表皮ヒアルロン酸合成制御機構の解明
研究分野 生 化 学 紹介教授 野水 基義 学位申請者 佐用 哲也 ヒアルロン酸(HA)は, N-アセチルグルコサミン(NAG)とグルクロン酸(GlcA)の二糖 が交互に結合した高分子多糖で, 哺乳動物の結合組織に広く存在するグリコサミノグ リカンのひとつである. その分子量は 107Da にも及び, 構成単糖のもつ水和能に加え, オーバーラップした三次元ネットワークが水分子を束縛することにより, 高い水和体 積を獲得する. HA は水和ゲルとして組織の水分保持を担い, また組織に機械的特性を 賦与する. 皮膚真皮に存在する HA は, 細胞外マトリックスを構築する分子として機 能する一方で, 表皮の HA は重層した表皮細胞の細胞間隙に存在して細胞間スペース を維持し, 細胞の増殖や分化に寄与する. 皮膚の HA は生体 HA の約 50%を占めるが, 加齢により減少することが知られている. 細胞の増殖・分化といった基本的な細胞機 能に関与するHA の減少は, 皮膚機能の低下に関わると考えられ, HA 代謝制御剤は抗 老化機能性素材として有望である. 皮膚HA の半減期は 0.5-1.5 日と短く, そのターンオーバーを担う HA 合成系は, 厳密 に制御されていると推測される. HA 合成酵素(HAS)には 3 種のアイソザイムが存在す ることが知られているものの(HAS1, HAS2, HAS3), ヒト表皮において, いずれのアイ ソザイムによりHA 合成が調節されているかは不明である. 本申請論文では, 表皮 HA の合成制御機構を明らかにするとともに, 抗老化機能性 素材につながる化合物を探索することを目的とし, 以下の 3 章において正常ヒト表皮 細胞おけるHA 合成制御因子の作用機構を詳細に検討した. 第1 章 HA 合成酵素遺伝子(HAS)発現変動による HA 合成制御 ヒト表皮細胞のHA 合成を惹起するサイトカインとして IFN-, 逆に抑制するサイト カインとしてTGF-を見出した. それぞれのサイトカイン刺激後の HAS 遺伝子の発現 について検討したところ, HA 合成変動と一致した発現挙動を示したのは, 表皮細胞で ドミナントに発現するHAS3 mRNA であった(Fig. 1). 表皮ヒアルロン酸合成を促進さ せるレチノイン酸の刺激でも HAS3 遺伝子の顕著な発現誘導が観察された. これらの 結果から表皮HA の合成は, 3 種の HAS 遺伝子のうち, 主に HAS3 遺伝子の発現を介し て制御されると考えられた.
HA 合成を促進する. TGF-が, 表皮細胞の 増殖を抑制し, 逆に線維芽細胞の増殖を促 進するサイトカインであることを考えあ わせると, TGF-による合成制御は細 胞増殖時にスペースを確保するという HA の役割を支持する応答であると考えられ た. ヒト表皮細胞により合成されるHA 分子 量サイズは, サイトカインによる刺激の有 無に関わらず, 高分子量(>1×106Da)であ った. したがって, HAS3 により合成される HA の分子量サイズは, 高分子であると考 えられた. 本研究により, 表皮細胞における HA の合成制御には, 3 種の HAS 遺伝子のうち主に HAS3 遺伝子の発現が重要な役割を担っていることが明らかになった. 表皮における 速い HA のターンオーバーを勘案すると, 合成機構だけでなく分解機構の解明も必要 である. 第2 章 レチノイン酸レセプターを介した HA 合成制御 レチノイン酸(RA)はビタミン A の活性本体で, 細胞増殖および分化の制御に重要な 役割を演ずるが, その前駆体はカロテンやク リプトキサンチンに代表される食餌性のプロビタ ミンA カロテノイドである. -カロテンが RA と同 様に, HAS3 mRNA の発現を誘導すること, 加えて その HA 合成誘導が レチノイン酸レセプター (RAR)拮抗薬である LE540 により阻害されたこと により, 表皮細胞におけるプロビタミン A から RA に至る代謝経路の存在が示された. 一方, 哺乳類 ではレチノイドに代謝されないルテイン, ゼアキ サンチンおよびアスタキサンチンなどのノンプロ ビタミンA カロテノイドにも HAS3 mRNA 発現誘 導活性およびその発現挙動と一致する HA 合成の 誘導が観察された. HA 合成に及ぼすルテインの効果は-カロテンと同様, LE540 で抑 制された. ルテインによる刺激後に RARE 依存の転写活性が増大したことから(Fig. 2), 0 2000 4000 6000 8000 Lutein (M) 0 2 5 10 ** ** S E A P a ct iv it y (a rb it ra ry c he m il um in es ce nc e un its )
Fig. 2 Effect of lutein on RAR activation. Significantly different from the control value; **P < 0.01 (William’s test). 0 2000 4000 6000 8000 Lutein (M) 0 2 5 10 ** ** S E A P a ct iv it y (a rb it ra ry c he m il um in es ce nc e un its ) 0 2000 4000 6000 8000 Lutein (M) 0 2 5 10 ** ** S E A P a ct iv it y (a rb it ra ry c he m il um in es ce nc e un its ) S E A P a ct iv it y (a rb it ra ry c he m il um in es ce nc e un its )
Fig. 2 Effect of lutein on RAR activation. Significantly different from the control value; **P < 0.01 (William’s test). 0 1 3 10 IFN-(ng / ml) TGF-(ng / ml) 0 0.3 1 3 HAS1 HAS3 G3PDH 2.4 kb 4.9 kb 2.0 kb
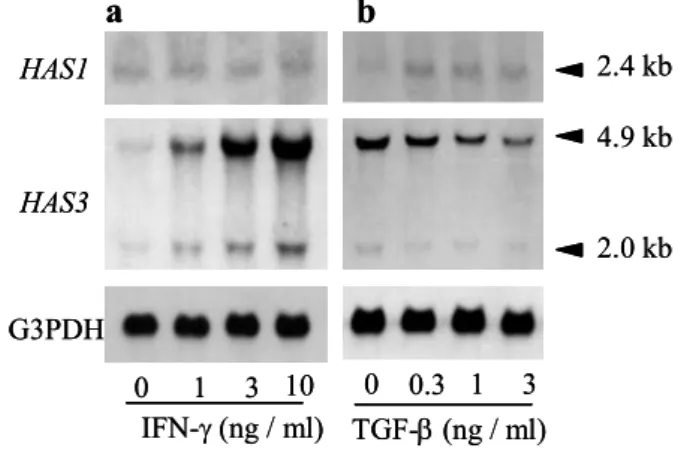
Fig. 1 The HAS3 transcript level of human keratinocytes was upregulated by IFN- but downregulated by TGF- in a dose-dependent manner. Human keratinocytes were cultured in various concentrations of IFN- (a) or TGF- (b).
0 1 3 10 IFN-(ng / ml) TGF-(ng / ml) 0 0.3 1 3 HAS1 HAS3 G3PDH 2.4 kb 4.9 kb 2.0 kb
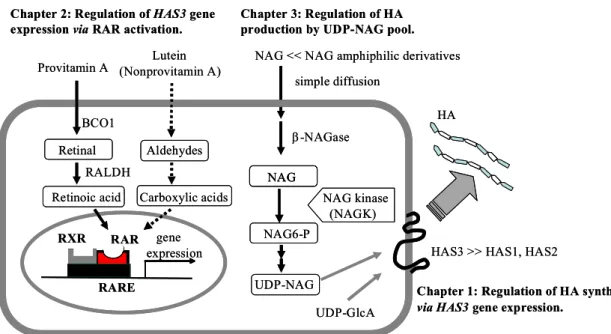
水素酵素(RALDH)の阻害剤であるシトラールにより抑制されたことから, ルテインは プロビタミンA 同様, 表皮細胞内でアルデヒド化合物に変換され, 続いて酸化反応を 受けてRA 類似体に代謝されることで生理活性を発揮させることが示唆された. 第3 章 細胞内 HA 基質プールの変動による HA 合成制御機構 表皮細胞のHA合成を高める化合物として, N-アセチルグルコサミン(NAG)を見出し た. 細胞内には複合糖質のリソソーム分解物として NAG が存在する. この NAG は, NAG キナーゼ(NAGK)により NAG6-リン酸に, 最終的には糖供与体である UDP-NAG に変換されることで, 糖タンパクや糖脂質の糖付加に再利用される.
表皮細胞でNAGK mRNA の発現を認め, さらに NAG の添加により細胞内 UDP-NAG 量が増加したことから(Fig. 3), NAG の HA 産生促進 作用は, 細胞内基質の増大によるものであることが 示された. 加えて, HA 生合成過程において基質の合 成が律速になっていると考えられた. 表皮細胞に RA と NAG を同時に作用させることにより, 相乗的 なHA 合成促進効果を認めたことから, HA のもう一 つの糖供与体である UDP-GlcA の細胞内プールは, HA 合成の律速にはなっていないことが示唆された. 次に, NAG の細胞内取り込みについても検討した. NAG の取り込みはグルコーストランスポーター (GLUT)を介するものでなく, 単純拡散により取り込まれると予想し, 細胞内で NAG へと変換可能な親油性のNAG グリコシド誘導体を合成・評価した. NAG グリコシド誘 導体はNAG に比して低濃度域で作用を示した. この結果は, NAG が単純拡散により細 胞内に取り込まれることを示唆した. これらの結果から, HA の合成調節が転写レベルだけでなく, 細胞や組織においては 細胞内基質レベルにおいても制御されることが示唆された. 以上, Fig. 4 に示すように, 表皮 HA 代謝に関わる新たな知見について記載した.
Fig. 3 Effect of NAG on UDP-NAG pools. 10 mM NAG U D P -N A G ( ng / g D N A ) 0 50 100 150 + - - Keratinocytes Fibroblasts
まず, 表皮の転写レベルにおける HA 合成の制御は主に HAS3 mRNA が担うことを 見出した. このことにより, 表皮 HA 合成制御剤開発において, HAS3 mRNA 発現を指 標にした薬剤探索が可能となることが期待される. 次に, RAR 活性化を機序とし, かつ安全性の高い HA 合成制御剤の候補として, プロ ビタミンA およびシクロヘキセン環とポリエン構造を有するノンプロビタミン A カロ テノイドを見出した. 最後に, HA 合成促進剤として細胞内基質プールを増大させる NAG を見出し, 転写 レベルによる制御だけではなく, 細胞内基質プールへ作用も HA 合成制御剤開発の重 要なストラテジーになることを示した. 膜透過を考慮した両親媒性 NAG 誘導体の開 発により, 効率的に HA 合成を亢進させることが可能なことも示した. 本研究結果の掲載 1) J. Invest. Dermatol., 118, 43-48 (2002).
