緒 言 菌類は,動植物遺体の分解者として,植物の養水分吸 収を補助する共生者として,また植物を枯死させる寄生 者として,森林生態系における養分循環や植物群落の形 成・発達に大きな影響を与えている(大園 2018).大型 菌類は,こうした様々な役割を果たす中で,栄養を獲得 しきのこ(子実体)を形成して繁殖する.この大型菌類 の子実体発生は,気温や降水量などの環境条件(例えば Sato et al. 2012)に加えて,植物の群集構成(Brown et
al. 2006),枯死材等の植物遺体(Rudolf et al. 2012),施 肥(Edwards et al. 2004)など,利用可能な資源の量や質 に大きく依存している.そのため,強度の撹乱によって 植生や植物遺体の現存量・質などの環境条件が劇的に変 化した立地では,その環境に適応して子実体を発生でき る大型菌類相も大きく変化する.例えば,山火事跡地で は無被害地区では見られない大型菌類が子実体を発生さ せており,その菌類相は無被害地区と大きく異なってい た(Horikoshi et al. 1986).さらに,山火事後の時間経過 とともに変化する地表の状態(焼土,リター,木材,雑 草等による被覆割合)に応じて,大型菌類相も遷移する ことが報告されている(McMullan-Fisher et al. 2002).
2000 年に噴火した有珠山の噴出物堆積地における大型菌類の発生消長
小長谷 啓介
1)*・宮本 敏澄
2)・玉井 裕
2)・矢島 崇
3) 1)国立研究開発法人 森林研究・整備機構 森林総合研究所 きのこ・森林微生物研究領域, 〒 305 8687 城県つくば市松の里 1 2)北海道大学大学院農学研究院,〒 060 8589 北海道札幌市北区北 9 条西 9 丁目 3)元 北海道大学大学院農学研究院,〒 060 8589 北海道札幌市北区北 9 条西 9 丁目Seasonal occurrence of macrofungi on denuded areas formed by thick volcanic
depo-sition of the 2000 eruption on Mt. Usu, Hokkaido, Japan
Keisuke O
BASE1)*, Toshizumi M
IYAMOTO2), Yutaka T
AMAI2), Takashi Y
AJIMA3)1) Department of Mushroom Science and Forest Microbiology, Forestry and Forest Products Research Institute, 1 Matsunosato, Tsukuba, Ibaraki 305 8687, Japan
2) Research Faculty of Agriculture, Hokkaido University, Kita-9, Nishi-9, Kita-ku, Sapporo, Hokkaido 060 8589, Japan 3) Former: Research Faculty of Agriculture, Hokkaido University, Kita-9, Nishi-9, Kita-ku, Sapporo, Hokkaido 060 8589, Japan
(Accepted for publication: 20 August, 2020)
Seasonal occurrence of macrofungi was investigated on denuded areas formed by thick volcanic deposition of the 2000 eruption on Mt. Usu, Hokkaido, Japan, from 2004 to 2006. Fungal species richness increased from 9 in 2004 to 23 in 2006. Ectomycorrhizal fungal species were found at the first time in autumn in 2005 in the study site; 4 and 8 species were found in 2005 and 2006, respectively. The members of saprotrophic fungal species such as Psathyrellaceae and Strophariaceae found in 2004 were continuously found and their species richness were moderately increasing with the passage of the years. The numbers of macrofungal species disappeared were 2 and 1 during 2004−2005 and 2005−2006, respectively, whereas those of newly appeared were high: 9 and 8 during 2004−2005 and 2005−2006, respectively. The results indicate that mac-rofungi which utilize resources from plant residue buried underground and/or from regenerated seedlings through symbi-otic associations propagated on the denuded areas, and the pattern of succession in macrofungal flora can be characterized as aggregation by the newly recruited species.
(Japanese Journal of Mycology 61: 81−90, 2020) Key Words― Disturbance, Ectomycorrhizal fungi, Saprotrophic fungi, Seedling, Volcano
論 文
また,火入れと伐採による攪乱を受けた立地では,日射 が強く比較的乾燥した環境下でも,おそらく木材内部に 生息していた菌類が,攪乱直後から木質の基質を利用し て子実体を発生させていたという報告もある(López-Quintero et al. 2012).強度の撹乱を受けた立地に定着し 子実体を発生させる菌種は,攪乱の種類や強度によって 規定される資源の量・質や環境条件,さらにはその時間 経過に伴う変化の影響を強く受けていると考えられる. 森林における大型菌類相を劇的に変化させる攪乱の一 つとして,火山噴火が挙げられる.日本では 2017 年時点 で 111 もの活火山が存在し(気象庁 2017),火山噴火に よって私達の社会活動はもとより,広範囲の森林生態系 が深刻な被害を受けてきた.火山噴火による爆風や堆積 物(軽石,火山灰など)によって植生が消失した立地には, 倒木等の多くの植物遺体が地表または地中に存在してい る.