日本産フクジュソウの植物学的研究(第2部)
26
0
0
全文
(2) ^mm^±^es w 2 gp B ) ^39^ ^ 2 ^ ^^jc^ 3 ^. Journal of Hokkaido University of Education (Section II B) Vol. 39, No. 2 March. 1989. Botanical Studies on Adonis amurensis Regel et Radde. in Japan (Part 2) ^ Tsunehiko NlSHIKAWA Biological Laboratory, Asahikawa College, Hokkaido University of Education, Asahikawa 070. i^fi^CT? t^SS — At? 'i 8i B:$:jt7^y^yr>®1tf^?£% (Hr.gis)^. J|13 j^. wm^±^w \v^±^m 33. Cultivars 331, Chromosomes Somatic chromosome number: Photo 12 showed that the chromosome numbers of Adonis cultivars were 2n = 16, 24, 32, and 40. Table 19 lists the chromosome numbers of 24 Adonis cultivars and their pollen fertility. Although 2n = 16 and 2n = 32 are prevalent in wild populations of Adonis species, 2n = 24 was seldom found and 2n = 40 has never been found. Of the 24 cultivars, 2n == 24 was found in seven cultivars, and 2n = 40 was found in only two cultivars. The above facts suggest that the basic chromosome number of genus Adonis is x = 8 (Darlington & Wylie, 1955 ; Fedorov, 1969). Meiosis : Of seven triploid cultivars (2n = 24), five cultivars were used in the meiosis observation. They were as follows : cvs. Kinshi, Beninadeshiko, Kmsekai, and Fukujukai, All of them showed irregular behavior. In most PMCs, univalent and multivalent were often seen in addition to bivalents. At diakinesis and metaphase I the configuration was so complicated in most of the PMCs that analysis of chromosome pairing was possible only in the two PCMs with an exceptionally simple configuration of cv. Fukujukai. The chromosome configurations found were IV + 5III + III + 21 (Photo 13-A) and 2IV + 511 + 61. At anaphase I and anaphase II the chromosomal bridges and lagging chromosomes were often observed (Photo 13-C and B). Triads, diads, and monoads were observed in addition to tetrads, and double nuclei in one cell of the tetrad were seen (Photo 13-D). Pollen grains were irregular in shape and lacked protoplasmic contents. However, considering that the chromosome number of "Adonis amurensis" has been reported to be n = 12 and 2n = 24 by senior authors (Ishikawa, ^ Continued from Jour. Hokkaido University of Education (Sect. II B) 39 (1): 1-35.. (D.
(3) Tsunehiko NlSHIKAWA. Photo 12, Somatic chromosomes of Adonis cultivars. A: 2n = 16 (cv. Hinomotoko). B : 2n = 24^ (cv. Beninadeshiko). C: 2n = 32 (cv. Garaku). D: 2n = 40 (cv. Fukurokuju). Scale indicates 10 ^ m.. 1916 ; Takamine, 1916 ; Kurita, 1955, 1957), it is interesting that the chromosome number of 12 + 12 was sometimes found in the anaphase I (Photo 13-E). This fact suggests that the previous reports on the meiosis may be based on horticultural plants.. 332. Morphology Shape of petals and seplas : In all cultivars, petals were mostly elliptic to obovate : sepals were mostly elliptic, often obovate except diploids in which sepals were mostly obovate, not elliptic (Table. 20, Photo 14). Petal color: On the basis of the inside color of petals, three groups were recognized : (1) reddish orange group, including cvs. Hinodeko (2n = 16), Hinomotoko (2n = 16), Beninadeshiko (2n = 24), and. Chichibuko (2n = 32). (2) pale yellow group, including cvs. Shiun (2n = 24) and Benten (2n = 32). (3) bright yellow to golden yellow group, including cvs. Kotobuki (2n = 16), Fukujukai (2n = 24), Kinsekai (2n = 24), Jitsugetsusei (2n == 24), Nadeshiko (2n = 24), &Msfet (2n = 24), GflmfeM (2n = 32),. (2).
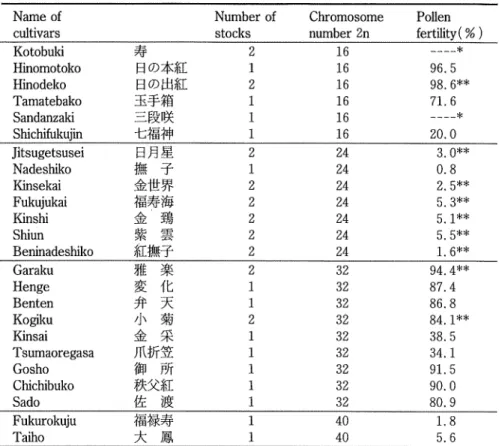
(4) Botanical Studies on Adonis amiirensis Regel et Radde in Japan (Part 2) Table 19. Chromosome number and pollen fertility otAdonis cultivars.. Number of. Name of cultivars. Kotobuki Hinomotoko. Hinodeko. Tamatebako. Sandanzaki Shichifukujin. Jitsugetsusei Nadeshiko Kinsekai Fukujukai. Kinshi. Shiun Beninadeshiko Garaku Henge Benten. Kogiku Kinsai. Tsumaoregasa. Gosho Chichibuko Sado. Fukurokuju Taiho. stocks. 2 1 2 1 1 1 2 1 2 2 2 2 2 2 1 1 2 1 1 1 1 1 1 1. B®2?. BO^ffi a^% ^^K -?tt Q^S. » -?. ^w a. Wf. m^. ^ \t. ^ ^. ^% ^ $R JTW^ ?l pft. a^a:. fe V. :?s%^. ±B. Pollen. Chromosome. fertility (%). number 2n. 16 16 16 16 16 16 24 24 24 24 24 24 24 32 32 32 32 32 32 32 32 32 40 40. „„„„*. 96.5 98.6** 71.6 ____*. 20.0 3.0** 0.8 2.5** 5.3** 5.1** 5.5** 1.6** 94.4**. 87.4 86.8 84.1** 38.5 34.1 91.5 90.0 80.9 1.8 5.6. * All stamens and pistils are transformed into petals. * * Average. Sado (2 = 32), ffew^e (2n = 32), Gosho (2n = 32), Kogiku (2n = 32), Kinsai (2n = 32), Tsumaoregasa (2n = 32), Fukwokuju (2n = 40) and Taiho (2n = 40). The outside color of the petals shows a good correlation with the ploidy. The triploids, tetraploids, and pentaploids were commonly yellow, tinged with purple or dark purple, whereas the diploids were reddish brown on the upper margin of the petals (Table 20). Sepal color: Except for the diploid cultivars, sepals were pale green inside and pale green or dark. green outside or pale yellow inside and dark purple to purple outside (Table 20). Table 20. Comparison of shape and color of petals and sepals among 4 polyploid groups Chromosome Shape of Shape of number 2n petals sepals. 16 24 32. inside. outside. yellow with reddish. obovate. elliptic to obovate. elliptic, often obovate. usually yellow, not fixed. 40. (3). inside. outside. brown on the upper margin. pale green. pale green to dark green. usually yellow, sometimes purple to dark purple. yellow to pale yellow. purple to dark purple.
(5) Tsunehiko NlSHIKAWA. 'w. ,ff. ^•- '^. .«?y. >.„.;-" d. it"^':. ^'. -'^<.".. ^. i' ^•Q^'V Ifa&ti^-. ^. :B. ft. -T-fy". ^'. »A If '". 51. 12. 10. ^. ^^,%r5y^< ^&E%.^'';q. .c r. <k>,;. ^:. 7. 10. 12. Photo 13. Meiotic chromosomes of cultivars with 2n =24. A : IV + 5III + III + 21. (cv. Fukujukai). B : Lagging chromosomes at late anaphase II (cv. Fukujukai). C : Chromosome bridge at anaphase I (cv. Fnknjnkai). D : Double nuclei in one cell of tetrad. E : Anaphase I, 12 + 12 chromosomes (cv. Fuknjukai). Scale indicates 10 /um.. The relation of sepal length to petal length: The diploid and triploid cultivars were characterized by having petals longer than sepals : the tetraploids were characterized by having petals subequal to sepals. Among them, the diploids were the shortest in length (Fig. 13). Number of flowers per stem '. The number of flowers per stem ranged from 1 to 9. However, it was very conspicuous that most of the tetraploids had one flower per stem, and that the triploids had 3 to 8, an average of 4.9 (Table 21).. Table 21. Number of flowers per stem Chromosome number 2n. 16 24 32 40. Number of cultivars. 6 7 9 2. Number of stocks. 1. 8 13 11 2. 0 0 8 0. No. of flowers per stem. 2. 3. 4. 5. 6. 7. 8. 9. 1 0 1 1. 3 2 1 0. 1 3 1 1. 1 4 0 0. 0 3 0 0. 1 0 0 0. 0 1 0 0. 1 0 0 0. 4.5 4.9 1.5 3.0. Fruit: The achenes were observed on 13 fertile stocks. In the diploids, achenes were smaller in size, paler in color, and thinner in hairiness than those in the rest.. Aggregate fruit of the diploids was different from that of the rest (Photo 15). In the diploids it. (4).