2) Biosci. Biotechnol. Biochem., 77, 1282-1286 (2013). 3) Skin Pharmacol. Physiol., 17, 77-83 (2004).
simple diffusion -NAGase NAG6-P UDP-NAG NAG kinase (NAGK) NAG UDP-GlcA
HAS3 >> HAS1, HAS2 HA RARE gene expression Provitamin A Retinal Retinoic acid (Nonprovitamin A) BCO1 RALDH RAR RXR Aldehydes Carboxylic acids
Fig. 4 Regulation of HA synthesis in keratinocyte.
Chapter 1: Regulation of HA synthesis
via HAS3 gene expression.
simple diffusion -NAGase NAG6-P UDP-NAG NAG kinase (NAGK) NAG NAG UDP-GlcA
HAS3 >> HAS1, HAS2 HA RARE gene expression Provitamin A Retinal Retinoic acid (Nonprovitamin A) BCO1 RALDH RAR RXR Aldehydes Carboxylic acids
Fig. 4 Regulation of HA synthesis in keratinocyte.
Chapter 1: Regulation of HA synthesis
謝辞 34
掲載論文 35
HA: hyaluronan
HAS: hyaluronan synthase
HABP: hyaluronan binding protein GAG: glycosaminoglycan
IFN-: interferon-gamma
TGF-: transforming growth factor-beta
RA: all-trans retinoic acid RAL: all-trans retinal RAR: retinoic acid receptor
RARE: retinoic acid responsive element SEAP: secreted alkaline phosphatase BCO1:-carotene 15, 15′-monooxygenase
RALDH: retinal dehydrogenase NAG: N-acetylglucosamine GlcN: glucosamine
GlcA: glucuronic acid Glc: glucose
UDP-NAG: uridinediphosphate-N-acetylglucosamine UDP-GlcA: uridine diphosphate glucronic acid NAGK: N-acetylglucosamine kinase
-NAGase: -N-acetylglucosaminidase
諸 論 ヒアル ロン 酸 ( HA)は , -N-アセチ ルグ ル コ サ ミ ン (NAG) と -D- グ ル ク ロ ン 酸 ( GlcA) の二 糖 が 交互 に 結 合し た 高 分子 多 糖 で, 哺乳 動 物 の結 合 組 織に 広 く 存在 す るグリ コサ ミノグ リカ ン( GAG) のひ とつ である ( Schem e 1). HA の分 子量は 107 Da に も 及 び, 糖 自 体 が も つ 水 和 能 に 加 え , オー バ ーラ ッ プし た 三次 元 ネッ ト ワー ク に水 分 子を 束 縛す る こと に より, 高 い 水 和体 積 を 獲得 す る. 生 体内 で は 水和 ゲ ル とし て 存 在し , 組 織 の 水分 維 持 や機 械 特 性に関 わっ ている. 組織の HA 濃 度は , 臍 帯で 最も高 く, 皮膚 およ び硝 子体が これ に続 く. しか しな がら絶 対量 では皮 膚に 存在 するHA が 圧倒的 に多 く, 全身 の約 50 %を占め る.1 ) 皮 膚は, 乾燥 や紫 外線 などの 外部 刺激 から生 体を 守る 最前 線にあ る. 皮膚 HA は , 組 織の 水 分保 持, 弾 力 性の 維 持だ け でな く, 多様 な 皮膚 生理 に 関わ る と考 え られ る . 例え ば, HA 合 成能 を亢 進さ せ たモ デル マ ウス では 創 傷治 癒が 促 進さ れる こ と 2 ), 他方でHA を 分解除 去す ると 表皮 細胞の 最終 分化 が促進 され るこ と3 )が 報告 され て おり, HA の細 胞増 殖や 分化 への 密接 な関 係 が示 唆さ れて いる . 申 請者 らは , UVB 照射後 の皮 膚炎症 過程 にお いて, 組 織 HA 量だ けで なく , そ の分子 量サ イズ が大 き く変動 する こと を示 した.4 ) した がっ て HA は , さ まざ まな 環境 に対 して , 柔軟 に その量 と分 子サイ ズを 変化 させ, 物 理的・生 理的な 特徴 を発 揮して いる と考 えら れ る. 皮膚の 組織 中で は, 0 .5- 1.5 日 5, 6 )とい う半 減期で 新たな HA に置 換さ れて いる こ とから, そ の活 発な 合成 と分解 の平 衡が , HA の機 能発 現に 重要で ある と考 えら れ る 7). 一 方, 皮膚の HA は加 齢に 伴い減 少す るこ とが報 告さ れて いる .8 -10 ) また, 加 齢に伴 い脱 アセチ ル化 され た HA が 増加 する こと,1 1 ) さ らに 光老化 皮膚 のエ ラス ト ーシス 部位 でHA が減少 する こと 12 )が 報告 されて いる. 細 胞の 増殖・分 化と いっ た 基本的 な細 胞機能 に関 与す るHA の 量的お よび 質的 変化は , 皮膚機 能の 低下 に関 わ ると考 えら れ, HA 代 謝制 御剤は 皮膚 老化 を予 防する 機能 的素 材とし て有 望で ある . HA 代謝を 担う HA の合成 機構 は, 他の GAG の それ とは大 きく 異な る ( Scheme 2) . HA 以外 の GAG は , コ アタ ンパク を有 する プロテ オグ リカ ン分子 とし て生 合成 さ れる. す なわ ち, リボ ソー ムで 翻訳 され たコ アタ ンパ クが粗 面小 胞体 に移 行し , 次 いで, ゴル ジ複 合体 に組み 込ま れた 多くの 酵素 群に より GAG 鎖 が 伸長 , 修 飾さ れ た後 に細 胞外 へ分 泌さ れる こと にな る. 一 方, HA 生合 成の 場は 細胞 膜で あり , 合 成に必 要な 酵素は HAS のみ であ る. 膜酵 素であ る HAS は , UDP- N- アセ チル グル コ サミン(UDP- NAG)と UDP- グル クロ ン酸( UDP- GlcA)の 2 つの 糖ヌク レオ チド を基 質 として, 2 糖の 繰り 返し 配列で ある HA を 細胞 外へ直 接伸 長さ せて いく.13 ) O H H HO H H OH H COOH O HO O H H H H NH H O OH COCH3
Scheme 1. Structure of hyaluronan (HA)
O H H HO H H OH H COOH O HO O H H H H NH H O OH COCH3 O H H HO H H OH H COOH O HO O H H H H NH H O OH COCH3
HAS を コ ードす る遺 伝子 (HAS1 , HAS2 , HA S3) は, 199 6 年以 降, ヒト およ びマ ウス で相次 いで 同定さ れた.14 -18 ) ヒ トの HA S1- 3 はア ミノ 酸レ ベル で互い に 64 .9- 78. 4 % の高い ホモ ロジー を持 ち, 何れ も 6 つの 膜貫通 ドメ イン と 1 つの膜 結合 ドメ イン か ら成 る. 近 年, 真 皮線 維芽 細胞 培養 系 にお いて , 細胞 外マ トリ ッ クス( E CM ) 構築 に 重要な サイ トカイ ンの T GF-が, 主に HAS2 遺 伝子発 現誘 導を 介して HA 合 成を 促 進する こと が明ら かに され てい る.19 ) 従 来, HA は 結合 組織の E CM を構 成す る分 子 と し て 知 ら れ, 皮膚 HA の 研究 対 象 は 主 に 真 皮 で あ っ た. そ の一 方 で , 1988 年 Tamm i ら の免疫 組織 染色 より , 重 層上皮 組織 の表 皮に も HA が存在 する こと が示 さ れた.20 ) 表皮 層で は, 基 底細 胞が 増殖 ・分 化を 経て , 脱 核・ 脱顆 粒 ・辺 縁体 形成 に より バリ ア機 能を 有 する 角層 が形 成 され, 表 皮 層全 体が 1-2 カ 月で ター ン オー バ ーする こと が知ら れて いる.21 ) 表皮 基底 層, 有棘 層のみ なら ず, 角層 にも HA が 存 在する こと が報 告さ れ,2 2 ) 表皮 細胞 自身 が HA を合 成す るこ とが示 され た. 表 皮 層の乾 燥重 量当た りの HA 量は , 真皮の それ の約 2 0 %と少 ない が, 主に 構造 タン パ クに よ り構 築 され る 真皮 結 合 組織 と 異な り, 細胞 が 密接 に 隣接 す る表 皮 細胞 間 の HA 濃 度は , 約 2 m g/m l に 達す る.23 ) 細 胞間ス ペー スが 限られ てい る表 皮では, HA の量 と 質の 両 方が, 細 胞の 生 存お よ び生 理 機 能発 現 に必 要 な水 環 境の 変 化に 関 係 Core protein Proteoglycan core protein mRNA
ER Golgi HAS mRNA HAS Sulfated GAG HA Elongation of sugar chains by HAS Core protein
Scheme 2. Sugar chain elongation of sulfated GAG and HA Proteoglycan
Elongation of sugar chains by glycosyltransferases
Core protein
Proteoglycan core protein mRNA
ER Golgi HAS mRNA HAS Sulfated GAG HA Elongation of sugar chains by HAS Core protein
Scheme 2. Sugar chain elongation of sulfated GAG and HA Proteoglycan
第 1 章 HA 合 成酵 素遺 伝子 ( HAS)発 現変 動に よる HA 合 成 制御 第 1 節 :序 論 創 傷治 癒 の 過 程 は, 炎 症 , 肉 芽 形 成 , 結 合 織 化の 大 きく 3 つの 過 程 か ら 成 る. その際, 真皮 HA は 肉芽形 成時 に増 加し , 細 胞遊 走・増 殖・ 血管 新生・ コラ ーゲン 分解 お よび 再 構築 の ため の 水 和ス ペ ース の 提供 な どに 寄 与す る.2 4 ) 線 維 芽細 胞培 養系で は, IL-1, T NF-, -FGF, P DGF, IGF-1 およ び T GF-など の炎 症性 サイ トカイ ンや増 殖因 子がHA の 合成を 惹起 する .25 -27 ) ヒ ト皮膚 線維 芽細 胞で は, ECM 構築に 重要な 役割 を担うT GF-が, 主に HAS2 m RNA の発 現誘導 を介 して HA の合 成を促 進させ るこ とが示 され た.1 9 ) 一 方, 表 皮に おいて は , EGF 刺激 後の HA 合 成惹起 に, Ha s2 m RNA の 発現誘 導を 伴う ことが , ラット 由来 表皮 株化細 胞を 用い た実 験で報 告され たが,28 ) ヒト 組織 ・細胞 にお ける その制 御機 構は 未だ不 明で ある. そこで 本研 究で は, ヒト 表皮 におけ る HA 合 成制御 機構 を明 らか にす る目的 で, ヒト由 来表 皮細胞 に発 現する HAS 遺伝子 を同 定す ると ともに , サイト カイ ン添 加 による HA 合 成変 動と HAS m RNA 発 現変 動の 挙動に つい て検 討した . さらに , 表 皮 HA の合 成を 亢進 するこ とが 知ら れる レチノ イン 酸( RA)2 9 )の 応答性 につ いて も 検討し た. 第 2 節 :実 験材 料お よび 実験 方法 1- 2- 1 培養 器具お よび 試薬 以下 の試 薬, キッ ト等 は , そ れぞれ 市販 品を 購入 し使用 した .