これらの資源を利用して子実体を形成する大型菌類 については,これまでに次の 1 例が報告されているのみ である.2000 年に噴火した有珠山の噴出物堆積地で行わ れた 2004 年の調査では,草地や道端などで見られるム ジナタケ(Lacrymaria lacrymabunda),ヒトヨタケ(Co-prinopsis atramentaria),ササクレヒトヨタケ(Coprinus comatus),ハタケシメジ(Lyophyllum decastes)など,い わゆる腐生菌類が火口付近の裸地部に発生していた (Obase et al. 2005).同様に裸地部で確認されたニガク リタケ(Hypholoma fasciculare)とウラベニガサ(Pluteus cervinus)については,直下に埋没する木材と菌糸で繋 がっていたことから,これらの資源を利用して繁殖して いたと推察している(Obase et al. 2005). 一方,火山噴出物堆積地では乾燥や土砂流出などの環 境ストレスが依然継続しているのにもかかわらず,噴火 活動の終息後にいち早くヤナギ科,カバノキ科,マツ科 などの木本植物が侵入し(例えば,後藤 1937, Tsuyuza-ki 1989),これらの個体は定着後すぐに外生菌根菌と共 生関係を築いている(Obase et al. 2007,小長谷 2009). 外生菌根菌は,マツ科,ブナ科,カバノキ科,ヤナギ科 など特定の樹木の細根部分に外生菌根と呼ばれる共生体 を作る菌類の総称であり,宿主植物と相利共生の関係を 構築して成長し,子実体を発生させる(Smith and Read 2010).2000 年に噴火した有珠山の噴出物堆積地におけ る調査(Obase et al. 2005)では,2004 年の時点で裸地 部における外生菌根菌の子実体発生は確認されていな い.しかし,およそ 300 年前に噴火した富士山の火山荒 原では,発達したミネヤナギ群落から外生菌根菌の子実 体が発生している(Nara et al. 2003)ことから,有珠山 においても,稚樹が成長すれば外生菌根菌の子実体が現 れると予想される. このように,火山噴出物堆積地では様々な大型菌類が 様々な資源を利用して子実体を発生させているが,噴火 後間もない遷移初期の段階において,大型菌類相の変化 を継時的に捉えた研究例はこれまでにない.そこで本研 究では,2000 年に噴火した有珠山の噴出物堆積地にお いて,2004 年に調査した既報の大型菌類に関する子実 体発生調査を 2006 年まで継続して行い,噴火から間も ない遷移初期の立地で繁殖する大型菌類相の経時変化を 明らかにした. 材料および方法 調査地は,北海道有珠山西山火口群の N-a 火口(42° 33 17 N 140°48 08 E)周囲の裸地部とその周辺部とし た.噴火前の西山斜面の植生は,カラマツやトドマツな どの針葉樹人工林,またはカンバ類やカエデ類などの広 葉樹天然林であった.西山火口周辺では,主に水蒸気と 共に噴出した噴泥と噴石が厚さ 50−300 cm 程度堆積し たため,前生植生が完全に消失した裸地が形成された. ただし,ガリーの形成により地表が大きく削れた箇所の 一部では,噴火前から定着していたと思われるオオイタ ドリの個体などが再生していた.裸地部には主にスギナ, オオイタドリ,オオヨモギ,アキタブキ,ヤマアワなど のシダ・草本種子植物が侵入していた(Fig. S1A).植 被率と植物種数は年経過とともに増加し,植被率は 12.2%(2003 年)から 57.1%(2006 年)へ,植物種数 は 27 種(2003 年)から 63 種(2006 年)へとなり,植 生は緩やかに回復していた(小長谷 2009).木本植物は, オノエヤナギ,エゾノバッコヤナギ,ドロノキが 2004 年にヘクタール当たり約 181 本から 325 本で優占して定 着しており,樹高 1 m を超える個体も現れていた(小 長谷 2009).一方,火口からやや離れた裸地の周辺部で は,おそらく爆風と噴出物による枝折れや樹幹の損傷が 見られたものの,多くの樹木個体は噴火後も生残してい た.また,噴出物が厚さ 10−50 cm 程度堆積していた ものの,噴火前から生息していたと思われる草本植物が 生残していた.特にオオイタドリの繁茂が顕著であり, 裸地に隣接する形でうっ閉した群落を形成していた.群 落の上層部は主に,オノエヤナギ,イヌコリヤナギ,シ ラカンバ,ミズナラ,クリ,カラマツなど成木個体が疎 に分布しており,下層部はオオイタドリ,オオヨモギ, クサソテツなどの多年生草本植物とシダ植物が繁茂して いた(Fig. S1B). 2005 年と 2006 年の 5 月から 11 月にかけて,裸地部 (Denuded area; DA) お よ び 周 辺 部(Circumference area; C)をそれぞれ約 1,000 m と 600 m 踏査し,進路
から約数メートルの範囲で確認された子実体の菌種とそ の発生時期および発生場所の環境を記載した(Fig. 1). 2004 年において,裸地に隣接する大型のオオイタドリ 群落やその他の生残していた前生植物を,裸地部と周辺 部の境界と定めた.境界として定めた前生植物の下(茎 または幹から 1 m の範囲内)から発生した子実体は, 周辺部から発生したものとみなした.踏査のルートは 2004 年の調査時(Obase et al. 2005)と同じとした.調 査はできるかぎり降雨から 2,3 日後に行い,調査間隔 は 1−2 週間であった.対象とする大型菌類は地表性の みとし,明らかに地上の木材から発生していた子実体は 除外した.確認された子実体は実験室に持ち帰り,光学 顕微鏡による形態観察を行った.種同定は以下の文献を 参考にした : Imai (1938a, 1938b),Smith (1951, 1972), Corner (1968),McNabb (1968),van de Bogart (1976, 1979 ),Bigelow (1982),Smith et al. (1983),Stangle (1983, 1989),Breitenbach and Kränzlin (1984, 1986,
1991 , 1995 , 2000),Kit van Waveren (1985),Kuyper (1986),Palm and Stewart (1986),Vilgalys and Miller (1987),今関・本郷(1987, 1989),Uljé and Bas (1988, 1991),Cha et al. (1992a, 1992b),Mueller (1992),Ba-nerjee and Sundberg (1995), Pegler et al. (1995), Vesterholt (1995, 2005),Cripps (1997),Young and Wood (1997),車(1999),Reid and Eicker (1999),Ak-ers and Sundberg (2001),吉見(2002),Kränzlin (2005). 各大型菌類の消長の記録から,次年度に新たに確認され た菌種数(新出種数)と,消失した種数(消失種数)を 算出した.また,各年において,全菌種数に対する裸地 部および周辺部に共通して出現した菌種数の割合を算出 した.
Fig. 1. Routes for survey of sporocarps occurrence in a denuded area (DA) and circumference areas (C) at Mt. Usu, Hok-kaido, Japan.
