カロテノイド:その多様性と普遍性が切り拓く新展開
篠村知子
帝京大学理工学部バイオサイエンス学科
〒320-8551 栃木県宇都宮市豊郷台1-1
Tomoko Shinomura
Diverse physiological roles of widespread carotenoids in photosynthetic organisms
Key words: carotenoid, carotene, xanthophyll, photoprotection, antioxidation Department of Biosciences, School of Science and Engineering, Teikyo University
1-1 Toyosatodai, Utsunomiya, Tochigi, 320-8551, Japan DOI: 10.24480/bsj-review.9b1.00133
カロテノイドの存在の普遍性
カロテノイドはテトラテルペノイド(C40)の一種であり, その構造からカロテン類とキサントフィ ル類に大別される。カロテン類は,炭素の共役2重結合からなる基本骨格(ポリエン鎖)とその両端に つくエンドグループが, 炭素と水素原子のみで構成される(図1A)。一方, キサントフィル類に分類 されるカロテノイドのエンドグループには, アルコール, ケトン, アルデヒド, カルボン酸, エポキ シド, ラクトンなどの酸素原子を含む官能基がある(図1B)(Niyogi et al., 2015; 眞岡, 2009)。カロテ ノイドの種類は非常に多く, 原核生物から動物や高等植物までを含む 683 種類の生物種から約1100 種類以上の天然カロテノイドが報告されている(Yabuzaki, 2017)。カロテノイドを生合成するのは植物 と藻類と, ごく限られた種類のバクテリアと菌類に限られることから, 様々な植物種や藻類種の生産 するカロテノイド組成やその合成酵素遺伝子を調べることで, 進化的な類縁関係や葉緑体の共生によ る進化の歴史が議論されてきた(三室ら, 2006)。動物は食物連鎖によってカロテノイドを摂取するの で, 動物種のカロテノイド組成を調べることは生態系の食物連鎖の関係を明らかにすることになる
(眞岡, 2009)。
図1 代表的なカロテノイドの構造 (A) β-カロテン;炭素と水素原子のみで構 成されるカロテン類の代表的なカロテノイド (B) ビオラキサンチン;分子内に酸素原子を 含む官能基をもつキサントフィル類の一種 (C) アスタキサンチン;強力な抗酸化作用を もち, 食品, 機能性サプリメント, 化粧品, 飼料および水産養殖分野で利用されているカ ロテノイド
カロテノイドの機能の多様性と進化
生物はこのように非常に多種類のカロテノイド分子を, どのような生理機能に利用しているのだろ うか。植物や藻類の光合成においては, 補助色素としての作用や, キサントフィルサイクルを介する
「光防御作用」や「抗酸化作用」などが明らかになりつつある(本総説集の高橋と西山 参照)。すな わち, カロテノイドは, シアノバクテリアや藻類や植物が光合成のための光を絶対的に必要とする反 応系において, 補助色素としての集光の役割や, 光を受け取ることで必然的に生じる過剰エネルギー やラジカルなどのストレス要因を緩和するための機能性分子の役割を果たしてきたと考えられる。さ らに, 食品栄養科学や医療分野でも, ヒトにおけるカロテノイドの欠乏症が夜盲症などの重篤な疾病 を引き起こすこと, β-カロテンなどを「ビタミンAの前駆体」(プロビタミンA)として摂取すると 小腸から吸収される際に酸化開裂酵素によりレチナール(ビタミンAの一種)に変換されてそれが視 細胞のロドプシンの発色団となること, ヒト網膜の最も光が集まる黄斑にはゼアキサンチンが集中的 に蓄積していることが知られている(三室ら, 2006)。これらの研究から, カロテノイドは生物進化の 非常に早い段階から, 光をエネルギー源として利用する反応系や, 光を情報媒体として利用する反応 系において, 生物が必ず利用してきた化合物分子であるといえるであろう。
一方, ヒトや類人猿には, 光とは一見関係がなさそうな脳組織にルテインなどのキサントフィル類
が蓄積することが報告されている(Erdman et al., 2015)。その他にも, ある種のカロテノイドがビタミ ンAに変換されるのではない機構(ノンプロビタミンA活性)で抗癌作用や抗動脈硬化作用を示すこ とが報告されており, その機能は, カロテノイドの「活性酸素の消去作用」, すなわち「抗酸化物質」
としての働きによると説明されている(三室ら, 2006; 宮下, 2009)。抗酸化作用のあるカロテノイドと して最も注目を集めているのはアスタキサンチン(図1C)であろう。アスタキサンチンはキサントフ ィル類の一種で, 単細胞性の鞭毛をもつ緑藻の一種であるHaematococcus pluvialisの休眠胞子に蓄積し, 強力な抗酸化活性をもつことがよく知られている(Kobayashi et al., 1997)。これらの微細藻類やある種 のバクテリアなどが生合成して蓄積したものを, 生態系のより上位の消費者のエビやカニやサケなど が摂取して鮮やかな赤色やピンク色の色素物質として蓄積する(矢澤,2009)。
このように, カロテノイドが非光合成生物を含む生物界全体に広く分布することや, その機能が光 受容関連の反応系にとどまらず多様な機能を示すことは, 光受容体フィトクロムの進化研究の飛躍的 拡大の道筋を連想させる。近年, フィトクロムタンパク質遺伝子に類似したフィトクロム様タンパク 質遺伝子が, シアノバクテリアにも(Ikeuchi & Ishizuka, 2008), 非光合成バクテリアにも見出され (Davis et al., 1999), フィトクロム進化の議論は, 一気に陸上植物の範囲を超えて非光合成バクテリア や菌類や後生動物を含むほぼすべての生物群がもつ酸素センサーとしてのヘムタンパク質にまで起源 を求めるほどに深化したことは記憶に新しい(Montgomery & Lagarias, 2002)。カロテノイドがさらに 興味深いのは, 多くの動物はカロテノイドを一部改変するとはいえ基本的には生合成せず, 分子その ものを摂取して利用することで進化を遂げてきたのである。こんなユニークな生体分子が他にあるだ ろうか。カロテノイド分子をめぐる進化は生命進化そのものの本質に迫る新しい知見を示すに違いな いと筆者は確信する。