Nor mal h um an for esk in k er atino cyt es (Kurabo 社 製) , Bo vine p it ur itary extr axt ( BPE ) (Kurabo 社 製), M CDB153 medium (Wak o P ur e Ch em ical 社 製), Recom bin ant h uman IL-1 TNF IL-8 お よび IL-10 (Intergen 社製 ), Recom bin ant h um an IFN- およ び T GF-1 (R& D 社 製), all-tran s ret ino ic acid (Sigma 社製 ), Sup erscript II (Gibco BML 社 製), GeneAmp PCR k it (Perk in- Elmer 社 製 ), Hum an gly cer aldeh y de-3- pho sp hat e dehy dro gen ase ( G3PDH) cDNA ( Clontech 社 製) , p SPT 18 ( Ro ch e 社 製), Dix igen in ( DI G) RNA labelin g k it, DI G n ucleic acid det er ction kit お よ び ainti-DI G alkalin e pho sph atase co njugat e ( Ro ch e 社 製) , Po sit iv ely ch ar ged ny lo n mem br an e ( Ro ch e 社 製) , RNaid kit ( Bio101 社製 ), Streptomyces h ya lu ro lyticu s 由 来 hy aluro nidase ( Seikagak u 社製) , D-[1 ,6-3H(N)] gluco sam ine hy dr och lo r ide (62 Ci/mmo l) (NE N Research Pr o duct s 社製) , Sepah adex G- 50 お よび Seph aro se CL-2 B (Ph arm acia 社 製) , DE -52 (Wh atman 社製) , Ch ugai Hyaluron an p late ( Ch ugai Diagno st ics Scien ce 社製) .
正常 ヒト表 皮細 胞を, 5 m g/l in sulin , 1 80 g/l hy drocort ison e, 14.1 m g/l O-ph o spho ry leth ano lam in e, 6.1 m g/l 2- am ino ethan ol, 100 n g/l E GFお よび0 .4 % (vo l/vo l) BP Eを 含有し たM CDB1 53培 地(0 .1 mM Ca2+) を 用い て24- well p lat eに播
種し, コン フル エン トの状 態に 至る まで培 養し た. 上記 培地 から E GFお よび BP E を除い た培 地を用 いて24 h 培養 した 後, サイ トカイ ンを 添加 した . サ イト カイン 存在下 でさ らに6 から2 4h培 養し た. サイト カイ ン添 加後6 h培 養し た細 胞から T otal RNAを 抽出し た. 一方 , 18 h 培養 した 細胞の 培養 上清 を採取 し、 その 上清 を 用いてHAの評 価を 実施 した . さ らに all-tran sRAにつ いて は, 添加2 4 h 後のtot al RNAおよ び培 養上清 を試 験に 供した .
角化 不溶 性膜の 評価 は, Sh imadaらの 方法30 )に従 った. 0 .25 %トリ プシ ンで 処理 した細 胞を1 .2 mM CaCl2およ び20M Calcium io noph ore含有 培地 で懸 濁した . こ
の懸濁 を37 ℃にて 2 h イン キュ ベート した 後, 2 % SDS含 有20 m M dith iothr eitol水 溶液中 で100 ℃ , 5 m in処 理し た. 角化 不溶 性膜形 成で , SD S耐性 とな った 細胞数 を血球 計算 盤を用 いて 計測 した. ヒト 皮膚 線維芽 細胞Detro it 551 (AT CC CCL100 )は , 10 % FBSを 含有 した ME M培 地を用 いて コンフ ルエ ント の状 態に至 るま で培 養した 後, Po ly( A)+ RNAを 抽出し た. 1- 2- 3 HA 合 成 能評 価およ び HA 定 量
I FN- , T GF- IL-1 , TNF-, IL-8 , IL-1 0 とと もに [3H] gluco sam ine (1 0 Ci/ml) を
表皮細 胞培 養系に 添加 し, 1 8 h 培養 した . HA 合成 能は , 培 養上 清 の hy aluron idase 感受 性画 分へ の 取り 込み 量を 評 価し た.3 1 ) 培養 上清 を h yaluron idase (1 .5 T RU/m l) で消化 した 後, Sephadex G- 50 カ ラム (1. 5 ×5 cm)に アプ ライ し, 分取 した v oid 画 分 の放射 活性 を測定 した. hy alur on idase 未 処理 および 処理 サン プルの 放射 活性 差を 算 出し, HA への 取り 込み 量とし た . HA 産生 量は , ヒ アル ロン 酸結合 タン パク を利 用 した Ch ugai hy aluron an p lat e を 用い , プ ロト コール に従 い評 価した 。
1- 2- 5 HAS3 cDNA の 単 離
ヒ ト HAS3 遺伝 子 cDNA は , RT-P CR およ び secon d P CR によ り単 離した . まず , ヒ ト 表 皮 細 胞 か ら 抽 出 し た Total RNA を 用 い , m RNA の シ ー ケ ン ス ( Gen Bank accessio n n um ber AF232 772) を元 に, 下記 のプ ライマ ーを 設計 し, RT-P CR を 行っ た. Hum an HAS3 sen se pr im er 5’ - GGAAAGCT T GGCAT GT ACCGCAACAG- 3’ ; Hum an HAS 3 ant isen se pr im er 5’ -AGAGGAGGGAGTAGAGGGAC -3 ’ (30 cycle) . 次 に, 得ら れた RT-PCR 産 物を アガ ロー ス電気 泳動 に供 し, ゲル から回 収し たサ イズフ ラク ション を鋳 型とし て, secon d P CR を下 記の pr im er を 用い て行っ た. Human HAS3 sen se pr im er 5’ - GGAAAGCT T GGCAT GGACC GCAACAG-3’ ; Hum an HAS 3 ant isen se primer 5’ -AGGGAAT T CGGAAGCAGGCGT AGGT G- 3’ (30 cy cle, 下 線部は HindI II お よびEco RI 制限 酵素 サイト ). 得 られ たP CR 産 物を アガロ ース ゲル 電気 泳動の 後, ゲルか ら切 りし精 製し た. 精製 した cDNA を Hin dII I/E coR I で 消化し , プラス ミド ベク ター (p SPT 18) HindI II /E co RI サイ トに 組み 込み , シー ケシ ング によ り, 目的 とする HAS3 cDNA が得 られた こと を確 認し た. DI G 標 識した ant isen se/sen se RNA プロー ブの 調製は, DI G RNA labelin g kit プロ トコー ルに 従い 実施 した.
1- 2- 6 ノザ ンブロ ット 解析
細胞よ り抽 出し た Total RNA を 0. 8 % fo rm aldehy de/agaro se ゲルを 用い た電 気泳 動 によ り 分離 し, ナイ ロ ンメ ン ブレ ン に転 写 し た. ハ イブ リ ダ イゼ ー ショ ン は DI G Nucleic Acid Detect ion Kit のプ ロト コール に従 い実 施し , 化 学発 光法 により 検出 し た.
1- 2- 7 統計 処理
統計解 析は SAS シ ステ ムによ り実 施し た. Dun nett's m ultiple comp ar ison test でコン トロ ールと の有 意性 を解 析し, 用量 依存 性の 評価 には William’s test を 用いた. p 値 0 .05 未満 を統 計的に 有意 とし た.
第 3 節 :実 験結 果
1- 3- 1 サ イ トカイ ンの 表皮 細胞 HA 合 成 に及 ぼす 影響
Co
ntrolIL-1TNF-IL-8 IL-10IFN-TGF-
3H d p m × 1 0 -5/w el l 0 2 4 6 8 * * Co
ntrolIL-1TNF-IL-8 IL-10IFN-TGF-
3H d p m × 1 0 -5/w el l 0 2 4 6 8 * * Co
ntrolIL-1TNF-IL-8 IL-10IFN-TGF-
3H d p m × 1 0 -5/w el l 0 2 4 6 8 * * Co
ntrolIL-1TNF-IL-8 IL-10IFN-TGF-
3H d p m × 1 0 -5/w el l 0 2 4 6 8 Co
ntrolIL-1TNF-IL-8 IL-10IFN-TGF-
Co
ntrolIL-1TNF-IL-8 IL-10IFN-TGF-
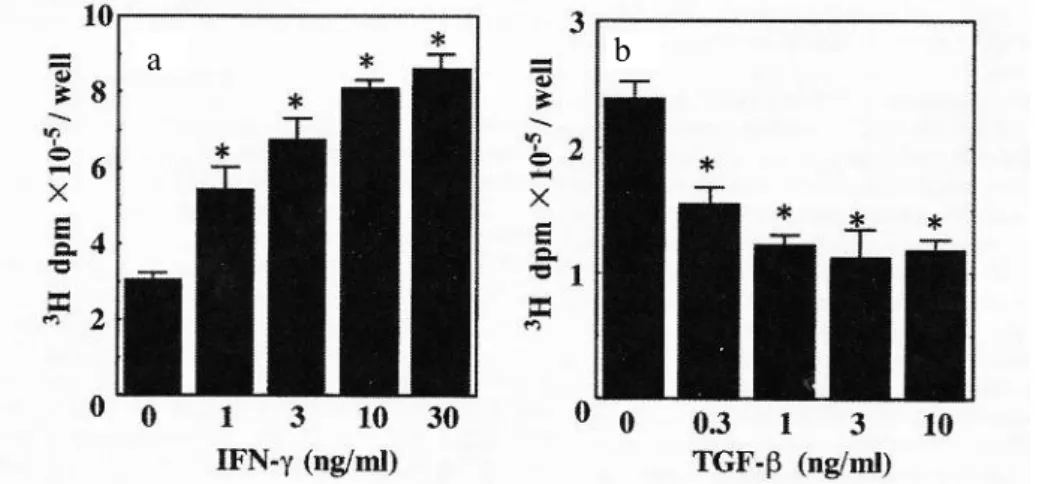
3H d p m × 1 0 -5/w el l 0 2 4 6 8 * * さらに, I FN-お よび T GF-の作 用に つい て用 量に対 する 応答 性を 検討 した とこ ろ, I FN-は 添加 1-10 n g/m l ま で用 量依 存的に HA 合成を 促進 した . 一方で , T GF- は添加0.3 n g/m l 以 上で HA 合 成を 抑制 し, 1.0 n g/m l 以 上の 添加濃 度で は , そ の作 用 が飽和 した(Fig. 1 -2) . 両サイ トカ イン 添加に より 培養 上清 中に蓄 積さ れる HA の 絶 対量を 定量 し (Table 1 -1) , IFN-お よび T GF-が実 際に HA 産生量 を変 動さ せて い るこ とを 確認 した. また , 本培 養条 件下 にお い て I FN- と T GF- は細 胞数 およ び 角化不 溶性 膜形成 に影 響を 及ぼ さず(Table 1-2), こ れら のサ イトカ イン の HA に 及 ぼす効 果は, 増殖お よび 分化 に依 存した 現象 では ないこ とが 示さ れた .