結 果 確認された菌類相 2004 年 に 19 回,2005 年 に 23 回,2006 年 に 20 回 調 査を行った.その結果,裸地部および周辺部において, 計 85 種の子実体発生が確認された(Table 1).外生菌 根菌では,キツネタケ属のウラムラサキ(Laccaria ja-ponica),L. proxima,キツネタケ(L. parva),ワカフサ タケ属のワカフサタケ(Hebeloma mesophaeum),アセ タケ属のクロトマヤタケ(Inocybe lacera),コツブタケ 属の未同定種(Pisolithus sp.),イボタケ属の Thelephora caryophylleaと ラ シ ャ タ ケ 属 の 未 同 定 種(Tomentella sp.)の計 8 種が裸地部で確認された.これらは全て, 裸地に定着した 4,5 年生のヤナギ・カンバ類またはミ ズナラの稚樹の根元および周辺から発生していた(Fig. S2).周辺部では,裸地部でも確認された L. proxima, キツネタケ,ワカフサタケ,T. caryophyllea に加え,キ シメジ属のカラマツシメジ(Tricholoma psammopus), フウセンタケ属のウスフジフウセンタケ(Cortinarius alboviolaceus),ワカフサタケ属のオオワカフサタケ(H. crustuliniforme),アセタケ属のザラツキキトマヤタケ(I. dulcamara),ウスツヤトマヤタケ(I. nitidiuscula),キ ヌハダトマヤタケ(I. fastigiata),ヤマイグチ属のヤマ イグチ(Leccinum scabrum),ニセショウロ属のハマニ セショウロ(Scleroderma bovista),ヌメリイグチ属のハ ナイグチ(Suillus grevillei),シロヌメリイグチ(S. vis-cidus),ベニタケ属の未同定種(Russula sp.)など多様 な外生菌根菌が確認された.これらは噴火後生残したオ ノエヤナギ,イヌコリヤナギ,カラマツ,クリなどの成 木の樹下で発生していた. その他の腐生菌類では,裸地部において Arrhenia 属 の A. griseopallida,シメジ属のハタケシメジ,ササクレ ヒトヨタケ属のササクレヒトヨタケ,ホコリタケ属の未 同定種(Lycoperdon sp.),コガネタケ属のコガネタケ (Phaeolepiota aurea),キララタケ属のキララタケ(Co-prinellus micaceus ),ヒトヨタケ属のヒトヨタケ,Lacry-maria属のムジナタケ,フミヅキタケ属のフミヅキタケ (Agrocybe praecox),ニガクリタケ属のニガクリタケ, スギタケ属のスギタケ(Pholiota squarrosa),ヒカゲタ ケ属の未同定種(Panaeolus sp.),Leucocybe 属のオシロ イシメジ(Leucocybe connata),ノボリリュウタケ属の 未同定種(Helvella sp.),チャワンタケ科の未同定種が 確認された(Table 1,Fig. S3).周辺部では,裸地部に おいても確認されたコガネタケ属,ササクレヒトヨタケ 属,ヒトヨタケ属,キララタケ属,クリタケ属,スギタ ケ属,ノボリリュウタケ属に加え,アカヤマタケ属(Hy-grocybe),ハイイロシメジ属(Clitocybe),ナラタケ属 (Armillaria),クリイロムクエタケ属(Macrocystidia), モリノカレバタケ属(Gymnopus),シラウメタケモドキ 属(Hemimycena),クヌギタケ属(Mycena),ウラベニ ガサ属(Pluteus),キツネノカラカサ属(Lepiota),ヒ メヒガサヒトヨ属(Parasola),ナヨタケ属(Psathyrella), コガサタケ属(Conocybe),モエギタケ属(Stropharia), シビレタケ属(Psilocybe),ケコガサタケ属(Galerina), Rickenella属,ツノマタタケ属(Guepinia),チャワンタ ケ属(Peziza),クロサイワイタケ属(Xylaria)など, 多様な菌類が確認された. 子実体の発生時期と消長 大きく分けて,6−7 月の初夏と 9−11 月の秋に子実 体発生が多く確認された(Fig. 2,Table S1).この傾向 は外生菌根菌およびその他の腐生菌で共通していた.主 に初夏に出現した菌種はモリノカレバタケ(Gymnopus dryophilus), コ ナ ヨ タ ケ(Psathyrella obtusata),Psilo-cybe sp.,Tomentella sp.,Helvella sp. などであった.秋 にのみ出現した菌種は,シロサカズキタケ(Clitocybe hydrogramma),ハタケシメジ,ナラタケ属,コガネタケ, ウスフジフウセンタケ,オオワカフサタケ,ウスツヤト マヤタケ,キヌハダトマヤタケ,ハナイグチ,シロヌメ リイグチなどであった.初夏と秋の二時期に出現した菌 種は,ウラムラサキ,キツネタケ,ササクレヒトヨタケ, ヒトヨタケ,ムジナタケ,ニガクリタケ,ワカフサタケ, ザラツキキトマヤタケ,クロトマヤタケ,ハマニセショ ウロなどであった. 裸地部における全ての大型菌類の種数は,2004 年の 9 種(Obase et al. 2005)から緩やかに増加し,2006 年に は 23 種が確認された(Table 2).裸地部における大型 菌類の消失種数は 2004 年から 2005 年にかけて 2 種, 2005 年から 2006 年にかけて 1 種と少なかったのに対 し,新出種数は 2004 年から 2005 年にかけて 9 種,2005 年から 2006 年にかけて 8 種と比較的多かった(Table 3).周辺部における大型菌類の種数は,2004 年の 36 種 (Obase et al. 2005)から,2005 年には 49 種まで増えた が,2006 年には 46 種に減少していた(Table 2).周辺 部における大型菌類の消失種数は 2004 年から 2005 年に かけて 8 種,2005 年から 2006 年にかけて 19 種であり, 新出種数は 2004 年から 2005 年にかけて 21 種,2005 年 から 2006 年にかけて 16 種であった(Table 3).周辺部 では,2004 年から 2005 年にかけては,新出種数の方が 消失種数よりも 13 種多かったが,2005 年から 2006 年 にかけては 3 種少なかった. 次に大型菌類を外生菌根菌とその他の腐生菌に分けて
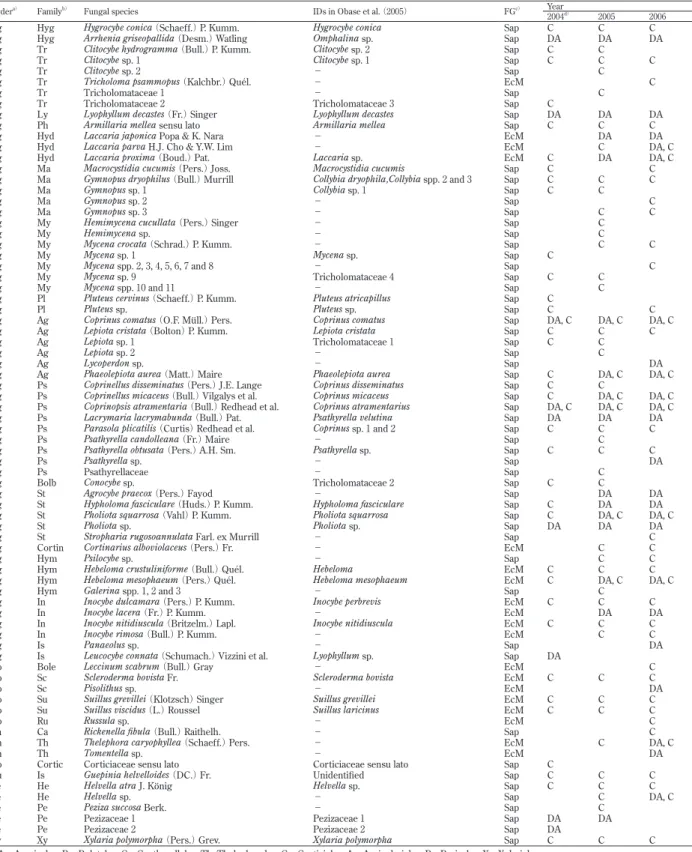
Table 1. Macrofungal species found in a denuded area (DA) and/or in its circumference areas (C) in the study site.
