熊本大学学位論文
新興薬剤耐性菌が産生するメタロ -β- ラクタマーゼ の加水分解機構の解明と非可逆的阻害剤の開発
2005 山口 佳宏
Probing the hydrolysis mechanism of metallo-β-lactamases and development of their irreversible inhibitors
2005
Yoshihiro Yamaguchi
Probing the mechanism of hydrolysis of metallo-β-lactamases and development of irreversible inhibitors
Yoshihiro Yamaguchi Metallo-β-lactamases (MBLs) are dinuclear Zn(II) metalloenzymes that catalyze the efficient hydrolysis of a wide range of β-lactam antibiotics including carbapenems and extended-spectrum cephalosporins. The emergence of appearance of these antibiotic-resistant bacteria raises a considerable public health concern and awareness regarding the proliferation and treatment of infectious diseases. More than twenty MBLs have been identified in clinical isolates of major gram-negative pathogens. Furthermore, MBLs show little or no susceptibility to clinically used β-lactamase inhibitors such as clavulanic acid, tazobactam, and sulbactam. Although there have been reported several of compounds that inhibit MBLs, at present no clinically useful inhibitors appear to be available.
One of the causes for this is the rate of MBL dissemination and diversity, which prevents the development of ΜΒL inhibitors. These facts clearly highlight the urgency and importance of improving our understanding of MBLs to design either new β-lactam antibiotics with improved activity against bacteria or inhibitors that are capable of extending the utility and lifetime of currently used antibiotics.
Therefore, the aims of this study were to clarify the common hydrolysis mechanism of MBLs using kinetics and X-ray crystallographic studies and to develop mechanism-based irreversible inhibitors for all MBLs and agents for detecting MBL-producing strains with high sensitivity and selectivity.
The new findings obtained in this study are summarized as follows.
1. Knowledge of the pH dependency in hydrolyzing enzymes is indispensable to elucidate
the reaction mechanism because the active sites generally contain acidic or basic
groups and one protonated form of the acid and base is catalytically active. In
pH-jump experiments, IMP-1, a member of the MBL family and named from a
Serratia marcescens isolate collected in Japan in 1991, was inactivated in an acidic
medium by the dissociation of Zn(II) from the holo-enzyme, where ligands
coordinating to Zn(II) ion(s) may be protonated. The pH dependence for the IMP-1
hydrolysis in the presence of large excess of Zn(II) ion revealed that no functional
group having a pK
abetween pH 5 and 9 was present. These results suggest that the
pK
avalue of a bridging OH
2to two Zn(II) ions (Zn1-OH
2-Zn2), which acts as a
nucleophile to the carbonyl carbon on the β-lactam ring, is lower than 5. It is thought
that the positive charges of the two Zn(II) ions are the main cause for lowering the pK
afor water as a nucleophile and assist in the deprotonation of the bridging OH
2.
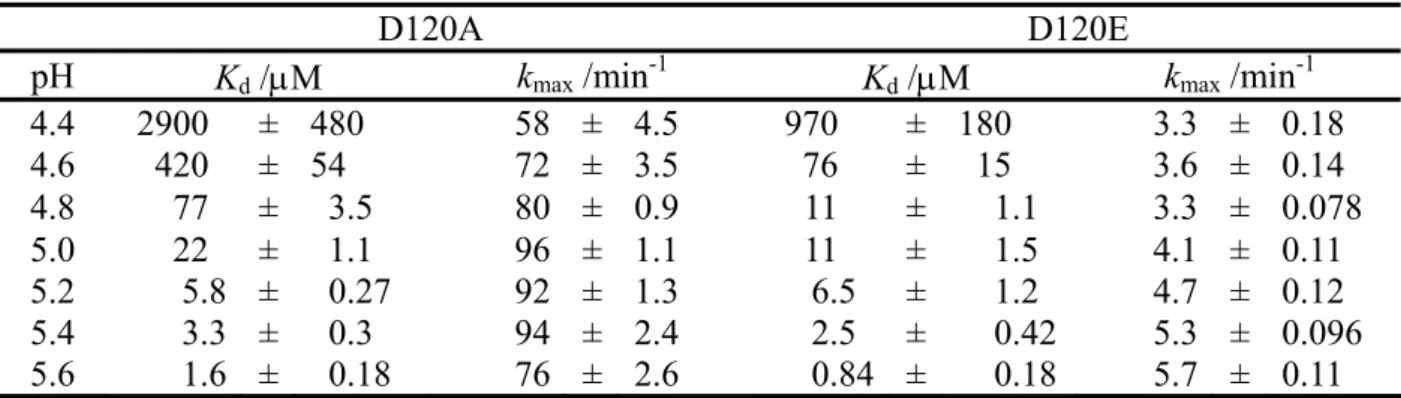
2. To probe the role of the well conserved Asp120 in the active site of IMP-1, site-directed mutagenesis was conducted. The two Asp120-mutants, D120(81)A and D120(81)E, in which asparatic acid was replaced by alanine and glutamic acid, were characterized using CD, metal analyses, steady-state kinetics, and X-ray crystallography. In the steady state kinetic experiments, D120(81)A and D120(81)E showed a remarkable reduction in activity, i.e. a decrease in k
catby 10
2-10
4fold and an increase in K
mof several fold to 10
2-fold. These results suggest that Asp120 in IMP-1 plays important roles in catalysis and substrate recognition. Furthermore, from pre-steady-state kinetic experiments, an anionic intermediate of nitrocefin having an absorption maximum around 650 nm was not observed for the IMP-1 mutants, suggesting that Asp120 in IMP-1 is not a proton donor to the anionic intermediate of nitrocefin. The pH dependence of the hydrolysis for the IMP-1 mutants indicated that Asp120 is not a factor in decreasing the pK
abelow 5 for the water bridging two Zn(II) ions. The three-dimensional structures of D120(81)E and Glu120(81)E were determined at a resolution of 2.0 and 3.0 Å, respectively. From the X-ray crystallography results, the structure of D120(81)E (the oxygen atom of Glu120(81)OE1 is presumably not coordinated to the bridging OH
2or OH
-), the oxygen atom of the carboxylate of Asp120 is essential for coordination to the bridging OH
2or OH
-by a hydrogen bond to achieve catalytic activity. Thus, the critical role of Asp120 is to orientate OH
2or OH
-, which acts as a nucleophile to the carbonyl carbon on the β-lactam ring.