Fig. 1-1 Effect of cytokeines on hyaluronan synthesis in cultured human keratinocytes. Conflurent human keratinocytes were incubated in the absence or presence of 10 ng/ml IL-1, TNF-, IL-8, IL-10, IFN-, or TGF- with [3H]glucosamine. The
incorporation of [3H]glucosamine into hyaluronan
during 18 h incubation period was determined. Each column represents the mean± SD of 4 separate experiments. *Significantly different from the control value; p < 0.01.
a b
a b
Fig. 1-2 The hyaluronan synthesis of human keratinocytes was upregulated by IFN - but down regulated by TGF- in a dose dependent manner. Conflurent human keratinocytes were incubated with fresh medium for 24 h followed by cultivating with [3H]glucosamine and various concentrations of
(a) IFN- (1-30 ng/ml) or (b) TGF- (0.3-10 ng/ml) for 18 h. The incorporation of [3H]glucosamine into
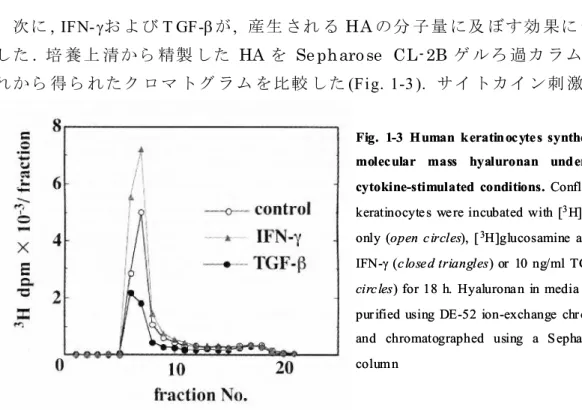
次に, IFN-お よび T GF-が, 産生 され る HA の分 子量 に及 ぼす効 果に つい て検 討 した. 培 養上 清から 精製 した HA を Se pharo se CL- 2B ゲ ルろ 過カ ラム に供し , そ れから 得ら れたク ロマ トグ ラム を比較 した(Fig. 1-3 ). サイ トカイ ン刺 激お よび
Fig. 1-3 Human keratinoc yte s synthesized high-molec ular mass hyaluronan under basal or cytokine-stimulated conditions. Conflurent huma n keratinocyte s we re incubated with [3H]glucosamine
only (open c ircles), [3H]glucosamine and 10 ng/ml
IFN-closed triangles) or 10 ng/ml TGF- (closed circ les) for 18 h. Hyalurona n in media fra ction wa s purified using DE-52 ion-exchange chromatography and chromatographed using a Sepharose CL-2B column
Fig. 1-3 Human keratinoc yte s synthesized high-molec ular mass hyaluronan under basal or cytokine-stimulated conditions. Conflurent huma n keratinocyte s we re incubated with [3H]glucosamine
only (open c ircles), [3H]glucosamine and 10 ng/ml
IFN-closed triangles) or 10 ng/ml TGF- (closed circ les) for 18 h. Hyalurona n in media fra ction wa s purified using DE-52 ion-exchange chromatography and chromatographed using a Sepharose CL-2B column
aKeratinocyte s were grown in the sam e conditions a s for Fig. 1-2. The number of cornified cells
forme d in response to calcium ionophore we re estimated as described in Materials and Methods. The results are the means of 4 wells±SD.
Table 1-2. Effect of IFN-and TGF- on total cell and cornified cell number Treatment
IFN- TGF-
Concentration (ng/ml)
Total cell number (×10-5 cells/well) Control 0 1 10 1 10
Cornified cell numbera
(×10- 5cells/well) 4.86±0.34 4.56±0.57 4.48±0.71 4.38±0.66 4.58±0.46 1.33±0.25 1.56±0.18 1.18±0.18 1.32±0.11 1.30±0.21
aKeratinocyte s were grown in the sam e conditions a s for Fig. 1-2. The number of cornified cells
forme d in response to calcium ionophore we re estimated as described in Materials and Methods. The results are the means of 4 wells±SD.
Table 1-2. Effect of IFN-and TGF- on total cell and cornified cell number Treatment
IFN- TGF-
Concentration (ng/ml)
Total cell number (×10-5 cells/well) Control 0 1 10 1 10
Cornified cell numbera
(×10- 5cells/well) 4.86±0.34 4.56±0.57 4.48±0.71 4.38±0.66 4.58±0.46 1.33±0.25 1.56±0.18 1.18±0.18 1.32±0.11 1.30±0.21 Table 1-1. Effect of IFN-and TGF- on hyaluronan production
Treatment IFN- TGF- Concentration (ng/ml) Hyaluronan productiona (ng/ml) Control 0 1 10 1 10 126±9 185±34* 473±14 * 64±5 * 64±4 *
aK eratinocytes were grown in the same condition as for F ig. 1-2. Hyaluronan content was measured as
described in M aterials and Methods. The data represent the mean±SD of 4 separate experiments. *Significantly different from the control value; p < 0.01.
Table 1-1. Effect of IFN-and TGF- on hyaluronan production Treatment IFN- TGF- Concentration (ng/ml) Hyaluronan productiona (ng/ml) Control 0 1 10 1 10 126±9 185±34* 473±14 * 64±5 * 64±4 *
Table 1-1. Effect of IFN-and TGF- on hyaluronan production Treatment IFN- TGF- Concentration (ng/ml) Hyaluronan productiona (ng/ml) Control 0 1 10 1 10 126±9 185±34* 473±14 * 64±5 * 64±4 *
aK eratinocytes were grown in the same condition as for F ig. 1-2. Hyaluronan content was measured as
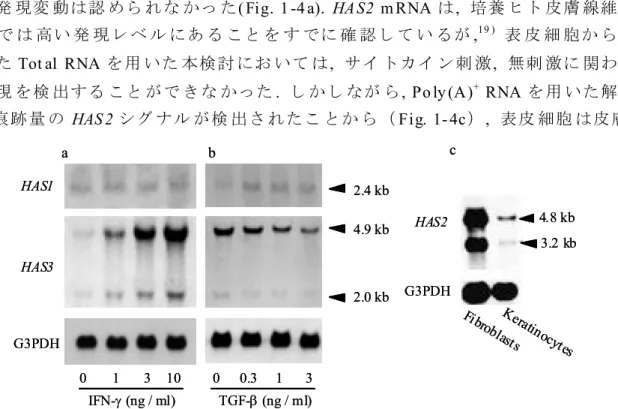
無刺激 のい ずれの 場合 も産 生さ れた HA は主 に vo id 画分 に溶 出し た. つま りそ れ らの分 子量は 1×106 Da より も大き いこ とを 示し ており , I FN-およ び T GF-は , い ずれも 高分 子 HA の産 生を 誘導す るこ とが わかっ た . 1- 3- 2 IFN- お よび TG F- の HAS 遺 伝 子 発 現に及 ぼす 影響 表皮 細胞の HA 合 成が これ まで報 告さ れて いる HAS ファ ミリー のい ずれ によ り 調節さ れて いる かを 調べ るた め, IFN-お よび T GF-で刺 激し た表 皮細 胞を 用い ノ ザンブ ロッ ト解析 を実 施し たHAS3 m RNA には約 4.9 k b の主 な転 写産 物と約 2 .0 k b の異 なる サイ ズの 転 写産 物が 存在 した. IFN-の 刺激 に より 両シ グナ ルの 強 度が 顕 著に上 昇し た ( Fig. 1-4 a) . 一 方, T GF-の添 加し た用量 に応 じて HAS3 の転 写レ ベ ルは減 少し た( Fig. 1-4 b). この HAS 3 m RNA の顕著 な発 現変 動に 対し, 2.4 k b の HAS 1 mRNA は T GF-刺激 後に わずか に発 現量 が上昇 する もの の(Fig. 1 -4 b), IFN- 添加 に よる発 現変 動は認 めら れな かっ た( Fig. 1 -4 a). HA S2 m RNA は, 培養 ヒト 皮膚 線維 芽 細胞 で は 高い 発 現レ ベ ル にあ る こと を す でに 確 認し て い るが,19 ) 表 皮 細 胞か ら 抽 出したTot al RNA を用 いた 本検討 にお いて は, サイ トカイ ン刺 激, 無刺 激に 関わ ら ず発現 を検 出する こと がで きな かった. し かし なが ら, Po ly (A)+ RNA を用 いた解 析 で, 痕跡量 の HAS 2 シグ ナル が検 出さ れたこ とか ら( Fig. 1- 4c), 表皮 細胞 は皮膚 線維芽 細胞 に比し て極 めて 低レ ベルで はあ るも のの, HAS2 m RNA を発 現し てい る 0 1 3 10 IFN-(ng / ml) TGF-(ng / ml) 0 0.3 1 3 HAS1 HAS3 G3PDH b a 2.4 kb 4.9 kb 2.0 kb 3.2 kb 4.8 kb Fibro
blastKeras tinocytes HAS2 G3PDH c 0 1 3 10 IFN-(ng / ml) TGF-(ng / ml) 0 0.3 1 3 HAS1 HAS3 G3PDH b a 2.4 kb 4.9 kb 2.0 kb 3.2 kb 4.8 kb Fibro
blastKeras tinocytes HAS2
G3PDH c
Fig. 1-4 The HAS3 transcript level of human keratinocytes was upregulated by IFN- but downregulated by TGF- in a dose-dependent manner. Human keratinocytes were cultured in various concentrations of IFN-(0-10 ng/ml) (a) or TGF- (0.3-3 ng/ml) (b) for 6 h and total RNA was extracted. Equal amount of samples (5 g/lane) were hybridized with HAS1 and HAS3 probes. (c) Poly(A)+ RNA extracted from confluent cells was used for northern blot analysis. Equal amounts of
ことが わか った. 以上 の結 果から, I FN-およ び T GF-の 刺激に よる 表皮 細胞の HA 合成 変動は , 主 にHAS3 m RNA の発 現変動 を介 して いる と考え られ た. 1- 3- 3 レ チ ノイン 酸 の HAS 遺 伝 子 発現 に及 ぼす 影響 表皮 HA 合 成を 惹起 する こと がすで に報 告さ れて いる レチ ノイ ン酸 ( RA)29 )を用 いて, RA が培 養表皮 細胞 の HA 合成 と HAS 遺伝子 発現 に及 ぼす効 果に つい て検 討 した. RA は, HAS3 m RNA の発 現を強 力に 誘導 する ととも に, HA 合 成を 顕著 に亢 進 さ せ た( Fig. 1 -5) . I FN- に よ る 応 答 と 同様 , HAS1 m RNA に 顕 著 な 誘導 を 認 め ず , HAS 2 m RNA は 本条件 では 検出 でき なかっ た.