本総説集の構成と今後の展開
本総説集は, 日本植物学会第81回大会(2017年9月, 東京理科大学 野田キャンパス)で開催さ
れたシンポジウム「カロテノイド:その多様性と普遍性が切り拓く新展開」の内容をもとに, 発表者 らの協力のもとにまとめたものである。このシンポジウムでは, まずカロテノイドの機能解析の最も 進んでいる光合成における機能解析や(本総説集の高橋と西山 参照), 合成経路探索の最先端研究が 紹介された(本総説集の高市 参照)。続いて進化系統上ユニークな材料であり産業応用も期待されて
いるEuglena gracilisのカロテノイド合成系の遺伝子探索や, カロテノイド合成と強光ストレス耐性と
の関連を議論し(本総説集の加藤と篠村 参照), カロテノイド合成経路探索の一連の研究が広く生物 界に分布するシトクロム P450 の機能解析の研究と結びついたことも紹介された(本総説集の玉木と 今石 参照)。さらに, 鞭毛をもつ微細藻類の眼点の主な組成がカロテノイドであることはよく知られ ているが, 近年, Chlamydomonas reinhardiiの走光性の研究においてカロテノイド欠損株を用いた興味 深い研究から, カロテノイドはレドックス制御を介する走光性の「高精度な光反射板」として機能す ることが明らかになったことが紹介された(本総説集の若林ら 参照)。本シンポジウム後半には, シ アノバクテリアを用いてカロテノイドを高蓄積させるための遺伝子改変の試みや(本総説集の島田ら 参照), 合成経路の代謝工学的な改変などの最新のアプローチが紹介された(梅野ら 大会要旨集 p.105)。カロテノイドの分析技術に関しては, HPLC解析やLC-MS解析が飛躍的進歩を遂げたことは言 を待たないが, ラマンスペクトル分光法の目ざましい発展により, 生きた細胞のカロテノイド分布を 無標識でイメージング解析することが可能になりつつあることが紹介された(鈴木ら 大会要旨集
p.105)。玉木博士, 梅野博士および鈴木博士らは他学会からの招待講演者であり, カロテノイド研究
の新展開を学会横断的に共有しつつ, 進化生物学の考察から将来の医療や化粧品やサプリメントなど の産業応用への新たな進展までをも議論する貴重な機会となった。
今日なお, 希少な種類のカロテノイドが, 大腸菌等を用いて合成して分析することで新規な種類の カロテノイドとして報告されている(Takemura et al., 2015)。生物はどのようにして, そしてどのよう な生理機能上の必要があって, かくも多様なカロテノイドを進化させてきたのだろうか。その解明は 緒に就いたばかりであり, 生物学的興味は尽きない。
謝辞
シンポジウムの企画に際してはオーガナイザーとして池内昌彦博士(東京大学総合文化研究科)に 多大な協力を仰いだこと, シンポジウム開催と総説集執筆に際して大会実行委員や編集委員の先生方, および発表者の方々に深謝することをここに記す。
引用文献
Davis, S. J., Vener, A. V., Vierstra, R. D. 1999. Bacteriophytochromes: phytochrome-like photoreceptors from nonphotosynthetic eubacteria. Science 286: 2517–2520.
Erdman, J. W. Jr., Smith, J. W., Kuchan, M. J., Mohn, E. S., Johnson, E. J., Rubakhin, S. S., Wang, L., Sweedler, J.V., Neuringer, M. 2015. Lutein and brain function. Foods 4: 547–564. doi: 10.3390/foods4040547
Ikeuchi, M., Ishizuka, T. 2008. Cyanobacteriochromes: a new superfamily of tetrapyrrole-binding photoreceptors in cyanobacteria. Photochem. Photobiol. Sci. 7: 1159–1167.
Kobayashi, M., Kakizono, N., Nishio, N., Nagai, S., Kurimura, Y., Tsuji, Y. 1997. Antioxidant role of astaxanthin in the green alga Haemtococccus pluvialis. Appl. Environ. Biotechnol. 48: 351–356.
眞岡孝至 2009.カロテノイドの構造と生物界における分布.宮下和夫(編)カロテノイドの科学と 最新応用技術. pp. 3-14. シーエムシー出版.東京
宮下和夫 2009.プロビタミンA活性とノンプロビタミンA活性.宮下和夫(編)カロテノイドの科学 と最新応用技術. pp. 107-115. シーエムシー出版.東京.
三室守,高市真一,富田純史 2006.高市真一(編)カロテノイド-その多様性と生理活性-.裳華 房.東京.
Montgomery, B. L., Lagarias, J. C. 2002. Phytochrome ancestry: sensors of bilins and light. Trends Plant Sci., 7:
357-366.
Niyogi, K. K., Wolosiuk, R. A. & Malkin, R. 2015. Photosynsthesis. In: Buchanan, B. B., Gruissem, W. & Jones, R. L. (eds.) Biochemistry & Molecular Biology of Plants, Second edition. pp. 508-566. John Wiley & Sons, Chichester, UK.
