熊本大学学位論文
NO 修飾ヌクレオシド :8 −ニトログアノシンと その関連化合物の生物活性に関する研究
2007
金子和義
Biochemical and physiological activities of 8-nitroguanosine and related compounds
Kazuyoshi Kaneko Nitric oxide (NO) plays important roles as a signaling molecule during diverse physiological and pathological events in circulatory, neuronal and
immunological/host defense systems. NO activates soluble guanylate cyclase to produce a second messenger cGMP which mediates down stream signaling pathways. On the other hand, NO also reacts with oxygen radical or heavy metals to form reactive nitrogen oxides that can react with lipids, proteins and nucleic acids to induce their chemical modifications such as oxidation, nitrososylation and nitration. Accumulating evidence has suggested that these NO-mediated modifications may participate in NO signaling.
8-Nitroguanosine is a representative guanosine nitration product. Recently it was found that 8-nitroguanosine production was increased in various inflammatory diseases.
Furthermore, 8-nitroguanosine has unique properties such as redox activity, mutagenic activity against RNA virus, which are not observed in parental guanosine. In this study we examined biochemical and physiological activities of 8-nitroguanosine and its related compounds. First, we established the method of 8-nitroguanine analogues synthesis.
Among them, we characterized a new compound 8-nitroguanosine 3’,5’-cyclic monophosphate (8-nitro-cGMP) focusing on its physical and chemical properties. Second, a mutagenic potential of 8-nitroguanosine was examined for mammalian cell. Finally, vascular signaling activities of 8-nitro-cGMP were investigated.
Synthesis and characterization of 8-nitroguanosine derivatives
8-Nitroguanosine derivatives were synthesized via two step reactions. First, guanosine derivatives were reacted with bromine to introduce bromo group at C8 position with yields of 20-40%. 8-Bromoguanosine derivatives were then reacted with sodium nitrite in dimethyl sulfoxide to replace bromo group with nitro group through nucleophilic substitution that gave yields of 5-20%. Accordingly, we could synthesize a series of compounds, e.g., 8-nitroguanine, 8-nitroguanosine, 8-nitroxanthine, 8-nitro-GMP and 8-nitro-cGMP. This method is simple and convenient, thus can be applicable for the syntheses of many guanine nucleotides and adenine nucleotides/nucleosides.
We found that N-glycosyl bond of 8-nitro-cGMP in physiological condition (pH 7.4, 37℃) is remarkably stable compared with other 8-nitroguanosine derivatives (8-nitroguanosine and 8-nitrodeoxyguanosine). Furthermore, 8-nitro-cGMP is highly resistant against phosphodiesterase-mediated hydrolysis of cyclic monophosphate moiety. These observations suggest that 8-nitro-cGMP possesses unique chemical properties in view of N-glycosyl and cyclic monophospate stabilities.
Mutagenicity of 8-nitroguanosine in mammalian cells
We investigated a mutagenic potential of 8-nitroguanosine in mammalian cells by measuring the occurrence of 6-thioguanine-resistant (TGr) mutants of AS52 cells containing a single functional copy of the bacterial xanthine-guanine phosphoribosyltransferase (gpt) gene. Treatment with authentic 8-nitroguanosine at concentrations of 10–1000 µM for 1 h significantly increased (by 6-8 times) the mutation frequency of the gpt gene without cytotoxic effects. Sequence analysis of the mutated gpt gene showed that 8-nitroguanosine
treatment induced a G-to-T transversion at position 86, which led to substitution of tryptophan with leucine. This mutation pattern differed completely from that determined for spontaneous TGr mutants, whose mutation was a 2-bp deletion. 8-Nitroguanosine treatment significantly increased levels of abasic sites in DNA, whereas it did not affect oxidative DNA damage, as evidenced by immunocytochemical determination of 8-oxodeoxyguanosine formation. These observations suggest that 8-nitroguanosine has mutagenic potential for mammalian cells and hence may play an important role in inflammation-associated carcinogenesis.
Unique vascular signaling activities of 8-nitro-cGMP
We investigated the effects of 8-nitro-cGMP on vascular function. 8-Nitro-cGMP induced a biphasic vascular responses: at high concentrations (>10 µM) it induced relaxation of the precotracted vascular strip, whereas it induced vasocnstriction at the concentration below 10 µM. By using cultured smooth muscle cells, 8-nitro-cGMP was found to activate cGMP-dependent protein kinase (PKG), indicating that vasorelaxation effect of 8-nitro-cGMP was mediated through PKG activation similar to cGMP. On the other hand, we found that the vasoconstriction effect of 8-nitro-cGMP was dependent on endothelial cells.
Interestingly, vessels from eNOS knock-out mice did not respond to 8-nitro-cGMP-mediated vasoconstriction. Electron spin resonance study revealed that 8-nitro-cGMP produced a significant amount of superoxide radical in the presence of eNOS and NADPH.
Furthermore, when vessels were pre-treated with scavengers of superoxide (SOD or Tiron), the vasoconstriction activity of 8-nitro-cGMP was largely suppressed. Taken together, 8-nitro-cGMP has a unique vascular signaling functions: the vasorelaxation activity at high concentrations mediated via PKG activation and the vasoconstriction activity at low concentrations by inducing eNOS uncoupling to produce superoxide.
Metabolism of 8-nitro-cGMP by thiol groups
We investigated the metabolic system of 8-nitro-cGMP. 8-nitro-cGMP reacted with cysteine which have thiol group. The reaction rate constant of 8-nitro-cGMP and thiol group was changed by pH, and it became the maximum around each thiol pKa. We examined the chemical reactim mechanisms of 8-nitro-cGMP with glutathione. The result showed 8-nitoro-cGMP was denitrated by glutathione, and formed an 8-thioallcoxy adduct via guanine C8 position and thiol group. Then, to examine whether 8-nitro-cGMP react with intracellular glutathione, 8-nitro-cGMP react with RAW 264 cell lysate treated BSO, a glutathione biosynthesis inhibitor. The result showed that nitrite in reaction solution decreased depend on BSO concentration. Indeed glutathione concentration in cell lysate was decreased by BSO. We also investigated the physiological function of adduct from 8-nitro-cGMP and glutathione by the organ bath system which thus indicated that the adduct has a significant vasorelaxation effect.