3. To develop effective inhibitors against IMP-1, two novel MBL inhibitors,
pentafluorophenyl 3-mercaptopropionate (PFMP) and 3-(3-mercaptopropionylsulfanyl)propionic acid pentafluorophenyl ester (MPAP) were
synthesized. These compounds have a thiol group for the coordination to Zn(II)
ion(s), and an activated ester capable of interacting with the Cys 221 or Lys224 residue
in the active site as a nucleophile. From gel-filtration and dialysis experiments, the
treatment of IMP-1 with each inhibitor resulted in a nearly 100% inhibition, indicating
that both inhibitors inhibited IMP-1 very rapidly and irreversibly. The MALDI-TOF
mass spectra of IMP-1 treated with each inhibitor showed an increase in mass,
corresponding to the sum of the mass of the acyl moiety that is removed from the
pentafluorophenol group from the inhibitor and of IMP-1. Moreover, to identify the
site of amino acid attachment and to determine the three-dimensional structure, the
X-ray crystallography of IMP-1 complexed with MPAP was carried out at a resolution of 2.6 Å. X-ray crystallography revealed that the thiol group bridges between the two Zn(II) ions in the active site and the terminal carbonyl group in the acyl moiety covalently bonds to the side chain N
ζatom of Lys224 to form an amide bond.
4. To devise a fluorometric method for detecting MBL-producing strains rapidly and selectively, a novel fluorescence detecting reagent, N-[2-(5-dimethylamino-naphthalene-1-sulfonylamino)ethyl]-3-mercaptopropionamide
(DansylC2SH) was synthesized. This compound contains a thiol group as a Zn(II) ligand and a dansyl group as a fluorophore and both groups are linked via ethylendiamine as a spacer. To examine the ability of the fluorescent detecting agent for IMP-1, the fluorescence emission spectra of DansylC2SH in the absence or the presence of IMP-1 were measured. When a IMP-1 buffered solution was added to a DansylC2SH solution, fluorescence enhancement was observed, indicating that DansylC2SH binds in the active site, and interacts with Trp64(28) near the active site.
As mentioned above, these findings shed new light on our understanding of the common hydrolysis reaction mechanism for, not only MBLs, but also binuclear Zn(II) hydrolases that have been discovered recently. The inhibition modes of the inhibitors for IMP-1 designed and synthesized in this study were clarified to be irreversible using kinetics, MALDI-TOF mass, and X-ray crystallographic studies. These inhibitors interacted with IMP-1 to form a covalent bond between the acyl group in the inhibitor and the side chain N
ζatom of Lys224, leading to irreversible inhibition. Thus, the strategy for the design of irreversible inhibitors and X-ray crystal results in this study would help in the design of mechanism-based inhibitors that target, not only all MBLs, but also on other metalloenzymes such as metallo matrix proteases.
Moreover, it is expected that the fluorescence detecting reagent, DansylC2SH, represents a
lead compound for the further development of detecting agents and specific inhibitors with a
high affinity and selectivity for all MBLs.
本論文で使用した略号一覧表
MBL Metallo-β-lactamase
MALDI-TOF MS Matrix assisted laser desorption ionization-time of flight mass spectrometer
MOPS 3-Morpholinopropanesulfonic acid MES 2-Morpholinoethanesulfonic acid
WT Wild-type
CD Circular dichroism
DMSO Dimethyl sulfoxide
Tris Tris(hydroxymethyl)aminomethane
PDB Protein Data Bank
MW Molecular weight
SDA-PAGE Sodium dodecyl sulfate-polyachrylamide gelelectrophoresis PFMP Pentafluorophenyl 3-mercaptopropionate
MPAP 3-(3-Mercaptopropionylsulfanyl) propionic acid pentafluorophenyl ester
TFA Trifluoroacetic acid
3-MPA 3-Mercapropropionic acid
Hepes 2-[4-(2-Hydroxyethyl)-1-piperazinyl]ethanesulfonic acid PCR Polymerase chain reaction
NCCLS National committee for Clinical Laboratory Standards DansylC2SH N-[2-(5-dimethylamino-naphthalene-1-sulfonylamino)ethyl]
-3-mercaptopropionamide
CA Carbonic anhydrase
本論文は学術雑誌に掲載された次の論文を基礎とするものである。
1. Dependence of Hydrolysis of β-lactams with a Zinc(II)-β-Lactamase Produced from Serratia marcescens (IMP-1) on pH and Concentration of Zinc(II): Dissociation of Zn(II) from IMP-1 in Acidic Medium
Masafumi Goto, Hisami Yasuzawa, Toshihiro Higashi, Yoshihiro Yamaguchi, Akiko Kawanami, Shiho Mifune, Hiromasa Mori, Hitoshi Nakayama, Kumiko Harada, and Yoshichika Arakawa