Ordera) Familyb) Fungal species IDs in Obase et al. (2005) FGc) Year
2004d) 2005 2006
Ag Hyg Hygrocybe conica (Schaeff.) P. Kumm. Hygrocybe conica Sap C C C Ag Hyg Arrhenia griseopallida (Desm.) Watling Omphalina sp. Sap DA DA DA Ag Tr Clitocybe hydrogramma (Bull.) P. Kumm. Clitocybe sp. 2 Sap C C
Ag Tr Clitocybe sp. 1 Clitocybe sp. 1 Sap C C C Ag Tr Clitocybe sp. 2 − Sap C
Ag Tr Tricholoma psammopus (Kalchbr.) Quél. − EcM C
Ag Tr Tricholomataceae 1 − Sap C
Ag Tr Tricholomataceae 2 Tricholomataceae 3 Sap C
Ag Ly Lyophyllum decastes (Fr.) Singer Lyophyllum decastes Sap DA DA DA Ag Ph Armillaria mellea sensu lato Armillaria mellea Sap C C C Ag Hyd Laccaria japonica Popa & K. Nara − EcM DA DA Ag Hyd Laccaria parva H.J. Cho & Y.W. Lim − EcM C DA, C Ag Hyd Laccaria proxima (Boud.) Pat. Laccaria sp. EcM C DA DA, C Ag Ma Macrocystidia cucumis (Pers.) Joss. Macrocystidia cucumis Sap C C Ag Ma Gymnopus dryophilus (Bull.) Murrill Collybia dryophila,Collybia spp. 2 and 3 Sap C C C Ag Ma Gymnopus sp. 1 Collybia sp. 1 Sap C C
Ag Ma Gymnopus sp. 2 − Sap C Ag Ma Gymnopus sp. 3 − Sap C C Ag My Hemimycena cucullata (Pers.) Singer − Sap C
Ag My Hemimycena sp. − Sap C
Ag My Mycena crocata (Schrad.) P. Kumm. − Sap C C Ag My Mycena sp. 1 Mycena sp. Sap C
Ag My Mycena spp. 2, 3, 4, 5, 6, 7 and 8 − Sap C Ag My Mycena sp. 9 Tricholomataceae 4 Sap C C
Ag My Mycena spp. 10 and 11 − Sap C Ag Pl Pluteus cervinus (Schaeff.) P. Kumm. Pluteus atricapillus Sap C
Ag Pl Pluteus sp. Pluteus sp. Sap C C Ag Ag Coprinus comatus (O.F. Müll.) Pers. Coprinus comatus Sap DA, C DA, C DA, C Ag Ag Lepiota cristata (Bolton) P. Kumm. Lepiota cristata Sap C C C Ag Ag Lepiota sp. 1 Tricholomataceae 1 Sap C C
Ag Ag Lepiota sp. 2 − Sap C
Ag Ag Lycoperdon sp. − Sap DA Ag Ag Phaeolepiota aurea (Matt.) Maire Phaeolepiota aurea Sap C DA, C DA, C Ag Ps Coprinellus disseminatus (Pers.) J.E. Lange Coprinus disseminatus Sap C C
Ag Ps Coprinellus micaceus (Bull.) Vilgalys et al. Coprinus micaceus Sap C DA, C DA, C Ag Ps Coprinopsis atramentaria (Bull.) Redhead et al. Coprinus atramentarius Sap DA, C DA, C DA, C Ag Ps Lacrymaria lacrymabunda (Bull.) Pat. Psathyrella velutina Sap DA DA DA Ag Ps Parasola plicatilis (Curtis) Redhead et al. Coprinus sp. 1 and 2 Sap C C C Ag Ps Psathyrella candolleana (Fr.) Maire − Sap C
Ag Ps Psathyrella obtusata (Pers.) A.H. Sm. Psathyrella sp. Sap C C C Ag Ps Psathyrella sp. − Sap DA
Ag Ps Psathyrellaceae − Sap C
Ag Bolb Conocybe sp. Tricholomataceae 2 Sap C C
Ag St Agrocybe praecox (Pers.) Fayod − Sap DA DA Ag St Hypholoma fasciculare (Huds.) P. Kumm. Hypholoma fasciculare Sap C DA DA Ag St Pholiota squarrosa (Vahl) P. Kumm. Pholiota squarrosa Sap C DA, C DA, C Ag St Pholiota sp. Pholiota sp. Sap DA DA DA Ag St Stropharia rugosoannulata Farl. ex Murrill − Sap C
Ag Cortin Cortinarius alboviolaceus (Pers.) Fr. − EcM C C
Ag Hym Psilocybe sp. − Sap C C Ag Hym Hebeloma crustuliniforme (Bull.) Quél. Hebeloma EcM C C C Ag Hym Hebeloma mesophaeum (Pers.) Quél. Hebeloma mesophaeum EcM C DA, C DA, C Ag Hym Galerina spp. 1, 2 and 3 − Sap C
