2 の減少に伴って有意に低下していた.逆に,NPC2 を高発現させるアデノウィルスを感染させたマウス では,胆汁中 NPC2 の上昇に伴い,コレステロー ルの胆汁中濃度は有意に上昇していた.NPC2 の高 発 現 に よ る コ レ ス テ ロ ー ル の 胆 汁 排 泄 亢 進 は ABCG5/G8 KO マウスでは認められなかったこと から,上述の in vitro の結果と一致して,マウス生 体内においても,胆汁中 NPC2 が ABCG5/G8 を介 したコレステロール排出を促進し,コレステロール の胆汁排泄を正に制御していると考えられた(Fig. 3).また,医学的必要性から採取されたヒトの胆汁 検体を用いて,胆汁中 NPC2 量と胆汁中コレステ ロール濃度を測定した結果,極めて強い正の相関関 係(相関係数 = 0.91)が認められたことから,マ ウスで見い出された制御機構はヒトにおいても当て はまる可能性が示唆された. 2-2-2. NPC1L1 による NPC2 の発現・分泌抑制 上述のように,分泌型 NPC2 は NPC1L1 のコレ ステロール輸送活性にほとんど影響を及ぼさなかっ たが,細胞内において NPC1L1 の類縁分子である NPC1と NPC2 の分子間共役が見い出されている ことや,19)NPC1L1 が細胞膜のみならず細胞内コン パートメントにも発現していることを踏まえて, NPC1L1と NPC2 の分子間共役について更なる検 討を行った.16)まず,NPC1L1 と NPC2 を高発現さ せた培養細胞(CHO-K1 細胞)を用いて免疫沈降 実験を行ったところ,両タンパク質の共沈が確認さ れた.また,免疫染色により両タンパク質が小胞体 において共局在する様子も観察された.さらに興味 深いことに,NPC1L1 の共発現により NPC2 のタ ンパク質発現量並びに分泌量がエゼチミブ非感受的 に著しく減少することも見い出された.この制御機 構が内因性タンパク質間でも生じ得るのかを検討す るために,両タンパク質を内因性に発現しているヒ ト肝臓がん由来の培養細胞である HepG2 細胞に NPC1L1を標的とした small interfering RNA(siRNA) を一過性に導入し,NPC2 の発現量・分泌量の変動
を解析した.その結果,NPC1L1 の発現量低下に 伴い,NPC2 のタンパク質発現量や分泌量の上昇が 観察された.この上昇は,NPC2 の mRNA 発現量 の変動を伴わないことから,NPC1L1 が転写以降 の段階で NPC2 タンパク質の発現量並びに分泌量 を負に制御していると考えられた.以上の一連の研 究により,肝臓に発現する NPC1L1 は,胆汁中か らのコレステロールの再吸収という直接的な機能に 加え,NPC2 の胆汁分泌を抑制することで間接的に ABCG5/G8 のコレステロール排出活性を抑制する という,異なる 2 つの機序によりコレステロールの 胆汁 排泄 を 負に 制御 し てい るこ と が示 唆さ れ た (Fig. 3). 3. 脂溶性ビタミンの消化管吸収制御機構 3-1. NPC1L1 によるビタミン E の消化管吸収制 御 コレステロール以外にも,様々な脂質や脂溶 性栄養素が胆汁酸ミセルに可溶化されて吸収される ことが知られている.しかしながら,その吸収機構 に関しては未解明な点が多い.一方で,脂溶性ビタ ミン の 1 つ であ り, 抗 酸化 物質 と して 知ら れ る a-トコ フェロール(ビ タミン E の主構成成 分) [Fig. 1(C)]はコレステロールと同様に消化管管腔 側膜から取り込まれたのち,細胞内でキロミクロン に組み込まれて体内に吸収されることが知られてお り,コレステロールとビタミン E の吸収経路には 共通性が見い出されている.21)そこで筆者は,a-ト コフェロールの消化管吸収に NPC1L1 が係わって いるのではないかと仮説を立てて検討することにし た.22)まず,上述 の in vitro 機能評価系を用 いて NPC1L1が a-トコフェロールの輸送活性を有して いるのか検討を行ったところ,a-トコフェロールの 細胞内取り込み量は NPC1L1 の高発現細胞で上昇 しており,その取り込み活性はエゼチミブによる阻 害を受けた.そこで次に生理的重要性の評価をすべ く,Wistar ラットを用いた a-トコフェロールの消 化管吸収実験を行った.その結果,エゼチミブの投 与により a-トコフェロールの消化管吸収量は有意 に低下することが明らかとなった.