Biol. Pharm. Bull., 26, 9-594 (2003)
2. Detection of Metallo-β-Lactamase (IMP-1) by Fluorescent Probes Having Dansyl and Thiol Groups
Hiromasa Kurosaki, Hisami Yasuzawa, Yoshihiro Yamaguchi, Wanchun Jin, Yoshichika Arakawa, and Masafumi Goto
Org. Biomol. Chem., 1, 17-20 (2003)
3. Probing the Role of Asp120(81) of Metallo-β-Lactamase (IMP-1) by Site-directed Mutagenesis, Kinetic Studies and X-ray Crystallography
Yoshihiro Yamaguchi, Takahiro Kuroki, Hisami Yasuzawa, Toshihiro Higashi, Wanchun Jin, Akiko Kawanami, Yuriko Yamagata, Yoshichika Arakawa, Masafumi Goto, and Hiromasa Kurosaki
J. Biol. Chem., 280, 20824-20832 (2005)
4. Irreversible Inhibition of Metallo-β-Lactamase (IMP-1) by
3-(3-Mercaptopropionylsulfanyl)propionic Acid Pentafluorophenyl Ester
Hiromasa Kurosaki, Yoshihiro Yamaguchi, Toshihiro Higashi, Kimitaka Soga, Satoshi Matsueda, Haruka Yumoto, Shogo Misumi, Yuriko Yamagata, Yoshichika Arakawa, and Masafumi Goto
Angew. Chem. Int. Ed., 44, 3861-3864 (2005)
目次
第
1
章 緒言···1
第
2
章 酸性溶液中におけるmetallo-β-lactamase (IMP-1)
からのZn(II)
イオンの解 離とβ-ラクタム加水分解活性のpH
依存性···9第
1
節 序論···9
第2節 結果
··· 11
2-1
酸性溶液中におけるIMP-1
の加水分解活性とZn(II)
イオン添加による 活性の回復··· 11
2-2
酸性溶液中におけるIMP-1
からのZn(II)
イオンの解離···12
2-3 イミペネムに対する加水分解の pH
依存性···14
第
3
節 考察···15
第4節 小括
···19
第3章 部 位 特 異 的 変 異 法 、 速 度 論 解 析 、
X
線 結 晶 構 造 解 析 を 用 い たmetallo-β-lactamase (IMP-1)
のAsp120(81)
の役割の検討···20
第1節 序論
···20
第
2
節 結果···22
2-1 Asp-120(81)変異体、D120(81)A
およびD120(81)E
の構築···22
2-2 IMP-1
変異体の基質加水分解に対する速度論的パラメータの決定···22
2-3 IMP-1
変異体の基質加水分解活性に対するpH
依存性···242-4 WT
とIMP-1
変異体の前定常状態におけるニトロセフィン加水分解···26
2-5 D120(81)A
とD120(81)E
のX
線結晶構造解析···28
2-6
アニオンによるD120(81)A
の阻害と阻害に対するアニオン依存性····32
第3節 考察
···33
第
4
節 小括···39
第
4
章3-(3-Mercaptopropionylsulfanyl)propionic acid pentafluorophenyl ester
によ るmetallo-β-lactamase (IMP-1)の非可逆的阻害···40
第
1
節 序論···40
第2節 結果
···42
2-1
新規阻害剤のIMP-1
に対する阻害能···42
2-2
ゲルろ過法による非可逆阻害性の検討···422-3
透析法による非可逆阻害性の検討···45
2-4
阻害剤処理IMP-1
のレーザーイオン化質量分析装置(MALDI-TOFMS)
による分子量測定···45
2-5 MPAP
で処理したIMP-1
のX
線結晶構造解析···47
第
3
節 考察···50
第4節 小括
···53
第5章 ダ ン シ ル 基 と チ オ ー ル 基 を 有 す る 蛍 光 プ ロ ー ブ に よ る
metallo-β-lactamase
の検出···54
第1節 序論
···54
第
2
節 結果···55
2-1 DansylC2SH
蛍光量子収率の決定···55
2-2 IMP-1
共存下のDansylC2SH
の蛍光スペクトル変化とIMP-1
との解 離定数の決定···552-3 Alubmin
及びCA
存在下におけるDansylC2SH
の蛍光スペクトルとIMP-1
存在下の蛍光スペクトルとの比較検討···58
2-4 apo IMP-1
存在下におけるDansylC2SH
の蛍光スペクトルとZn(II)イオン
の効果···59
2-5
トリプトファン励起光(280 nm)によるDansylC2SH
共存下のIMP-1
の蛍光スペクトル変化···60
2-6 DansylC2SH
によるIMP-1
阻害の検討···62
第
3
節 考察···63
第4節 小括
···65
第6章 総括 ···67
実験の部···71
謝辞
···88
参考文献···90
第
1
章 緒言人類が
1940
年代半ばにペニシリンの工業的大量生産に成功して以来、各種の抗生 物質の探索や抗菌薬の開発が意欲的に推し進められ、細菌感染症の治療は著しく進歩 した。β-ラクタム剤は、細菌の外膜の主要な構成要素であるペプチドグリカンの生合 成に関与するペニシリン結合蛋白質(PBP)
のtranspeptidase
機能を阻害するので、極め て選択毒性に優れ現在も汎用されている 1。だがこれらの薬剤の使用に伴い、耐性を 持つ菌株が増加している。細菌の耐性機構は抗菌薬の標的部位の変化、酵素による薬 剤の不活化、細胞内への薬剤透過性の低下、細胞外への薬剤排出などが挙げられる。そこで耐性菌の克服とより広い抗菌スペクトルを求めて、第
2
、3
世代へと抗菌薬の 開発が行われた。1980
年代の中頃から、グラム陽性菌から陰性菌までに幅広い抗菌力 を発揮するカルバペネム薬が臨床で使用され始め、β-
ラクタム剤の中でも切り札的な 存在として扱われた2。しかし1990
年代に入ると、カルバペネム薬に耐性をもつ病原 菌が世界各地で報告され、日和見感染症や院内感染の起炎菌として問題となっている。β-ラクタム剤を不活化する酵素、β-lactamase
はβ-ラクタム剤を加水分解することに より、抗菌活性を消失させる。β-lactamase
はアミノ酸配列の相同性からclass A
からD
に 分 類 さ れ て お り 、 酵 素 の 活 性 中 心 が セ リ ン 残 基 で あ るclass A
、C
、D
のserine-β-lactamase
とZn(II)
イオンであるclass B
のmetallo-β-lactamase (MBL)
の2
種類 が知られている3。MBL
は1966
年に非病原菌であるBacillus cereus
から初めて発見さ れ4、1985
年にPseudomonas maltophilia
からL1
が発見された5。1991
年に初めて病原 菌であるBacteroides fragilis
からCcrA
6、Aeromonas hydrophilia
からCphA
7、1994
年にSerratia marcescens
からIMP-1
が発見された8。この時期からMBL
産生菌の迅速な同 定法が報告され9、急速にMBL
産生菌が単離されるようになり、現在までに20
種類 を超えるMBL
が同定されている10。さらに
MBL
は保存されているアミノ酸配列から、subclass B1からB3
まで分類されている 11。Fig. 1 に代表的な
MBL
で保存されているアミノ酸配列と活性中心をsubclass
に分けて比較した。本論文においてアミノ酸番号はclass B β-lactamase (BBL)
numbering
に従い番号付けした。また( )の中はmatured
酵素におけるアミノ酸番号を示す11。例えば
IMP-1
のAsp120(81)
は、BBL numbering
では120
番目、matured IMP-1
では81
番目のアミノ酸番号を示す。Subclass B1 (..HxHxD..)
BcII DVIITHAHADRIGGI CcrA TFIPNHWHGDCTGGL IMP-1 GSISSHFHSDSTGGI VIM-2 RAVSTHFHDDRVGGV BlaB MNIATHSHDDRAGGL IND-1 AVFATHSHDDRAGDL
Subclass B2 (..NxHxD..)
CphA EVINTNYHTDRAGGN
Subclass B3 (..HxHxDH..)
L1 LILLSHAHADHAGPV FEZ-1 ILLISHAHFDHAAGS
116 118 120
116 118 120
116 118 120
A)
Subclass B1 CcrA12
Subclass B2 CphA13
Subclass B3 L114
B)
Fig. 1. A) Comparison of alignment of the amino acid sequences between subclasses
of MBLs
11. B) Comparison of the active site of the three subclasses
12-14.