Ag In Inocybe dulcamara (Pers.) P. Kumm. Inocybe perbrevis EcM C C C Ag In Inocybe lacera (Fr.) P. Kumm. − EcM DA DA Ag In Inocybe nitidiuscula (Britzelm.) Lapl. Inocybe nitidiuscula EcM C C C Ag In Inocybe rimosa (Bull.) P. Kumm. − EcM C C Ag Is Panaeolus sp. − Sap DA Ag Is Leucocybe connata (Schumach.) Vizzini et al. Lyophyllum sp. Sap DA
Bo Bole Leccinum scabrum (Bull.) Gray − EcM C Bo Sc Scleroderma bovista Fr. Scleroderma bovista EcM C C C Bo Sc Pisolithus sp. − EcM DA Bo Su Suillus grevillei (Klotzsch) Singer Suillus grevillei EcM C C C Bo Su Suillus viscidus (L.) Roussel Suillus laricinus EcM C C C
Bo Ru Russula sp. − EcM C
Ca Ca Rickenella fibula (Bull.) Raithelh. − Sap C Th Th Thelephora caryophyllea (Schaeff.) Pers. − EcM C DA, C Th Th Tomentella sp. − EcM DA
Co Cortic Corticiaceae sensu lato Corticiaceae sensu lato Sap C
Au Is Guepinia helvelloides (DC.) Fr. Unidentified Sap C C C Pe He Helvella atra J. König Helvella sp. Sap C C C Pe He Helvella sp. − Sap C DA, C Pe Pe Peziza succosa Berk. − Sap C
Pe Pe Pezizaceae 1 Pezizaceae 1 Sap DA DA
Pe Pe Pezizaceae 2 Pezizaceae 2 Sap DA
Xy Xy Xylaria polymorpha (Pers.) Grev. Xylaria polymorpha Sap C C C a)Ag: Agaricales; Bo: Boletales; Ca: Cantharellales; Th: Thelephorales; Co: Corticiales; Au: Auriculariales; Pe: Pezizales; Xy: Xylariales.
b) Hyg: Hygrophoraceae; Tr: Tricholomataceae; Ly: Lyophyllaceae; Ph: Physalacriaceae; Hyd: Hydnangiaceae; Ma: Marasmiaceae; My: Mycenaceae; Pl: Pluteaceae; Ag: Agari-caceae; Ps: Psathyrellaceae; Bolb: Bolbitiaceae; St: Strophariaceae; Cortin: Cortinariaceae; Hym: Hymenogastraceae; In: Inocybaceae; Is: Incertae sedis; Bole: Boletaceae; Sc: Sclerodermataceae; Su: Suillaceae; Ru: Russulaceae; Ca: Cantharellaceae; Th: Thelephoraceae; Cortic: Corticiaceae; He: Helvellaceae; Pe: Pezizaceae ; Xy: Xylariaceae. c)Functional group of fungi. EcM: Ectomycorrhizal, Sap: Saprotrophic.
見ると,2004 年に裸地部において外生菌根菌の子実体 は確認されなかったが(Obase et al. 2005),2005 年の秋 からウラムラサキ,L. proxima,ワカフサタケ,クロト マヤタケの 4 種が初めて発生し,2006 年には 8 種が確 認された(Table 2).周辺部では,2004 年には既に 8 種 の外生菌根菌の子実体が確認され(Obase et al. 2005), 以後種数は緩やかに増加し,2006 年には 15 種が確認さ れた.外生菌根菌の消失種数は,周辺部において 2004 年から 2005 年にかけて 1 種確認されたのみで,その他 の年の間では裸地部,周辺部ともに消失した種は確認さ れなかった(Table 3).外生菌根菌の新出種数は裸地部, 周辺部ともに 2004 年から 2005 年にかけて 4 種,2005 年から 2006 年にかけて 4 種であった. その他の腐生菌についてみると,裸地部では 2004 年 に 9 種が確認され,以後種数は緩やかに増加し,2006 年には 15 種が確認された(Table 2).一方,周辺部で は 2004 年に既に 28 種が確認され(Obase et al. 2005), 2005 年には 38 種に増えたが,2006 年には 31 種に減少 していた.裸地部における腐生菌の消失種数は,2004 年から 2005 年にかけて 2 種,2005 年から 2006 年にか けて 1 種と少なく,新出種数は 2004 年から 2005 年にか けて 5 種,2005 年から 2006 年にかけて 4 種と比較的多 かった(Table 3).周辺部における腐生菌の消失種数は, 2004 年から 2005 年にかけて 7 種,2005 年から 2006 年 にかけて 19 種であり,新出種数は 2004 年から 2005 年 にかけて 17 種,2005 年から 2006 年にかけて 12 種で あった(Table 3).2004 年から 2005 年にかけては,新 出種数の方が消失種数よりも 10 種多かったが,2005 年 から 2006 年にかけては 7 種少なかった. 裸地部と周辺部の菌類相の比較 両立地で共通して確認された菌種は,全 85 種のうち 11 種(外生菌根菌が 4 種,その他の腐生菌が 7 種)で 少なかった.裸地部と周辺部で共通して確認された菌類
Fig. 2. Sporocarps occurrence of ectomycorrhizal (EcM) and other saprophytic (Sap) fungi from 2004 to 2006 in a denuded area (DA) and its cirdumference areas (C) in the study site. Data in 2004 are referred from Obase et al. (2005).
Table 2. The numbers of macrofungal species found in a denuded area (DA) and/or in its circumference areas (C) in the study site.