In conclusion, the present study showed that nitration of guanosine provides unique biochemical and physiological functions that may contribute to NO-mediated physiological and pathological phenomena. Particularly, 8-nitro-cGMP is a novel cGMP analogue that can be formed endogenously. Therefore further investigation of signaling activities and metabolic pathways of 8-nitro-cGMP is now warranted.
NO 修飾ヌクレオシド :8 −ニトログアノシンと その関連化合物の生物活性に関する研究
2007
金子和義
Biochemical and physiological activities of 8-nitroguanosine and related compounds
Kazuyoshi Kaneko
NO修飾ヌクレオシド:8−ニトログアノシンとその関連化合物の生物活性に関す る研究
目次
第1章 序論 1
第2章 8-ニトログアノシンヌクレオチドの合成とそのキャラクタリゼーション 6
第1節 研究の背景および目的 6
第2節 実験結果 7
第1項 8-ニトログアノシンヌクレオチドの合成 7
第2項 8-ニトロ-cGMPのモル吸光係数の測定 15
第3項 8-ニトロcGMPのN-グリコシド結合の安定性 15 第4項 8-ニトロcGMPのホスホジエステラーゼ(PDE)耐性の測定 17
第3節 考察 18
第3章 8-ニトログアノシンの変異原性の解析 22
第1節 研究の背景 22
第2節 研究の目的 23
第3節 実験結果 24
第1項 8-ニトログアノシンのAS52細胞に対する変異誘発作用 24
第2項 8-ニトログアノシンによる酸化的DNA損傷の解析 26 第3項 8-ニトログアノシンによるゲノムDNA上の脱塩基部位形成作用 27
第4節 考察 28
第4章 8-ニトロ-cGMPの血管機能に対する影響の解析 31
第1節 研究の背景および目的 31
第2節 実験結果 31
第1項 ラット頸動脈に対する8-ニトロ-cGMPの作用 31 第2項 血管内皮非依存性の弛緩反応 33
第3項 8-ニトロ-cGMPによるPKGの活性化 34
第4項 血管内皮依存性の収縮反応 35
第5項 内皮型NO合成酵素(eNOS)欠損マウス血管を用いた解析 36
第6項 8-ニトロ-cGMPによるeNOSアンカップリングを介したスーパーオ
キサイド生成 38
第7項 8-ニトロ-cGMP誘発性血管収縮反応に対するスーパーオキサイド消
去剤の影響 39
第8項 8-ニトログアノシンによる血管収縮反応 39
第3節 考察 40
第5章 8-ニトロ-cGMPとSH化合物との反応の解析:新規8-チオアルコキシ
-cGMP誘導体の生成とその血管弛緩作用
第1節 研究の背景および目的 42
第2節 実験結果 42
第1項 8-ニトロ-cGMPとアミノ酸との反応の解析 42
第2項 8-ニトロ-cGMPとSH化合物の反応産物の解析 45
第3項 8-ニトロ-cGMPのRAW264細胞溶解液による分解 47
第4項 8-チオアルコキシ-cGMPの血管機能に対する影響 49
第3節 考察 50
第6章 総括 52
実験の部 56
参考文献 72
謝辞 80
本論文で使用した略号一覧表
BSO buthionine sulfoximin
8-nitro-cGMP 8-nitroguanosine 3’,5’-cyclic monophosphate 8-Br-cGMP 8-bromoguanosine 3’,5’-cyclic monophosphate cGMP guanosine 3’,5’-cyclic monophosphate
DMF N,N-dimethylformamide DMSO dimethyl sulfoxide
DTCS N-(dethiocarboxy)sarcosine
EDRF endothelium-derived relaxing factor GMP Guanosine 5’-monophosphate
GDP Guanosine 5’-diphosphate GTP guanosine 5’-triphosphate
HPLC high performance liquid chromatography IFN-γ interferon -γ
IL-1β interleukin-1β IL-4 interleukin-4 LPS lypopolysaccharide MS mass spectrometry NOS nitric oxide synthase PBS phospate bufferd saline PDE phosphodiesterase
PKG cGMP dependent protein kinase sCG soluble guanylate cyclase
SDS sodium dodecyl sulfate TNF-α tumor necrosis factor-α
VASP vasodilator-stimulated phosphoprotein
発表論文リスト
(1)
Sawa T., Akaike T., Ichimori K., Akuta T., Kaneko K., Nakayama H., Stuehr D J., and Maeda H. Superoxide generation mediated by 8-nitroguanosine, a highly redox-active nucleic acid derivative. Biochem. Biophys. Res. Commun. 311, 300-6 (2003).
(2) Kuniyasu A., Kaneko K., Kawahara K., and Nakayama H. Molecular assembly and subcellular distribution distribution of ATP-sensitive potassium channel proteins in rat hearts. FEBS Let.t 552, 259-63 (2003).
(3) Sawa T., Zaki M.H., Okamoto T., Akuta T., Kaneko K., Tokutomi Y., Kim-Mitsuyama S., Ishima Y., Kobayashi A., Yamamoto M., Fujii S., Arimoto H.
and Akaike T. Potential involvement of physiologically formed 8-nitroguanosine 3',5'-cyclic monophosphate in nitric oxide-induced signal transduction. Nature Chemical Biology (Article), in revision.
(4) Kaneko K., Akuta T., Sawa T., Kim H W., Fujii S., Okamoto T., Nakayama H., Ohigashi H., Murakami A., and Akaike T. Mutagenicity of 8-nitroguanosine, a product of nitrative nucleoside modification by reactive nitrogen oxides, in mammalian cells. Cancer Sci., in revision.