MBL
のsubclass B1
は、His116-x-His118-x-Asp120
の配列を有しており、BcII
では活 性中心に1
つのZn(II)
イオンを含むが、それ以外は2
つのZn(II)
イオンを含む 15。Subclass B2
は、Asn116-x-His118-x-Asp120の配列を有しており、活性中心にZn(II)イ
オンを1
つ含むが、2
つ目のZn(II)
イオンの結合は活性を阻害することが知られてい る16。Subclass B3は、His116-x-His118-x-Asp120-His121の配列を有しており、活性中 心に2
つのZn(II)
イオンを含む14。Subclass
間の活性中心の構造は、subclass B1
につ いては詳細に後述するが、subclass B2では、1つのZn(II)イオンが Zn2
サイトに結合 し、Zn1
サイトにはZn(II)
イオンが結合せず、またsubclass B3
では、His121
がZn2
サイトの
Zn(II)イオンに配位していることが X
線結晶構造解析の結果から明らかにされた
(Fig. 1B)
13, 14。MBL
であるIMP family
とVIM family
は主として日和見感染で問題となるP.
aeruginosa
やS. marcescens
から発見されている8, 10, 17。IMP
遺伝子、VIM
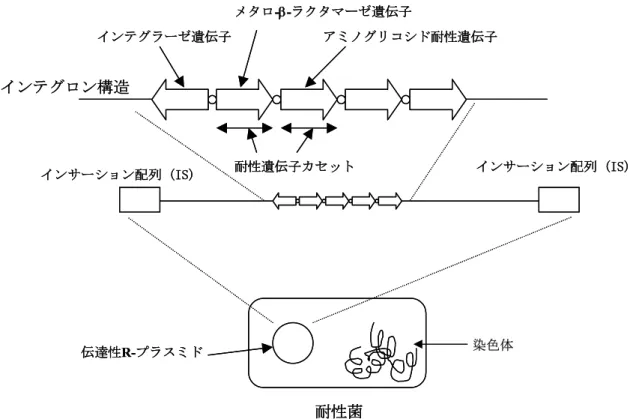
遺伝子は伝 達性プラスミド中に存在するインテグロン構造にカセットとして組み込まれている(Fig. 2)
。インテグロン構造はカセット組み込み・離脱のためのインテグラーゼとカセット遺伝子発現のための
promotor
領域を含むため、水平的に他の細菌に伝播すること ができる18。またカセット遺伝子としてアミノグリコシド耐性遺伝子なども含むこと から、インテグロンを有する細菌は獲得性の多剤耐性菌となる。IMP
遺伝子、VIM
遺 伝子は日本のみならず世界中に拡散しており、またIMP family
はIMP-1
からIMP-13
まで、VIM familyはVIM-2
からVIM-8
まで少しずつアミノ酸配列に変異が入りなが ら発見されることから、特に警戒する必要がある遺伝子である10。そのためIMP family
および
VIM family
に有効な阻害剤開発は急務である。その他のMBL
産生菌の世界中への拡散は
IMP family
およびVIM family
に比べると少ないが、臨床で単離されてい ることから、すべてのMBL
に有効な阻害剤を開発するために、すべてのMBL
に共 通な加水分解機構および基質・阻害剤認識機構を原子・分子レベルで詳細に解明する ことが重要であると考える。インサーション配列 (IS) インサーション配列 (IS) インテグラーゼ遺伝子
メタロ-β-ラクタマーゼ遺伝子
アミノグリコシド耐性遺伝子
耐性遺伝子カセット
インテグロン構造
伝達性R-プラスミド
耐性菌
インサーション配列 (IS) インサーション配列 (IS)
インテグラーゼ遺伝子
メタロ-β-ラクタマーゼ遺伝子
アミノグリコシド耐性遺伝子
耐性遺伝子カセット
インテグロン構造
伝達性R-プラスミド
耐性菌
Fig. 2. The structure of integron mediated the gene encoded IMP family.
CcrA
の3
次元構造と活性中心をFig. 3
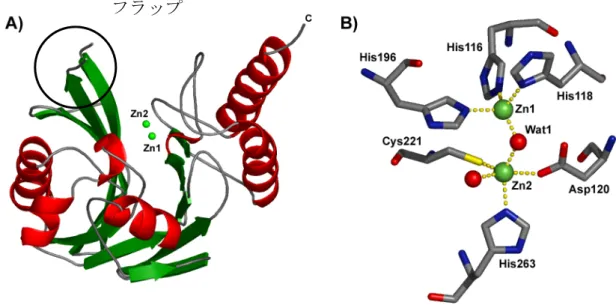
に示す12。MBL
は特徴としてαβ/βαのサンド イッチ構造を持ち、β-
シートの間にZn(II)
イオンが1
つまたは2
つ広くて浅い溝に存 在することがX
線結晶構造解析により報告されている12。この構造は報告されている すべてのMBL
で見られる。また活性部位は広くて浅い溝の底にあるため、様々なβ-
ラクタム剤を収容することができる。そのためほとんどすべてのβ-ラクタム剤を加水 分解することができると考えられている。また活性中心近傍には「フラップ」と呼ば れるヘアピンループが存在し、基質認識に重要であることが知られている(Fig. 3A)。Zn(II)
イオンに対する活性中心の配位構造は、Fig. 3B
で示しているように、1
つのZn(II)
イオン(Zn1)は3
つのHis
と架橋した水分子とで四面体構造をとっており、他のZn(II)
イオン(Zn2)
はもう1
つの水分子とAsp
を頂点として、Cys
,His
と架橋した水分子と でできる三角両錘構造をとっている。染色体
Fig. 3. A) The overall structure of CcrA (PDB code, 1ZNB
12). B) The active site of CcrA
MBL
の加水分解機構は、単核Zn
酵素であるBcII
と、複核Zn
酵素であるCcrA
で 提案されている。Fig. 4で示すようにBcII
では、Zn(II)イオンに配位したOH
2がOH
- となってβ-ラクタムのカルボニル炭素を求核攻撃し、テトラヒドラル中間体形成後,活性中心にある
Asp
がプロトンシャトルの役割をし、β-ラクタム環が加水分解される
19。一方
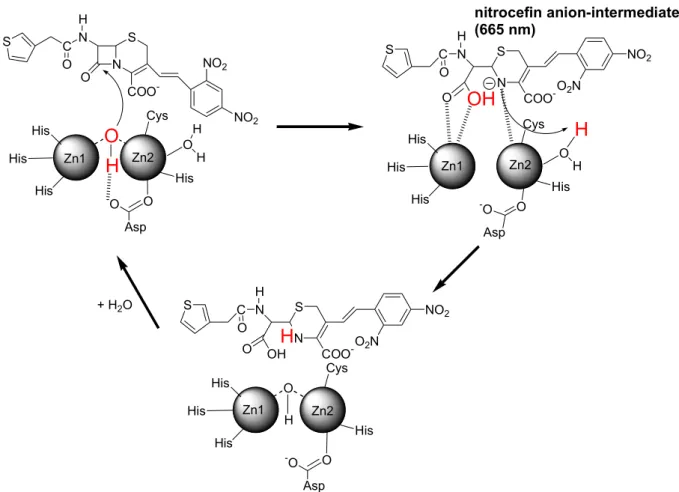
CcrA
では、2
つのZn(II)イオンに架橋した OH
2がOH
-になりBcII
と同じよう に求核攻撃をする。その後Zn2
がアニオン中間体を安定化し、この中間体にプロトン が付加されることによりβ-ラクタム環が加水分解される。このとき活性中心にあるAsp
は求核攻撃の補助をすると考えられている(Fig. 5)12, 20-23。しかし、未だ詳細な反 応機構は明らかにされていない。フラップ
O O Asp His
Zn1 O H
H
+H2O
His His
H
N O OHS
COO-
NO2 O2N
N S C
O O
Asp His
H
Zn1 His
N S
O COO- HN
S C
NO2
NO2 His
OH
O H
O O
Asp H His
Zn1 His
N S
O COO- C N
S
NO2
NO2 H
O
His
nitrocefin (391 nm) nitrocefin tetrahedral-intermediate
hydrolyzed nitrocefin (491 nm)
O
O H
Fig. 4. Proposed mechanism of nitrocefin hydrolysis by the mono-Zn(II) MBL (BcII).