Takemura, M., Maoka, T., Misawa, N. 2015. Biosynthetic routes of hydroxylated carotenoids (xanthophylls) in Marchantia polymorpha, and production of novel and rare xanthophylls through pathway engineering in Escherichia coli. Planta 241: 699-710.
Yabuzaki, J. 2017. Carotenoids Database: structures, chemical fingerprints and distribution among organisms.
Database box004: 1-11, doi:10.1093/database/bax004
矢澤一良(編) 2009. アスタキサンチンの科学. 成山堂書店. 東京.
H. Takahashi-1
光 合 成に おけ るカ ロ テノ イド の 機 能
髙 橋 拓 子 , 西 山 佳 孝
埼 玉 大 学 大 学 院 理 工 学 研 究 科 生 命 科 学 部 門
〒338-8570 埼 玉 県 さ い た ま 市 桜 区 下 大 久 保 255
Hiroko Takahashi1, Yoshitaka Nishiyama1
Roles of carotenoids in photosynthesis
Keywords: carotenoids, energy quenching, photosystem, photoprotection, repair of PSII
1Department of Biochemistry and Molecular Biology, Saitama University 255 Shimo-Okubo, Sakura-ku, Saitama 338-8570, Japan
DOI: 10.24480/bsj-review.9b2.00134
1. は じ め に -光 合 成 と カ ロ テ ノ イ ド-
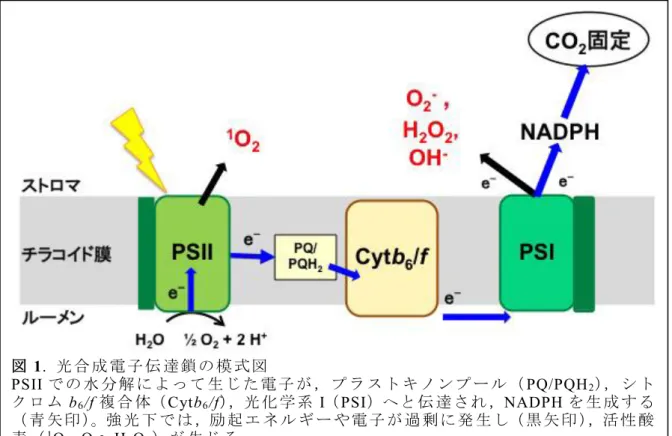
光 合 成 細 菌 か ら 陸 上 植 物 に 至 る ま で す べ て の 光 合 成 生 物 は カ ロ テ ノ イ ド を 合 成 す る こ と が で き ,多 様 な カ ロ テ ノ イ ド を 有 し て い る (三 室 et al. 2006)。ま た ,光 合 成 生 物 で は カ ロ テ ノ イ ド は 主 に チ ラ コ イ ド 膜 に 存 在 す る 。チ ラ コ イ ド 膜 に は ,光 化 学 系 複 合 体 な ど 様 々 な タ ン パ ク 質 複 合 体 が 存 在 し て お り ,光 エ ネ ル ギ ー 変 換 や 電 子 伝 達 が 起 こ る 。光 化 学 系 II(PSII)で 光 エ ネ ル ギ ー が 化 学 エ ネ ル ギ ー に 変 換 さ れ る と き , 水 分 子 か ら 電 子 が 引 き 抜 か れ ( 水 分 解 反 応 ), 酸 素 が 発 生 す る 。 電 子 は プ ラ ス ト キ ノ ン プ ー ル ,シ ト ク ロ ム b6/f 複 合 体 を 経 て 光 化 学 系 I(PSI)に 伝 達 さ れ る 。 PSI で 光 エ ネ ル ギ ー 変 換 が 起 こ り , 電 子 が NADP+に 伝 達 さ れ ,NADPH が 生 成 す る
( 図 1)。光 エ ネ ル ギ ー は 光 合 成 の 駆 動 に 必 須 で あ る が ,過 剰 な 光 エ ネ ル ギ ー は 活 性 酸 素 を 誘 発 し , タ ン パ ク 質 や 核 酸 , 生 体 膜 な ど に 酸 化 傷 害 を 及 ぼ す 。
カ ロ テ ノ イ ド は ,8 個 の イ ソ プ レ ノ イ ド が 直 鎖 状 に 結 合 し た C40H56を 基 本 骨 格 と す る 有 機 化 合 物 で あ り ,炭 化 水 素 で あ る カ ロ テ ン と ,酸 素 を 含 む 官 能 基 が 付 い た キ サ ン ト フ ィ ル に 大 別 さ れ る 。カ ロ テ ノ イ ド は 光 エ ネ ル ギ ー を 吸 収 す る が ,ク ロ ロ フ ィ ル の よ う に 光 化 学 反 応 を 起 こ さ な い 。ま た カ ロ テ ノ イ ド は ,励 起 エ ネ ル ギ ー を 熱 に 変 換 し て 過 剰 エ ネ ル ギ ー を 散 逸 し た り ,ラ ジ カ ル 化 合 物 と 電 子 授 受 し て ラ ジ カ ル を 消 去 し た り す る 。す な わ ち ,光 合 成 で は カ ロ テ ノ イ ド は 集 光 や 光 防 御 ,抗 酸 化 と い っ た 役 割 を 担 っ て い る 。 本 稿 で は , 酸 素 発 生 型 光 合 成 に お け る カ ロ テ ノ イ ド の 様 々 な 役 割 に つ い て 述 べ る と と も に ,カ ロ テ ノ イ ド の 光 防 御 機 能 に 関 し て 新 た な 知 見 を 紹 介 す る 。
2. 光 合 成 装 置 に 結 合 す る カ ロ テ ノ イ ド