第 4 節 :考 察
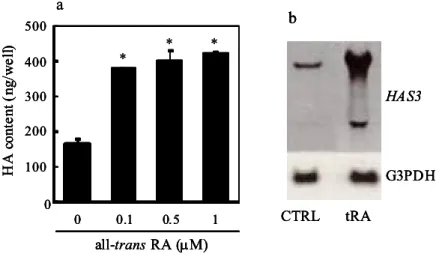
HAS に は 3 種 のア イソ ザイム が存 在す る ( HAS1, HAS2 , HAS3 ). 本研 究では , ヒ ト表皮 細胞の HA 合成 を惹 起する サイ トカ インと して I FN-, 逆に 抑制 する サイ ト カイン とし てT GF-を 見出 した . こ れら サイ トカ インに よる HA 合 成挙 動と 一致 し た発現 挙動 を示 す HAS 遺 伝子 が HAS3 m RNA で ある ことも 明ら かに した . さ らに , サイト カイ ン以 外の 表 皮 HA 合成 促進 因子 とし て, RA に つい て検 討し たと ころ , RA は , IFN-と 同様顕 著に HAS3 m RNA 発現量 を増 加さ せるこ とを 見出 した .
真 皮線 維芽 細胞 でド ミナ ン トに 発現 し, か つ HA 合 成を 主に 調節 する 合 成酵 素 0 100 200 300 400 500 0 0.1 0.5 1 H A c ont ent ( ng /w el l) all-trans RA (M)
Fig. 1-5 The HAS3 transcript level of human keratinocytes was upregulated by all-trans RA. (a) Human keratinocytes were cultured in various concentrations of all-trans RA (0-1.0 M) for 24 h and hyaluronan production was measured as described in Materials and Methods. The data represent the mean±SD of three separate experiments. *Significantly different from the control value; p < 0.01. (b) Keratinocytes were incubeted for 24 h in the presence of all-trans RA at a concentration of 1.0 M. Total RNA was extracted and subjectied to northern blot as described in Materials and Methods. Similar results were obtained in two independent experiments.
a HAS3 G3PDH CTRL tRA b * * * 0 100 200 300 400 500 0 0.1 0.5 1 H A c ont ent ( ng /w el l) all-trans RA (M)
Fig. 1-5 The HAS3 transcript level of human keratinocytes was upregulated by all-trans RA. (a) Human keratinocytes were cultured in various concentrations of all-trans RA (0-1.0 M) for 24 h and hyaluronan production was measured as described in Materials and Methods. The data represent the mean±SD of three separate experiments. *Significantly different from the control value; p < 0.01. (b) Keratinocytes were incubeted for 24 h in the presence of all-trans RA at a concentration of 1.0 M. Total RNA was extracted and subjectied to northern blot as described in Materials and Methods. Similar results were obtained in two independent experiments.
a HAS3 G3PDH CTRL tRA b 0 100 200 300 400 500 0 0.1 0.5 1 H A c ont ent ( ng /w el l) all-trans RA (M) 0 100 200 300 400 500 0 0.1 0.5 1 H A c ont ent ( ng /w el l) all-trans RA (M)
Fig. 1-5 The HAS3 transcript level of human keratinocytes was upregulated by all-trans RA. (a) Human keratinocytes were cultured in various concentrations of all-trans RA (0-1.0 M) for 24 h and hyaluronan production was measured as described in Materials and Methods. The data represent the mean±SD of three separate experiments. *Significantly different from the control value; p < 0.01. (b) Keratinocytes were incubeted for 24 h in the presence of all-trans RA at a concentration of 1.0 M. Total RNA was extracted and subjectied to northern blot as described in Materials and Methods. Similar results were obtained in two independent experiments.
遺伝 子が HA S2 であ る 19 )のに 対し, 表皮 細胞 で主 に発 現し てい る遺 伝子 は HAS 3 mRNA で あった . このこ とか ら, 皮膚 組織 の表 皮と真 皮で は, HAS の発 現様 式が 異 なると 考え られた. 線維芽 細胞 で HAS2 m RNA 発現を 誘導 する T GF-は , 表 皮細 胞 では HAS 3 m RNA 発 現を 抑制し , HA 合 成能 を低 下させ た. 真皮 にお ける T GF-は , 創傷 治 癒時 の 真皮 マ トリ ッ クス 再 構築 に 中心 的 な役 割 を担 い, 線 維芽 細 胞の 増 殖 を促進 する.32 , 33 ) 一方, 表皮 では , 基 底層 から 有棘層 にか けて T GF-が 恒常 的に 発 現し て お り,3 4, 3 5 ) 表 皮 細 胞増 殖 に対 し, 抑 制 的に 働 く .3 6 ) 同 一 のサ イ ト カイ ン が 表皮と 真皮 で逆 の HA 合成 制御 を担 うこ とは , 応答 性の 異な るア イソ ザイ ムの 存 在を 示唆 する もの であ る. ま た , そ れ ぞれ の細 胞に 対す る T GF- の異 なる の 合 成制 御は組織 の再 構築 , ある いは 細胞 増殖 の際 の水 和ス ペー スの 確保 など の HA の複雑 な役 割を担 う応 答性 と考 えられ る. I FN-は , 活性化 T 細胞 から放 出さ れる 代表的 なサ イト カイン であ り , そ れ自 体 は表 皮細 胞の 増 殖を 抑制 す る.3 7 ) そ の 一方 で細 胞 増殖 を促 進す る T GF-の誘 導 等 を介 し て 表 皮 過 増殖 を 伴う さ まざ ま な 病変 に も関 与 す る. 急性 湿 疹に お け る 病 理 学 的 特 徴 の 表 皮 海 綿 状 浮 腫 の 形 成 に 関 与 す る サ イ ト カ イ ン の 一 つ と し て IFN- が知 られ てお り, 急性 湿疹 の病 変部に おい て HAS 3 m RNA の 顕著 な発 現上 昇 と E-カ ドヘ リン の減少 が示 され た.4 0 ) 海 綿状 浮腫で の HA の 病態生 理的 な役 割は , 表皮 細 胞間 ス ペー ス を広 げ る こと で 細胞 間 接着 を 減弱 さ せる こ とで あ る. し た が って海 綿状 組織に おけ るHA 量 の増 加は, 有 害化合 物の 濃度 を低下 させ るた めの 生 体適応 過程 である と推 察さ れる.
第 2 章 レ チノ イン 酸レ セプ ター を介 した HA 合 成 制御 第 1 節 :序 論 第 1 章 にお いて , RA は, HAS3 遺伝 子発 現を誘 導し , 表 皮細 胞の HA 合 成を促 進 させる こと を示し た. RA は ビタ ミン A の 活性 体で あり , 細 胞の分 化増 殖の 制御 な ど重要 な生 理活 性を 有す る. さら に尋 常性 乾癬 , 一部 角化性 疾患 , 尋常 性座 瘡お よ び悪性 腫瘍 の薬物 治療 にも 用い られる. 美 容面 でも in vivo お よび iv vitro 両 面か ら 多く の 研究 が なさ れ, 欧米 で はシ ワ 等の 光 障 害皮 膚 の改 善 剤と し て認 可 され て い る 42). しかし なが ら, RA は その 効果だ けで なく , 副 作用 として 皮膚 刺激 , 落屑 性 紅斑お よび 掻痒 等を 引き 起こ す. T adak i ら は, RA がシ ワに改 善効 果が ある 一方 , 特に 日本 人皮 膚 に対 する 刺 激性 が高 いこ と を指 摘し てい る.43) こ のこ とか ら, RA にかわ る安 全性が 高く, か つ有 効な 改善剤 が望 まれ てい る. RA の 前駆 体は, カロ テンお よび ク リプ トキサ ンチ ンに 代表さ れる 食餌 性の プロビ タミ ン A カロ テノ イドで ある . プ ロビ タミ ン A は小 腸上皮 細胞 に吸 収さ れ た後, カ ロテ ン-15,1 5’ -モ ノオ キシ ゲナ ー ゼ(BCO1) によ り 中央 開裂 して レ チナ ー ルに変 換, さら にレ チナー ルデ ヒド ロゲナ ーゼ ( RAL DHs)によ り酸 化さ れ, RA を 生 成する.44 ) all- tran s RA は, 核内 レセ プタ ーで あるレ チノ イン 酸レ セプ ター( RARs) と結合 し, 9- cis RA レセ プタ ーの レチノ イン 酸 X レセプ ター ( RXRS) とヘ テロ ダイ マ
ーを形 成し て DNA のレ チノ イン 酸応答 配列 ( RARE) に結 合し てそ の下 流の 遺伝 子 発現を 制御 する. 近 年, HAS2 遺伝 子は RAR シグナ ルの 直接 のター ゲッ ト遺 伝子 で あるこ とが 報告さ れた.45 ) しか し一 方で, RA の HA 合 成促 進作 用に, extracellular
sign al-regulated kinase 1/2 (E RK1 /2) 活性 化を 介した 二次 的な 応答が 関与 する こと が 示唆さ れて いる.46 ) 本研究 では, 経 口摂 取によ り RA へ変換 され るプ ロビタ ミ ン A カロ テノ イド が , 表皮細 胞の RAR シグナ ルを 介して HA 合成 を誘導 する か否 かにつ いて 検討 した . さらに 哺乳 類では レチ ノイ ドへ 変換さ れな いノ ンプロ ビタ ミン A カ ロテ ノイド 47 ) の RAR 活性 化を 介し た HA 代 謝に 及ぼ す効果 につ いて も検 討した . 第 2 節 :実験 材料 およ び実 験方 法 2- 2-1 実 験器 具お よび 試薬
so lut ion (Mo ss 社製 ), T RI zo l r eagent (In vit ro gen 社 製) , T ran scr iptor Fir st Str an d cDNA Synth esis k it ( Ro she), p RARE -TA- SE AP お よび p SE AP2 - basic v ecto r お よびE scApe SE AP chem iluminescence detect ion kit ( Clont ech 社製 ), Fugen e 6 tranf ectio n r eagent ( Ro she).