Year
Functional group and the numbers of macrofungal species found in each areaa)
EcM Sap Total
DA C Both DA C Both DA C Both
2004b) 0 8 0 9 28 2 9 36 2
2005 4 11 1 12 38 5 16 49 6
2006 8 15 4 15 31 6 23 46 10
a) Functional group of fungi. EcM: Ectomycorrhizal, Sap: Saprotrphic. The numbers of macrofungal species found in both DA and C are indicated as Both .
の割合は,2004 年の 4.7%から,2005 年の 10.2%,2006 年の 16.9%と全体的には低いものの,緩やかに増加して いた(Table 4).外生菌根菌およびその他の腐生菌別に 見ても同様の傾向であった. 考 察 2000 年に噴火した有珠山の噴出物堆積地の裸地部で は,次年に消失した菌種は,2004−2005 年では 2 種, 2005−2006 年では 1 種と少なかったのに対して,新た に 確 認 さ れ た 菌 種 は 2004−2005 年 で は 9 種,2005− 2006 年では 8 種と多かった.従って,年経過とともに 新たな種が加わる形で菌類相は推移していた.裸地部で は,菌類が利用できる資源,例えば木材など枯死した植 物遺体が土壌中に大量に埋没されていることが考えられ た.また,そのような資源を利用して裸地環境下で子実 体を発生する菌種は限られていると考えられるので, 2004−2006 年の短い期間の中では,成長に必要な資源 を失って消失する菌種は少なかったと考えられた.また, 噴火後いち早く定着したヤナギ属,ハコヤナギ属,カン バ属の稚樹と共生関係を築いていた外生菌根菌が,2005 年秋から初めて子実体を発生したこと,そしてその後も 継続して子実体を発生し続けていたことも,菌種数の増 加に寄与していた.稚樹のサイズの増加に伴い,外生菌 根菌の子実体が発生し始めることは,他の遷移初期段階 の立地においても確認されている(Jumpponen et al. 2002).以上のように,大量の植物遺体等の資源の存在 と外生菌根性稚樹の発達が,裸地部における大型菌類相 の経時変化パターンを規定する主な要因と考えられた. これに対し,周辺部では単年のみ発生する菌種が多く, そのため消失種数,新出種数ともに高い値を示しており, 各年で確認された種数は増減を繰り返していた.森林内 では,毎年必ず子実体発生を行う大型菌類は少なく,多 くの種が年単位で散発的に発生するという報告がある (Roberts et al. 2004).また,森林内の数十年間に渡る 子実体発生調査によると,各年において新規に発見され た菌種が多く,種数累積曲線は頭打ちにならなかったと 報告している(Watling 1995, Tofts and Orton 1998).周 辺部における子実体発生の挙動は,以上の報告にある森 林内での挙動に類似していた.
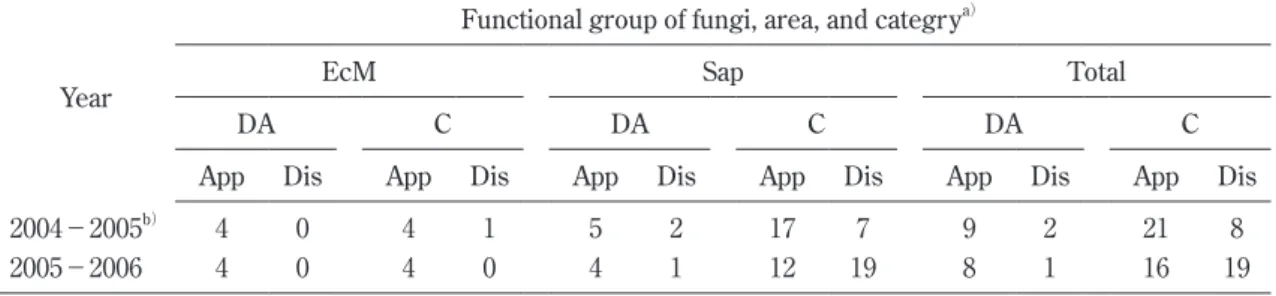
Obase et al.(2005)にあるように,2000 年有珠山噴出 物堆積地の裸地部と周辺部では,①資源の質の違い(周 Table 3. The numbers of macrofungal species newly appeared or disappeared during 2004 to 2005 and 2005 to
2006 in a denuded areas (DA) and its circumference areas (C) in the study site.
Year
Functional group of fungi, area, and categrya)
EcM Sap Total
DA C DA C DA C
App Dis App Dis App Dis App Dis App Dis App Dis
2004−2005b) 4 0 4 1 5 2 17 7 9 2 21 8
2005−2006 4 0 4 0 4 1 12 19 8 1 16 19
a) EcM: Ectomycorrhizal, Sap: Saprotrophic, App: Appearance, Dis: Disappearance. b) Data in 2004 are referred from Obase et al. (2005)
Table 4. The ratio (%) of the numbers of macrofungal species found both in a denuded area and its circumference area in the study site.