近 年 の 結 晶 構 解 析 技 術 の 進 歩 に よ っ て ,光 化 学 系 複 合 体 お よ び シ ト ク ロ ム b6/f 複 合 体 に 結 合 す る カ ロ テ ノ イ ド の 種 類 や 数 が 明 ら か に な っ て き た 。ま た ,シ ア ノ バ ク テ リ ア で は カ ロ テ ノ イ ド を 結 合 す る 可 溶 性 タ ン パ ク 質 も あ る 。光 合 成 装 置 に 結 合
H. Takahashi-2
す る カ ロ テ ノ イ ド の 分 子 数 は ,ク ロ ロ フ ィ ル 分 子 数 の 約 20%に 相 当 す る 。こ れ ら の カ ロ テ ノ イ ド は 主 に β-カ ロ テ ン や キ サ ン ト フ ィ ル 類 で あ り ,反 応 中 心 複 合 体 に は 主 に β-カ ロ テ ン が 結 合 し ,集 光 性 複 合 体 に は 主 に キ サ ン ト フ ィ ル 類 が 結 合 す る が ,そ の 分 子 種 は 生 物 種 に よ っ て 異 な る 。こ の 章 で は ,結 晶 構 解 析 に よ り 明 ら か に な っ た カ ロ テ ノ イ ド の 分 子 種 と 局 在 性 を 述 べ る 。
2-1. 光 化 学 系 反 応 中 心 複 合 体 に 結 合 す る カ ロ テ ノ イ ド
PSII の 構 は , 好 熱 性 シ ア ノ バ ク テ リ ア か ら 単 離 さ れ た PSII 複 合 体 の 結 晶 構 解 析 か ら 明 ら か に な っ て い る (Ferreira et al. 2004, Kamiya and Shen 2003, Loll et al.
2005, Umena et al. 2011)。PSII は 2 量 体 で 存 在 し ,PSII 単 量 体 あ た り 16分 子 の カ ロ テ ノ イ ド が 結 合 す る 。 そ の う ち 11分 子 は ト ラ ン ス 型 β-カ ロ テ ン で あ る (Loll et al.
2005) 。反 応 中 心 を 形 成 す る D1,D2 サ ブ ユ ニ ッ ト に は そ れ ぞ れ 1 分 子 の カ ロ テ ノ イ ド が 結 合 し て い る 。D2 サ ブ ユ ニ ッ ト に 結 合 し て い る β-カ ロ テ ン は , 電 子 供 与 体 と し て シ ト ク ロ ム b559 を 介 し た 電 子 移 動 に 関 与 す る と 考 え ら れ て い る (Kamiya and Shen 2003)。 ま た , 反 応 中 心 集 光 性 サ ブ ユ ニ ッ ト CP43 に 3 分 子 ,CP47に 5 分 子 の カ ロ テ ノ イ ド が 結 合 す る 。そ の 他 ,周 辺 サ ブ ユ ニ ッ ト の 近 傍 に も カ ロ テ ノ イ ド が 結 合 し て い る 。
一 方 PSIに 関 し て は , 好 熱 性 シ ア ノ バ ク テ リ ア で は 3 量 体 PSI, 陸 上 植 物 で は 単 量 体 PSI の 結 晶 構 が 明 ら か に な っ て い る (Amunts et al. 2007, Jordan et al. 2001)。 好 熱 性 シ ア ノ バ ク テ リ ア で は ,PSI 単 量 体 あ た り 22 分 子 の カ ロ テ ノ イ ド が 結 合 し て お り , 電 子 密 度 か ら そ の 分 子 種 が β-カ ロ テ ン だ と 推 測 さ れ て い る (Jordan et al.
図 1. 光 合 成 電 子 伝 達 鎖 の 模 式 図
PSII で の 水 分 解 に よ っ て 生 じ た 電 子 が , プ ラ ス ト キ ノ ン プ ー ル (PQ/PQH2), シ ト ク ロ ム b6/f複 合 体(Cytb6/f),光 化 学 系 I(PSI)へ と 伝 達 さ れ ,NADPHを 生 成 す る
( 青 矢 印 )。強 光 下 で は ,励 起 エ ネ ル ギ ー や 電 子 が 過 剰 に 発 生 し( 黒 矢 印 ),活 性 酸 素 (1O2, O2-, H2O2) が 生 じ る 。
H. Takahashi-3
2001) 。22分 子 の う ち ,10 分 子 は PSI 反 応 中 心 の PsaA/Bサ ブ ユ ニ ッ ト に 結 合 し て い る 。 残 り の カ ロ テ ノ イ ド は , ル ー メ ン 側 サ ブ ユ ニ ッ ト PsaF と PsaJ, お よ び 単 量 体 同 士 の 隣 接 面 に 位 置 す る PsaI,PsaL,PsaM,PsaK サ ブ ユ ニ ッ ト に 結 合 し て い る (Jordan et al. 2001)。PSI と PSII の 反 応 中 心 複 合 体 の 構 の 類 似 性 が 指 摘 さ れ て い る が ,反 応 中 心 ク ロ ロ フ ィ ル に 対 す る カ ロ テ ノ イ ド の 配 置 も 両 者 の 間 で 類 似 し て い る (Nelson and Yocum 2006)。
2-2. 集 光 性 複 合 体 に 結 合 す る カ ロ テ ノ イ ド