(6) Botanical Studies on Admits amurensis Regel et Radde in Japan (Part 2). ®2n=l6 X2n=2A A2n=32 02n=^0. -4048. 12mm. PETAL LENGTH - SEPAL LENGTH. Fig. 13. Scatter diagram showing the relationship of "petal length minus sepal length" to "sepal length" of Adonis cultivars.. H Photo 14. Shape of sepals and petals. A : cv. Tamatebako (2n == 16). B : cv. Sandanzaki (2n. =16). C : Schichifukujin. (2n = 16). D : cv. Fukujukai. (2n == 24). E : cv. Fukw'okuju (2n = 40). F : cv. Beninadeshiko (2n = 24). G : cv. Shiun (2n = 24). H cv. Kinshi (2n = 32). Scale indicates 2cm.. B Photo 15. Aggregate fruits of Adonis cultivars. A: 2n = 16 (cv. Hinomotoko). B : 2n = 24 (cv. 7Cws/;.(). C: 2n = 32 (cv. Henge). D: 2n = 40 (cv. Fitkurokuju). Scale indicates 10mm.. was globose and smaller than. that in the rest. A similar tendency was shown in the aggregate fruit of. wild Honshu diploids collected at Towada (Photo 6-B, 2). Hairiness of leaves: No differences were observed among the cultivars in the hairiness of leaves. They were commonly subglabrous to glabrous beneath, and glabrous above.. Flowering time : The flowering time of the diploids was later than that of the rest. Table 22 shows flower opening date in 1982 in Asahikawa in a cultivated condition. As seen in the table, the di-. ploids blossomed from late April to early May; the rest finished the flowering by late April. Pollen fertility : Table 19 and Photo 16 show pollen fertility. It was interesting that the pollen fertility of the even number polyploids was higher than that of the oddpolyploids. In the former, the pollen fertility was 75.0% on the average and the latter only 3.5% on the average. But it was exceptionally low, being from 20 to 39%, in cvs. Shichifukujin (2n = 16), Tsumaoregasa (2n == 32), and Kinsai (2n = 32). In some flowers of pentaploid cultivars, solid anthers sometimes contained gigantic pol-. (5).
(7) Shichinohe (Aomori Pref. Hirosaki (Aomori Pref. ) Asahikawa (Hokkaido) Toyoura (Hokkaido). Kitami (Hokkaido). § Maruseppu (Hokkaido). 2 Towada (Aomori Pref.. Nishine (Iwate Pref.). Gosho Fukurokuju Taiho. Tsumaoregasa. Kogiku Kinsai. Benten. Chichibuko Sado. Garaku Henge. Jitsugetsusei ^ Nadeshiko § Kinsekai Fukujukai Kinshi 0 Shiun Beninadeshiko. Shichifukujin. Tamatebako Sandanzaki. Kotobuld Hinomotoko Hinodeko. localities of wild plants. Names of cultivars and. 16 16 16 16 32 32 32 32. 16 16 16 16 16 16 24 24 24 24 24 24 24 32 32 32 32 32 32 32 32 32 40 40. Chromosome number 2n. Date of flower opening. +. +. + +. +. +. +. +. +. +. + +. +. +. +. +.. + +. + +. +. 14 16 18 20 22 24 26 28 30. April. +. Table 22. Date of flower 'opening in 1982 of Adonis cultivars and wild Adonis cultivated in Asahikawa.. +. +. 8 10 12 +. May. I. ?. 0. z.
(8) Botanical Studies on Adonis amurensis Regel et Radde in Japan (Part 2). Photo 16. Pollen grains otAdonis cultivars. A: 2n = 16 (cv. Hinodeko). B : 2n = 24 (cv. Kinsekai). C: 2n = 32 (cv. Garaku). D: 2n = 40 (cv. Taiho). Scale mdicates 40 //m, len grams which were about 3.5 times as large as the normal-size, and also empty anthers were often observed. I considered that this resulted from disturbed meiosis in relation to their irregular meiosis.. 4. Discussion and Conclusion. 41. Basic chromosome number In the present studies, four kinds of chromosome numbers for Adorns amurensis were observed, that is, 2n = 16, 24, 32, and 48. As shown in Table 23, the chromosome numbers of the genus Adonis are almost all 2n = 16 and 32, so that the basic number is believed to be x = 8 (Darlington & Wylie, 1955 ; Fedorov, 1969). Thus, the series of the chromosome number mentioned above are the diploid, the triploid, the tetraploid and the hexaploid. Sometimes, however, a problematic issue occur-. red. Ishikawa (1916), Takamine (1916), and Sugiura (1913) reported that the meiotic chromosome numbers were n = 12 and n = 20 for the Japanese materials under the names of A. davurica, and A. vernalis var. amurensis, respectively. If this were so, the basic number should have been considered to be x = 4 (Kurita, 1957), and the series mentioned above should be altered to be the tetraploid, the hexaploid, the octaploid, and the dodecaploid. In the present studies, the pollen fertility of the 2n = 24 specimens used was not so high as expected, being about 1% on the average. In addition to this, the chromosomal behavior in the meiosis was considerably irregular. These results support the conclusion that 2n = 24 specimens are the triploid but not the hexaploid, because in the 2n = 16 and 2n = 32 specunens which were supposed to be. (7).
(9) Tsunehiko NlSHIKAWA Table 23. The chromosome numbers of the species in the genus Adorns Taxa Adonis aestivalis. Chromosome numbers. 2n=32. References Langlet 1927. Gregory 1941. 1). Kurita 1963.. A.. aestivalis. ssp. aestivalis. aestivalis ssp. parviflora A. aestivalis A.. A.. ssp. squarrosa amurensis. n=16. 2n=32. Shlangena 1976.. 2n=16, 32, 48. Shlangena 1976.. 2n=32 2n=16. Shlangena 1976. Sokolovskaya 1966. Lee 1967. SUangena 1976. Kurita 1955. Kurita 1957. Kurita 1963. Shlangena 1976.. 2n=24 A.. annua. A.. annua ssp. cuspiana apemna. A. A.. autumnalis. Poganetal. 1?80. 4> Mui-ml978. 4) Hindakokova 1978. 4> Bhatetal. 1972.. 2n=32 2n=16 2n=16 2n=32. Marchietal. 1974. 4) Langlet 1927. Langlet 1927. Gregory 1941,., 1). DekTyl947. 1). A.. chrysocyatha. 2n=16 n=16. A.. dahurica. 2n=24. A. A. A.. flammea leiosepala. pyrenaica. 2n=32 2n=16 2n=16. A.. sibirica. 2n=16. Shragerl980. 6),. A. A. A. A.. sibiricus ex Ledeb. tianschanica. 2n=16 2n=16 2n=16 2n=16. Rostovtseva 1976. 4) Shlangena 1976. Shlangena 1976. Langlet 1927.. twkestanica vernalis. Shlangena 1976. Mehra & Ramanandan 1972. Ishikawa 1916. Takamine 1916. Langlet 1927. Langlet 1927. Shlangena 1976. Langlet 1927. Kupferl974. 3^,. Shlangena 1974. 3) Shlangena 1976. Kartashova etal. 1974. 4). Turessonl938,, 1). Larsenl954. " Skalinska. et al. 1964.„. n=8 A.. vernalis var. amurensis. 2n=40. A. A.. villosa wolgensis. 2n=16 2n=16. D. 4). See Frdorov 1969. See Goldblatt 1981.. Zhukoval967. 2) Shlangena 1976. Uhrikoval978. 4). Zickler &. Lambert 1967. z). Sugiura 1931. Sugiura 1936. Shlangena 1976. Shlangena 1976.. 2) See Moore 1973. 3) See Moore 1977.. 5) See Goldblatt 1984.. (8). 6) See Goldblatt 1985..
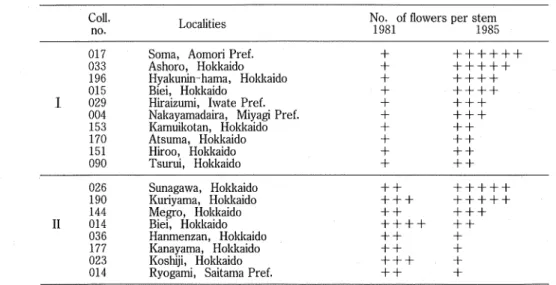
(10) Botanical Studies on Adonis amwensis Regel et Radde in Japan (Part 2). the diploid and the tetraploid, the pollen fertility was absolutely high, ranging from 91.8% to 97.9%, and the pollen production was normal in both quantity and quality. In the interpretation of the chromosome number of the Japanese materials of "Fukujuso", there were some problems, as Hisauchi (1941) and Hara (personal communication) pointed out, because the materials used by cytologists might be those derived from a commercial source', that is, from the hybridizations since the Edo Period (A. D. 1603-1867). In the cases of Ishikawa, Takamine, and Sugiura, their materials may be one of cultivars, which had easily been obtained from common markets or private gardens, as shown in Chapter 3. Again, it is further proof that the 2n = 24 specimens seldom occur in wild populations but commonly occur in cultivars.. 42. Morphology There are some discrepancies between the 2n = 16 Hokkaido specimens and the 2n = 16 Honshu specimens in morphological characteristics : the ratio of sepal length to petal length, number of flowers per stem, cross section of stem, and hairiness of leaves. Furthermore, a difference is found in the initiated seasons of the meiotic divisions : middle September with the the former, and middle December to early January with the latter. From the facts mentioned above, the 2n = 16 Hokkaido specimens are seen as clearly different from the 2n == 16 Honshu specimens. In the case of the 2n = 32 specimens, it may be said that the Hokkaido specimens and the Honshu specimens are included in the same category. The 2n = 32 specimens, however, have two forms in terms of number of flowers per stem ; one is a one-flowered form, and the other is a multi-flowered form. Since the one-flowered form of 2n = 32 specunens changed to the multi-flowered form, and the reciprocal change was also found in cultivation (Table 24), the number of flowers is not fixed but is changeable in this case, which is different from the case of the 2n = 16 specimens. Thus, I do not think that the number of flowers per stem is essential in the former case. Table 24. No. of flowers on a stem m 1981 and 1985 Coll. T nn»i;t.;B» No'-.of flowers per stem no. i.ucauues ^gg^ - ^ggg. 017 Soma, Aomori Pref. + ++ 033 Ashoro, Hokkaido + 196 Hvakunin-hama, Hokkaido + 015 Biei, Hokkaido +. I 029 Hu-aizumi, Iwate Pref. +. 004 Nakayamadaira, Miyagi Pref. + 153 Kamuikotan, Hokkaido + 170 Atsuma, Hokkaido + 151 Hiroo, Hokkaido + 090 Tsurui, Hokkaido +. 026 Sunagawa, Hokkaido + + 190 Kuriyama, Hokkaido +++ 144 Megro, Hokkaido + + H 014 Bief, Hokkaido + 036 Hanmenzan, Hokkaido + + 177 Kanayama, Hokkaido + + 023 Koshiji, Hokkaido +++ 014 Ryogami, Saitama Pref. ++ + indicates one flower. I : the group of one-flowered form in 1981.. II: the group of multi-flowered form in 1981.. (9).