Year Functional group of fungi a)
EcM Sap Total
2004b) − 5.7 4.7
2005 7.1 11.1 10.2
2006 21.1 15.0 16.9
a) EcM: Ectomycorrhizal, Sap: Saprotrophic. b) Referred from Obase et al. (2005)
辺部ではカラマツや広葉樹類の落葉,落枝などのリター が存在するのに対し,裸地部ではリター層は無く,地中 の木材等の植物遺体が主な資源),②環境条件の違い(裸 地部は周辺部に比べて日照に曝され,乾燥しやすいため, 子実体を発生できる菌種が限られている;O dell et al. 1999, Ogaya and Peñuelas 2005),そして③植物群集構造 の違い(裸地部では 0 から 5 年生のヤナギ類の稚樹が定 着していたのに対し,周辺部では噴火以前に定着してい た様々な樹種による成木個体が生残しているため,子実 体 を 発 生 す る 外 生 菌 根 菌 が 多 様;Jumpponen et al. 2002),などが要因となって,大型菌類相は大きく異なっ ていた.しかし,裸地部と周辺部で共通して子実体を発 生していた大型菌類の種数の割合は年経過とともに増加 傾向にあった.これは周辺部で発生していた種が徐々に 裸地部で発生し始めていたためである.裸地部で子実体 を発生していた菌種が周辺部でも発生し始めた例は 2005 年から 2006 年にかけての L. proxima の 1 例のみで あるのに対し,周辺部から裸地部に進出した例はキツネ タケ,L. proxima,コガネタケ,キララタケ,スギタケ, ワカフサタケ,T. caryophyllea,Helvella sp. の 8 種で確 認された.このうち外生菌根菌 4 種については,裸地部 に定着する稚樹が外生菌根菌の子実体発生に必要な炭素 源を供給できる個体サイズにまで成長したことが大きな 要因と考えられる.その他の腐生菌では,例えばコガネ タケは徐々に子実体の発生個所を広げて,裸地部にまで 侵入した様子が確認された(Fig. S3B).菌糸体や胞子 分散で生息域を広げたことが要因と考えられた. 以上のように,2000 年有珠山噴出物堆積の裸地部で は,年経過とともに大型菌類の種数が緩やかに増加し, 周辺部の大型菌類が少しずつ裸地部で繁殖し始めている 状況を捉えることができた.今後は,地中に存在する木 材等の資源の枯渇,植物遺体の蓄積,植生の発達など, 年経過に伴う資源の量・質の変化や土壌の養水分環境の 変化などの諸要因を反映して,子実体を発生する大型菌 類相は大きく変化していくと予想される.火山噴火に対 する大型菌類の応答パターンを理解するには,例えば噴 火から植生が回復して森林が再生していくまでの,より 長期的な視点に立ったモニタリング調査とともに,他の 噴火跡地における研究事例を積み重ねていくことが重要 である. 摘 要 2000 年に噴火した有珠山の噴出物堆積地において, 2004 年に調査した既報告の大型菌類の発生調査を 2006 年まで継続して行い,噴火から間もない遷移初期の立地 で繁殖する大型菌類相の経年変化を明らかにした.菌種 数は,2004 年の 9 種から緩やかに増加し,2006 年には 23 種が確認された.外生菌根菌は 2005 年秋に初めて 4 種が確認された.2006 年も新たに 4 種が発生し,計 8 種が確認された.その他の腐生菌は,2004 年に確認さ れていたナヨタケ科,モエギタケ科,キシメジ科の菌類 が継続して発生しており,種数は年経過とともに緩やか に増加した.次年度に消失した菌種は,2004−2005 年 では 2 種,2005−2006 年では 1 種と少なかったのに対 して,新たに確認された菌種は 2004−2005 年では 9 種, 2005−2006 年では 8 種と多かった.噴出物堆積地では, 埋没した植物遺体や更新稚樹を資源として利用する菌類 が繁殖しており,その菌類相は年経過とともに新たな種 が加わる形で推移していた. 引用文献
Akers BP, Sundberg WJ (2001) Lepiotaceae of Florida, IV. Stenosporic species of Lepiota s. str. Mycotaxon 80 : 469−479
Banerjee P, Sundberg WJ (1995) The genus Pluteus sec-tion Pluteus (Pluteaceae, Agaricales) in the midwest-ern United States. Mycotaxon 53: 189−246
Bigelow HE (1982) North American species of Clitocybe, Part I. Nova Hedwigia Beiheft 72: 1−280
Breitenbach J, Kränzlin F (1984) Fungi of Switzerland. Volume 1 Ascomycetes. Verlag Mykologia, Lucerne, Switzerland
Breitenbach J, Kränzlin F (1986) Fungi of Switzerland. Volume 2 Non gilled fungi. Mykologia, Lucerne, Swit-zerland
Breitenbach J, Kränzlin F (1991) Fungi of Switzerland. Volume 3 Boletes and agarics 1st part. Mykologia Lu-cerne, Switzerland
Breitenbach J, Kränzlin F (1995) Fungi of Switzerland. Volume 4 Agarics 2 nd part. Mykologia, Lucerne, Switzerland
Breitenbach J, Kränzlin F (2000) Fungi of Switzerland. Volume 5 Agarics 3 rd part. Mykologia Luzern, Swit-zerland
Brown N, Bhagwat S, Watkinson S (2006) Macrofungal diversity in fragmented and disturbed forests of the Western Ghats of India. J Appl Ecol 43: 11−17 車柱榮(1999)北海道産ナラタケの分類と生態.日菌報
40: 155−164
and morphological characteristics of Armillaria mel-lea complex in Hokkaido: A. sinapina and two new species, A. jezoensis and A. singula. Mycoscience 35: 39−47
Cha JY, Sung JM, Igarashi T (1992b) Biological species and morphological characteristics of Armillaria mel-lea complex in Hokkaido: A. ostoyae and A. bulbosa. Res Bull Exp For Hokkaido Univ 49: 185−194 Corner EJH (1968) A monograph of Thelephora
(Basidio-mycetes). Nova Hedwigia Beiheft 27: 1−110
Cripps CL (1997) The genus Inocybe in Montana aspen stands. Mycologia 89: 670−688
Edwards IP, Cripliver JL, Gillespie AR, Johnsen KH, Schol-ler M, Turco RF (2004) Nitrogen availability alters macrofungal basidiomycete community structure in optimally fertilized loblolly pine forests. New Phytol 162: 755−770
後藤春利(1937)十勝岳爆発後十年間の植生遷移(火山 爆発と植生).日林誌 19: 537−550
Horikoshi T, Tateishi T, Takahashi F (1986) Changes of fungus flora after fires in Pinus densiflora forest. Trans Mycol Soc Jpn 27: 283−295
Imai S (1938a) Studies on the Agaricaceae of Hokkaido I. J Fac Agr Hokkaido Imp Univ 43: 1−178
Imai S (1938b) Studies on the Agaricaceae of Hokkaido II. J Fac Agr Hokkaido Imp Univ 43: 179−378 今関六也・本郷次雄(1987)原色日本真菌類図鑑(I).