また,その後の 検討で,a-トコフェロール以外のビタミン E 同族 体[Fig. 1(C)]に関しても NPC1L1 の基質となる ことが明らかとなっている.23)これらの結果は, NPC1L1が従来考えられてきたような選択的なス テロールトランスポーターではないことを示す重要 な知見であるとともに,エゼチミブの服用により, ビタミン E 吸収量が低下する可能性を示唆する臨 床的にも興味深い知見である. 3-2. NPC1L1によるビタミン K1の消化管吸収 制御 ビタミン K は,1930 年代に g-グルタミル カ ル ボ キ シ ラ ー ゼ ( gamma-glutamyl carboxylase; GGCX)の補酵素として同定された脂溶性ビタミ ンである.その後の研究により,ビタミン K は肝 臓においてビタミン K サイクルを循環することで 再利用され,その過程にて GGCX の補酵素として ビタミン K 依存性血液凝固因子を活性化すること により,血液凝固作用を示すことが明らかになっ た.近年では,ビタミン K による骨代謝の制御や 血管壁の石灰化抑制など,その多岐に渡る生理作用 が明らかとなり注目を集めている.24)しかしなが ら,その消化管吸収機構については長い間不明で あった.天然のビタミン K には植物体で合成され るフィロキノン(ビタミン K1)[Fig. 1(D)]と細 菌などの微生物が合成するメナキノン類(ビタミン K2)があるが,われわれが食事から摂取するビタ ミン K の 90%以上はビタミン K1である.25)筆者 は,ビタミン K1の消化管吸収が NPC1L1 の生理的 基質であるコレステロールやビタミン E と同様に 胆汁依存的であること,26)ビタミン K 1の生理的な 吸収部位が NPC1L1 の発現部位と同様に小腸上部 であること27)に着目し,ビタミン K 1の消化管吸収 を NPC1L1 が担うという仮説の下検討を行った.28) NPC1L1 のin vitro 機能評価系を用いてビタミン K1の取り込み実験を行ったところ,NPC1L1 高発 現細胞でコントロール細胞と比較して,エゼチミブ 感受性のビタミン K1取り込みの上昇が認められ た.そこで,NPC1L1 が生理的にビタミン K1の消 化管 吸収 に 係わ っ てい るの か 検討 する た めに , NPC1L1 KO マウスを用いた消化管吸収実験を行っ た.その結果,NPC1L1 KO マウスではビタミン K1吸収量は WT マウスの約 30%にまで低下してい た.また,WT マウスにエゼチミブを投与するとビ タミン K1の吸収量は NPC1L1 KO マウスと同程度 にまで低下する一方で,NPC1L1 KO マウスではエ ゼチミブの効果は認められなかった.これらの結果 か ら , ビ タ ミ ン K1 の 消 化 管 吸 収 の 大 部 分 を NPC1L1が担っていること,エゼチミブが生理的 状況下においても NPC1L1 依存性のビタミン K1吸
Fig. 4. Proposed Mechanism of the Drug Interaction between Ezetimibe and Warfarin2,28)
Vitamin K1is absorbed by NPC1L1 in the intestine and delivered to the liver. In the liver, vitamin K1activates clotting factors through circulating vitamin K
cy-cle and regulates blood coagulation. Warfarin is an anticoagulant drug that inhibits the vitamin K cycy-cle. Ezetimibe, an NPC1L1 inhibitor clinically used for dys-lipidemia, inhibits intestinal absorption of vitamin K1in addition to cholesterol. When warfarin and ezetimibe are administered together, the anticoagulant eŠect of
warfarin is apparently enhanced by the reduction in hepatic vitamin K1level due to the ezetimibe-related malabsorption of vitamin K1.