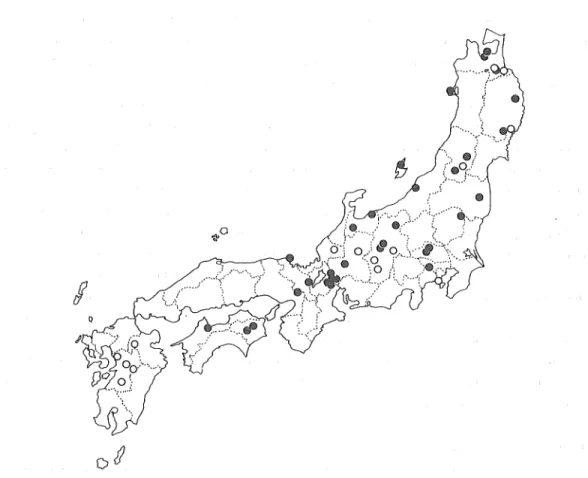
(11) Tsunehiko NlSHIKAWA. 43. Geographical distribution The 2n = 32 specmiens occupy northern, central, and south-westem Hokkaido, while the 2n = 16 Hokkaido specimens are distributed mainly in the north-eastem of Hokkaido (Fig. 2), which is characterized by light snowfall in winter, and a cool climate in summer. In some localities of Hokkaido, such as near Mt. Yubari, Notoro-misaki, Rushin, and'Atsunai, where 2n = 32 specimens are dominant or prevailing, 2n == 16 specimens prefer special habitats : they are limited to open fields or on the edges of coniferous or deciduous forests, where the soil is frozen at the early flowering time, while the 2n = 32 specimens were found on the edges of deciduous forests or in the interior of deciduous forests, where the soil is not frozen and the snow often remains here and there at early flowering time. In the case of Notoro-misaki, 2n = 16 specimens shared their habitats with 2n == 32 specimens, in which the former established their population on the top of grassland exposed to severe winter conditions such as strong seasonal winds, frozen soil, etc. ; but the latter were established in the deciduous forest, not conditioned by frozen soil in the interior part of Notoro-misaki. The 2n = 16 Honshu specimens are distributed mainly on the Pacific Ocean side of Honshu, where snowfall is usually light, and they were usually found in gentle slopes on open grassland and under sparse lower shrubs, but not on deciduous forests floors. According to my observation, under cultivation in Asahikawa, dormant buds of the 2n == 16 Hoshu specimens were below the soil surface till the snow disappears m early April, while those of the rest appear on the earth in late November and throughout winter. Thus, judging from the evidence of distribution range, habitat preference, and habits of dormant buds passing the winter season in all cytological races, it is suggested that one of the factor? affecting their distribution is snowfall, becuse the snowfall has a protective effect on plants due to the coldness and the drying in winter season by being covered with snow. Identification of herbarium specimens was carried out based on the above-mentioned results. About 170 specimens were used but cultivars were omitted. As a result, herbarium specimens could be divided into three groups : the 2n = 16 Hokkaido plants, the 2n = 16 Hoshu plants, and the 2n = 32 plants (cf. appendix 1). This examination of herbarium specunens was particularly useful for elucidating the geographical distribution of 2n = 16 and 2n = 32 Honshu plants (Fig. 14). The most important facts obtained m the examination of living plants and the survey of the herbarium specimens were as follows : (1) the range of the 2n = 32 plants clearly overlaps the ranges of the. 2n == 16 Hoshu plants and the 2n = 16 Hokkaido plants, (2) the range of the 2n = 16 Hokkaido plants extends along the Okhotsk Sea side to the Pacific Ocean side of Eastern Hokkaido, and also occurs in Sakhalin, (3) the range of the 2n =:: 16 Honshu plants extends from Aomori Pref. to Kumamoto Pref. southward, in which the plants are found in the mountain range, and as well as occurs in Korea, and (4) the 2n = 16 plants of Hokkaido and Honshu are considered to be the relict, and the 2n = 32 plants to be endemic, in Japan. The reason for such an overlap of distribution range is assumed to be that the southern race of the Honshu plants had probably come across the northern race of the 2n = 16 Hokkaido plants in some places of Honshu Proper in a certain geologic period, and that the hybridization might have occurred between them there. In the case of the Adonis, the 2n = 16 Honshu plants extend from Aomori Pref. to Kumamoto Pref., while the 2n = 16 Hokkaido plants are found mainly in NE Hokkaido and near Mt.. (10).
(12) Botanical Studies on Adorns amurensis Regel et Radde in Japan (Part 2). 0. s. Fig. 14. Distribution map of Adonis multiflora and A. ramosa based on herbarium specimens. 0: A. multiflora. • : A. ramosa. Yubari across the Hidaka mountains, in which their populations are isolated from the 2n = 16 population. Such a distribution range might have been established by the repetition of migration-movment associated with the climatic oscillations. The migration-movement courses could be a repetition of those from the south to the north, or vice versa, along mountain systems which help the migrationmovement from the north to the south and often disturb the migration-movement from the east to the west,. When plants migrate southward or northward, they are influenced by photoperiodism, which plays an important role in the geographical distribution because the natural day lengths vary with latitude as well as with the seasons. Sensibility to photopreiodism of a plant is one of the genetic characteristics. (Oka, 1958 ; Panji & Srinivans, 1959 ; Ray & Alexander, 1966 ; Takahashi & Yasuda, 1970 ; McMillan, 1970). In the present studies, I showed that the beginning of meiotic divisions in the 2n = 16 Hokkaido plants is middle September, and that of the 2n == 16 Honshu plants is late December to early January. Furthermore, the distribution ranges of the Hokkaido plants and the Honshu plants were shown to be the northern part and the southern part of Japan, respectively. This fact shows that early meiotic plants correspond to plants of the northern range, while late ones correspond to those of the southern. (11).
(13) Tsunehiko NlSHIKAWA range. The same result was observed in Caltha plants in Japan (Nishikawa, 1987) : the beginning of meiotic divisions is middle January to early February in C. fistulosa of the northern race, and is March in C. palustris of the southern race. A similar tendency was observed in other species : Dendranthema arcticum (n. race) vs. D. arcticum subsp. maekawanum (s. race), Cirsiwn pectinellum and C. kamtschaticum (n. race) vs. Cirsium aomorense and C. heiianum (s. race) (Compositae), and Aconitum yezoense (n. race) vs. A. japonicum (s. race) (Ranunculaceae) (Nishikawa, unpublished). Most of the southern races have their southern limit of distribution m central or northern Honshu and have their northern limit in Hokkaido. Such a difference in ranges has been caused by the migration-movement along mountain systems northward or southward so that the flower-opening clock of plants could have deviated from the physical clock of the habitat. However, the plants may conserve the same photo-. periodic response m different habitats as that in the previous habitats due to genetic heredity. This fact serves as an explanation of the synchronous initiation of meiosis of Adonis plants collected from Nagano Pref. and Aomori Pref. in a cultivated condition in Ashikawa, which is located to the north of those habitats (Fig. 5). The same results were observed in the 2n = 16 Hokkaido plants and the 2n = 32 plants. Considering the present distribution ranges, the geographical difference in photoperiodism might reflect different histories of invasion into the habitat of each of the plants. Thus, it is sug-. gested from the time of meiosis and the distribution pattern of plants in Japan that the relationship between the differentiation time of flower buds and the day length was a very useful approach in under-. standing the distribution route of the plants. 44. Taxonomical treatments As akeady mentioned above, I found that the Japanese Adorns was divided in to three groups based on the chromosome number and morphology. Among them, the 2n = 16 plants found in Hokkaido are surely A. amurensis var. puberula Honda or "Kitami-fukujuso". Furthermore, the photo of Gorovoy and Gurzenkov (1969) of A. amurensis Regel et Radde, as well as Regel's original description and figure, is clearly identical with those of the 2n = 16 Hokkaido plants. A. amurensis Regel et Radde has been known as a diploid plant with 2n = 16. outside Japan (Sokolovskaya, 1966 ; Lee, 1967 ; Shlangena, 1976 ; Probatova & Sokolovskaya, 1981). Compared with the 2n = 16 Hokkaido plants, the 2n = 16 Honshu plants were distributed from northern Honshu to Kyushu in Japan, and also in Korea. This plant looks superficially like A. ramosa Franchet (1894). But this plant should be referable to neither A. amurensis Regel et Radde nor a genuine A, ramosa Franchet, which was described as a new species having many flowers per stem and was collected from N. Japan by U. J. Faurie. The 2n = 16 Honshu plants were different from A. amurensis Regel et Radde ( = A. amurensis Regel et Radde var. puberula Hnda) m that they had many flowers per stem, glabrous leaves, and shorter sepals ; they were separated from A. ramosa Franchet, having shorter sepals and an open cavity in a stem-cross, although many flowers are borne on stem. In addition, the 2n = 16 Honshu plants differ cytologically from a genuine A. ramosa Franchet, which is 2n = 32 in chromosome number. From the facts mentioned above, I think that the 2n = 16 Honshu plant is a new species, Adonis multiflora Nishikawa et Ko. Ito (Photo 17-A). This new species covers Honshu, Kyushu and Korea in its range, and is an endemic plant of the Japanese Korean element.. (12).