保育社,大阪
今関六也・本郷次雄(1989)原色日本真菌類図鑑(II). 保育社,大阪
Jumpponen A, Trappe JM, Cázares E (2002) Occurrence of ectomycorrhizal fungi on the forefront of retreat-ing Lyman Glacier (Washington, USA) in relation to time since deglaciation. Mycorrhiza 12: 43−49 気象庁(2017)火山噴火予知連絡会による新たな活火山の
選 定 に つ い て.(http://www.jma.go.jp/jma/press/ 1706/20c/new-volcano170620.html,アクセス日:2020 年 3 月 28 日)
Kit van Waveren E (1985) The Dutch, French and British species of Psathyrella. Rijksherbarium, Leiden, Neth-erlands
Kränzlin F (2005) Fungi of Switzerland Volume 6 Russula-ceae. Mykologia, Lucerne, Switzerland
Kuyper TW (1986) A revision of the genus Inocybe in Eu-rope. 1. Subgenus Inosperma and the smooth-spored species of subgenus Inocybe. Persoonia Suppl 3 : 1−
247
López-Quintero CA, Straatsma G, Franco-Molano AE, Boekhout T (2012) Macrofungal diversity in Colom-bian Amazon forests varies with regions and regimes of disturbance. Biodivers Conserv 21: 2221−2243 McMullan-Fisher SJ, May TW, Keane PJ (2002) The
mac-rofungal community and fire in a Mountain Ash for-est in southern Australia. Fungal Divers 10: 57−76 McNabb RFR (1968). The Boletaceae of New Zealand. N
Z J Bot 6: 137−176
Mueller GM (1992) Systematics of Laccaria (Agaricales) in the Continental United States and Canada, with discussions on extralimital taxa and descriptions of extant types. Fieldiana Bot 30: 1−158
Nara K, Nakaya H, Hogetsu T (2003) Ectomycorrhizal sporocarp succession and production during early primary succession on Mount Fuji. New Phytol 158: 193−206
小長谷啓介(2009)2000 年有珠山噴出物堆積地におけ る植生再生初期動態と菌根共生に関する研究.北大 演習林研報 66: 1−31
Obase K, Tamai Y, Miyamoto T, Yajima T (2005) Macro-fungal flora on the volcano Usu, deforested by 2000 eruptions. Eurasian J For Res 8: 65−70
Obase K, Tamai Y, Yajima T, Miyamoto T (2007) Mycor-rhizal associations in woody plant species at the Mt. Usu volcano, Japan. Mycorrhiza 17: 209−215
O dell TE, Ammirati JF, Schreiner EG (1999) Species richness and abundance of ectomycorrhizal basidio-mycete sporocarps on a moisture gradient in the Tsu-ga heterophylla zone. Can J Bot 77: 1699−1711 Ogaya R, Peñuelas J (2005) Decreased mushroom
pro-duction in a holm oak forest in response to an experi-mental drought. Forestry 78: 279−283
大園享司(2018)菌類生態学.共立出版,東京
Palm ME, Stewart EL (1986) Typification and nomencla-ture of selected Suillus species. Mycologia 78: 325− 333
Pegler DN, Læssøe T, Spooner BM (1995) Brirish puff-balls, earthstars and stinkhorns: An account of the British gasteroid fungi. Royal Botanic Gardens, Kew Reid DA, Eicker A (1999) South African fungi 10: new
re-cords and some new observations. Mycotaxon 73 : 169−197
Roberts C, Ceska O, Kroeger P, Kendrick B (2004) Mac-rofungi from six habitats over five years in Clayoquot
Sound, Vancouver Island. Can J Bot 82: 1518−1538 Rudolf K, Morschhauser T, Pál-Fám F (2012)
Macrofun-gal diversity in disturbed vegetation types in North-East Hungary. Open Life Sci 7: 634−647
Sato H, Morimoto S, Hattori, T (2012) A thirty-year sur-vey reveals that ecosystem function of fungi predicts phenology of mushroom fruiting. PloS one 7: e49777 Smith AH (1951) The North American species of
Naema-toloma. Mycologia 43: 467−521
Smith AH (1972) North American species of Psathyrella. Mem NY Bot Garden 24: 1−633
Smith AH, Evenson VS, Mitchel DH (1983) The veiled species of Hebeloma in the western United States. University of Michigan Press, Ann Arbor
Smith SE, Read DJ (2010) Mycorrhizal symbiosis. Aca-demic press, Tokyo
Stangl J (1983) Inocybe nitidiuscula (Britz.) Sacc. Int J Myc Lich 1: 161−168
Stangl J (1989) Die Gattung Inocybe in Bayern. Hoppea 46: 5−388
Tofts RJ, Orton PD (1998) The species accumulation curve for agarics and boleti from a Caledonian pine-wood. Mycologist 12: 98−102
Tsuyuzaki S (1989) Analysis of revegetation dynamics on the volcano Usu, northern Japan, deforested by 1977−1978 eruptions. Am J Bot 76: 1468−1477 Uljé CB, Bas C (1988) Studies in Coprinus-I. Subsections
Auricomi and Glabri of Coprinus section Pseudocopri-nus. Persoonia 13: 433−448
Uljé CB, Bas C (1991) Studies in Coprinus-II. Subsection Setulosi of section Pseudocoprinus. Persoonia 14 : 275−339
van de Bogart F (1976) The genus Coprinus in western north America, Part I. Section Coprinus. Mycotaxon 4: 233−275
van de Bogart F (1979) The genus Coprinus in western north America, Part Ⅲ . Section Atramentarii. Myco-taxon 10: 155−174
Vesterholt J (1995) Hebeloma crustuliniforme and related taxa - notes on some characters of taxonomic impor-tance. Symb Bot Ups 30: 129−137
Vesterholt J (2005) The genus Hebeloma. Fungi of North-ern Europe vol. 3. Narayana Press, Gylling, Denmark Vilgalys R, Miller OK (1987) Morphological studies on
the Collybia dryophila group in Europe. Trans Br My-col Soc 88: 461−472
Watling R (1995) Assessment of fungal diversity: macro-mycetes, the problems. Can J Bot 73(S1): 15−24 吉見昭一(2002)日本産ニセショウロ属.日菌報 43:
3−18
Young AM, Wood AE (1997) Studies on the Hygrophora-ceae (Fungi, Homobasidiomycetes, Agaricales) of Australia. Aust Syst Bot 10: 911−1030