収を阻害し得ることが明らかとなった. 3-3. ワ ル フ ァ リ ン 療 法 の 修 飾 因 子 と し て の NPC1L1 エゼチミブは,ビタミン K アンタゴ ニストである抗血液凝固薬のワルファリン[Fig. 1 (E)]と併用すると,ワルファリンの作用増強[血 液凝固時間の指標であるプロトロンビン時間(pro-thrombin time; PT)が延長する]をもたらすこと が報告されている.29,30)しかしながら,この機序は 不明であった.筆者は,NPC1L1 がビタミン K1吸 収の大部分を担うという上述の結果から,エゼチミ ブのビタミン K1吸収阻害による肝臓中ビタミン K1 濃度の低下が,エゼチミブとワルファリンの薬物間 相互作用を説明し得ると考え,Wistar ラットへの 薬物投与実験を行った.28)その結果,ワルファリン 単独投与群(以下,単独投与群)に比べ,エゼチミ ブとワルファリンの併用群(以下,併用群)におい てより顕著な PT 延長が観察され,また肝臓中ビタ ミン K1濃度は併用群において有意に減少してい た.ビタミン K1を経口投与することで併用群の肝 臓中ビタミン K1濃度を単独投与群と同程度まで上 昇させた際には,併用群における PT 延長は消失し たことから,エゼチミブとワルファリンの薬物間相 互作用はエゼチミブによるビタミン K1吸収阻害に 起因した肝臓中ビタミン K1濃度の低下によること が示された(Fig. 4). さらに,ケースレポートとしての報告例のみで あったエゼチミブとワルファリンの薬物間相互作用 について,その発生頻度を東京大学医学部附属病院 の電子カルテ情報を基に調査した結果,ワルファリ ンを服用していた解析対象者 42 名中 37 名におい て,エゼチミブの服用の前後で PT-INR 値(PT の 国際標準比)の延長が確認された.この結果は,両 薬物間の相互作用が特異体質的なものではなく,ほ ぼすべての患者で起こり得ることを示しており,上 述のラットを用いた薬物投与実験の結果を支持する ものであった. 最近筆者は,電子カルテ情報を用いた後方視的な 調査により,合成副腎皮質ホルモン剤であるプレド ニゾロン[Fig. 1(F)]をワルファリンと併用する と,エゼチミブと同様に,高頻度で PT-INR 値が 延長することを見い出した.31)この薬物間相互作用 の機序はいまだ明らかではないが,エゼチミブとワ ルファリンの相互作用メカニズムを踏まえて,プレ ドニゾロンが NPC1L1 の発現量や機能活性になん らかの影響を与えているのではないかと考えて in vitro で検討を行った.その結果,プレドニゾロン は NPC1L1 の転写をグルココルチコイド受容体 a 依存的に阻害し NPC1L1 の mRNA 発現量を低下さ
せること,また,ビタミン K1の輸送活性もエゼチ ミブほどではないが,抑制し得ることが明らかと なった.すなわち,プレドニゾロンとワルファリン の薬物間相互作用メカニズムとして,プレドニゾロ ンによる NPC1L1 の発現量低下や機能阻害による 消化管ビタミン K1吸収の低下が考えられた. これら一連の研究により,ワルファリン療法の修 飾因子として,これまでに知られていたワルファリ ンの代謝酵素である CYP2C9,ワルファリンの標 的分子である Vitamin K epoxide reductase complex subunit 1,ビタミン K の代謝酵素である CYP4F232) に加えて,ビタミン K1の吸収トランスポーターで ある NPC1L1 の重要性が示唆され,治療域が狭く 細かな薬用量調整が必要なワルファリン療法におい て重要な知見を提供することとなった. 4. LDL による薬物の体内動態制御 4-1. 脂質運搬体としての LDL リポタンパ ク質は血液中に存在する脂質複合体であり,主にリ ン脂質,アポタンパク質,コレステロール,トリグ リセリドによって構成されている.比重の違い(リ ポタンパク質中のトリグリセリドの比率の違い)に より,キロミクロン,very low-density lipoprotein, low-density lipoprotein ( LDL ) , high-density lipoproteinの大きく 4 つに分類される.