1 第1章 序論
一酸化窒素(nitric oxide: NO)は電荷が中性の小さな分子であり、分子内に不 対電子を一個含むフリーラジカルである。そのため生体内において細胞膜を通 って自由に他の細胞に拡散し、生体内の多様な反応に関わる細胞間情報伝達因 子として働いている。NOの生理作用に関しては1980年Furchgottらにより発 見された内皮由来血管弛緩因子(endothelium-derived relaxing factor : EDRF) の研究により幕が開いた1。その後、1986 年FurchgottおよびIgnarroらによ りEDRFの本体はNOであることが証明された2,3。現在までNOの生理作用は 幅広く研究され血管内皮やマクロファージだけでなく循環系、中枢/末梢神経 系、免疫系の調節に関わっていると報告されている4,5。
生体内においてNOはNO合成酵素(NO synthase:NOS)によりL-アルギニン から産生される。NOSはこれまで神経型NOS(neuronal NOS:nNOS)、誘導型 NOS(inducible NOS:iNOS)、および内皮型NOS(endothelial NOS:eNOS)の3 つのアイソフォームが同定されている6。
このうちnNOSとeNOSはそれぞれ主に神経細胞の細胞質、血管内皮細胞の 細胞質膜に特異的かつ恒常的に発現しており、細胞内のカルシウム濃度に依存 して NO 合成活性を発揮する 7。このためこの2つは構成型 NOS(constitutive
NOS:cNOS)とも呼ばれる。これに対してiNOSはマクロファージをはじめとし
て、上皮細胞、血管平滑筋細胞、心筋細胞、肝細胞など様々な細胞に発現し、
細胞質内に局在する。iNOSはその名前からもわかるように平常時の細胞では発 現が認められず、細胞外からの様々な刺激により転写レベルで発現が誘導され8、 また他の2つのアイソフォームとは異なり NO 合成はカルシウム濃度に依存し ない 7,9。したがって、ひとたび発現が誘導されると長時間にわたり、cNOS と 比較して多量のNOを産生し続ける。iNOSを誘導する因子としては細菌の外膜 構成成分であるリポポリ多糖(lipopolysaccharide: LPS)、炎症性サイトカインで あ る イ ン タ ー フ ェ ロ ン-γ (interferon-γ:IFN-γ) 、 イ ン タ ー ロ イ キ ン -1β(interleukin-1β:IL-1β)および tumor necrosis factor-α(TNF-α)などがある
10,11。逆に発現を抑制する因子としてはIL-4、transfoming growth factor-βなど
が知られている 12,13。これらに加え、補酵素であるテトラハイドロビオプテリ ン、基質であるL-アルギニンの合成・分解なども NO産生調節に関与すること が報告されている14,15。
NO の生物活性の発現において細胞内のシグナル経路としてこれまでもっと
2
もよく研究されてきたのが、可溶性グアニル酸シクラーゼ(soluble guanylate cyclase; sGC)の活性化を介したcGMP(guanosine 3’,5’-cyclic monophosphate) をセカンドメッセンジャーとする経路である(図1)。すなわちNO は sGC 内の ヘム鉄へ結合し、sGC の立体構造を活性化型へと変化させ、GTP(guanosine 5’-triphosphate)から cGMP を生成する。cGMPはcGMP 依存性キナーゼを活 性化し、下流のリン酸下シグナルへと情報を伝達する。例えば血管系の場合、
血管内皮で生成したNOは平滑筋細胞へ移行し、sCGを活性化する。その結果 生成したcGMPが平滑筋の弛緩を誘導する。
一方、NOは共存する酸素ラジカルや金属と反応し、化学反応性に富む活性窒 素酸化物へと変換される。活性窒素酸化物はさらに核酸、タンパク質、脂質な どと反応し、それらを酸化したり、ニトロソ化(NO 付加反応)やニトロ化(NO2
付加反応)したりする。このような物質による生体分子の化学修飾はこれまで生 体分子の構造を変化させ、その機能の低下・失活を引き起こす生体損傷反応と
3
いう側面から研究されてきた。ところが、近年、生成した修飾産物が、元の生 体分子にはない新しい生物活性を獲得し、NOの新たなシグナル分子として様々 な生物活性に関与することが分かってきた。
図2にはこれまでに同定されたニトロ化生体分子の構造を示す16-22。ニトロ チロシンはこれまでパーオキシナイトライトなどの活性窒素酸化物のバイオマ ーカーとして生体内検出が行われてきた。しかしニトロチロシンそのものは化 学的・生物学的反応性に乏しく、生体内のニトロ化を反映する単なるバイオマ ーカーとして位置づけられている。つまりニトロチロシンの生成だけでは、ニ トロ化による生物学的効果を説明できないというのが一般的なコンセンサスで ある。
それに対して核酸成分の化学修飾は、その遺伝子変異作用との関連から注目 されている。例えば、8-ニトログアニンあるいは 8-ニトログアノシンは、活性 窒素酸化物であるパーオキシナイトライトによるグアニン塩基の修飾体として
4
Ohshimaらにより報告された23。その後、8-ニトログアニンは、好中球由来の
MPO の触媒により過酸化水素と亜硝酸イオンとの反応からも生成することが 明らかとなり24、炎症反応における新しい修飾塩基として注目されている。