(14) Botanical Studies on Adonis amurensis Regel et Radde m Japan (Part 2). Photo 17. A : Adorns multiflora Nishikawa et Ko. Ito. B : A. ramosa Franchet. C : A. amurensis Regel et Radde. Scale indicates 2cm.. The resemblance of the one-flowered 2n = 32 plants to A. amnrensis Regel et Radde has been pointed out: the 2n = 32 plants bear single flower or more per stem, and the habit which bears flowers on a stem was variable through cultivation (Table 24). Moreover, when the 2n = 32 plants have a smgle flower, the secondary stem issued from sheaths bears two or three branches (Photo 17-B), and its appearance is similar to that of A. amurensis (Photo 17-C). Although Franchet might have referred one-flowered plants (with 2n =r 32 ?) to A. amurnesis Regel et Radde, and he gave a new name, A. ramosa Franchet, to. the multi-flowered plants in his studies, Makmo (1901) did not recognize any differences between A. ramosa Franchet and A. amurensis Regel et Radde because the flower numbers were variable. Makino's opinion has been accepted by most authors of current floral manuals. But judging from his enumeration of the specimens and its distributional range, and from my examination of his specimens in MAK, A. amurensis sensu Makmo is also surely the 2n == 32 plants. This fact indicates that Franchet, Makino, and many Japanese taxonomists discussed the classification of A. amurensis Regal et Radde without any considen-ation of its cytology. It is clear that the 2n == 32 plants show some morphological and geographical deviation from genuine A. amurensis Regel et Radde by consistently having many flowers on a stem with multi-branched stem and 32 chromosomes, and that they are endemic to Japan, as far as is known. Furthermore, Gorovoy and Gurzenkov (1969) pointed out that the Japanese Adonis was referable to A. ramosa Franchet, and Kitagawa (1971) accepted their opinion and called A. ramosa "Edauchifukujuso" in Japanese. But for the above-mentioned reasons, those treatments are not correct for the. (13).
(15) Tsunehiko NISHIKAWA. Japanese Adonis. In addition, A. ramosa identified by Gorovoy and Gurzenkov (1969) in Kunashiri Isl. should correspond to 2n = 32 plants.. In conclusion, Japanese Adonis shuold be classified into the following 3 taxa: (1) A. amwensis Regel et Radde in Bull. Soc. Nat. Mosc. 33 (1) : 35, tab. II. fig. 1, 2, a, b.. (1861). Syn: A. amurensis var. puberula Honda in Bot. Mag. Tokyo 52 : 49. (1939). syn. nov. It is characterized by having one flower on a stem and hairy leaves. Japanese name : Kitami-fukujuso (Honda, 1939), Ichige-fukujuso (Kitagawa, 1971), Fukujuso (acut. jap.). Chromosome number : 2n = 16 (Nishikawa and Ito, 1978).. Distribution: NE Hokkaido. (2) A. multiflora Nishikawa et Ko. Ito, sp. nov. in Jour. Jap. Bot. 64 : 50-53 (1989). Planta habitu inter Adonem amurensem Regel et Radde et A. ramosam Franchet. Differt ab A. amurense floribus pluribus, plerumque 3-4, non unifloribus, petalis duplo longioribus quam sepalis, foliis toto glabris ; ab A. ramosa petalis longioribus quam sepalis, caule vacuo, chromosomatibus diploideis vel 2n = 16.. Perennis. Caudbc brevis, crassus. Caulis ca. 3—25 cm altus sub anthesis, elongatus ad 30-40 cm altum post anthesin, teres, vacuus vel non solidus, crassus vel saepe tenuis, divaricatus e base, ramosi 3—4, foliaque glabra. Squamae ad caulis basm subvaginatae, membranaceae, ca. 2—3 cm longae, inferiores semper nudae, superiores interdum folia parvo terminatae. Folia petiolata, supra viridia, infra glaucescentia, temata ; pinnis 2—3 pinnatisectis, laciniis lanceolatis, oblongis, integris, acuminatis vel acutis. Stipulae parvae, 3-5 mm longae. Sepala ovata, oblonga vel oblongo-lanceolata, glaberrima, ca 1/2 plo longiora quam petiolis, 0.5-2 cm longa. Petala cuneato-oblonga vel spathulata vel elliptica, laete flava, antice saepe irregulariter denticulata, distantia, 10—20 mm longa, 3-5 mm lata. Carpella matura globosa, densa pubescentia, sub apice stylo tenui carpello adpresso terminata. Receptaculum oblongo-cyrindricum, 0.5 mm longum, glabrum. Acheniorum capitulum subglobosum vel globosum. Achenia compressa, 4-5 mm longa, 3-3.5 mm lata, in stylum rostriforme porrectum apice hamatum attenuata. Chromosomatum numerum 2n == 16. Pl. IV—V. Pr. V—VI.. Hab. : in partibus silvaticis et in paludis herbosis fluviorum. Typus : Shimodaira, Towada C., Aomori Pref., 24 April 1982, T. Nishikawa. un Herb. SAPT conservatur.. Japanese name: Michinoku-fukujuso. Chromosome number : 2n = 16 (Nishikawa and Ito, 1979). Distribution : Honshu, Kyushu, and Korea. (3) A. ramosa Franchet in Bull. Soc. PhUomath. 6 : 91. (1894). Syn. : A. amurensis auct. jap. p. m. p.. A. amwensis var. yezoensis Kudo in Hokkaido Yakuyo Shokubutsu Zui. p. 44. (1922). A. am.ure.nsis var. uniflorus Makino in Bot. Mag. Tokyo 14 : 97. (1901). A. amurensis var. ramosus (FT.) Makino in Bot. Mag. Tokyo 14: 97. (1901). The plant is characterized by having a multi-flower or one-flower per stem and sparsely hairy leaves underneath.. (14).
(16) Botanical Studies on Adonis amurensis Reget et Radde in Japan (Part 2) Japanese name : Fukujuso, Edauchi-fukujuso (Maldno 1901). Chromosome number : 2n = 32 (Nishikawa and Ito, 1978, 1979). Distribution : Hokkaido, Honshu, Shikoku, endemic.. 45. Cross experiment In some examples of Fi, that is, A. ramosa x A. mnltlflora and A. amurensis x A. multiflora, plants bore one or more flowers per stem (see Table 13). The one-flowered habit of A. amurensis might be supposed to be the same as that of the Fi plants ; but, in fact, this case is not correct as regards details, as shown in Fig. 15. An Fi of A. amnremis x A. multiflora, about half of the total 55 plants were a one-flowered form. However, the leaf arrangement of the Fi is not the same as A. amurensis (Table 25). In A. anuirensis the leaf arrangement is opposite on the primary branch ; in Fi it is alternate, although the stem leaves are alternate in both species. From the facts mentioned above, one-flowered Fi may be nearer to A. multiflora in leaf arrangement than to A. amurensis, which is the mother of F^ A similar phenomenon was observed in an Fi case of A. ramosa x A. multiflora, but in this case it is not surprising that Fi plants were nearer to A. ramosa, because the parent shared. A. amurensis. with 2n=16. with 2n=32. with 2n=32. with 2n=16. A. multlflora. A. ramosa. A. ramosa. Fig. 15. Branching and leaf arrangement. the same characteristics in the leaf arrangement. In the Pi case of A. ramosa x A. amwensis, all of the Pi plants bore one flower per stem and, of course, the leaf arrangement was opposite, very like A. amurensis (Table 25). A, ramosa, the third parent of a triangular cross experiment, was characterized by a multi-flowered form with alternate leaf arrangement on the primary branches and by a 32 chromosome number.. Table 25. No. of flowers on a stem and leaf arrangement of hybrids. Cross A. A. A.. amurensis x ramosa x ramosa x. No. of flowers A. A. A.. multiflora multiflora amurensis. one or more one or more one. Primary branch.. (15). leaf arrangement* alternate alternate opposite.