これら 4 つのリポタンパク質のうち,LDL はコレステロー ルやトリグリセリドなどの脂質を LDL 受容体を介 して末梢組織に運搬する役割を担っている.LDL コレステロールの血液中濃度は遺伝的な要因や生活 習慣などによって大きく変動することが知られてい るが,その濃度が高いと,生活習慣病や動脈硬化症 を始めとする様々な脂質関連疾患の発症につながる とされている.このような背景から,臨床において は,生活習慣病の発症リスクを評価する代表的なバ イオ マー カ ーの 1 つ と して ,多 く の健 康診 断 で LDLコレステロール値(濃度)が検査されている. 4-2. 薬物運搬体としての LDL 上述のよう に,従来 LDL はコレステロールやトリグリセリド などの脂質を各組織に運搬する粒子とみなされてき たが,近年,ビタミン E やビタミン K などの一部 の栄養素も LDL に分布して体の各組織に運ばれる ことが明らかとなり,様々な生体内物質の運搬に LDLが係わる可能性が見い出されている.筆者 は,疾患治療のために用いられる薬物にも脂溶性の 高いものが数多く存在することに着目し,これら薬 物が服用後,LDL に分布して血液中を循環し, LDL 受容体を介して体内動態制御を受ける可能性 を考えて検討を行った.33) まず,薬物がリポタンパク質へ分布し得ることを 明らかにするため,様々な物性の薬物をマウスに経 口投与したのち,血液を採取し,サイズ排除クロマ トグラフィーを用いて各リポタンパク質分画を分離 した.その後,各分画中の薬物濃度を測定した.そ の結果,従来の報告と一致して,血液中の主たる薬 物結合タンパク質であるとされているアルブミンを 含むタンパク質分画に多数の薬物が検出された一方 で,LDL 分画に分布する薬物も多く(解析した全 42薬物のうち 10 薬物)存在した.なお,以降の記 載では,主にタンパク質分画に検出された薬物を 「アルブミン結合型薬物」,主に LDL 分画に検出さ れた薬物を「LDL 分布型薬物」と記載する. 上記の結果を踏まえ,どのような特徴を有する薬 物が LDL に分布するのか明らかにするため,薬物 をその水溶性及び生体内での代謝速度(膜透過性若 しくは未変化体の尿中排泄率より算出)により 4 つ のクラスに分類する Biopharmaceutics Drug Dispo-sition Classiˆcation System (BDDCS)34)を用いて,
解析した薬物をクラス分類した.その結果,LDL 分布型薬物の多く(10 薬物中 7 薬物)はクラス 2 に分類された.クラス 2 は水溶性が低く(脂溶性が 高く),代謝速度が速い薬物群であることから,こ のような特徴を有する薬物が比較的 LDL に分布し 易い傾向であることが明らかとなった.ただし,ク ラス 2 の薬物でも LDL への分布率が低い薬物はあ ること,また,他のクラスでも LDL への分布率が 高い薬物が存在することなどから,薬物の LDL へ の分布率を規定するファクターについては今後の更 なる検討が必要である. 4-3. LDL受容体を介した薬物の体内動態制御 LDL は LDL 受容体を介したエンドサイトーシス により組織(細胞内)に取り込まれる.そこで, LDL に分布した薬物が LDL 受容体を介して細胞内 に取り込まれるのかについて,LDL 受容体を高発 現又はノックダウンした培養細胞(Hepa1-6 細胞又 は Huh-7 細胞)を用いた in vitro 薬物輸送実験に より検討した.その結果,LDL に分布した薬物の 細胞内取り込みは LDL コレステロールと同様に,
Fig. 5. A Novel Concept in Drug Behavior: LDL-mediated Drug Transport33)
Drugs in circulation are typically categorized into unbound free form or a protein-bound form, and only the free form is thought to move into tissues. Based on our ˆndings, we propose that the LDL-associated form should be distinguished from the protein-bound form because of its ability to transfer into tissues.