緑 藻 類 や 陸 上 植 物 で は ,膜 内 在 性 の light-harvesting complex II (LHCII) が PSII 集 光 性 タ ン パ ク 質 複 合 体 と し て 機 能 し て い る 。LHCII は ,3 量 体 を 形 成 す る 主 要 な 集 光 性 複 合 体 ( 陸 上 植 物 で は Lhcb1-3) と , 単 量 体 で 存 在 す る マ イ ナ ー サ ブ ユ ニ ッ ト
( 陸 上 植 物 で は Lhcb4-6)に 大 別 さ れ ,そ れ ぞ れ PSII 反 応 中 心 複 合 体 と 超 分 子 複 合 体 を 形 成 し て い る (Minagawa and Takahashi 2004, Nield et al. 2000)。LHCII 3 量 体 の 結 晶 構 は 明 ら か に な っ て お り ,単 量 体 あ た り 3 本 の 膜 貫 通 部 位 を も つ ア ポ プ ロ テ イ ン に ,8 分 子 の ク ロ ロ フ ィ ル a,6 分 子 の ク ロ ロ フ ィ ル b,4 分 子 の カ ロ テ ノ イ ド が 結 合 す る 。ま た ,こ の 4 分 子 の カ ロ テ ノ イ ド は キ サ ン ト フ ィ ル で あ り ,そ の 内 訳 は 1 分 子 の ル テ イ ン ,1 分 子 の ネ オ キ サ ン チ ン ,2 分 子 の キ サ ン ト フ ィ ル サ イ ク ル カ ロ テ ノ イ ド( 後 述;ゼ ア キ サ ン チ ン ,ヴ ィ オ ラ キ サ ン チ ン ,ア ン テ ラ キ サ ン チ ン の い ず れ か ) で あ る (Liu et al. 2004) 。
PSI集 光 性 タ ン パ ク 質 複 合 体 light-harvesting complex I (LHCI) は , 紅 藻 を 含 む 藻 類 や 陸 上 植 物 で PSI集 光 装 置 と し て 機 能 し て い る 。エ ン ド ウ(Pisum sativum)に お け る PSI -LHCI結 晶 構 解 析 (Amunts et al. 2007, Ben-Shem et al. 2003, Qin et al.
2015) に よ り ,LHCI の サ ブ ユ ニ ッ ト(Lhca1-4)は ,Lhca1 と Lhca4,Lhca2 と Lhca3 の 2 つ の ヘ テ ロ ダ イ マ ー が 並 列 に PSI 単 量 体 の 片 側 に 結 合 す る こ と が 示 さ れ て い る 。4 つ の LHCI サ ブ ユ ニ ッ ト で は , ク ロ ロ フ ィ ル a(45 分 子 ), ク ロ ロ フ ィ ル b
(12 分 子 ),β-カ ロ テ ン(4 分 子 ),ル テ イ ン(5 分 子 ),ヴ ィ オ ラ キ サ ン チ ン(4 分 子 ) が 同 定 さ れ て い る (Qin et al. 2015)。
2-3. シ ト ク ロ ム b6/f 複 合 体 に 結 合 す る カ ロ テ ノ イ ド
シ ト ク ロ ム b6/f 複 合 体 は ,キ ノ ン 結 合 部 位 Qo,Qi が 内 向 き に な る よ う に 2 量 体 を 形 成 し て い る (Kurisu et al. 2003, Stroebel et al. 2003)。 単 量 体 あ た り 1 分 子 の β- カ ロ テ ン が , シ ト ク ロ ム b6 サ ブ ユ ニ ッ ト に 結 合 し て い る (Kurisu et al. 2003)。
2-4. カ ロ テ ノ イ ド を 結 合 す る 水 溶 性 タ ン パ ク 質
上 記 の 複 合 体 は い ず れ も チ ラ コ イ ド 膜 タ ン パ ク 質 で あ る が ,カ ロ テ ノ イ ド を 結 合 す る 可 溶 性 タ ン パ ク 質 が 存 在 す る 。 そ の 一 種 に オ レ ン ジ カ ロ テ ノ イ ド プ ロ テ イ ン (orange carotenoid protein; OCP) が あ り , シ ア ノ バ ク テ リ ア に 広 く 保 存 さ れ て い る (Holt and Krogmann 1981, Kirilovsky and Kerfeld 2012)。OCP は ,7 本 の α ヘ リ ッ ク
H. Takahashi-4
ス か ら な る N末 端 ド メ イ ン と ,2 本 の αヘ リ ッ ク ス と 7 本 の β 鎖 か ら な る β バ レ ル を 含 む C 末 端 ド メ イ ン か ら な り , ド メ イ ン を ま た ぐ よ う に 1 分 子 の カ ロ テ ノ イ ド が 結 合 し て い る (Kerfeld et al. 2003, Leverenz et al. 2015, Wilson et al. 2010)。 そ の カ ロ テ ノ イ ド は ,3'-ヒ ド ロ キ シ エ キ ネ ノ ン で あ る が ,エ キ ネ ノ ン ,ゼ ア キ サ ン チ ン , カ ン タ キ サ ン チ ン も 結 合 し て 役 割 を 補 填 す る こ と が 知 ら れ て い る (Leverenz et al.