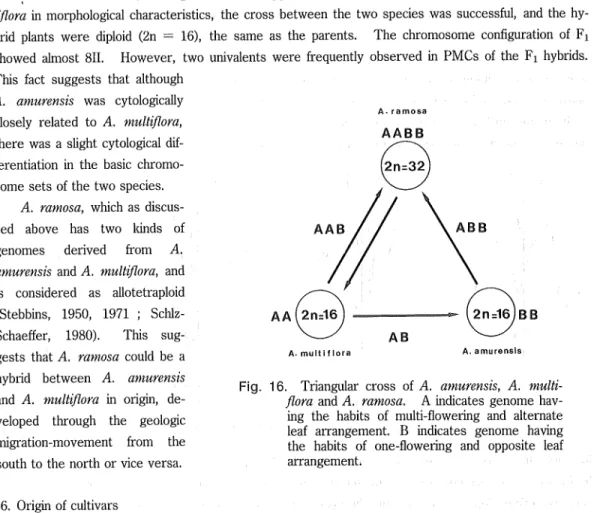
(17) Tsunehiko NISHIKAWA Now, we will return to the number of flowers per stem. A. multiflora has the genome (A) which affects the habit of setting multi-flower and alternate in leaf arrangement in A. ramosa, whereas A. amurensis has the other one, (B), which is responsible for setting one flower and opposite in A. ramosa (Fig. 16). Again it is assumed that A. ramosa has two genomes : genome A which corresponds to the habit of setting multi-flower, and genome B which corresponds to that of setting one-flower. Based on the assumptions mentioned above, it is considered to be reasonable that A. ramosa derived from A. amurensis and A. multiflora (Fig. 16). As mentioned above, although there were apparent differences between A. amurensis and A. multiflora in morphological characteristics, the cross between the two species was successful, and the hybrid plants were diploid (2n = 16), the same as the parents. The chromosome configuration of Fi showed aknost 811. However, two univalents were frequently observed in PMCs of the Fi hybrids.. This fact suggests that although A. amurensis was cytologically A • ramosa. closely related to A. multiflora,. there was a slight cytological differentiation in the basic chromosome sets of the two species. A. ramosa, which as discus-. AAB. sed above has two lands of. ABB. genomes derived from A. amurensis and A. multiflom, and is considered as allotetraploid. (Stebbms, 1950, 1971 ; Schlz-. AA(2n=16. Schaeffer, 1980). This suggests that A. ramosa could be a hybrid between A. amnrensis and A. multiflora in origin, de-. veloped through the geologic. A. multi flora. A. amurensis. Fig. 16. Triangular cross of A. amurensis, A. multiflora and A. ramosa. A indicates genome hav-. ing the habits of multi-flowering and alternate. migration-movement from the. leaf arrangement. B indicates genome having the habits of one-flowering and opposite leaf. south to the north or vice versa.. arrangement.. 46. Origin of cultivars Although it is possible that the triploid arose from the diploid through fertilization of unreduced gamete, the triploids examined were large, stout, and multi-flowering plants, unlike the A. amurensis and A. multiflora, and were very close to A. ramosa m general appearance. The triploids were always multi-flowering as well. Moreover, considering the fact that the 2n = 24 and 40 have seldom been found in wild populations, and that these numbers were counted in seven cultivars among the 24 ones in the present studies, it is plausible that the triploids seem to have been produced by crossmg the tetra-. ploids and A. multiflora (the diploid) either artificially or naturally since the Edo Period. In assuming the diploid parent for the pairing, A. amurensis should be excluded from the parents because of the flowering number and flowering time. From the facts mentioned above, I have tried to explain the ori-. (16).
(18) Botanical Studies on Adonis amurensis Regel et Radde in Japan (Part 2). gin of Adorns cultivars : (1) diploid cultivars descending from A. mnltiflora (cvs. Kotobuki, Hinomotoko, Hinodeko, Tamatebako, Sandanzaki, and Shichifukujin), (2) tetraploid ones descending from A. ramosa (cvs. Garaku, Henge, Benten, Kogiku, Kinsai, Tsumaoregasa, Gosho, Chichibuko, and Sado), and (3) tri- and pentaploid ones derived from the hybrid between A. multiflora and A. ramosa (cvs. /('tsusetsusel, Nadeshiko, Kinsekai, Fukujukai, Kmshi, Shiun, Beninadeshiko, Fukurokuju, and Taiho), Each of the cultivars are mainly characterized by intermediate or transitional morphological characteristics between native species and between cultivars, and by the emergence of new characteristics through long cultivation techniques. (1) Diploid cultivars bear some flowers on a stem ; sepals are obovate in outline, pale to dark green in color, and shorter than the petals ; the fruit is almost globose. (2) Tetraploid cultivars bear a sole flower on a stem ; sepals are tinged with purple to dark purple color. outside and are similar to the petals in length. (3) Triploid and pentaploid cultivars bear some flowers on a stem ; sepals are somewhat shorter than the petals. They are similar to A. ramosa in general appearance, but pollen fertility is lower than that of A. ramosa.. Acknowledgements. I wish to express my cordial thanks to Prof. Dr. K. Ito, Graduate School of Environmental Science, Hokkaido University, for his guidance through the present work. I also wish to thank Prof. Dr. T. Saho, Prof. Dr. K. Tsutsui, Prof. Dr. T. Yoshida, and Associate Prof. Dr. H. Fukuda, Hokkaido University, former Prof. Dr. S. Sato and Prof. Dr. H. Tachibana, Hokkaido University of Education, for their land advice during this study. Thanks are due to the late Prof. Dr. Emer. of Tokyo University Dr. H. Hara, Prof. Emer. of Ybkohama National University Dr. M. Kitagawa, and Associate Prof. Dr. Harada, Hirosaki University. I am grateful to the following persons who kindly assisted me in collectmg samples or sent me materials : Drs. S. Mitsuta and A. Kmoshita, Kyoto University ; Mr. Y. Kormo, Obihiro University of Agriculture and Vetermary Medicine ; Mr. 0. Nakamura, Saitama Museum of Natural History ; Mr. B. Kosuga, Chichibu City ; the late Mr. H. Hinoto, Nishine Town ; Messrs. M.. Mori and T. Takegahara, Sanbongi Agricultural High School; Mr. M. Hashimoto, Hachinohe City ; Mr. S. Mochizuki, Maruko Town ; Mr. T. Maruyama, liyama City ; Mr. F. Ito, lida City; Mr. H. Okuhara, Matsumoto City ; Mr. H. Kobayashi, Nemuro City. My hearty thanks are due to the curators of her-. braria (SAPT, KYO, MAK, TI, TNS, and SHIN). I would like to give special thanks to all the members of Biological Laboratory of Asahikawa College, Hokkaido University of Education, for their generous assistance in this study.. References. Bhat. B. K.. S. K. Bakshi, & M. K. Kaul. 1972. In IOPB chromosome number reports XXXVIII. Taxon 21 : 671-. 684, Briggs, D. B. and S. M. Walters. 1984. Plant variation and evolution. 2nd. ed. 412 pp. Cambridge Univ. Press, London,. (17).
(19) Tsunehiko NlSHIKAWA Darlington, C. D. & A. P. Wylie. 1955. Adonis, in Chromosome atlas of flowering plants. 2nd ed. p. 25. George Alien & Unwin Ltd. London. Fedorov, A. A. ed. 1969. Adonis, in Chromosome numbers of flowering plants, pp. 599-600, Academy of Sciences of the USSR, Moscow. Franchet, A. 1894. Les Adorns vivaces et leur repartition g6ographique. Bull. Soc. PhUom. Paris, Ser. 8, 6: 80-93.. Goldblatt, P. ed. 1981. Adonis, in Index to plant chromosome numbers 1975-1978. Monographs in Systematic Botany 5 : 428. Goldblatt, P. ed. 1984. Adorns, in Index to plant chromosome numbers 1979-1981. Monographs in Systematic. Botany 8: 323. Goldblatt, P. ed. 1985. Adonis, in Index to plant chromosome numbers 1982-1983. Monographs in Systematic. Botany 13: 168. Gorovoy, P. G. & N. N. Gurzenkov. 1969. Adonis ramosa Franch. (Ranunculaceae), a new species for the flora of the USSR and some critical remarks on the Far Eastern species of Adonis L. Bot. Zhum. 54: 139-143. Grant, V. 1981. Plant Speciation. 2nd ed. 563 pp. Columbia Univ. Press, New York. Grant, W. F. ed. 1984. Plant Biosystematics. 674 pp. Academic Press, Toronto. HeB, H. E., E. Landolt, & R. Hirzel. 1970. Adonis, in Flora der Schweiz. Bd. 2, pp. 101-102. Birkhauser Verlag, Basel. Hisauchi, K. 1941. Botanical notes XII. J. Jap. Bot. 1 7 : 473-476. (In Japanese). Honda, M. 1939. Nuntia ad Floram Japoniae. XXXVII. Bot. Mag. Tokyo 52 : 49. Ishikawa, M. 1916. A list of the number of chromosomes. Bot. Mag. Tokyo 30 : 404-448. Izumimoto, Y. 1862. Honzoyosho. (In Japanese). Kitagawa, M. 1971. Fukujuso no Gakumei. Shokubutsu Saishu News. No. 56 : 77. (In Japanese). Kitamura, S. & G. Murata. 1961. Adorns, in Colored illustrations of herbaceous plants of Japan (Choripetalae). p. 234. Hoikusha, Osaka. (In Japanese). Kudo, Y. & T. Suzaki. 1922. Hokkaido Yakuyo Shokubutsu Zui. p. 44. (In Japanese). Kurita, M. 1955. Cytological studies in Ranunculaceae IV. The karyotype analysis in Actaea and some other genera.. Jap. J. Genet. 30 : 124-127. (In Japanese with English summary). Kurita, M. 1957. Chromosome studies in Ranunculaceae VI. Karyotypes of sc< genera. Rep. Biol. Inst. Ehime Univ. No. 3 ; 9-15. Kurita, M. 1963. Chromosome studies in Ranunculaceae. XXI. Karyotypes of Myosnrus and Adorns. Mem. Ehime Univ., H (Sci.) Ser. B. 4: 487-492. Langlet, 0. F. J. 1927. Beitrage zur Zytologie der Ranunculazeen. Svensk Bot. Tidslff. 21 : 1-17. Lee, Y. N. 1967. Chromosome numbers of flowering plants in Korea (1). J. Korea Cult. Res. Inst. 11 : 455-478.. Lewis, W. H. ed. 1979. Polyploidy. 583 pp. Plenum Press. New York. Maluno, T. 1901. Observations on the flora of Japan. Bot. Mag. Tokyo 14: 97.. Makino, T. 1940. Adonis, in An Illustarated Flora of Nippon, with the cultivated and naturalized plants, p. 553. Hokuryukan, Tokyo. (In Japanese). Masuda, K. (Taneldya, K.) 1829. Somoku-ldhin-kagami. (In Japanese). Matsuda, 0. 1960. Hanagoyomi. p. 12. Shakai-shisosha, Tokyo. (In Japanese). Matsudaira, S. 1846. Hyakka-baiyokou. (In Japanese). McMillan, C. 1970. Photoperiod in Xanthium population from Texas and Mexico. Amer. J. Bot. 57 : 881-888. Mehra, P. N., & P. Ramanandan. 1972. Cytology of some W. Hunalayan Ranunculaceae. Cytologia 37 : 281-296. Mizuhara, S., S. Kato & K. Yamamoto. eds. 1981. Nihon-dai-saijilu, Shin-nen, Shokubutsu. pp. 186-187. Kodansha, Tokyo. (In Japanese). Mizuno, M. 1681. Kadankomoku. (In Japanese). Moore, R, J. ed. 1973. Adonis, in Index to plant chromosome numbers 1967-1971. Regnum Veg. 90 : 184. Moore, R. J. ed. 1977. Adonis, in Index to plant chromosome numbers 1973 / 74. Regnum Veg. 96 : 181. Murai, C. 1984. Adorns, in Tsukamoto, Y. ed. Genshoku Kakiengei-daijiten. pp. 725-726. Yokendo, Tokyo. (In. (18).