MBL
阻害剤は、1996
年から、trifluoromethyl alcohols and ketones
24、hydroxamates
25、thiols
19, 26-34、thioester 誘導体 28, 35-38、cysteinyl peptides39、biphenyl tetrazoles40, 41、mercaptocarboxylates
28, 39, 42、1β-methylcarbapenem
誘導体43, 44、2,3-disubstituted succinic acid
誘 導 体 45、tricyclic natural products
46、N-arylsulfonyl dydrazones
47、synthetic
cephamycin
誘導体48が報告されているが、いずれも可逆的な阻害剤であり、また数種類の
MBL
しか阻害しない。個々の阻害剤は酵素が違うと阻害の強さが異なることか ら、それぞれ異なった酵素-
阻害剤結合様式をもつと考えられる。MBL
阻害剤を開発 するためには、これらの酵素の1) Zn(II)イオンおよび近隣のアミノ酸残基、 2)
疎水性 残基と阻害剤との結合様式をX
線結晶構造解析などにより分子レベルで活性中心のZn1 Zn2 His Zn2
Zn1 Zn2 His
His
Zn1 His
N S
O
COO-
NO2 NO2 C N
S
H
O
Cys H OH
O
-O Asp
O
H
His HisCys His
His
-O O Asp His
H
N O OHS
COO-
NO2 O2N
N
S C H
O
O H
His
His -O O
Asp N O
OH
S
COO-
NO2 O2N
N
S C H
O
Cys
His O
H
H
+ H2O
nitrocefin anion-intermediate (665 nm)
Fig. 5. Proposed mechanism of nitrocefin hydrolysis by the di-Zn(II) MBL (CcrA)
詳細な構造を明らかにした上で設計することが重要である。
MBL-
阻 害 剤 複 合 体 のX
線 結 晶 構 造 解 析 は 現 在 ま で に 、CcrA
と 阻 害 剤4-morpholinoethanesulfonic acid (MES)の複合体
49、CfiA とtricyclic natural products
の 複 合 体 46、IMP-1
と 阻 害 剤mercaptocarboxylate (2-[5-(1-tetrazolyl-methyl)thien-3-yl]
-N-[2-(mercaptomethyl)-4-(phenylbutyrylglycine)])
の複合体 42 および 2,3-dissubstitutedsuccinic acid
誘導体の複合体 45、Chryseobacterium meningosepticum
由来であるBlaB
とD-captopril
の複合体50、Legionella gormanii 由来であるFEZ-1
とD-captopril
の複 合体 51について報告されている。新しい阻害剤の開発はこれらのX
線結晶構造解析 の結果に立脚し、合理的に分子設計する必要があると考えられる。感染菌の
MBL
産生の有無を確認することは感染症治療の初期段階において極めて重要である。現在知られている
MBL
の検出方法として耐性遺伝子の塩基配列を利用 するPCR
法があるが、高度な技術と特殊試薬を必要とするため臨床検査で日常的に 行うのは困難である 74。2000年にArakawa、Goto
らは、メルカプト酢酸ナトリウムディスク
(SMA)
とセフタジジム(CAZ)
のディスクを並置するダブルディスク法を報告した75。これは、NCCLS法76に従いミューラーヒントン培地に
MacFarland 0.5
の 菌液を綿棒で塗布し、MBL
に対して阻害作用を示すSMA
ディスクとセフェム系抗 菌剤であるCAZ
のディスクを並置して一晩置き、ディスク周辺の発育阻止帯の変化 からMBL
の産生の有無を識別する方法である。この方法は、臨床検査で比較的簡単 に使用することが可能だが長時間を要するという欠点があった。これらの背景の下、本研究は日本で発見され、その耐性遺伝子がプラスミド上に存
在する
IMP-1
を対象として以下の研究を行なった。第2
章ではIMP-1
からのZn(II)
イオンの解離機構をイミペネム加水分解活性の
pH
依存性で検討し、第3
章ではすべ てのMBL
の活性中心に保存されているAsp120
のIMP-1
変異体を調製し、pH
依存性、速度論的解析および
X
線結晶構造解析によりAsp120
の役割を明らかにした。さらに第
4
章ではIMP-1
の三次元構造に基づいて、IMP-1
に対し非可逆的に阻害する新規阻害剤を設計、合成し阻害活性ならびに阻害様式を速度論的解析、
MALDI-TOF MS
、X
線結晶構造解析により解明した。第5
章では簡便にかつ短時間で検査できる蛍光プローブ法で
IMP-1
産生菌を同定するために新規蛍光剤を設計、合成し蛍光スペクトル測定および阻害活性測定から新規蛍光剤の特徴づけを行なった。以下に得られた知見を 詳述する。
第
2
章 酸性溶液中におけるmetallo-β-lactamase (IMP-1)からの Zn(II)イオンの解離
とβ-
ラクタム加水分解活性のpH
依存性第
1
節 序論加水分解酵素の研究において、活性の
pH
依存性を調べることは詳細な反応機構を 知る上で重要な研究手法の1
つである。これは酵素の加水分解反応が、活性中心を構 成している酸性または塩基性側鎖をもつアミノ酸のpK
aと密接に関係しているためで ある。特に活性中心に金属イオンを含有する場合、金属イオンのリガンドへの配位は リガンドのプロトン化と金属イオンとの競争反応によって達成される。金属イオンが 加水分解反応機構に参加している場合、2 つの点に注意を払う必要がある。第一に、金属イオンの解離がリガンドのプロトン化状態に依存するため、
pH
によるリガンド からの金属イオンの解離の検討を行なう必要があること。第二に、加水分解酵素の反 応物(
基質)
への攻撃種は、金属イオンに配位しているOH