2015, Punginelli et al. 2009, Wilson et al. 2011)。OCP の 相 同 タ ン パ ク 質 と し て , ヘ リ カ ル カ ロ テ ノ イ ド プ ロ テ イ ン(helical carotenoid protein; HCP)と い う フ ァ ミ リ ー も 存 在 し ,OCP の N 末 端 ド メ イ ン 部 分 の み を も ち 、 そ こ に カ ロ テ ノ イ ド が 結 合 す る と い う 構 が 予 測 さ れ て い る (Melnicki et al. 2016)。
3. カ ロ テ ノ イ ド の 集 光 機 能
集 光 色 素 と し て 機 能 す る カ ロ テ ノ イ ド は ,ク ロ ロ フ ィ ル と と も に 集 光 性 タ ン パ ク 質 に 結 合 し て い る 。ク ロ ロ フ ィ ル の 励 起 寿 命 が 数 ナ ノ 秒 で あ る の に 対 し ,カ ロ テ ノ イ ド の 励 起 寿 命 は お よ そ 10ピ コ 秒 と 短 い (三 室 et al. 2006)。 そ の た め カ ロ テ ノ イ ド は ,近 く の ク ロ ロ フ ィ ル に 励 起 エ ネ ル ギ ー を 渡 し ,ク ロ ロ フ ィ ル の 補 助 色 素 と し て 機 能 し て い る 。集 光 性 複 合 体 お よ び 反 応 中 心 複 合 体 に 結 合 す る カ ロ テ ノ イ ド は す べ て 近 傍 に ク ロ ロ フ ィ ル が 存 在 し て お り ,集 光 補 助 あ る い は 過 剰 エ ネ ル ギ ー の 散 逸
( 後 述 ) に 寄 与 し て い る と 考 え ら れ る 。
4-1. 光 ス ト レ ス か ら の 保 護 -エ ネ ル ギ ー の 消 去-
カ ロ テ ノ イ ド の 保 護 機 能 は ,分 子 の 励 起 状 態 を 緩 和 し て エ ネ ル ギ ー を 散 逸 さ せ る 作 用 と ,化 学 反 応 に よ っ て フ リ ー ラ ジ カ ル を 消 去 す る 作 用 に 大 別 さ れ る 。こ こ で は , 過 剰 エ ネ ル ギ ー の 散 逸 メ カ ニ ズ ム に つ い て 述 べ る 。
基 底 状 態 の ク ロ ロ フ ィ ル は 一 重 項 で あ る が , 励 起 す る と 三 重 項 状 態 へ 遷 移 す る 。 こ の と き ,ス ピ ン 交 換 反 応 な ど で 励 起 エ ネ ル ギ ー が 酸 素 分 子 へ 移 動 す る と 一 重 項 酸 素 が 発 生 す る 。三 重 項 励 起 状 態 で は 、カ ロ テ ノ イ ド の エ ネ ル ギ ー 準 位 は ク ロ ロ フ ィ ル の エ ネ ル ギ ー 準 位 よ り 低 い た め ,ク ロ ロ フ ィ ル か ら カ ロ テ ノ イ ド へ の エ ネ ル ギ ー 移 動 が 起 こ り ク ロ ロ フ ィ ル の 励 起 状 態 は 緩 和 さ れ る 。一 方 ,ク ロ ロ フ ィ ル か ら エ ネ ル ギ ー を 受 け 取 っ た カ ロ テ ノ イ ド は 励 起 後 に 熱 を 放 散 し て 基 底 状 態 へ と 戻 る 。こ れ に よ り ,ク ロ ロ フ ィ ル か ら 酸 素 分 子 へ の エ ネ ル ギ ー 移 動 が 妨 げ ら れ ,一 重 項 酸 素 の 発 生 が 抑 制 さ れ る 。一 方 ,カ ロ テ ノ イ ド は 一 重 項 酸 素 自 体 の 消 去 に も 寄 与 し て い る 。 エ ネ ル ギ ー 準 位 の 差 に よ り 一 重 項 酸 素 か ら カ ロ テ ノ イ ド へ の エ ネ ル ギ ー 移 動 が 起 こ り ,励 起 さ れ た カ ロ テ ノ イ ド は 熱 を 放 散 し て 基 底 状 態 へ と 戻 る 。カ ロ テ ノ イ ド の エ ネ ル ギ ー 準 位 は , 共 役 二 重 結 合 の 数 に 依 存 し て い る (Hudson et al. 1982)。有 機 溶 媒 中 で は ,カ ロ テ ノ イ ド の 共 役 二 重 結 合 数 が 9 以 上 に な る と ,一 重 項 酸 素 消 去 度 が 増 加 す る こ と が 知 ら れ て い る 。 こ れ は , 共 役 二 重 結 合 数 が 10 を 超 え る と , 一 重 項 酸 素 の 励 起 状 態 よ り も エ ネ ル ギ ー 準 位 が 下 が る た め で あ る (三 室 et al. 2006)。
キ サ ン ト フ ィ ル サ イ ク ル は ,ク ロ ロ フ ィ ル の 励 起 状 態 の 消 去 に 重 要 な 役 割 を 担 っ
H. Takahashi-5
て い る 。キ サ ン ト フ ィ ル サ イ ク ル は ,LHCIIに 結 合 す る ヴ ィ オ ラ キ サ ン チ ン が ア ン テ ラ キ サ ン チ ン を 経 て ゼ ア キ サ ン チ ン へ と 変 換 さ れ る 機 構 で ,こ れ に 伴 い エ ネ ル ギ ー 準 位 が 下 が り ク ロ ロ フ ィ ル か ら 励 起 エ ネ ル ギ ー を 受 け 取 る (Demmig-Adams and Adams 1992, Yamamoto 1979, Yamamoto et al. 1962)。 そ の た め , 過 剰 な 励 起 エ ネ ル ギ ー が 熱 と し て 消 去 さ れ る 。こ の キ サ ン ト フ ィ ル 変 換 反 応 は ,ヴ ィ オ ラ キ サ ン チ ン・