(20) Botanical Studies on Adonis amwensis Regel et Radde in Japan (Part 2) Japanese). Nihon-fukujuso-no-kai & B. Nakamura. 1978. Fukujuso. 76 pp. Nyusaiensu-sha, Tokyo. (In Japanese).. Nishikawa, T. 1987. Chromosome numbers and geographical distribution of Caltha (Ranunculaceae) in northern Japan. Mem. Natn. Sci. Mus., Tokyo, No. 20 : 91-101. (In Japanese with English summary). Nishikawa, T. 1989. A new species of Adonis m Japan. J. Jap. Bot. 64 : 50-53. (In Japanese with English summary). Nishikawa, T. & Ko. Ito. 1978. New chromosome numbers of Adorns aimirensis Regel et Radde of Hokkaido. J. Jap. Bot. 53 : 33-43. Nishikawa, T. & Ko. Ito. 1979. The chromosome numbers of Adonis amwensis Regel et Radde (sensu lato) of northern Honshu. J. Jap. Bot. 54: 353-362. Nishikawa, T. & Ko. Ito. 1985. An experimental hybridization of Adorns amurensis group and morphological comparisons of cultivars. J. Jap. Bot. 60; 79-89.. Ohwi, J. 1975. Adonis, in Flora of Japan CNew edition), p. 614. Shibundo, Tokyo. (In Japanese). Ohwi, J. & M. Kitagawa. 1983. Adorns, in New Flora of Japan, pp. 697-698. Shibundo, Tokyo. (In Japanese).. Oka, H. I. 1958. Photoperiodic adaptation to latitude in rice varieties. Phyton 11 : 153-160. Okuhara, H. 1985. Some new or noteworthy plants found in Nagano Prefecture (4). Bull. Bot. Soc. Nagano. No. 18 : 3. (In Japanese).. Panje, R. R. & K. Srinivasan. 1959. Studies in Saccharwn spontaneum. The flowering behavior of latitudinally displaced populations. Bot Gaz. 1 20: 193-202. Probatova, N. S. & A. P. Sokolovskaya. 1981. Kariologicheskoe issledovanie sosudistykh rastenij ostrovov Dal'nevostocnogo gosudarstvennogo morskogo sapovednika. Sb. Cvetkaye Rastenija Ostrovov Dalnevostochnogo Morskogo Sapovednika. pp. 92—114. (In Russian). Ray, P. M. & W. E. Alexander. 1966. Photoperiodic adaptation to latitude in Xanfhium strumaritim. Amer. J. Bot.. 53: 806-816. Regel, E. 1861. Plantae Raddeanae. pp. 32-38. t. 2. f. 1, 2, a, b. Schulz-Schaeffer, J. 1980. Cytogenetics, Plant, Animals, Humans. 446 pp. Spring-Verlag, New York. Shimizu, T. 1982. Adorns, in Satake, Y., J. Ohwi, S. Kitamura, S. Watari & T. Tominari. eds., Wild Howers of. Japan, Herbaceous plants (including dwarf subshrubs). p. 75. Heibonsha, Tokyo. (In Japanese). Shimizu, M. 1955. Fukujuso-no-Hanashi. Engei-techo 5 ; 3-5. (In Japanese). Shlangena, Z. E. 1976. The chromosome numbers of some Adorns L. species in the USSR (Ranunculaceae). Bot.. Zhum. 61 : 1603-1608. (In Russian). Sokolovskaya, A, P. 1966. Geografichesko rasprostranenie poliploidnykh vidov rasteniy. (Issledovanie flory Primors-. kogo kraya). Vestnik Leningr. Univ. Ser. 3, Biol. 1966 : 92-106. (In Russian with English summary). Stebbins, G. L. 1950. Variation and Evolution in Plants. 643 pp. Columbia Univ. Press, New York. Stebbins, G. L. 1971. Chromosomal evolution in higher plants. 216 pp. Edward Arnold Ltd, London. Sugiura, T. 1931. A list of chromosome numbers in angiospermous plants. Bot. Mag. Tokyo 45 : 353-355. Sugiura, T. 1936. Studies on the chromosome numbers in higher plants, with special reference to cytokinesis I.. Cytologia, Fuji; Jub. Vol. : 845-849. Takahashi, R. & S. Yasuda. 1970. Genetics of earliness and growth habit in barley. Barley Genet. II: 388-408. Takamine, N. 1916. Uber die ruhenden und die prasynaptischen Phasen der Redukutionsteilung. Bot. Mag. Tokyo. 30: 293-303, Tamura, M. 1962. Petiolar anatomy in the Ranunculaceae. I. Structure of the proper part of petioles. Sci. Rep. Osaka Univ. 11 : 19-47. Tateoka, T. 1973. A taxonomic study of the Poa macrocalyx complex, with particular reference to the populations in eastern Hokkaido. Bot. Mag. Tokyo 86: 213-228.. Tateoka, T. 1974. Phytogeographical studies of Calamagrostis saclialmensis (Gramineae). II. Origin and dispersal. Bot. Mag. Tokyo 87 : 149-164. Tateoka, T. 1983. Plant Speciation and Taxonomy. 269 pp. Yokendo, Tokyo. (In Japanese).. (19).
(21) Tsunehiko NlSHIKAWA Tokyo Astronomical Observatory ed. 1986. Rikanenpyo (Science Calendar). 1007 pp. Maruzen, Tokyo. (In Japanese).. Wang, W. & S. Wang. 1980. Adonis, in Flora Reipublicae Popularis Sinicae. 28 : 246-255. (In Chinese). Yoshimura, K. & G. Sakurai. 1968. Adonis, in Ishii, R. & Y. Inoue. eds. Encyclopedia of Horticulture. 1 : 35-37.. Seibundoshinkosha, Tokyo. (In Japanese).. Appendix 1. Adonis amurensis Regel et Radde Nom. Jap. Kitami-fukujuso, Ichige-fukujuso.. Distribution. Japan (NE. Hokkaido), Saldialin, E. Siberia, Korea, and Manchuria. Specunens examined.. Hokkaido. Soya Prov. : Utanobori (M. Yamanoi, 29 Apr. 1972 TNS). Abashiri Prov. : Tokoromura (H. Iwamoto, 20 Apr. 1936 SAPT; H. Iwamoto, 16 Apr. 1936 TNS ; H. Iwamoto Herb. No. 86, 19 Apr. 1936 TI-Holotype of var. ptiberula ; H. Iwamoto Herb. No. 86, 20 Apr. 1936 TI-Holotype of var. pubemla). Kushko Prov. : Tenneru,. Kushiro-cho. (K. Takita, No. Ill, 19 Apr. 1981 KYO ; K. TaMta No. 124, 5 May 1981 KYO). Shibecha (M. Tatewaki, 20 Jun. 1920 SAPT), Chanai (M. Tatewaki, 29 May 1936 SAPT). Kamioboro (M. Tatewaki, 25 May 1936). Nemuro Prov. : Nemuro (D. Hoshi, 30 Apr. 1911). Bettoga (M. Tatewaki, 18 Jun. 1936). Adonis multiflora Nishikawa et Ko. Ito, sp. nov. Nom. Jap. Michinoku-fukujuso. Distribution. Japan (Honshu and Kyushu) and Korea. Specunens examined.. Honshu. Tohoku Distr. Aomori Pref. : Hachinohe (R. Yamazald, May 1902 MAK). Shiwa. Towada (U. Narita, 15 Apr. 1946 TI). Hirota, Rikuzentakada (G. Toba, 14 Apr. 1905 MAK). Yamagata Pref. : Kaminoyama (T. Takano, 12 Apr. 1970 TNS). Chikayama-fukazawa (T. Nagasawa, Apr. 1882 TNS). Kanto Distr. Kanagawa Pref. : Yokohama (?, 15 Mar. ? MAK). Yokohama-kawashima (K. Hisauchi, 25 Mar. 1916 TI). Chubu Distr. Nagano Pref. ; Otaki (M. Kawakatsu, Apr. 1955 KYO). Kawachi, Achi (P. Ito, 11 Apr. 1982 SHIN). Kurimoto, Kisofukushima (H. Okuhara, 27 Apr. 1982 SHIN). Near Edatachi-toge, China (M. Togashi, 13 May 1972 TI). Kamieda 0. Kasahara, May 1939 TI). Hokuriku Distr. Konehashi, Katsuyama (S. Watanabe, 3 May 1974 KYO). Kyushu. Kumamoto Pref. : Naidaijin-yama (Y. Shimada, 11 Mar. 1954 TNS). Mt. Noke-eboshiyama, Itsuld (K. Maebara, 30 Mar. 1916 KYO ; N. Kurosaki, 20 Apr. 1975 KYO ; ? 31 Mar. 1916 MAK). Araoyama (H. Kamitsuma, 1 Apr. 1908 MAK). Mt. Iwaudo-yama, Izumi (M. Hotta, 3 May 1963 KYO). Oita Pref. : Harano Minamiyama (K. Ikebe, 30 Mar. 1926 KYO) Korea. Kangnug, Kyonggi-Do (0. Yongsok, 29 Apr. 1949). Nankanzan (T. Mori, 8 Jun. 1921 TI). Koryo (R.. Takagi, 12 Apr. 1929 TI). Adonis ramosa Franchet Nom. Jap. Edauchi-fukujuso, Fukujuso. Distribution. Japan (Hokkaido, Honshu, and Shikoku) Specknens examined. Hokkaido. Kamikawa Prov. : Asahiliawa (G. Katsuge, MAK). Arashiyama, near Asahikawa (H. Iwamoto Herb. No. 86-b, 2 Jun. 1937 TI). Sankakuyama, Higashitakasumura (H. Iwamoto Herb. No. 86-c, 15 Jun. 1937 TI). Sorachi. Prov. : Iwamisawa (N. Kinashi, No. 49, May 1902 KYO). IshUrari Prov. : Noppro 0. Doi, 22 May 1932 KYO). Maruyama, Sapporo 0'. Tokubuchi, 22 May 1891 SAPT ; H. Takeda, 3 May 1908 MAK ; H. Takeda, 17 May 1908 MAK ; ? Apr. 1905, TNS). Moiwayama, Sapporo (K. Inagaki, 28 May 1928 SAPT ; Fujita et al. Apr. 1928 SAPT; H. Takeda, 9 May 1908 MAK). Sapporo (K. Miyabe, Apr. 1882 SAPT ; Y. Tokubuchi, 26 May MAK ; Y. Tokubuchi, Jun. 1891 SAPT ; K. Miyabe, Apr. 1878 TI ; H. Takeda, 5 May 1909 TI; May 10 1885 MAK ; T. Hori, 10. (20).