我々はこの8-ニトログアニンに注目し研究を行ってきた。すなわち、Akaike らは、作成した抗 8-ニトログアニン抗体を用いた免疫学的手法によりインフル エンザ感染マウスの気管支上皮細胞に 8-ニトログアニンが生成していることを 明らかにし 25、内因性の 8-ニトログアニンを初めて同定した。その後、様々な 炎症病態でその炎症組織において 8-ニトログアニンの生成が亢進している例が 報告されている。興味深いことにこれらの病態の多くが発がんのリスクを伴う 病態であり、上記の持続的な炎症による発がんが 8-ニトログアニンの生成を介 していることを示唆している点である。さらに興味深いことに、8-ニトログアノ シンはP450還元酵素やNOSの共存下、スーパーオキサイドラジカルを生成す る、いわゆるレドックス活性分子であることが明らかとなった。元のグアノシ ンにはこのようなレドックス活性は認められず、ニトロ化修飾による新しい機 能が付加されたことを示している。
ところで、8−ニトログアニンの生成についてはいくつかのグループより報告 されている。しかし、生体内においてグアニン塩基は、グアニン、グアノシン だけでなく、グアノシン-5’-一リン酸(guanosine 5’ –monophosphate: GMP)や、
cGMP など様々な形で存在する。さらにグアニンヌクレオチドは、遺伝子構成 成分としての機能に加え、cGMPの様なシグナル活性分子でもある。
このような研究背景をふまえ、本研究では 8-ニトログアノシンに関して、以 下の4つのテーマで研究に取り組んだ。
1. 8-ニトログアニン誘導体は 8-ニトログアニン研究において有用なツール となり得る。そこで 8-ニトログアニン、8-ニトログアノシン、およびそのヌク レオチドの合成法を確立した。また、新規化合物である 8-nitroguanosine 3’,5’-cyclic monophosphate (8-ニトロ-cGMP)に関してはその物理化学的性質を 解析した。
2. 8-ニトログアニンの生成が発がんのリスクを伴う病態と関連することに
注目し、哺乳類細胞に対する変異頻度を調べた。合わせて、その変異スペクト ルを解析した。また、8-ニトログアノシンの変異誘発作用のメカニズムを調べる
ために 8-ニトログアノシンによる DNA の酸化的損傷と脱塩基部位の形成を調
べた。
5
3. 8-ニトロ-cGMP は細胞内の NO 産生に依存して生成する全く新規の
cGMP 誘導体である。このものが、血管機能に対してどのような作用があるの かは全く分かっていない。そこで、本研究ではorgan bathの系を用いて血管機 能に対する8-ニトロ-cGMPの作用ならびにそのメカニズムを解析した。
4. 8-ニトロ-cGMP の生体内での代謝経路は全く分かっていない。そこで8-
ニトロ-cGMPと反応する生体分子を探索した。
6
第2章 8-ニトログアノシンヌクレオチドの合成とそのキャラクタリゼーショ ン
第1節 研究の背景および目的
8-ニトログアニンの合成法として、これまで(1)グアニンをパーオキシナイト ライトで処理する方法2326、(2) 8-ブロモグアニンを亜硝酸ナトリウム共存下、
DMSO中160℃で加熱する方法27 、および(3) 8-アミノグアニンを酸性条件下、
亜硝酸ナトリウムと反応させる方法が報告されている28 29。(1)のパーオキシナ イトライトの反応では重炭酸にニトロ化を高める作用が報告されている 30。一
方、8-ニトログアノシンの合成は8-ブロモグアノシンをDMSO中25、あるいは
DMF 中 31、亜硝酸イオンと反応させる方法が報告されている。上記(2)(3)の方 法は8-ニトログアノシンのN-グリコシド結合の不安定性のために適用できない。
一方で、生体内においてはグアノシンにリン酸が付加したGMP、GDP、GTP、
環状リン酸が付加したcGMPなどのヌクレオチドがあるがこれらグアニンヌク レオチドのニトロ化体の合成はこれまでほとんど検討されていない。そこで本 研究では、8-ニトログアニンヌクレオチドの効率の良い合成法の確立を目的とし た。具体的にはグアニン骨格の 8 位にブロモ基を導入し、ついでニトロ基と置 換する方法について検討した(図1)。図2には今回合成した8-ニトログアニン関 連化合物の構造式を示す。この中で8-ニトロ-cGMPは新規化合物である。そこ
でこの 8-ニトロ-cGMP についてはモル吸光係数ならびに N-グリコシド結合の
安定性について解析した。さらに環状リン酸を加水分解する酵素であるホスホ ジエステラーゼに対する8-ニトロ-cGMPの感受性について検討した。
7 第2節 実験結果
第1項 8-ニトログアノシンヌクレオチドの合成
1-1. 8-ニトログアノシンおよび8-ニトログアニンの合成
8-ニトログアノシンは既報にしたがって合成した25。8−ニトログアノシンの
N-グリコシド結合は不安定であり、ニトロ化反応中に分解してリボースと8-ニ
トログアニンが副成する。したがって8-ニトログアニンは8-ブロモグアノシン と亜硝酸イオンの反応の副産物として得た。反応溶液をHPLCで分離した結果 を図3に示す。得られた8-ニトログアノシンおよび8-ニトログアニンはそれら のUVスペクトルならびにマススペクトルにより確認した。(図4)。収率は反応 に用いた8-ブロモグアノシンに対して8-ニトログアノシンが4.8 %、8-ニトロ グアニンが10.8 %であった。
8 1-2 8-ニトロキサンチンの合成
8-ブロモキサントシンはキサントシンをブロモ化した後、HPLC により精製 して得た(図5)。得られた8-ブロモキサントシンはマススペクトルにより確認し
た(図 6)。8-ブロモキサントシンの収率は用いたキサントシンに対して 21 %で
あった。
9
8-ブロモキサントシンをDMSO中、亜硝酸ナトリウムと反応させた結果、8- ニトロキサントシンは得られず、8-ニトロキサンチンのみが生成した。これは 8-ニトロキサントシンのN-グリコシド結合が不安定であり、ニトロ化反応中に
10
分解してリボースと 8-ニトロキサンチンが生成したためと考えられる。精製の HPLCチャートを図7に示す。8-ニトロキサンチンの生成はその分子量(197)よ り確認した(図8)。8-ニトロキサンチンの収率は反応に用いた8-ブロモキサント シンに対して44.2 %であった。