デ エ ポ キ シ ダ ー ゼ に よ り 触 媒 さ れ ,そ の 酵 素 活 性 は チ ラ コ イ ド 膜 ル ー メ ン の 酸 性 化 に よ っ て 活 性 化 す る 。
シ ア ノ バ ク テ リ ア で は ,OCP が 過 剰 な 励 起 エ ネ ル ギ ー の 散 逸 を 担 っ て い る (Kirilovsky and Kerfeld 2012, Wilson et al. 2006)。 エ ネ ル ギ ー 散 逸 機 構 は , キ サ ン ト フ ィ ル サ イ ク ル と 同 様 に 分 子 の 励 起 状 態 の 解 消 で あ る が ,ク ロ ロ フ ィ ル で は な く シ ア ノ バ ク テ リ ア 集 光 色 素 で あ る ビ リ ン 色 素 か ら 励 起 エ ネ ル ギ ー を 受 け 取 る 。OCP は , 結 合 し て い る 3'-ヒ ド ロ キ シ エ キ ネ ノ ン に よ り 光 を 吸 収 し ,460 - 480 nm で 吸 収 極 大 を 示 す 。こ の 光 吸 収 に よ っ て ,OCP は エ ネ ル ギ ー 散 逸 状 態 が 可 能 な 活 性 型 へ と 変 換 さ れ る 。 そ の 分 子 メ カ ニ ズ ム は 以 下 の 通 り で あ る :(i) 細 胞 質 中 に 存 在 す る 不 活 性 型 OCPが ,青 色 光 や 白 色 光 の 照 射 に よ り 活 性 化 へ 変 換 す る (Wilson et al. 2006)。活 性 型 OCP で は ,OCP 内 の カ ロ テ ノ イ ド の 配 位 ア ミ ノ 酸 が 変 化 し , そ れ に 伴 っ て N 末 端 ド メ イ ン の α ヘ リ ッ ク ス に 結 合 し て い た C 末 端 ド メ イ ン が 外 れ , 分 子 内 に 隠 れ て い た カ ロ テ ノ イ ド が 外 側 に 露 出 す る よ う な 構 へ と 変 化 す る (Leverenz et al.
2015)。 さ ら に こ の 構 変 化 で ,C 末 端 ド メ イ ン か ら 外 れ た N 末 端 ド メ イ ン が , 集 光 性 複 合 体 フ ィ コ ビ リ ソ ー ム の コ ア 部 分 に あ る ア ロ フ ィ コ シ ア ニ ン 3 量 体 の シ リ ン ダ ー 構 に 入 り 込 ん で 結 合 す る (Harris et al. 2016)。 こ の と き , カ ロ テ ノ イ ド は ビ リ ン 色 素 へ 接 近 す る こ と で 励 起 状 態 を 解 消 し , 過 剰 エ ネ ル ギ ー が PSII 反 応 中 心 へ 移 動 す る の を 抑 制 し て い る と 考 え ら れ る (Leverenz et al. 2015)。 こ の エ ネ ル ギ ー 消 去 機 構 は 可 逆 的 で あ り , 細 胞 内 で は fluorescence recovery protein に よ っ て 不 活 性 型 へ と 戻 る (Boulay et al. 2010)。
集 光 性 複 合 体 で 得 た 光 エ ネ ル ギ ー 量 の 指 標 と し て ,ク ロ ロ フ ィ ル 蛍 光 収 率 が 用 い ら れ る 。光 エ ネ ル ギ ー が 光 化 学 反 応 に 用 い ら れ る と ク ロ ロ フ ィ ル 蛍 光 収 率 は 低 下 し
「 消 光 状 態 」と な る 。光 化 学 反 応 以 外 の 要 因 で ク ロ ロ フ ィ ル 蛍 光 収 率 が 低 下 す る 現 象 は ,非 光 化 学 的 消 光(nonphotochemical quenching, NPQ)と 呼 ば れ る 。キ サ ン ト フ ィ ル サ イ ク ル や OCP に よ る 光 エ ネ ル ギ ー の 消 失 は , 励 起 エ ネ ル ギ ー の 熱 変 換 で あ り , 光 化 学 反 応 に よ る 励 起 エ ネ ル ギ ー の 消 失 で は な い た め NPQ に 分 類 さ れ る (Niyogi and Truong 2013)。一 般 的 に ,強 光 条 件 で は NPQ が 高 く な り ,光 化 学 系 の 保 護 に 重 要 な 役 割 を 担 っ て い る こ と が 知 ら れ て い る 。
4-2. 光 ス ト レ ス か ら の 保 護 -フ リ ー ラ ジ カ ル の 消 去-
前 章 で は 化 学 反 応 を 伴 わ な い 励 起 エ ネ ル ギ ー の 消 去 に つ い て 述 べ た が ,こ こ で は 化 学 反 応 と し て カ ロ テ ノ イ ド が フ リ ー ラ ジ カ ル を 消 去 す る メ カ ニ ズ ム に つ い て 述 べ る 。
H. Takahashi-6
強 光 下 で は ,光 合 成 電 子 伝 達 反 応 に お い て 過 剰 量 の 電 子 が 生 成 さ れ ,そ の 一 部 が 酸 素 を 還 元 す る こ と に よ り 活 性 酸 素 が 生 じ る 。酸 素 分 子 が 一 電 子 還 元 さ れ る と ス ー パ ー オ キ サ イ ド (O2-) , ス ー パ ー オ キ サ イ ド が さ ら に 還 元 さ れ る と 過 酸 化 水 素
(H2O2),フ ェ ン ト ン 反 応 に よ り 過 酸 化 水 素 が 還 元 さ れ る と ヒ ド ロ キ シ ル ラ ジ カ ル