(22) Botanical Studies on Adonis amurensis Regel et Radde in Japan (Part 2) May 1943 TI). Kobetsuzawa-bankei, Sappro CM. Mizushima, 29 Apr. 1944 TI). Juuniken-nosawa, Sapporo (M, Mizushima, 29 Apr. 1944 TI). Mt.Teine (H. Yanagisawa, 2 May 1916 SAPT). Near Sankakuyama, Sappro (M. Tatewaki, 30 Apr. 1920 SAPT). Chitose River (M. Tatewalu, 16 May 1929 SAPT). Chitose-fukajo, Chitose (M. Tatewaki, 11 May 1940 SAPT). Abashiri Prov. : Shiitokoro, Oketo (M. Tatewald, 2 May 1950 SAPT). Tokachi Prov. : Urahoro (K. Hori, 30 May 1896 SAPT). Hagigaoka, upper Otofuke (M. Tatewaki, 2 May 1950 SAPT). Kushiro Prov. : Pirikaneppu, Akan (M. Tatewaki, 11 May 1950 SAPT). A lake side of Akan, Akan (S. Sansho? 12 May 1975 KYO). Iburi Prov. : Tomakomai Experimental Forest M. Tatewaki, 8 May 1929 SAPT). Muroran (K. Miyabe, 27 Apr. 1894 SAPT). Sokuryo-zan (D. Hoshi, May 10 1940 SAPT). Iburi (K. Nakano, 1895 TNS). Hidaka Prov. : Mt. Apoi (M. Tatewaki, Jun. 1927 SAPT, Apr. 28 1928 SAPT). Samani (M. Tatewaki, May 21 1927 SAPT). Saruru (M. Tatewaki, 19 May 1951 SAPT). Hidakamonbetsu (M. Tohyama, 5 May 1966 SAPT). Maruyama, Shoya (V. Tateishi, 24 Apr. 1975 TI). Oshima Prov. : Shirikishinai (M. Hara, 19 Apr. 1980 KYO). Motomachi, Hakodate (Takeda, 7 May 1905 TI). Hakodate (F. B. Greatrex, 27 Apr. 1894 SAPT). Vicinity of Hakodate (F. B. Greatrex, May 1916 SAPT). Hakodate-hiU (I. Yamamoto, 19 Apr. 1924 SAPT). Hiyama Prov. : Gamushi (M. Tatewaki et at., 23 May. 1955 SAPT). Honshu. Tohoku Distr. Aomori Pref. : Asamushi, Aomori (Faurie, Apr. 1902 KYO ; 1897 KYO). Gonohe (Faurie, 10 May 1886 KYO). Natsudomari, Hiranai (K. Hosoi, 16 Apr. 1950 MAK, 27 Mar. 1953 MAK). Akita Pref. : Ogadaijima (H. Muramatsu, 26 Mar. 1930). Hatamura, Oga Peninsula (S. Muramatsu, 20 Mar. 1930 TI). Iwate Pref. Akka, Iwaizumi (E. Ito & Togashi, 25-26 May 1967 TNS). Orikabe-mura (Apr. 1900 MAK). Yamagata Pref. : Numasawa (S. Okuyama, 19 Apr. 1946 MAK). Yamadera, Yamagata (K. Takahashi, 5 Apr. 1967 MAK). Fukushima Pref. : Monokura, Soma (E. Haga, 25 Mar. 1973 TI). Kanto Distr. Tochigi Pref. : Nasu, Akuto^ (S. Kitamura, 3. Mar. 1971 KYO). Tokyo Metro. : Musashi-asakawa (K. Hisauchi, 16 Mar. 1941 TNS). Takaosan, Asakawa (M. Mizushima, 19 Mar. 1967 MAK). Saitama Pref. : Sawaguchi, Yoshda (M. Togashi, 5 Mar. 1972 TI). Urashima, Ryogami (T. Yamazalu, 6 Mar. 1959 TI). Chubu Distr. Gifu Pref. : Karita-yama, Obuse (Y. Matsuda, 2 May 1982 SHIN). Karasawagawa, Hata (Y. Matsuda, 30 Apr. 1978 SHIN ; F. Yokouchi, 30 Apr. 1978 SHIN). Shirakabaguchi, Matsumoto (Sakakibara, et al. 29 Apr. 1970 SHIN). Hakamakoshi (T. Shimizu, 8 May 1966 SHIN). Hokuriku Distr. Toyama Pref. : Maruyama, Iguchi (N. Satomi & M. Togashi, 14 Apr. 1958 SHIN & TNS ; N. Satomi & M. Togashi, 14 Apr. 1956 TI; S. Satomi, 5 Apr. 1954 TNS). Sanmenshirataki, Asahi (T. Ucharawa, 10 Apr. 1960 TNS). Yahiko (I. Ito, 4 Apr. 1951 TNS). Nyukawa-Tadaramine, Sado (S. Okuyama, 13 May 1954 TNS). Kinki Distr. Osaka Pref. : Nakashukuno, Nose (K. Seto, 28 Mar. 1968 KYO). Kyoto Pref. : Midono, Yasaka (G. Murata, 25 Apr. 1970 TI, TNS & KYO). Ohara, Kyoto (0. Suzuka & S. Kudo, 9 Mar. 1967 KYO ; 0. Suzuka, Mar. 1935 KYO). Shiga Pref. : Ryozenyama, Maebara (G. Murata 16 May 1982 KYO ; H. Yamarnoto, 19 Apr. 1931 KYO). Oike-dake, Taga (T. Kodama, 29 Apr. 1963 TNS ; M. Hutoh, 29 Apr. 1963 TNS ; G. Murata & N. Fukuoka 29 Apr. 1963 KYO, TI & MAK ; M. Togashi, 30 Mar. 1967 TI; G. Murata, 14 May 1970 KYO & TNS). Mie Pref: Fujiwara-dake (H. Hara, 5 May 1956 TI; T. Shimizu, 1 May 1955 KYO ; K. Inami, Mar. 1959 TNS ; S. Okuyama, 4 Apr. 1953 TNS ; Z. Tashiro, 3 Apr. 1937 KYO ; Y. Sonoda, 15 May 1937 KYO ; H. Takahashi et al. 4 Apr. 1978 KYO ; G. Murata, 1 May 1955 TI, 4 May 1969 KYO, 14 Apr. 1970 MAK, SHIN, TNS & KYO ; 9 May 1971 KYO & SHIN ; G. Murata & H. Nishimura, 30 Mar. 1968 KYO & TI; K. Yato, 3 Apr. 1938 TI; N. Satomi 8 Apr. 1962). Shikoku. Shikoku Distr. Ehime Pref. : Tanbara (M. Kurose, 25 Mar. 1937 MAK). Tokushima Pref. : Jinryo (N. Satomi, 25 Mar. 1952 TNS & TI; T. Yamanaka, 10 May 1957 KYO ; Satomi, 8 Apr. 1962 MAK). Tsurugi-san (S. Fukushima, 24 Mar. 1960 TNS). Appendix 2 Locality names English • • • Japanese. jtrti^-IE. (21).