その他 の試 薬につ いて は, 第 1 章で 記載し たも のを 用い た.
2- 2- 2 細胞 培養
ヒト表皮 細胞 は第 1 章で記 載し た方 法に従 い培 養し た. 24- well プレ ート およ び 12- well プ レー トに播 種し た細 胞がコ ンフ ルエ ント に達し た後 , 0. 04 %(v /v ) BPE を 含有し, hy dro cort ison e お よび E GF を除去 した 培地 に変換 し, 評価 化合 物を 添加 し た. HA 産 生量 の評価 は 24 - well プ レート の培 養上 清を用 い, 細胞 画分 は RT-P CR に よる評 価に 用いた. 12- well プレー トの 細胞 は, レポ ーター アッ セイ に用 いた.
2- 2-3 RT- PCR 法
表皮 細胞か ら T RIzo l 試薬 によ り Total RNA を 抽出し , 逆転写 反応 は T ran scr ipto r Fir st Str an d cDNA Synth esis k it プロト コー ルに 従い実 施し た. HAS 3, G3PDH, BCO1 , RAL DH1, RALDH2 およ び RALDH3 m RNA 検出 に用 いる P CR プライ マー セッ トは , primer 3 を利 用し , エ キソ ンジャ ンク ショ ンに設 計し た.
HAS 3 sen se: 5’ - CT GCACCT GCT CAT T CAGAG- 3’ , HAS 3 anti- sen se: 5’- GAGT CGCACACCT GGAT GT A-3 ’, G3PDH sen se: 5 ’- ACCACAGT CCAT GCCAT CAC-3 ’, G3PDH ant i- sen se: 5’ -T CCACCACCCT GT T GCT GTA- 3’, BCO1 sen se: 5 ’- ACAGAGAT GT GAAGGAGGGAAG- 3’ , BCO1 ant i- sen se: 5’ -T CCAAACT CAGACACCACAAT C-3 ’, RAL DH1 sen se: 5’ -CTCTGCCAGG TAGAAGAAGGAG-3 ’, RAL DH1 ant i- sen se: 5’ - GT GGAGA GCAGT GAGAGGAGT T-3 ’ RAL DH2 sen se: 5 ’- AGAACT CAGAGAGT GGGAGAGT GT-3 ’ RAL DH2 ant i- sen se: 5’ -AT GTAGAGT GCACT GAGT GGT GT T-3’ RAL DH3 sen se: 5 ’- GGA GCAGGT CTACT CT GAGT T T GT-3 ’ RAL DH3 ant i- sen se: 5’ -AAGCGTAT T CACCTAGT T CT CT GC-3 ’
2- 2- 4 HA 定 量
培養 上 清HA の定 量は , hy aluro nan bin din g protein ( HABP )を用 い た Rilla ら4 8 )の サ
ンドイ ッチ アッセ イ法 に従 った. 96 well プレ ート に HABP をコ ーテ ィン グし, 標 準 サ ンプ ル お よ び評 価 サ ン プ ルを ア プ ラ イし た. 室 温 にて 2 h 静 置 の 後, EZ -L in k maleim ide act iv ated hor ser adish p er ox idase kit を用 いて ペル オキシ ダー ゼ標 識し た HABP ( HRP- HABP)を 反応 させ, サ ンドイ ッチ を形 成させ た. 次に , HRP の基 質で あ る3,3 ',5, 5'-tetr am ethy lbenzidin e (T M B) substr ate so lution を 添加 し, 450 n m の 吸光 度 を測定 した. サ ンプ ル中 の HA 濃度 は, 標準 サンプ ルに より 作成し た検 量線 を用 い て算出 した.
2- 2- 5 DNA 定 量
DNA 量の 定量 は, John son -Wint ら 4 9 )に よる 蛍光 試薬( 3, 5- diam ino ben zo ic acid)
(DABA)を 用い た手法 で実 施し た. 24- well プ レー ト上 の細 胞を 8 0 % 冷 エタ ノー ル で固定 した 後, DABA ( 400 m g/m l)を添 加し , 60 ℃, 45 m in イン キュ ベー トした . 反 応後, 1 N HCl を 0 .5 m l 添 加し, 蛍 光強度 (励 起波 長:42 0 nm, 蛍光 波長 :540 n m) を 測定し た. 標準 サン プルに より 作成 した検 量線 より DNA 量を 算出 した .
2- 2- 6 レポ ーター アッ セイ 法
RARE ト ラン ス活 性化の 評価 は, 2 コ ピー の RARE シー ケンス およ び Secr eted alkalin e pho sphat ase ( SE AP) レポー ター 遺伝 子が 組み込 まれた p RARE-TA- SE AP ベ クタ ーを 用い た. ま た バッ クグ ラン ド 測定 用は プロ モ ータ ーを 欠 く p SE AP2- basic ベクタ ーを 用いた. 1 2- well プレ ートに 培養 した 表皮細 胞に , Fugen e 6 トラ ンス フ ェク シ ョン 試 薬を 用 いて そ れ ぞれ の ベク タ ーを 導 入し た. トラ ン スフ ェ クシ ョ ン 24 h 後 に ル テイ ン を添 加 し, さらに 2 4 h 培養 し た. 培 養 上 清の SE AP 活性 は , EscAp e SE AP chem iluminescence detect ion システ ムを 用い て評 価した .
2- 2- 7 統計 処理
チン だけ でな く, ノン プロ ビタ ミ ン A に 属す るル テイ ン, ゼア キ サン チン およ び アスタ キサ ンチン の存 在下 で亢 進した( Fig. 2-1 a) . 一 方, リコ ピン , フィト エン , -イオノ ンお よび--イオノ ンに はそ の効 果を認 めな かっ た( それぞ れの 処理 群の HA 産生量, 254 , 1 83, 27 4, 274 n g/well v s コ ント ロー ル群 の HA 産 生量 , 288 n g/m l) . こ れらの こと から, HA 合 成を 促進す るに はシ クロヘ キセ ン環 とポリ エン 部の 両方 を 併せ持 つ構 造が必 要と 考え られ た. これ らカ ロテ ノイド による HA 合成 変動 の挙 動 は, 誘導さ れる HA S3 遺 伝子 の発 現挙 動と一 致し た( Fig. 2- 1 b) .
Fig. 2-1 Effects of carotenoids on hyaluronan synthesis in keratinocytes. Confluent human keratinocytes were incubated for 48 h in the absence or presence of -carotene, -cryptoxanthin, lutein, zeaxanthin, and astaxanthin at a concentration of 5 M. (a) Hyaluronan content was measured as described in Materials and Methods. Values represent mean±SD of 3 replicate assays. Significantly different from the control value; *P < 0.05, ***P < 0.001 (Dunnett’s test). (b) Total RNA was extracted and subjected to RT-PCR. The PCR products were visualized in a 2% agarose gel stained with ethidium bromide.
-C
arotene Lutein Zeaxant hin -C rypto xanthin Asta xanthin Control H ya lu ro na n co nt en t ( ng /w el l) a b 0 200 400 600 800 1000 1200 -C
arotene LuteinZeaxanth i n -C rypto xanthin Astaxan thin Control *** *** *** *** * HAS3 G3PDH
Fig. 2-1 Effects of carotenoids on hyaluronan synthesis in keratinocytes. Confluent human keratinocytes were incubated for 48 h in the absence or presence of -carotene, -cryptoxanthin, lutein, zeaxanthin, and astaxanthin at a concentration of 5 M. (a) Hyaluronan content was measured as described in Materials and Methods. Values represent mean±SD of 3 replicate assays. Significantly different from the control value; *P < 0.05, ***P < 0.001 (Dunnett’s test). (b) Total RNA was extracted and subjected to RT-PCR. The PCR products were visualized in a 2% agarose gel stained with ethidium bromide.
-C
arotene Lutein Zeaxant hin -C rypto xanthin Asta xanthin Control H ya lu ro na n co nt en t ( ng /w el l) a b 0 200 400 600 800 1000 1200 -C
arotene LuteinZeaxanth i n -C rypto xanthin Astaxan thin Control *** *** *** *** * HAS3 G3PDH
Fig. 2-1 Effects of carotenoids on hyaluronan synthesis in keratinocytes. Confluent human keratinocytes were incubated for 48 h in the absence or presence of -carotene, -cryptoxanthin, lutein, zeaxanthin, and astaxanthin at a concentration of 5 M. (a) Hyaluronan content was measured as described in Materials and Methods. Values represent mean±SD of 3 replicate assays. Significantly different from the control value; *P < 0.05, ***P < 0.001 (Dunnett’s test). (b) Total RNA was extracted and subjected to RT-PCR. The PCR products were visualized in a 2% agarose gel stained with ethidium bromide.
-C
arotene Lutein Zeaxant hin -C rypto xanthin Asta xanthin Control H ya lu ro na n co nt en t ( ng /w el l) a b 0 200 400 600 800 1000 1200 -C
arotene LuteinZeaxanth i n -C rypto xanthin Astaxan thin Control *** *** *** *** * HAS3 G3PDH -C
arotene Lutein Zeaxant hin -C rypto xanthin Asta xanthin Control H ya lu ro na n co nt en t ( ng /w el l) a b 0 200 400 600 800 1000 1200 -C
arotene LuteinZeaxanth i n -C rypto xanthin Astaxan thin Control *** *** *** *** * HAS3 G3PDH -Carotene -Cryptoxanthin Lycopene phytoene -Ionone -Ionone Astaxanthin Lutein Zeaxanthin -Carotene -Cryptoxanthin Lycopene phytoene -Ionone -Ionone Astaxanthin Lutein Zeaxanthin
表 皮細 胞が プ ロビ タ ミ ン A か ら RA に至 る 代謝 経 路を 有 する かを 調 べる た め , BCO1 およ び RALDHs の発現 を転 写レ ベル で検討 した . 表皮 細胞は BCO 1 mRNA を発現 し, RALDHs( RALDH1 -3) のう ち RALDH3 をド ミナ ントに 発現 した (Fig. 2-3 ).