1-3 8-ニトロ-GMPの合成
GMP と臭素の反応液を HPLC により分離し、その 37 分から 52 分を
11
8-Br-GMPを含むフラクションとして回収した(図9)。8-Br-GMPの生成はその 分子量 (441, 443)より確認した(図10)。8-Br-GMPの収率は用いたGMPに対 して22.8 %であった。
ついで 8-Br-GMP を亜硝酸ナトリウムと反応させた。つづいて 8-ニトロ -GMPを逆相HPLCによる3段階の操作で精製した。それぞれの段階のHPLC チャートを図11 に示す。8-ニトロ-GMPの生成はその分子量 (408)より確認し た(図12)。8-ニトロ-GMPの収率は用いた8-Br-GMPに対して16.1 %であった。
12
13 1-4 8-ニトロ-cGMPの合成
cGMPと臭素の反応液をHPLCにて分離し、38分から65分に溶出するフラ クションを回収した(図13)。8-Br-cGMPの生成はその分子量(423, 425)より確 認した(図14)。8-Br-cGMPの収率は用いたcGMPに対して31.3%であった。
ついで 8-Br-cGMPを亜硝酸ナトリウムと反応させた。8-ニトロ-cGMPは逆 相 HPLC を用いた3段階の操作で精製した。それぞれの段階の HPLC チャー トを図 15 に示す。8-ニトロ-cGMP の生成はその分子量(390)より確認した(図 16)。 8-ニトロ-cGMP の収率は、反応に用いた8-Br-cGMP に対して21.3 %
14 であった。
15
第2項 8-ニトロ-cGMPのモル吸光係数の測定
図17Aに示すように、pH7.4の緩衝液中で8-ニトロ-cGMPは、250, 270, 397 nmにピークを持つ吸収スペクトルを示した。すでに8−ニトログアニンや8−ニ トログアノシンについて報告のあるように、397 nm の吸収は 8−ニトログアニ ン骨格に特有の吸収である。この 397 nm の吸収を、段階希釈した 8-ニトロ -cGMPに対して測定した結果を図17Bに示す。この関係より、8-ニトロ-cGMP
の397 nmにおけるモル吸光係数が10,000と算出された。これまでに報告のあ
る8−ニトログアニン関連化合物のモル吸光係数をあわせて表1に示した32。
第3項 8-ニトロ-cGMPのN-グリコシド結合の安定性
逆相 HPLC により 8-ニトロ-cGMP および関連物質を分離し、電気化学検出
器で検出した例を図18Aに示す。このように、本実験条件下では、4つの関連 化合物、すわなち、8-ニトロキサンチン、8-ニトログアニン、8-ニトログアノシ ン、および8-ニトロ-cGMPを完全に分離することができた。さらに、8-ニトロ
16
-cGMPの検出限界について、標準試料を用いて検量線を作成した結果を図18B
に示すが、注入絶対量として25 fmolの感度まで検出可能であった。
さらに、8-ニトロ-cGMPおよび8-ニトログアノシンのN-グリコシド結合の安 定性について調べた結果を図19に示す。8-ニトログアノシンは、pH 7.4、37 °C でインキュベーションすることにより、時間依存的に加水分解された。このと き、半減期が約22.4時間と見積もられた。一方、この条件下で8-ニトロ-cGMP は全く分解を受けず、そのN-グリコシド結合が非常に安定であることが明らか となった。DNA上の8-ニトログアニンは、半減期約4時間で加水分解されるこ とが報告されている 33。従って、環状リン酸構造は、8-ニトログアノシンの N- グリコシド結合を著明に安定化させる作用があることが分かった。
17
第4項 8-ニトロ-cGMPのホスホジエステラーゼ(PDE)耐性の測定
図20には、HPLC-電気化学検出器を用いたauthenticな8-ニトロ-cGMP, 8- ニトロ-GMP, 8-ニトログアニンの分離例、および8-ニトロ-cGMPを0および1
mU/ml のホスホジエステラーゼ 1(PDE1)と反応させた溶液の分離例を示す。
PDE は 3’,5’-環状ヌクレオチドを加水分解して 5’-リン酸ヌクレオチドを生成す
る酵素であり、生体内で環状ヌクレオチドの寿命を規定する主要な酵素である
34,35。図20のように、8-ニトロ-cGMPは、PDE1と反応後もそのままで残って
いること、また、8-ニトロ-GMPに一致する部分には全くピークが現れていない ことから、分解反応は起こっていないことが示された。図21にはPDE1、PDE5 による反応を、cGMP、8-Br-cGMP、8-ニトロ-cGMPについて調べた結果をま
とめた。8-Br-cGMPは、PDEに対して耐性であることが知られているが、本実
験でも確認できた。さらに本実験において、8-ニトロ-cGMP も、PDE1および PDE5に対して耐性であることが明らかとなった。
18 第3節 考察
本実験では-8-ニトログアニン誘導体の合成法として、まずグアニン骨格のC8 位にブロモ基を導入し、ついでそのブロモ基を亜硝酸イオンで求核置換反応す る方法について検討した。ブロモ化については、今回検討したキサントシン、
GMP、cGMPのいずれも収率が20-30 %程度であった。このように収率が低く なった原因として、溶媒抽出における回収ロスが考えられる。今後は固相抽出 などより効率の良い抽出・精製法の検討が必要である。
今回検討した亜硝酸イオンによる求核置換反応を利用したニトロ基の導入法
19