(23) Tsunehiko NlSHIKAWA Hokkaido Soya Prov.. ^^ST. Ofuntarumanai, Utanobori. ^.sw^w'y^^'i-^. Okajuna, Esashi. -K^BTW. w^±w. Omisaki, Wakkanai Soya-misaki, Wakkanai Kamikawa Prov.. ^fW^SW. ±JIIS/r. ASN-4'^. Chuo, Shimukappu. wiir?yffipu ABjnmxwn. Han-menzan, near Asahikawa. Higashi-asahikawa, Asahikawa Higashi-furen, Furen Ikutora, Mmamifurano Kamifurano. ±g?±±glJ WllrWJ^i. Kami-shibetsu. Shibetsu. Kamuikotan, Asahikawa Kanayama, Minami-furano. ±JIIBTiSE&. Koshiji, Kamikawa. ^'SrtiB?!. Nisshin, Nayoro Noei, Minami-furano. IW. Okikineushi, Biei. jww^ms. Pon-ubun, Asahikawa Seinan-mappu, Toma Sorachi Prov.. Kuruki-rindo, Yubari Minami-yoshino, Sunagawa. Mogami, Kurisawa. S?U» CT. S^S/T f'mi^'iS^W WWf^faW !?JW^± ^rti'y.i'^ajiSWS. Near Shuuparo-ko, Yubari. ^Sf^^R. Nishikizawa, Yubari. ^grms ^ ?j? ^grm^gi. Numanoswa, Yubari. Okukashim, Yubari. ^^TOMuU. Onikubi-yama, Yubari. ^gTp^-y^y®^. Osoushi-no-sawa, Yubari Sakuraoka, Kuriyama Shibun, Iwamizawa. Shirakin, Yubari Abashm Prov.. gsW^fi. gjirRTl?^ ^5grpe^ if?£;T. wgw^a s E. Akisato-3-ku, Okoppe Baro, Yubetsu. feSRW-?^. Chirai, Saroma. i^gsm^-. Gifu.Tokoro. Hokko, Okoppe. iJISiJlilPtsRW ^BBKBTSF1. Hon-mazawa, Yubetsu Iwato, Ikutawara. Kaigancho, Abashiri Kamimo, Nishiokoppe. jiS±nrr±tlA®. Kami-sakkm, Takinoue. i.m'swi 4 y 2 E. 4-2, Kamiyubetsu. (22).
(24) Botanical Studies on Adonis amurensis Regel et Radde in Japan (Part 2). IS'JBTfrtStt. Kerochi, Yubetsu. ssmw^. Kowa, Rubeshibe. i-im^ww. Maruseppu. ^WiWWi. Muri, Maruseppu Nakaokoppe, Nishiokoppe. •;?im*imwffi. Near Hon-mazawa, Yubetsu. fr^N-SE. Nishiku, Shirataki Nishi-okoppe.. ?rt^W. Notoro-misaki, Abashiri Saroma. ffl^rf]±[S) feSFsW. Saroma-ko, Saroma. fe@rBW-9-a-7M. Omagari, Abashiri. igijBrs^. Shibushi, Yubetsu Shoei, Rubeshibe. &SW±fS. issmw^^; it±B?g^T. Taisei, Saroma. Takmoyu, Rubeshibe Takinoshita, Takinoue Tomioka, Engaru. NiglJBTrt)^ H?Tti3PJ^. Tsubetsu Ubaranai, Abashu-i. MSW?N^. Utsutsu, Monbetsu. AJ|Ttl^t&. Wakamatsu. Kitami. ^b^^^^-i i?TWA^Ii sm^ff. Wakamatsu-suki-jo, Kitami. Yobito-hanto, Abashiri Kushiro Prov.. Beppo, Kushiro. ^.fwwmw. Kussharo-ko, Teshikaga. ^S?4^SiJ. Naka-onbetsu, Onbetsu Shimo-setsuri, Tsurui. ^?-@. Teshikaga. I^T. Nemuro Prov.. smwww. Nakanishibetsu, Bekkai. tg^rWftEB. Nishiwada, Nemuro Nemuro. 'IS^rpjfH® a^?. Tobai, Nemuro Hidaka Prov.. mm^. Heiu, Samani. Hidaka-suki-jo, Hidaka. ^^njHA^. Hyakunin-hama, Erimo. x.t3^BT§j. Meguro, Erimo. Nioi, Biratori Nishi-horobetsu, Urakawa. WWIBSWiiim yrsw±m!-. Near Hidaka-bokujo, Urakawa Okaribe, Niikappu. ^OfeBTCT +KIT ^{H^i. Shoya, Erimo Tokachi Prov. Aikoku, Obihiro. (23).
(25) Tsunehiko NISHIKAWA. s-^w&w. Ashyoro-ko, Ashyoro. mwrrt. Atsunai, Urahoro. nsijinmN lA^BTTtl^ ^BTSig. Furumai, Makubetsu. Hiroo Ishizaka.Taiki Kami-atsunai, Urahoro Katsuhira, Urahoro. mwrrjii±. Kawakami, Urahoro. ^rtiJIIE. Kawanishi, Obihu-o. Kisen, Shintoku. ^nrBXU-J. Kunimiyama, Memuro. ffiwmuj. Maruyama, Urahoro. Mikage, Shimizu Minami-9-go, Toyokoro. 44LFW. Nakasatsunai. 9^. ?fflrtbBT?£ ?f8Wrnajws. Near Kitamachi, Urahoro Near Maruyama, Urahoro. mww. Nukanai, Makubetsu. ±^wss S^OTI^. Oda, Taiki Rawan, Ashoro Rushin, Urahoro Rushin-onsen, Urahoro. mm£fi 44LFW^-?&E. Sakae, Makubetsu Shinrin-koen, Nakasatsunai. »9BTm^. Shintoku. mmw ^ISrt^iE. Shinwa, Makubetsu Taisho, Obihiro Taitomi, Urahoro. ifcXBr+%. Tofutsu, Toyokoro Urahoro. Ishikari Prov. Shikotsu-ko, Chitose. S^ff. :f^rt?^®ffl nsimI?»-?&E. Shinrin-koen, Ebetsu Shokubutsuen, Sapporo. Iburi Prov. Raiba, Noboribetsu Rebunge, Toyoura. ss.m^ff lgljrt?5f^ jl?il?[irfflj^. inffwuu. Sokuryozan, Muroran Takaoka, Astuma. iffimTiss?. Tomiura, Toyoura Toyoura. Shiribeshi Prov. Shirozumi, Kuromatsunai Hiyama Prov.. ISffiBTrPN. f^3?. m^wQix. It^sjf. ±?^.W. Kaitorima, Taisei. mxgwri^uj. Kaminoyama, Assabu. i.^sw'gs. Miyakoshi, Kaminokuni. (24).
(26) Botanical Studies on Adonis amwensis Regel et Radde in Japan (Part 2). 4-^illT4]S. Nakasato, Imagane. ^K\. Okushiri Isl. Oshima Prov.. ^BTtflW ^BTCtlj. Nakayama-toge, Ono Niyama, Ono. Honshu Aomori Pref.. *iW ffinwsnin +?BTpitl. Ainai-gawa, Soma. Fujishima, Towada Herai, Shingo. +S]BTp»Nrt. Horanai, Towada. Ichinowatari, Hirosaki. ^BUTp— ^ ?^. Kirita, Towada. -NtlBrtrfSEB -bFBTNl^W. Nantozawa, Shichinohe. !ew±?m. Omagoshi, Iwasald. -NOBBrf3T;iP. Shimodaira, Towada. -N{]Bar?ft]. Shiwa, Towada. AFTfiSM. Tanesashi, Hachinohe Yakeyama, Towadako Iwate Pref.. 5FJHBT4"1'^. Chusonji, Hiraizumi Obuke. Nishine Miyagi Pref.. ».l-?Tp4'^. Nakayamadaira, Naruko. ^3.^. Saitama Pref.. w^&w. Hekiitazawa, Ryogami. mWQ^±!S-^R. Hinataooya-Kurasawa, Ryogami Isama, Yoshida. ^GBfWa sSyWm^R ft5<:Ttl?t^. Sawaguchi, Yoshida Takikoshizawa, Ryogami Urayama, Chichibu. m. Nagano Pref.. ^W^^S^f. China, Ankokuji. ?(fflfflTjwrF[ij Rsti-ffift. Eboshiyama, Sanada. Gowa, Achi. ^^^a^BTM^. Harano, Kisofukushima. vamws^f ^lllT^hgN!. Hofukuji, Shiga Hokuryuike, liyama. @;S^±*A. Kamihoniri, Takeishi. W§-^'? ±BrMit!lS[ij. Osaka, Okaya Tokko-san, Ueda. t». Mie Pref. Fujiwara-dake, Fujiwara. (25).
(27)
図



+4
関連したドキュメント
The JSC strives to promote sport and improve physical health of school children through the following activities; management of sport facilities such as the New
〃o''7,-種のみ’であり、‘分類に大きな問題の無い,グループとして見なされてきた二と力判った。しかし,半
[r]
INTERNATIONAL MASTER’S PROGRAM IN JAPANESE HUMANITIES INTERNATIONAL DOCTORATE IN JAPANESE HUMANITIES
JAPAN STUDIES PROGRAMS IN ENGLISH AT THE GRADUATE SCHOOL OF HUMANITIES THE INTERNATIONAL MASTER’S PROGRAM (IMAP) IN JAPANESE HUMANITIES AND THE INTERNATIONAL DOCTORATE (IDOC)
Introduction to Japanese Literature ② Introduction to Japanese Culture ② Changing Images of Women② Contemporary Korean Studies B ② The Chinese in Modern Japan ②
関西学院大学産業研究所×日本貿易振興機構(JETRO)×産経新聞