2のH
+が解離したOH
-である と考えられており、金属イオンの配位水のpK
a を検討しなければならない。つまり、加水分解酵素にとって、如何に攻撃種である
OH
2のpK
a を下げるかが重要となる。OH
2のpK
aを下げる1
つの手段として正電荷を持ったZn(II)イオンへの配位がある。
これが多くの加水分解酵素の活性中心に
Zn(II)
イオンを含む理由の1
つである。遊離 のOH
2のpK
aは14.0
であるが、Zn(II)-OH2のpK
aは10.0
と下がる。これはZn(II)イオ
ンの正電荷にOH
2の電子が引き付けられる事でH
+が解離しやすいと考えられる。さ らにZn(II)-OH
2にリガンドが配位することでZn(II)-OH
2のpK
aは下がることがわかっ ている。例えば、carboxypeptidase A
は重要な加水分解酵素であるが、そのZn(II)-OH
2の
pK
aは6.1
であると推定されている52。本研究対象である
MBL
の多くは複核Zn
酵素であり、β-
ラクタム環の加水分解を触 媒している。β-ラクタム環のカルボニル炭素への求核試薬は2
つのZn(II)イオンに架
橋して配位している配位水(Zn1-OH2
-Zn2)だと考えられている
12, 14。MBLであり単核Zn
酵素であるBcII
は前述したように、β-
ラクタム環の加水分解に2
つのH
+の関与を 示し、その1
つはZn(II)イオンに配位している Zn1-OH
2でpK
aは5.6
と考えられてい る19。もう1
つのH
+はすべてのMBL
に保存されているAsp
で同じpK
aを持つとされ た19。しかしMBL
で複核Zn
酵素であるCcrA
では、活性のpH
依存性や溶媒同位体 効果による評価を行なったが、pK
aが5
から9
の間にある官能基は存在せず、2
つのZn(II)イオンに架橋した Zn1-OH
2-Zn2
のpK
aは5.0
以下であることが報告されている22, 23。これらのことから単核
Zn
酵素のBcII
と複核Zn
酵素のCcrA
では違う反応機構が提案された53。
しかし
BcII
とCcrA
のpH
依存性の研究では、Zn(II)
リガンドのプロトン化状態のこ とが考慮されていない。このことは(1) リガンドがプロトン化されることでZn(II)イ
オンが酵素から脱離し活性が低下したこと、(2)
活性種であるOH
-(Zn-OH
-)
がプロトン化され
OH
2(Zn-OH
2)になると活性が低下すること、以上 2
つの問題点が重なり、正当な評価ができていないと考えた。
そこで、MBLであり複核
Zn
酵素であるIMP-1
のpH
依存性を調べるために、1
)pH
によるIMP-1
からのZn(II)
イオン脱離の検討2)(1)を基にしたイミペネム加水分解活性の pH
依存性を行なった。この結果を基に、
IMP-1
の加水分解機構のみならず、複核Zn
酵素の反 応機構を解明することが本研究の目的である。Fig. 6. Time-course of absorbance change at 298 nm at 30 ºC for hydrolysis of imipenem on the addition of 10 nM IMP-1 in 50 mM Tris buffer(pH 7.4) to 100 µM imipenem in 0.2 M acetic acid-sodium acetate buffer(pH 5.1)-0.5 M NaCl. After 900 s, Zn(NO
3)
2was added and the measurement was continued.
第
2
節 結果2-1 酸性溶液中における IMP-1
の加水分解活性とZn(II)イオン添加による
活性の回復
酸性溶液中における
IMP-1
のイミペネム加水分解活性は、100 µM
のイミペネムを 含む酢酸緩衝液 (0.2 M, pH 5.1, 0.5 M NaCl)にIMP-1
溶液 (Tris-HCl buffer, 50 mM, pH7.4, 0.5 M NaCl)
を10 nM
になるように加え、298 nm
の吸光度の減少から測定した。イ ミペネムの加水分解は、最初の100
秒の間ですばやく起こったが、時間の経過と共に 急速に活性の減少が観察され、100
秒後には時間変化に伴う吸光度変化は一定となり、反応が終了したかのように見えた(Fig. 6)。
Fig. 7. Dependence of activity, k
2, for hydrolysis of imipenem by IMP-1 on the concentration of Zn(NO
3)
2at pH 5.0.
しかし今回用いたイミペネムがすべて分解された場合、その吸光度変化は
1.0
である と予想されるが、400
秒後では0.1
の吸光度変化しか変化していなかった。そこで、900
秒後にZn(NO
3)
2を100 µM
になるように反応溶液に加えると、酵素活性が回復し、298 nm
の吸光度の急速な減少が観察された(Fig. 6)
。2-2
酸性溶液中におけるIMP-1
からのZn(II)
イオンの解離2-1
から、IMP-1
は酸性溶液中でホロ酵素からZn(II)
イオンの解離によって活性が消失することがわかった。このことから活性のある
Zn
結合型と活性のないZn
解離型の 間に平衡があると考え、酸性溶液中におけるイミペネム加水分解活性のZn(II)
イオン 濃度依存性を調べた。IMP-1
はpH 4.0
から9.0
の緩衝液中で30 ºC
で10
分間、様々なZn(II)
イオン濃度であらかじめインキュベートされ、イミペネム加水分解の初速度v
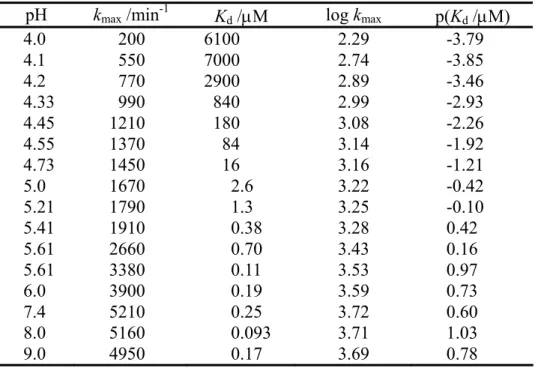
initTable 1. The dissociation constants of Zn(II) from IMP-1 and the limiting k
maxfor the hydrolysis with concentration of Zn(II) determined using imipenem as a reporter substrate.