は比較的温和な条件(70℃)で進行し、N-グリコシド結合を維持したままでニト ログアニンヌクレオシド・ヌクレオチドを合成できた。ただし、8-ニトロキサ ントシンについては、おそらくN-グリコシド結合が非常に不安定であるために 合成中に速やかに分解したものと考えられた。実際、8-ニトロキサンチンが生 成していた。一方8-ニトログアノシンに比べ、8-ニトロ-cGMPは4倍程度収率 が高かったが、これは、生成した8-ニトロ-cGMPが安定なために、効率よく精 製できたためと考えられる。実際、8-ニトロ-cGMP の N-グリコシド結合は 8- ニトログアノシンのそれに比べ非常に安定であることが分かった。以上より、
今回検討した方法により 8-ニトログアニン、8-ニトログアノシン、8-ニトロキ サンチン、8-ニトロ-GMP、および8-ニトロ-cGMPの合成に成功した。本合成 法は 8-ニトログアニン誘導体の簡便な合成法として有用であると考えられる。
また、本合成法は他のグアニンヌクレオチド(GDPやGTP)やアデニンヌクレオ シド・ヌクレオチドのニトロ誘導体合成に対しても適用が可能であろう。
8-ニトロデオキシグアノシンの N-グリコシド結合の安定性については、これ までに複数のグループが報告している。DNA中の8-ニトログアニンの安定性に ついて、Ohshima らはpH 7.4、37℃では約 4時間の半減期で加水分解すると 報告している33。一方、Chenらは、pH 7.0-7.5においてパーオキシナイトライ ト処理したウシ胸腺 DNAからの 8-ニトログアニンの脱プリン化は、約 1 時間 の半減期で起こることを報告している 36。同様の結果(半減期=約 1 時間)を、
Tretyakowaらはパーオキシナイトライト処理したプラスミドDNAについて報
告している27。SodumとFialaらは、8-ニトロデオキシグアノシンのN-グリコ シド結合の安定性について、pH 7.0、-20℃で44時間、0℃で7時間、室温では 10分、37℃で3分以下の半減期であると報告している29。このように8-ニトロ グアニンとデオキシリボースとの間のN-グリコシド結合は非常に不安定である。
これに対し2’位が水酸基となったリボース誘導体ではN-グリコシド結合が安 定化されることが報告されている。すなわちSodumとFialaらは、8-ニトログ アノシンのN-グリコシド結合の半減期はpH 7.0、37℃で5時間であり、8-ニト ロデオキシグアノシンと比べて 100 倍以上安定であると報告している 29。 Masudaらは、RNA中の8-ニトログアニンの安定性について調べ、pH 7.4、37℃ で 6時間インキュベーションしても、約 5 %が加水分解されるにすぎないと報 告している37。今回筆者が調べた結果では、8-ニトログアノシンはpH 7.4、37℃
で6時間のインキュベーションにより約15 %加水分解されたことより、半減期
20
は約22.4時間と見積もられた。Sodum、Fialaらの測定値との違いは、pH(7.0 と7.4)、塩濃度(0.5と 0.05 Mのリン酸ナトリウム)などの影響が考えられる。
いずれにしても 2’位の水酸基は、N-グリコシド結合の安定性に寄与しているこ とが分かる。今後、2’位の水酸基以外の化学修飾による N-グリコシド結合の安 定性向上なども検討することは興味深いことと思われる。
ところで、本研究において、3’,5’-環状リン酸構造を持つ 8-ニトロ-cGMP が N-グリコシド結合の加水分解に対して非常に安定であることが分かった。すな
わち、pH 7.4、37℃において検討した68時間までは、ほとんど分解しないこと
が分かった。このことから 3’,5’-環状リン酸構造は 8-ニトログアニンの N-グリ コシド結合を劇的に安定化させることが分かった。
細胞内で生成した環状ヌクレオチドは、通常ホスホジエステラーゼ(PDE) の作用によって速やかに5’-リン酸ヌクレオチドへと分解される34 35。現在まで にPDEのアイソフォームとしてPDE1からPDE11までが報告されている3435。 これらのアイソフォームは基質(cGMP か cAMP)に対する選択性や発現組織 などに違いがある。今回用いたPDE1はcGMPとcAMPをほぼ同等に加水分解 し、その活性はカルモジュリンにより増強される38。一方、PDE5はcGMP選 択的なPDEとして知られている35。これまでにPDEによる加水分解に耐性な アナローグとして、さまざまな cGMP誘導体が合成されている。特に C8 位に 化学修飾したもの、例えばブロモ化やアルキル化は PDE 耐性な cGMP アナロ ーグとして薬理学的な研究に用いられている38。しかし、C8位のニトロ化の影 響についてはこれまで報告がなかった。今回の実験により、C8位がニトロ化さ れると、少なくともPDE1とPDE5に対して著明に耐性になることが分かった。
興味深いことに、従来のC8位修飾cGMPと異なり、この8-ニトロ-cGMPは生 体内のニトロ化反応により生成すると考えられるため、このような生体内での ニトロ化により、生体内での代謝がPDE非依存的になることが予想されること である。実際に、医学薬学研究部・本微生物学分野における最近の成果として、
生体内における8-ニトロ-cGMPの生成が証明された。従って、今後は、8-ニト ロ-cGMPの生体内での代謝メカニズムを調べることが、興味ある課題である。
以上の結果をまとめると、グアニン骨格のC8位にブロモ基を導入し、ついで そのブロモ基を亜硝酸イオンと求核置換反応させることで、各種 8-ニトログア ニン誘導体の合成に成功した。この方法は他のグアニンヌクレオチド、アデニ ンヌクレオチド・ヌクレオシドのニトロ化反応にも適用が期待できる。今後の
21
課題としては、各精製段階における効率を上げ、全体の収率を向上させること が必要である。
新規化合物である 8-ニトロ-cGMP の N-グリコシド結合の安定性について検 討した結果、3’,5-’環状リン酸構造がN-グリコシド結合を強力に安定化さる作用 があることを見出した。一方、cGMPのC8位へのニトロ基の導入は、ホスホジ エステラーゼ耐性を付与するユニークな効果がもたらすことが分かった。