LDL 受容体を高発現させると増加し,逆に LDL 受 容体をノックダウンすると減少した.このことから, LDL 分布型薬物も LDL 受容体を介して細胞内に取 り込まれることが明らかとなった.そこで,薬物の 体内動態制御における LDL 受容体の生理的重要性 を検討するために,LDL 受容体に遺伝子変異を有 するマウス(LDLR 変異マウス)を用いて薬物投 与実験を行った.なお,LDLR 変異マウスでは, LDL 受容体が正常に働かないため,血液中の LDL が組 織 (肝 臓) に 取り 込ま れ ず, 結果 と して , LDL コレステロールの血液中濃度が WT マウスと 比較して顕著に上昇していることを確認している. 薬物投与実験の結果,アルブミン結合型薬物の血液 中濃度は WT マウスと LDLR 変異マウスでほとん ど違いが認められなかった一方で,LDL 分布型薬 物の血液中濃度は,LDLR 変異マウスで顕著に高 い値を示した.また,アデノウィルスを用いた in vivo 遺伝子導入技術により,LDLR 変異マウスの 肝臓に正常な(野生型の)LDL 受容体を高発現さ せたマウス(LDLR レスキューマウス)では,血 液中に蓄積していた LDL が肝臓に取り込まれ, LDL 分布型薬物の血液中濃度が低下することが明 らかとなった.これら一連の結果から,LDL 受容 体は LDL 分布型薬物の組織移行を制御することで 薬物の血液中濃度並びに体内動態の制御に係わるこ とが明らかとなった.薬物の体内動態を予測する上 で,これまでは血液中でタンパク質に結合していな い非結合型薬物のみが組織移行可能であり,タンパ ク質結合型薬物は組織移行しないと考えられてい た.しかし,一連の研究により,これまでタンパク 質結合型薬物に分類されていた LDL 分布型薬物も LDL 受容体を介して組織移行し得ることが明らか となり,薬物動態学に新たな視点をもたらす重要な 知見が得られた(Fig. 5). 4-4. LDL コレステロール低下療法による血液 中薬物濃度の変動 現在,LDL コレステロール 値の低下を目的とした脂質異常症の治療は,食事療 法に加えて,コレステロール合成阻害薬(スタチン) の単独又はエゼチミブなど他の作用機序の薬剤を併 用す る 薬物 療 法が 基本 と なっ てい る .し かし , LDL受容体関連分子に遺伝子変異を有する家族性 高コレステロール患者では,上記の食事療法や薬物 療法だけでは LDL コレステロール値を十分に低下 させることができないために,LDL を選択的に吸 着するカラム(デキストラン硫酸セルロースカラム) を用いて,血液中から LDL を直接除去する治療法 (LDL アフェレシス)が行われる.筆者はヒトにお いても LDL が薬物の運搬を担うのか明らかにする ために,LDL アフェレシス療法を受けている患者 さんの協力を得て,LDL アフェレシスの前後で, 服用している薬物の血液中濃度が変動するか否かを 解析した.33)その結果,アルブミン結合型薬物の血
液中濃度はアフェレシス療法によりほとんど影響を 受けなかった一方で,LDL 分布型薬物の血液中濃 度は,LDL コレステロールと同様に,LDL アフェ レシス療法により大きく低下することが明らかと なった.これらの結果は,ヒトにおいても LDL が 薬物の運搬体として機能していることを示唆すると ともに,LDL コレステロール低下療法に伴う血液 中薬物濃度の変化に注意を促す知見となった. 5. おわりに 以上,筆者が進めている脂溶性物質の体内動態制 御メカニズムの解明と,その理解に基づいて実施し た臨床検体や電子カルテ情報を用いた臨床研究に関 して,現在までの成果を紹介させて頂いた.一連の 研究により,コレステロールの体内動態を制御する 分子(NPC1L1 やリポタンパク質代謝に係わる分 子)がコレステロールのみならず,他の脂溶性栄養 素や脂溶性薬物の体内動態制御にも係わることが明 らかとなってきた.現在,NPC1L1 やリポタンパ ク質代謝関連タンパク質を標的とした脂質異常症治 療薬が臨床使用されているが.35)これらの薬物はコ レステロールのみならず他の脂溶性物質の体内動態 (血液中・組織中濃度)にも影響を及ぼす可能性が あり注意が必要である.また,個々の脂質の体内レ ベル変動を考慮するだけでは説明できない生体応答 (病態の発症や進行)は非常に多い.しかし,上述 のような脂質間の連関も考慮して,包括的に脂質の 体内レベル変動を測定し,変動している脂質の活性 を統合して解析することで,複雑な生体応答をより 精度よく予測できると期待される.このような解析 には ,近 年 急速 に発 展 して いる メ タボ ロー ム 解 析36,37)やシステムズバイオロジーによる解析38)が有 用であろう.