pH k
max/min
-1K
d/µM log k
maxp(K
d/µM)
4.0 200 6100 2.29 -3.79
4.1 550 7000 2.74 -3.85
4.2 770 2900 2.89 -3.46
4.33 990 840 2.99 -2.93
4.45 1210 180 3.08 -2.26
4.55 1370 84 3.14 -1.92
4.73 1450 16 3.16 -1.21
5.0 1670 2.6 3.22 -0.42
5.21 1790 1.3 3.25 -0.10
5.41 1910 0.38 3.28 0.42
5.61 2660 0.70 3.43 0.16
5.61 3380 0.11 3.53 0.97
6.0 3900 0.19 3.59 0.73
7.4 5210 0.25 3.72 0.60
8.0 5160 0.093 3.71 1.03
9.0 4950 0.17 3.69 0.78
から酵素濃度[E]Tで除することで分子活性
k
2(v
init/[E]
T)を得た。 Fig. 7
はpH 5.0
における
Zn(II)
イオン濃度に対するk
2のプロットを示す。外部からのZn(II)
イオンがない時の
v
0は82 min
-1であったが、IMP-1の分子活性k
2はZn(II)イオン濃度の増加により上
昇し、Zn(II)
イオン濃度が0.5 µM
のときは320 min
-1となり、20 µM
のときほぼプラト ーに達した (k2= 1500 min
-1) (Fig. 7)。
この挙動は酸性溶液中においては、
Zn(II)
イオンが活性中心から解離して、活性の あるZn
結合型と活性のないZn
解離型の間の平衡が存在するという仮定と一致するこ とを示す。活性から求めたIMP-1
からのZn(II)
イオンの解離定数K
dと飽和活性k
maxは、Eq. 9 (p74参照)の非線形最小二乗法を使って求めた。得られた速度論的パラメー タ
K
dとk
maxをTable 1
に示す。解離定数K
dはpH 5.0
から4.0
に減少するにつれて大き く増加した。kmaxと、pH と比較するためにK
dの常用対数をとった-logK
dをpH
に対 してプロットした(Fig. 8)
。Fig. 8
で示すように、pH
に対する-log K
dのプロットはpH
Fig. 8. Dependence of dissociation constant (K
d) (○) and limiting rate constant (k
max) (□) for imipenem by IMP-1 on pH.
5.0
以下で傾きが4.0
の直線を得た。さらにFig. 8
の変曲点から、IMP-1からのZn(II)
イオンの解離に関与している官能基のpK
aは5.3
であることがわかった。2-3 イミペネムに対する加水分解の pH
依存性活性変化から得られた
IMP-1
からのZn(II)イオンの解離定数 K
dを2-2
で得ることが できたので、IMP-1
活性のpH
依存性をそれぞれのpH
におけるK
dの25
倍量のZn(NO
3)
2の濃度で基質であるイミペネムの濃度を変えて分子活性
k
2を求めた。使用した緩衝液 は、pH 4.0
~5.6
では酢酸緩衝液(0.2 M, 0.5 M NaCl)
、pH 5.6
~9.0
ではMTEN
緩衝液で あった。データはMichaelis-Menten
の式(Eq. 10、p74参照)を使って速度論的パラメー タK
mとk
catを求めた。Fig. 9
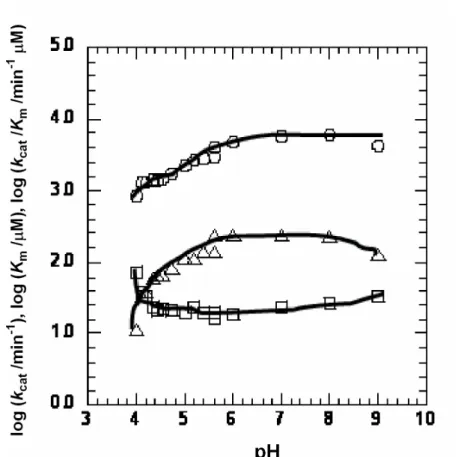
で示したように、イミペネムのlog k
catに対するpH
プロットはpH 7.0
と8.0
Fig. 9. pH profiles of log (k
cat/min
-1) (○), log (K
m/µM) (□) and log (k
cat/K
m/min
-1µM) (∆) for the hydrolysis of imipenem by IMP-1 under sufficient concentration of Zn(II) at 30 ºC.