22
第3章 8-ニトログアノシンの変異原性の解析
第1節 研究の背景
微生物感染、化学物質の暴露、あるいは物理的な刺激などが原因となる慢性 炎症が様々な臓器・組織において発がんのリスクを増加させることが明らかと なってきた。そのような例としてHelicobacter pylori (H.pylori) 感染による萎 縮性胃炎と胃がん、潰瘍性大腸炎、クローン病などの炎症性大腸疾患と大腸が ん、ウイルス性慢性肝炎と肝がん、肝吸虫感染と胆管がん、バレット食道と食 道がんなどが挙げられる 39-41。炎症組織では活性化されたマクロファージ、リ ンパ球が浸潤し、それら自身、あるいは間質細胞に成長因子や血管新生因子な どのサイトカインの産生を促す。これら炎症細胞はまた大量に活性酸素や窒素 酸化物を産生する。これら活性酸素、窒素酸化物は核酸、タンパク質、細胞膜 と反応して酸化、ニトロ化、ニトロソ化修飾する 42,43。これらの修飾により生 体分子の生化学的な機能に影響を与える。もしこれらの修飾が適切に修復や代 謝を受けずに蓄積すると、遺伝子変異やアポトーシスの原因となる43。
活性窒素酸化物は核酸と反応し8-ニトログアニンや5-ニトロ-4-グアニジノヒ ダントインなどのニトロ化修飾塩基を生成する 44-46。実際、インフルエンザ感 染マウスの気管支上皮細胞中に 8-ニトログアニンが生成していることが抗8-ニ トログアニン抗体を用いた免疫化学的手法により初めて同定された25。その後、
炎症に関連した様々な病態において 8-ニトログアノシンの生成が亢進している ことが明らかとなってきたが、興味深いことにそれらの病態の多くが発がんリ スクと関連したものである。すなわち、Terasakiらは高頻度に肺がんへと進展 することが知られている特発性肺線維症の患者の気道上皮に顕著に高レベルの 8-ニトログアニン生成を報告している 47。8-ニトログアニンの免疫染色像は iNOSと局在が一致することから、このニトロ化修飾にはiNOSが重要な働きを していることが示唆された。また肺がん患者の扁平上皮がんにおいても 8-ニト ログアニンの生成の亢進が見られた。肺がんのリスク因子である喫煙が尿中 8- ニトログアニンを有意に増加させることも報告されている48。 8-ニトログアニ ン生成の亢進はさらに、H.pylori感染胃上皮49、胆管がん組織 50、慢性C型肝 炎患者の組織中51にみとめられている。
DNA中のグアニンニトロ化とその発がんへの関わりについては、in vitroで 研究されてきた。すでに2章で述べたように、DNA中の8-ニトログアニンは不
23
安定であり、速やかに DNA から遊離して脱塩基部位を形成する。そのため G
→Tトランスバージョン変異を誘発すると考えられている。また、8-ニトログア ニンを含むオリゴデオキシヌクレオチドを用いた検討から、8-ニトログアニンは それ自身もG→Tトランスバージョンを引き起こす可能性が示唆されている52。 しかしながら DNA 中の 8-ニトログアニンが実際に細胞内で変異原性を引き起 こすかはまだ分かっていない。
上述した免疫化学的解析により、グアニンニトロ化は主として細胞質に起こ っていることが示唆されている 25,47,53。これらの事実はヌクレオチドプールや RNA 中の成分として 8-ニトログアノシンが存在していることを示唆している。
8-ニトログアノシンの N-グリコシド結合は安定である 37。さらに、8-ニトログ
アノシンはセンダイウイルスなどの RNA ウイルスに対して変異誘発作用を示 すことが報告されている 53。 特に 8-ニトログアノシン処理したウイルス RNA の変異スペクトルは C→Uトランジッション変異であり、ごくまれに G→Uト ランスバージョン変異が見られた。また、8-ニトログアノシンは NADPH-シト クロムP450還元酵素や NO synthase:NOS のような還元酵素と共存すること でスーパーオキサイドを産生し 25,54、酸化ストレスを誘導することも報告され ている。したがって 8-ニトログアノシンの形成は単なる核酸の化学修飾ではな く、その生物活性から発がんメカニズムに関与する可能性がある。
第2節 研究の目的
本研究では 8-ニトログアノシンが哺乳類細胞に対して変異誘発性を示すか、
また変異誘発性を示す場合、どのような変異スペクトルを示すかを明らかにす ることを目的とした。変異原性の解析には AS52 細胞を用いた。この AS52 細 胞はチャイニーズハムスター卵巣(CHO)細胞のヒポキサンチングアニンホスフ ォリボシルトランスフェラーゼ遺伝子欠損株に、大腸菌のキサンチングアニン ホスフォリボシルトランスフェラーゼ(gpt)遺伝子をゲノム DNA に導入したも のである 55。このgpt 遺伝子に変異が起こると 6-チオグアニンに耐性になるこ と(TGr変異細胞)を利用して変異原性を解析することができる。このAS52細胞 を8-ニトログアノシンで処理し、出現するTFr変異株のgpt遺伝子をシークエ ンス解析して変異スペクトルを調べた。また AS52 細胞を 8-ニトログアノシン 処理した時に誘発されるDNAの酸化的損傷と脱塩基部位の形成も調べた。
24 第3節 実験結果
第1項 8-ニトログアノシンのAS52細胞に対する変異誘発作用
本実験条件下におけるTGr変異細胞の自然発生頻度は4.5 1.5/106 cellsであ った。AS52細胞を変異原性を持つ酸化剤であるH2O2 (100 µM)で処理すると、