ノンプ ロビ タミ ン A が どのよ うな 機構 で HA S 遺伝 子発 現を 介し た HA 合成 を惹 RA LD H 1 RA LD H 2 RA LD H 3 RALDHs keratinocytes testis ker atin ocy tes fibr
oblastskidney stomach
BCO1
Fig. 2-3 BCO1 mRNA and RALDH3 mRNA are expressed in human keratinocytes. Total RNA was subjected to RT-PCR. The PCR products were visualized in a 2% agarose gel stained with ethidium bromide.
RA LD H 1 RA LD H 2 RA LD H 3 RALDHs keratinocytes testis ker atin ocy tes fibr
oblastskidney stomach
BCO1 RA LD H 1 RA LD H 2 RA LD H 3 RALDHs keratinocytes testis RA LD H 1 RA LD H 2 RA LD H 3 RALDHs keratinocytes testis ker atin ocy tes fibr
oblastskidney stomach
BCO1 ker atin ocy tes fibr
oblastskidney stomach
BCO1
Fig. 2-3 BCO1 mRNA and RALDH3 mRNA are expressed in human keratinocytes. Total RNA was subjected to RT-PCR. The PCR products were visualized in a 2% agarose gel stained with ethidium bromide.
0 2 4 6 0 0.3 1 3 5 Lutein (M) D N A c on te nt ( g/ w el l)
c
Fig. 2-4 The hyaluronan synthesis of human keratinocytes was upregulated by lutein in a dose dependent manner. Confluent human keratinocytes were incubated with various concentrations of lutein (0.3-5 mM) for 48h. (a) Hyaluronan content was measured as described in Materials and Methods. Values represent mean±SD of 3 replicate assays. Significantly different from the control value; **P < 0.01 (William’s test). (b) Total RNA was extracted and subjected to RT -PCR. The PCR products were visualized in a 2% agarose gel stained with ethidium bromide. (c) DNA content was measured as described in Materials and Methods. Values represent mean± SD of 3 replicate assays.
HAS3 G3PDH b 0 0.3 1 3 5 Lutein (M)
a
0 400 800 1200 0 0.3 1 3 5 H ya lu ro na n co nt en t ( ng /w el l) Lutein (M) ** ** ** 0 2 4 6 0 0.3 1 3 5 Lutein (M) D N A c on te nt ( g/ w el l)c
0 2 4 6 0 0.3 1 3 5 Lutein (M) 0 2 4 6 0 0.3 1 3 5 Lutein (M) D N A c on te nt ( g/ w el l)c
Fig. 2-4 The hyaluronan synthesis of human keratinocytes was upregulated by lutein in a dose dependent manner. Confluent human keratinocytes were incubated with various concentrations of lutein (0.3-5 mM) for 48h. (a) Hyaluronan content was measured as described in Materials and Methods. Values represent mean±SD of 3 replicate assays. Significantly different from the control value; **P < 0.01 (William’s test). (b) Total RNA was extracted and subjected to RT -PCR. The PCR products were visualized in a 2% agarose gel stained with ethidium bromide. (c) DNA content was measured as described in Materials and Methods. Values represent mean± SD of 3 replicate assays.
起す るの かを 調べ るた め, 評価 した ノン プロ ビ タミ ン A の うち , ルテ イン を用 い て検討 を進 めた. ル テイン は, 添加 1 -5 M で用 量依 存的な HAS3 m RNA 発現 誘導 を 伴って HA 産 生を 亢進し た( Fig. 2 -4 a, b). 本 実験条 件下 にお いてル テイ ンは 細胞 の ウェル 当た りの DNA 量に 影響 しなか った こと から, こ の HAS3 m RNA 発現 量の 増 加に細 胞増 殖が関 与し ない こと を示し た( Fig2- 4 c) .
2- 3- 2 ルテ インに よ る RAR 活 性 化
ノンプロ ビタ ミン A の ルテ インに よる HA 合 成誘 導に RARs の 関与 の有 無を 明 らかに する ため, RAR パ ンアン タゴ ニス トであ る L E54 050 )を 用い た. LE 540 の添 加
により, プ ロビ タミ ン A の -カ ロテ ンのみ なら ず, ル テイン の HA 産 生促 進作 用 も抑制 され た( Fig. 2 -5a). コン トロ ール 群の HA 合成 も L E54 0 存在 下で 一部 阻害 さ れた. さ らに , ルテ イン の RARs 活性 化を レポ ータ ーア ッセ イで 評価 した と ころ , HA 合 成を 亢進 する 濃度域 にて RARE 作 動に よる SE AP 活性 を用量 依存 的に 上昇 さ せた( Fig. 2 -5 b) . したが って ルテ イン自 体あ るい はそ の代謝 物が , 表皮 細胞 の RAR リガン ドと して機 能す るこ とが 示され た. 0 2000 4000 6000 8000 S E A P a ct iv it y Lutein (M) 0 2 5 10 ** b ** (a rb it ra ry c he m il um in es ce nc e u ni ts ) 0 100 200 300 Lutein Control H ya lu ro na n co nt en t ( ng /w el l) ** *** *** -Carotene a
Fig. 2-5 Effect of lutein on RAR activation. (a) Following 30min incubation of human keratinocytes in the presence (open columns) or absence (closed columns) of 0.2 M LE540, -carotene or lutein were added to the culture at a concentration of 5 M and incubated for 24 h. Hyaluronan content was measured as described in Materials and Methods. Values represent mean±SD of 3 replicate assays. Significantly different from the value of LE540 untreated cells; **P < 0.01, ***P <0 .001 (Dunnett’s test). (b) Keratinocytes transfected with pRARE-TA-SEAP reporter vector and an internal control vector were treated with the indicated concentration of lutein (2-10 M) for 24 h. SEAP activity was measured by the EscApe SEAP Chemiluminescence system. Values represent mean±SD of 3 replicate assays. Significantly different from the control value; **P < 0.01 (William’s test).
0 2000 4000 6000 8000 S E A P a ct iv it y Lutein (M) 0 2 5 10 ** b ** (a rb it ra ry c he m il um in es ce nc e u ni ts ) 0 100 200 300 Lutein Control H ya lu ro na n co nt en t ( ng /w el l) ** *** *** -Carotene a 0 2000 4000 6000 8000 S E A P a ct iv it y Lutein (M) 0 2 5 10 ** b ** (a rb it ra ry c he m il um in es ce nc e u ni ts ) 0 100 200 300 Lutein Control H ya lu ro na n co nt en t ( ng /w el l) ** *** *** -Carotene a
2- 3- 3 ルテ イン の HA 合 成誘 導に 対する シト ラー ルの 効果 表皮細胞 はレ チナー ルを レチ ノイン 酸へ と変 換する 能力 を有 する.51 )RALDHs の 阻害薬 であ るシト ラール 52 )を 用い, ル テイ ンの HA 合成 誘導 に及ぼ す影 響を 検討 し た. シトラ ール は, RA 刺 激によ る HA 合成 誘導 には 影響し なか った が, ルテ イン 存 在下で の HA 合成 誘導を 阻害 した (Fig. 2 -6) . こ の結 果から , ルテイ ンは プロ ビタ ミ ンA の代 謝と 同様 , 表 皮細 胞によ りア ルデ ヒド に変換 され , 引 き続き RALDH 依 存 的 な 酸 化 反 応 に よ り レ チ ノ イ ン 酸 様 の カ ル ボ ン 酸 へ と 代 謝 さ れ, そ の 代 謝 物 が RAR の リガ ンドと して 機能 する と考え られ た( Fig. 2- 7). H ya lur ona n cont ent (n g/ w el l) RA Lutein Control RAL 0 200 400 600 800 ** *** n.s. n.s. H ya lur ona n cont ent (n g/ w el l) RA Lutein Control RAL 0 200 400 600 800 ** *** n.s. n.s.
Fig. 2-6 Citral inhibits the effect of lutein on hyaluronan synthesis. Following 30 min incubation of human keratinocytes in the presence ( open columns) or absence (closed columns) of 20 M citral, 10 nM retinoic acid (RA), 50 nM retinal (RAL) and 10 M lutein were added to the culture and incubated for 48 h. Hyaluronan content was measured as described in Materials and Methods. Values represent mean±SD of 3 replicate assays. Significantly different from the value of citral untreated cells; **P < 0.01, ***P < 0.001 (Dunnett’s test). Citral Lutein Aldehydes RARE transactivation LE540 Carboxylic acids
HAS3 mRNA induction
Hyaluronan production
Monooxygenase?
A ldehyde dehydrogenase
Fig. 2-7. Possible metabolic transformation of lutein in keratinocytes. Citral Lutein Aldehydes RARE transactivation LE540 Carboxylic acids
HAS3 mRNA induction
Hyaluronan production
Monooxygenase?