の間で最大となったが、pH 4.0と
5.0
付近で減少した。logK
mのプロットは、pH 4.8 と6.0
で最小であり、pH 4
付近で急激に上昇した。これらの結果からlog K
m/k
catのプ ロットは、pH 6.0から8.0
の間でプラトーになり、酸性側と塩基性側で縁となるよう な、ベル型となった。第
3
節 考察加水分解酵素の活性の
pH
依存性の研究は、一般的に活性中心は酸性または塩基性 アミノ酸残基を含んでおり、それらのプロトン化状態が触媒的な活性発現に重要であ るため、詳細な反応機構を解明する上で必要不可欠である。MBL
によるβ-
ラクタム剤の加水分解の
pH
依存性はいくつか報告されている19, 52-55。これらのpH
依存性の測 定法において、Zn(II)
イオン濃度は、酵素濃度の10
倍53または500
倍以上19緩衝液に 加えられている。BcII活性のpH
依存性を報告した論文では、酸性領域で傾き2
の直 線が得られ、この結果からBcII
の加水分解反応機構は、2
つの解離できる残基、Zn(II)
イオンに配位したZn1-OH
2 と、加水分解にとって重要な残基として提案されているAsp
が関与していると提案された54。しかしながら金属酵素にとって、金属イオンは 酵素機能発現には活性中心に保存されるべきであり、pH
によるZn(II)イオンに配位し
ているリガンドのプロトン化状態を調べる必要がある。pH 7.4
の条件下にあるIMP-1
がpH 5.1
に調整された基質溶液と混合され、反応が始まると基質に対する活性は
100
秒以内ですばやく減少し、減少した活性はその後維 持された(Fig. 6)。しかし、反応が開始してから900
秒後にZn(II)イオンを添加したと
ころ、酵素活性が回復したことから、Zn(II)
イオンが酸性溶液中でIMP-1
から解離し たことがわかった(Fig. 6)。これは、pH 7.4
の酵素溶液がpH 5.0
の緩衝液に移された時、ある速い変化が起こったため、数
100
秒で酵素活性が不安定な活性になったと考えら れる。これらのことから、酸性溶液中、30 ºCでは最初の100
秒以内にEq. 1
が起こっ ていると考えた。active form non-active form + Zn(II) Eq. 1 K
d酸性領域において、活性から求められた
Zn(II)イオンの解離定数 K
dのpH
依存性は、4
つのプロトン化がZn(II)
イオンの解離に関与して、それら官能基のpK
aは5.3
と考え られる(Fig. 8)。このZn(II)イオンリガンドのプロトン化は、酸性溶液中においてみか
けの
IMP-1
の不活性化に強く関係していた。アミノ酸側鎖のZn(II)
イオンに対する配位は
pH
に強く支配され、1 つ以上のリガンドが協奏的に金属イオンを捕捉する役割をしている。遊離の
His
のpK
aは5.97
であるが、このことからHis
のイミダゾールNδ
がpK
aより低いpH
でプロトン化されることで、酵素分子からZn(II)
イオンが解離 したと考えられる。IMP-1
のZn
含量は酵素1
分子あたり2
つであるが、本研究における活性がないIMP-1
のZn
含量は不明である。しかし、IMP-1 を含むMBL
では、1つのZn(II)イオ
ンだけで活性があると報告されている56-58。IMP-1
からのZn(II)
イオンの解離をEqs. 2, 3
で示す。E-Zn
2E-Zn + Zn
E-Zn E + Zn
K
d1K
d2Eq. 2 Eq. 3
不活性なフリーの酵素濃度[E]、mono-Zn(II)型酵素濃度[E-Zn]と
di-Zn(II)型酵素濃度 [E-Zn
2]
はEqs. 4-6
で示す。[E] = [E]
T1 + K
d1[Zn] +
K
d1K
d2[Zn]
2[E-Zn] = [E]
T[Zn]/K
d21 + K
d1[Zn] +
K
d1K
d2[Zn]
2[E-Zn
2] = [E]
T[Zn]
2/K
d1K
d21 + K
d1[Zn] +
K
d1K
d2[Zn]
2Eq. 4
Eq. 5
Eq. 6
非線形最小二乗法を用いて
Eq. 9
の回帰曲線を当てはめたところ(Fig. 7)、実験値と理 論曲線が一致したことから、di-Zn(II)
型IMP-1(E-Zn
2)
から2
つのZn(II)
イオンが解離して、フリーの
IMP-1(E)になったため、 IMP-1
活性が不活性化されたと考えられる。こ れらのことからE-Zn2
、E-Zn
およびE
をそれぞれ検出することはできなかったが、IMP-1
に結合した2
つのZn(II)イオンが協同的に IMP-1
から解離したことが示唆された。それゆえに、本研究で得られた
K
dは、Eq. 3
のK
d2に対応すると考えられる。また酸性領域で、
pH
に対する-log Kdのプロットの傾きが4.0
であったことから、プ ロトンが解離できるアミノ酸残基がZn(II)
イオンと結合していることがわかった。も っともプロトン化されそうな残基は、2
つのZn(II)イオンに配位している Zn1-OH
2-Zn2、
pK
aが5.97
であるHis
のイミダゾールのNδ
、pK
aが3.96
であるAsp
のカルボキシル基 のOδ1
またはOδ2
が考えられる。IMP-1の活性中心には4
つのHis
と1
つのAsp
がZn(II)
イオンリガンドとして含まれている。しかし遊離のAsp
のOδ1
またはOδ2
のpK
aが
3.96
と5.3
より低いことから、IMP-1
に結合したZn(II)イオンは、 His
のイミダゾー ルのNδ
が、そのpK
aより低いpH
でプロトン化されることで、酵素から放出されたと 考えられる。過剰の
Zn(II)
イオン存在下、イミペネムの加水分解に対するlog k
catは、Fig. 9
で示 すように、pH
に依存した。Good buffer
であるMOPS
とMES
は、Fitzgerald
らがCcrA
で報告したように、これらの化合物がIMP-1
でも阻害したことから活性に影響が出た ため使用しなかった27。logk
catは、pHが下がるにつれて減少したが、pH 4と5
の間 の傾きは0.6
であった。この触媒反応において、求核試薬は、2
つのZn(II)
イオンに架 橋しているZn1-OH
2-Zn2
であると予想される。Fig. 9のプロットから、IMP-1の加水 分解機構に関与する官能基のpK
aは5
より低いと考えられる。この架橋した水分子のpK
aの値が低いことがMBL
の特徴の1
つである。BcII
とCcrA
の活性中心は、それぞ れZn(II)
イオンをそれぞれ1
つ、2
つ含んでおり、Zn1-OH
2またはZn1-OH
2-Zn2
の状 態をとっていると考えられている 12, 16, 53。Mono-Zn(II)酵素であるhuman carbonic
anhydrase II
のZn(II)
イオンに配位しているZn-OH
2のpK
aは6.8
であり、酵素に結合し ていない遊離なZn-OH
2イオンのpK
aの10
より低くなっている59。またCcrA
では、今回の研究と同様に
pH 5
から9
の間でpK
aをもつ官能基はないと報告されている22, 23。 これらのことから、Zn(II)
イオンに配位したOH
2のpK
aは、2
つのZn(II)
イオンが架橋 したZn1-OH
2-Zn2
の脱プロトン化を助けることで、mono-Zn(II)酵素より di-Zn(II)酵素
の方が低くなると考えられる。第
4
節 小括本章では、
MBL
であるIMP-1
の詳細な反応機構を理解するため、IMP-1
からのZn(II)
イオン解離のpH
依存性と活性のpH
依存性について解析した。以下に得られた結果 を要約する。(1) IMP-1
は酸性溶液中において、Zn(II)
イオンが解離することで活性が消失することがわかった。
(2) IMP-1
からのZn(II)イオンの解離は pH
に依存しており、Zn(II)イオンの解離に関 係する官能基は4
つあり、それらのpK
aは5.3
であることがわかった。(3) IMP-1
活性のpH
依存性から、pH 5
から9
の間でpK
aをもつ官能基は存在せず、β-ラクタム環のカルボニル炭素を求核攻撃すると考えられている Zn1-OH
2-Zn2
の架橋している水のpK
aは5
以下であることがわかった。以上の結果から、