今後は,このような視点に立って,様 々な in vitro・in vivo 実験と in silico 解析を組み合 わせることで,脂溶性物質の摂取量変動が生体にも たらす影響をそのメカニズムも含めて明らかにして いきたい.また,これまでと同様に臨床的な解析も 組み合わせることで,研究成果の臨床への還元・応 用を目指したい. 謝辞 本稿で紹介した一連の研究は,東京大学 医学部附属病院で行われたものであり,終始多くの 御指導御鞭撻を賜りました鈴木洋史教授並びに高田 龍平講師に厚く御礼申し上げます.また,研究の遂 行にあたり,多大なる御協力・御指導を頂きました 正田純一先生(筑波大学),小倉正恒先生(国立循 環器病研究センター研究所)を始めとする共同研究 者の先生方,並びに,薬剤部研究室の皆様に感謝申 し上げます. 利益相反 開示すべき利益相反はない. REFERENCES
1) Stillwell W., ``An Introduction to Biological Membranes: Composition, Structure and Function,'' 2nd ed., Elsevier Science, Amster-dam, 2016, pp. 1590.
2) Yamanashi Y., Takada T., Suzuki H., Biol. Pharm. Bull, 41, 110 (2018).
3) Yamamoto H., Yamanashi Y., Takada T., Mu S., Tanaka Y., Komine T., Suzuki H., Mol. Pharmacol., 96, 4755 (2019).
4) Toyoda Y., Takada T., Umezawa M., Tomura F., Yamanashi Y., Takeda K., Suzuki H., FASEB BioAdvances, 1, 283295(2019). 5) Altmann S. W., Davis H. R. Jr., Zhu L. J.,
Yao X., Hoos L. M., TetzloŠ G., Iyer S. P., Maguire M., Golovko A., Zeng M., Wang L., Murgolo N., Graziano M. P., Science, 303, 12011204(2004).
6) Temel R. E., Tang W., Ma Y., Rudel L. L., Willingham M. C., Ioannou Y. A., Davies J. P., Nilsson L. M., Yu L., J. Clin. Invest., 117, 19681978(2007).
7) Berge K. E., Tian H., Graf G. A., Yu L., Gri-shin N. V., Schultz J., Kwiterovich P., Shan B., Barnes R., Hobbs H. H., Science, 290, 17711775 (2000).
8) Lee M. H., Lu K., Hazard S., Yu H., Shulenin S., Hidaka H., Kojima H., Allikmets R., Sakuma N., Pegoraro R., Srivastava A. K., Salen G., Dean M., Patel S. B., Nat. Genet., 27, 7983 (2001).
9) Garcia-Calvo M., Lisnock J., Bull H. G., et al., Proc. Natl. Acad. Sci. USA, 102, 8132 8137 (2005).
10) Yamanashi Y., Takada T., Suzuki H., J. Pharmacol. Exp. Ther., 320, 559564 (2007). 11) Cohen J. C., Pertsemlidis A., Fahmi S., Es-mail S., Vega G. L., Grundy S. M., Hobbs H. H., Proc. Natl. Acad. Sci. USA, 103, 1810
1815(2006).
12) Myocardial Infarction Genetics Consortium Investigators, Stitziel N. O., Won H. H., Mor-rison A. C., et al., N. Engl. J. Med., 371, 20722082(2014).