A ldehyde dehydrogenase
第 4 節 :考 察 本研 究に より, プ ロビ タミン A に 属する -カロ テン のみ ならず , ノン プロ ビタ ミ ンA で ある ルテイ ンも RAR 依 存的 に表皮 細胞 の HA 合 成を 亢進す るこ とが 明ら か になっ た. -カロ テン は重 要なビ タミ ン A 源で あり, 2 つの レチニ ル基 から 構成 される . 小 腸の粘 膜中で BCO1 によ り中央 開裂 し, 2 分子 のレ チナー ルを 生成 する . さ らに こ のレチ ナー ルがRAL DHs に より 活性型 レチ ノイ ドであ る RA に 変換 され る. BCO15 3, 54), RALDH55 )と も に 表皮 で の 発現 が 確認 さ れ てい る. 本 研究 で 用い た 培 養ヒ ト 正
第 3 章 細 胞内 HA 基 質 プー ルの 変動 によ る HA 合 成制 御機 構 第 1 節 :序 論 表皮細 胞の HA 合 成能を 高め る天 然物質 の探 索を 行っ たとこ ろ, N- アセ チル グル コサミ ン( NAG) にそ の作 用を 見出し た. NAG は , カ ニ, エビ などの 甲殻 類の 外殻 を 起源 と する 天 然多 糖 類キ チ ン を構 成 する 単 糖で あ り, す で に食 品 分野 で 広く 応 用 されて いる.64 ) 生体 にお いて は, HA の他 にヘパ リン およ びヘ パラ ン硫 酸な どの 硫 酸化 GAG の 構成 単糖 であ ると同 時に , 様 々な 糖タ ンパク , 糖脂質 を修 飾す るア ミ ノ糖 であ る. NAG が 糖 供与 体と して 機能 す るに は, 高 いエ ネル ギー を持 た なけ れ ば な ら な い た め, ヌ ク レ オ チ ド に 活 性 化 さ れ る 必 要 が あ る . 糖 ヌ ク レ オ チ ド の UDP -NAG は , グ ルコ ー スト ラ ンス ポ ータ ー( GL UT ) に より 取り 込 まれ た グル コ ー スから, そ の 2 -5 %が ヘキソ サミ ン経 路を経 て生 合成 される .65 ) 近 年, UDP- NAG を 糖供与 体と して O- NAG 転移 酵素 により 触媒 され る O-グ リコ シル化 の修 飾部 位が , タン パク のリ ン酸 化 部位 と同 一, あ るい はそ の近 傍 であ るこ とか ら, NAG は 細 胞 内シグ ナル 伝達に おい ても 重要 な役割 を担 うこ とが解 明さ れつ つある.66 , 67 ) 本研究 では, 培 養表 皮細胞 にお ける NAG の HA 合成 促進 機序を , RA の 作用 点と 比較す るこ とで検 討し た. 第 2 節 :実 験材 料お よび 実験 方法 3- 2- 1 培養 器具お よび 試薬
Human sk in f ibro blasts ( Detr iut 551, AT CC CCL11 0) (AT CC 製 ), Rer ino l およ び Retino ic acid ( Sigm a 社 製) , N-acetylgluco sam in e ( NAG) ( Seikagak u 社製 ), Eagle’s m in im al essential medium (M EM ) (I CN Bio medicals 社製 ), Fet al bo vine ser um ( FBS) (I rv in e Scient if ic 社 製) , 3, 5 - diam in o benzo ic acid ( Sigm a 社 製) , DC protein assay k it ( Bio r ad 社製 ), So dium[35S] Sulfat e (1 00 m Ci/mm ol) ( Am er sh am Pharm acia Biotech 社 製 ) , UDP- [14C] GlcA (30 0 m Ci/mmo l) お よ び [1,6 -3H( N)] N- acety lgluco sam ine(60Ci/mm ol)(Am erican Radio labeled Chem icals
社製) , Partisp here SAX (W hatm an 社製) , UDP -N-acety lgluco sam in e ( Kyo wa 社 製).
その他 の試 薬につ いて は, 第 1 章 および 第 2 章で 記載し たも のを 用いた .
3- 2- 2 両親 媒性 NAG 誘 導 体 の合 成
ル(n =2 , 4 , 5 , 6, 8, 12 ) と のグリ コシ ル化 を行 い, それ ぞれ 55 -75 %の 収率で -グリ コシドB を得た . グリ コシ ドの 脱アセ チル 化に より , 目 的とす る 1 位 に親 油性 を もつ NAG 誘 導体 を定 量的 に得た .68 ) 3- 2- 3 細胞 培養 ヒト 表皮 細胞の 培養 は第 1 章に 記載 した 方法で 実施 した 。ヒト 皮膚 線維 芽細 胞 (Detro it 55 1)は , 10 % FBS 含有 ME M 培 地を 用い て 24 - well プレ ート に播 種した . コ ンフル エン トに達 する まで 培養 の後, FBS 不 含培 地で 培地 交換を おこ ない 24 h 馴 化 させた. 次 に評 価化 合物を 添加 し, 24 h 後 の培 養上 清中 HA 量を 定量 した 。 3- 2- 4 DNA 定 量 DNA 量の 定量 は, 第 2 章で 記載し た方 法に 準じて 実施 した . 3- 2- 5 HA 定 量 培養 上清 中の HA 量は , 第 1 章 で記 載し た方法 に準 じて 実施 した. 3- 2- 6 HAS 活 性測 定法 HA 合 成酵 素活性 の測 定は , It ano らの 方法 4 1 )に 従い実 施し た. NAG (5 m M)
あるいは all- tran s Retino ic acid ( RA) (1 M)で 24 h 刺激し た表 皮細 胞を回 収し , 0.5 mM dith iothr eiro l お よび 0.2 5 m M sucro se を含 有す る 10 m M HEP E S-NaOH (pH 7 .1 )中で 超音 波破 砕した . 次に , 細 胞破 砕画 分を超 遠心 (105 ,000 g, 1 h)す ること によ り粗膜 画分 を得 た. UDP -[14C] GlcA(10Ci/m l), 5 m M dith ioth reito l, 15 m M M gCl2, 0 .2 mM UDP- GlcA, 1. 0 m M UDP- NAG を 含 む 2 5 m M
HEPE S- NaOH (p H 7 .1 ) に粗 膜画 分 を添 加 する こ とに より 酵 素反 応 を開 始し (total vo lum e 20 0 l), 37℃ で 1 h イ ンキ ュベ ートし た. 酵素 反応 は, 2 % so dium do decy l sulf ate 存在下 で 10 0℃ , 1 min 処 理する こと に停 止した . 酵素反 応物 を Sep h a dex G- 5 0 ゲル ろ過 カラム ( 1 5 x 5 0 m m ) に 供し, そ の素通 し画 分の
放射活 性を 測定し た.
3- 2- 7 硫酸 化 GAG 合 成能 評価
硫酸化 グリ コサミ ノグ リカ ン(硫 酸化 GAG) の de n ovo 合成 能は , [3 5S] Sulf ate の培 養上清 GA G 画分 への 取り 込み活 性に より 評価し た. 培養 表皮 細胞 に NAG ( 5 m M) , あるい は RA (1 M) を添 加し, [35S] Sulfat e (2 0 Ci/ml)と とも に 48 h 培養 した, 培 養
上清 をプ ロナ ーゼ で 消化(30 µg/ml) した 後, Seph adex G- 50 ゲル ろ過 カラ ム( 15x5 0 mm)に 供し , そ の素 通し 画分の 放射 活性 を測定 した .
3- 2- 8 NAG K cDNA の 単 離
ヒト N- acety lgluco samine kinase ( NAGK) 遺伝 子の cDNA は RT -P CR よ り単離 した 。 ヒ ト 表 皮 細 胞 か ら 抽 出 し た Tot al RNA を 用 い , m RNA の シ ー ケ ン ス ( Gen Ban k accessio n n um ber AJ242 910 )を 元に 下記 のプ ライ マー を 設計 し, RT -P CR を 行っ た . Hum an NAGK sen se p r imer 5’ - GGAAAGCT T GGCACACGAT CCGAGGT C- 3’ ; Human NAGK ant isen se p rimer 5’- AGGGAAT T CGGA GCCGCCT CTAGGT T G- 3’ ( 30 cy cle, 下 線部は Hind III およ び E coR I 制限 酵素 サイ ト) . 得 られ たP CR 産物 をア ガロ ース ゲ ル電気 泳動 の後, ゲ ルから 切り し精 製した . 精製 した cDNA を HindI II /E co RI で 消 化し, プラ スミ ドベ クター (p SPT 18) HindI II /E co RI サイト に組 み込 んだ . シ ーケ シ ングに より, 目 的と する NAGK cDNA が得 られた こと を確 認し, DI G RNA labelin g kit プ ロト コルに 従い , DI G 標 識し た antisen se/sen se RNA プロ ーブ を調製 した .
3- 2- 9 ノザ ンブロ ット 解析
ノザン ブロ ット解 析は, 第 1 章 で記 載した 方法 に準 じて 実施し た .
3- 2- 10 細胞 内 UDP- NAG の 定 量
し, 予め作 成し た検 量線を もと に定 量をお こな った .
3- 2- 11 NAG の 細 胞 内 取り 込み 評価
24- well プレ ート に播 種し た表 皮細胞 をコ ンフ ルエ ント の状 態まで 培養 し, 培 地 を除去 の後, 細 胞表 面をグ ルコ ース 不含 Krebs- Rin ger リ ン酸 緩衝液 (KRP )で 洗浄 し た. 細胞に , [3H] GlcN (1Ci/m l KRP, 2 0 nM)ある いは [3H] NAG (1Ci/m l KRP, 1 7 nM)
を添 加 し, そ れ ぞれ の 添加 群 にグ ル コー ス ト ラン ス ポー タ ー阻 害 薬で あ るサ イ ト カラシン B を 添加 した (0-3 0 M). 37 ℃で 10 m in 静置後 , 上 清を 除去 し, ice-cold PBS で 細胞 表面 を 3 回 洗浄 した . 0 .2 % SD S 溶 液で 細胞を 溶解 させ , 液 体シ ンチ レ ーショ ンカ ウンタ ーで 放射 活性 を測定 した.
3- 2- 12 統計 処理
統計解 析は SAS シ ステ ムによ り実 施し た. Dun nett's m ultiple comp ar ison test でコン トロ ールと の有 意性 を解 析し, 用量 依存 性の 評価 には William’s test を 用いた. p 値 0 .05 未満 を統 計的に 有意 とし た. 第 3 節 :実 験結 果 3- 3- 1 NAG に よ る 表皮 細胞 の HA 合 成促 進作 用 単糖の HA 合成 に及ぼ す影 響を 把握 するた め, グル コー ス, ガラ クト ース, マ ン ノース, グ ルク ロン 酸およ び NAG をそ れぞ れ最 終濃 度 10 m M でヒ ト表 皮細 胞に 添 加した とこ ろ, NAG 添 加群 のみ 顕著 に HA 合 成が 促進 された ( Fig. 3 -1) . NAG が 作 用を発 現す る添加 濃度 域に つい て調べ たと ころ, 添 加 3 mM より 1 0 m M ま で用 量 依存的 に HA 合成 が亢進 され た( Fig. 3- 2a). 0 0.2 0.4 0.6 Con
trol Glc Gal Man NAG GlcA
H A co nt en t ( g /w el
l) Fig. 3-1 Effects of monosaccharides on HA
production in keratinocytes. Normal human keratinocytes were cultured with glucose, galactose, mannose, NAG or glucuronic acid at the concentrations of 10 mM for 24 h and hyaluronan production was measured as described in Materials and Methods
.
. Values represent mean± SD from three individual wells. 00.2 0.4 0.6
Con
trol Glc Gal Man NAG GlcA
H A co nt en t ( g /w el l) 0 0.2 0.4 0.6 Con
trol Glc Gal Man NAG GlcA
H A co nt en t ( g /w el
l) Fig. 3-1 Effects of monosaccharides on HA