変異頻度は 36.5 2.8/106 cells まで上昇した。8-ニトログアノシンは 10-1000 µMの範囲で細胞毒性を示さなかった(図1)。この濃度範囲でAS52細胞を1時 間処理すると、TGr変異細胞の発生頻度が顕著に上昇した(図2)。すなわち、1000 µM 8-ニトログアノシンにより、AS52細胞の変異頻度はコントロールに比べ6 倍に上昇した。この変異頻度の上昇は100 µM H2O2による変異誘発作用に匹敵 するものであった。以上の結果より、8-ニトログアノシンがAS52細胞に対して 強力な変異誘発作用を示すことが示唆された。
25
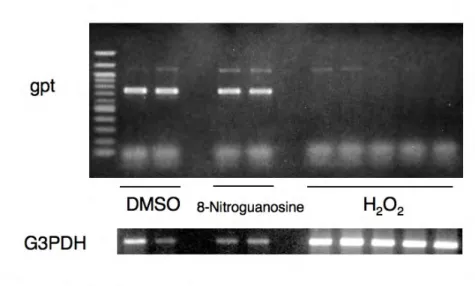
8-ニトログアノシンにより誘発された gpt 遺伝子の変異スペクトルを解析し た。TGr変異細胞から得られたgpt cDNAをアガロースゲル電気泳動した結果 を図 3 に示す。自然発生株および 8-ニトログアノシン誘発変異株ともに約 0.7 kbpのRT-PCR産物が得られた。しかしH2O2処理に由来するTGr変異株から
はRT-PCR産物は得られなかった。この原因としては、変異がgpt遺伝子のプ
ロモーター領域に起こることによって転写活性が損なわれたか、あるいは今回 の実験条件では検出できない異常な転写産物ができていることが考えられる。
シークエンス解析の結果(表1)、正常なAS52細胞においてはgpt遺伝子に全 く変異は見られなかった。自然発生TGr変異細胞では373-374番の2塩基対欠 損のみが起こっていた。これはフレームシフト変異を起こしている可能性があ る。それに対して8-ニトログアノシン処理によるTGr変異細胞を解析したとこ ろ、その変異はすべて86番のG→Tトランスバージョン変異であることが分か った。この遺伝子変異は Trp→Leu のアミノ酸置換をもたらす変異である。今 回調べた中では、gpt遺伝子中に大きな欠損、挿入、タンデム型、複数点変異は いずれのTGr変異細胞にも見られなかった。
26
表 1. TGr変異株の gpt遺伝子変異スペクトルと予想されるアミノ酸置換
試薬 位置 塩基変化 標的配列 アミノ酸置換
DMSO (n=10)
373-374 2-bp deletion GAC TAT GTT Frame shift
8-ニトログア ノシン
(n=15)
86 G:C to T:A CAA TGG AAA 29 Trp to Leu
第2項 8-ニトログアノシンによる酸化的DNA損傷の解析
8-ニトログアノシン処理による酸化的DNA損傷への影響を、酸化的DNA損 傷のマーカー・8-オキソデオキシグアノシン56の生成を指標として調べた。図4 に示すように H2O2処理は有意に 8-オキソデオキシグアノシンの染色を上昇さ せた。これよりH2O2処理が酸化的DNA損傷を誘発していることが示唆された。
これに対し、8-ニトログアノシン処理では 8-オキソデオキシグアノシンの染色 はほとんど増加しなかった。したがって 8-ニトログアノシンは本実験条件下で はAS52細胞に対して酸化的DNA損傷をほとんど誘発しないことが示された。
27
第3項 8-ニトログアノシンによるゲノムDNA上の脱塩基部位形成
DNA 中の脱塩基部位は変異誘発性であり、X→チミンへの突然変異を誘発す る 57。ここでX は脱塩基部位を示す。8-ニトログアノシン処理により AS52 細 胞ゲノムDNAの脱塩基部位数が変化するかどうか解析した。表2に示すように コントロール細胞中の脱塩基部位数に比べて、H2O2処理細胞では脱塩基部位数 がわずかに増加していた。興味深いことに、8-ニトログアノシンで処理した細胞 では、H2O2処理した細胞よりもさらに多くの脱塩基部位が形成していた。した がって、8-ニトログアノシンにはゲノムDNA中の脱塩基部位を増加させる作用 があることが明らかとなった。
28
表 2. AS52 細胞の 8-ニトログアノシンによる脱塩基部位の増加
脱塩基部位数/105 ヌクレオチド 処理後経過時間
コントロール H2O2 8-ニトログアノシン
5時間 0.92 ± 0.16 1.80 ± 0.09* 2.53 ± 0.09**, ***
8時間 0.18 ± 0.16 1.48 ± 0.07* 2.51 ± 0.30**, ***
H2O2と8-ニトログアノシンはともに終濃度100 µMで用いた.
*P <0.002, **P <0.001(vs コントロール), ***P <0.005 compared with H2O2 treatment.
第4節 考察
iNOSによる過剰なNOの産生が起こると、NOが直接、もしくはパーオキシ ナイトライトや二酸化窒素を経てDNA, RNA, ヌクレオシド/ヌクレオチド, タ ンパク質, 脂質をニトロ化修飾する 42,44。 先に述べたように特発性肺繊維症患 者の再生上皮など、がんのリスクを高める病変ではグアニンのニトロ化修飾が 亢進している。しかしながら、このニトロ化修飾がどのように変異原性、発が ん性に関与しているかはまだはっきりと分かっていない。本研究により 8-ニト ログアノシンが AS52 細胞に対して強い変異原性を示すことが初めて明らかと なった。シークエンス解析により、8-ニトログアノシンはgpt遺伝子の86番の G→Tトランスバージョン変異を引き起こすことが明らかとなった。この遺伝子 変異はTrp→Leuのアミノ酸変異をもたらす変異であり、gptタンパク質の機能 変化を引き起こしていることが予想される。この変異のパターンは 373-374 の 2 塩基対欠損を起こす自然発生TGr 変異細胞とは異なっていた。以上の結果か ら、8-ニトログアノシンは哺乳類細胞に対して強い変異誘発作用を持つことが示 された。
G→Tトランスバージョン変異は一般にヒトのがんにおけるsomaticな変異に おいて観察される 58,59。この変異は 8-オキソデオキシグアノシン 60や、その酸 化物 (スピロイミノジヒダントイン、グアニジノヒダントイン、イミノアラント インなど) の生成を伴ったグアニンの酸化的な傷害で起こる61。8-ニトログアノ