13) Yamanashi Y., Takada T., Suzuki H., Phar-macogenet. Genomics, 19, 884892 (2009). 14) Yu L., Li-Hawkins J., Hammer R. E., Berge
K. E., Horton J. D., Cohen J. C., Hobbs H. H., J. Clin. Invest., 110, 671680 (2002). 15) Yamanashi Y., Takada T., Yoshikado T.,
Shoda J., Suzuki H., Gastroenterology, 140, 16641674(2011).
16) Yamanashi Y., Takada T., Shoda J., Suzuki H., Hepatology, 55, 953964(2012).
17) Takada T., Yamanashi Y., Suzuki H.,Nippon Yakurigaku Zasshi, 139, 5660 (2012). 18) Takada T., Yakugaku Zasshi, 133, 451461
(2013).
19) Liscum L., Sturley S. L., Biochim. Biophys. Acta, 1685, 2227 (2004).
20) Klein A., Amigo L., Retamal M. J., Morales M. G., Miquel J. F., Rigotti A., Zanlungo S., Hepatology, 43, 126133 (2006).
21) Yamanashi Y., Takada T., Kurauchi R., Tanaka Y., Komine T., Suzuki H., J. Athero-scler. Thromb., 24, 347359 (2017).
22) Narushima K., Takada T., Yamanashi Y., Suzuki H., Mol. Pharmacol., 74, 4249 (2008).
23) Takada T., Suzuki H.,Mol. Nutr. Food Res., 54, 616622(2010).
24) Willems B. A. G., Vermeer C., Reuteling-sperger C. P. M., Schurgers L. J., Mol. Nutr. Food Res., 58, 16201635 (2014).
25) Booth S. L., Suttie J. W.,J. Nutr., 128, 785 788(1998).
26) Shearer M. J., Mcburney A., Barkhan P., Vitam. Horm., 32, 513542 (1974).
27) Hollander D., Rim E., Muralidhara K. S., Am. J. Physiol., 232, E69E74 (1977). 28) Takada T., Yamanashi Y., Konishi K.,
Yamamoto T., Toyoda Y., Masuo Y., Yamamoto H., Suzuki H., Sci. Transl. Med., 7, 275ra23(2015).
29) Ritchie S. R., Orr D. W., Black P. N.,Eur. J. Gastroenterol. Hepatol., 20, 572573 (2008). 30) Merck / Schering-Plough Pharmaceuticals, ``Zetia (Ezetimibe) tablets prescribing infor-mation.'': 〈http://www.accessdata.fda.gov/ drugsatfda _ docs / label / 2006 / 021445s014lbl. pdf〉, cited 20 May, 2019.
31) Ito S. M., Yamanashi Y., Takada T., Suzuki H., Circ. J., 83, 471480 (2019).
32) Glurich I., Burmester J. K., Caldwell M. D., Heart Fail. Rev., 15, 239248 (2010). 33) Yamamoto H., Takada T., Yamanashi Y.,
Ogura M., Masuo Y., Harada-Shiba M., Suzuki H., Sci. Rep., 7, 633(2017).
34) Benet L. Z., Broccatelli F., Oprea T. I.,AAPS J., 13, 519547 (2011).
35) Kinoshita M., Yokote K., Arai H., et al., Committee for Epidemiology and Clinical Management of Atherosclerosis, J. Athero-scler. Thromb., 25, 846984 (2018).
36) Takada T., Yamamoto T., Matsuo H., Tan J. K., Ooyama K., Sakiyama M., Miyata H., Yamanashi Y., Toyoda Y., Higashino T., Nakayama A., Nakashima A., Shinomiya N., Ichida K., Ooyama H., Fujimori S., Suzuki H., Sci. Rep., 8, 11147 (2018).
37) Toyoda Y., Takada T., Suzuki H.,Sci. Rep., 6, 24586 (2016).
38) Uchida S., Asai Y., Kariya Y., Tsumoto K., Hibino H., Honma M., Abe T., Nin F., Kura-ta Y., FuruKura-tani K., Suzuki H., KiKura-tano H., Inoue R., Kurachi Y., J. Physiol. Sci., 69, 433451 (2019).