(OH・)が 生 成 す る 。カ ロ テ ノ イ ド は ,こ れ ら の ラ ジ カ ル と 電 子 の 授 受 を 行 っ て ラ ジ カ ル を 消 去 す る こ と が で き る (El-Agamey et al. 2004)。 し か し , ラ ジ カ ル 消 去 の 機 構 や 効 率 に つ い て は 不 明 な 点 が 多 い 。ま た ,カ ロ テ ノ イ ド は ラ ジ カ ル に よ る 不 飽 和 脂 肪 酸 の 過 酸 化 を 抑 制 す る (El-Agamey et al. 2004)。
5. カ ロ テ ノ イ ド に よ る 光 化 学 系 の 保 護
カ ロ テ ノ イ ド は ,強 光 下 で 励 起 ク ロ ロ フ ィ ル や 一 重 項 酸 素 を 消 光 し た り,光 合 成 電 子 伝 達 反 応 か ら 生 じ た 活 性 酸 素 を 消 去 し た り し て 酸 化 ス ト レ ス 傷 害 を 防 い で い る と 考 え ら れ る 。こ の 機 能 は ,具 体 的 に 光 合 成 の ど の プ ロ セ ス で 光 酸 化 ス ト レ ス か ら の 保 護 に 寄 与 し て い る の だ ろ う か 。強 光 下 で は 光 合 成 活 性 が 容 易 に 低 下 す る 。こ の 現 象 は 光 阻 害 と 呼 ば れ , 主 に 光 感 受 性 の 高 い PSIIの 失 活 が 原 因 と な る 。
5-1. PSII の 光 阻 害 と 活 性 酸 素 の 関 係
PSII の 光 阻 害 機 構 に は ,い く つ か 仮 説 が あ る 。例 え ば ,ア ク セ プ タ ー サ イ ド 説 で は ,強 光 下 で PSII の 反 応 中 心 で 発 生 し た 一 重 項 酸 素 が ,PSII 反 応 中 心 の D1 タ ン パ ク 質 を 直 接 損 傷 し,PSII を 失 活 さ せ る と 考 え ら れ て い る (Hideg et al. 1994, Keren et al. 1997, Vass et al. 1992)。こ の 場 合 ,一 重 酸 素 発 生 抑 制 に 寄 与 す る カ ロ テ ノ イ ド は , PSII の 損 傷 緩 和 に 寄 与 す る こ と が 考 え ら れ る 。一 方 ,近 年 提 唱 さ れ て き た Two-step 説 で は ,水 分 解 反 応 を 触 媒 す る 酸 素 発 生 複 合 体 マ ン ガ ン ク ラ ス タ ー が 直 接 光( 特 に UV や 青 色 光 )を 吸 収 し て 損 傷 し ,そ れ が 引 き 金 と な っ て PSII の 反 応 中 心 が 損 傷 す る こ と が 考 え ら れ て い る (Murata and Nishiyama 2018, Nishiyama and Murata 2014, Ohnishi et al. 2005)。Two-step 説 お よ び そ の 関 連 研 究 か ら , 活 性 酸 素 は PSII に 直 接 損 傷 を 及 ぼ す の で は な く,PSII の 修 復 を 阻 害 す る こ と が 示 さ れ て い る 。PSII は ,強 光 下 で D1 タ ン パ ク 質 が 損 傷 を 受 け る が ,修 復 機 構 に よ っ て や か に 修 復 さ れ 活 性 を 維 持 し て い る 。具 体 的 に は ,損 傷 を 受 け た D1 タ ン パ ク 質 の 分 解 ,D1 タ ン パ ク 質 の 新 規 合 成 ,D1 タ ン パ ク 質 の PSII 複 合 体 へ 挿 入 と プ ロ セ シ ン グ を 経 て PSII が 修 復 さ れ る (Jarvi et al. 2015, Murata and Nishiyama 2018, Nishiyama and Murata 2014)。
D1 タ ン パ ク 質 は,チ ラ コ イ ド 膜 上 に 結 合 し た リ ボ ソ ー ム で ペ プ チ ド が 伸 長 さ れ な が ら PSII 複 合 体 に 挿 入 さ れ る (Tyystjarvi et al. 2001)。 シ ア ノ バ ク テ リ ア Synechocystis sp. PCC 6803( 以 下 Synechocystis)で は ,強 光 下 で の D1 タ ン パ ク 質 の 新 規 合 成 は,ス ー パ ー オ キ サ イ ド や 過 酸 化 水 素 ,一 重 項 酸 素 に よ っ て 翻 訳 伸 長 反 応 の 過 程 で 阻 害 さ れ る こ と が 示 さ れ て い る (Nishiyama et al. 2004, Nishiyama et al.
2001)。さ ら に,翻 訳 伸 長 反 応 に 関 与 す る 翻 訳 因 子 EF-G お よ び EF-Tu が 活 性 酸 素 に よ っ て 特 定 の シ ス テ イ ン 残 基 が 酸 化 さ れ 失 活 す る (Kojima et al. 2009, Kojima et al.
H. Takahashi-7
2007, Nagano et al. 2012, Yutthanasirikul et al. 2016)。 加 え て , こ れ ら の 翻 訳 因 子 の 酸 化 標 的 シ ス テ イ ン 残 基 を セ リ ン に 置 換 す る と , D1 タ ン パ ク 質 の 新 規 合 成 が 促 進 し , 光 阻 害 が 緩 和 す る (Ejima et al. 2012, Jimbo et al. 2018)。 ま た,活 性 酸 素 消 去 系 酵 素 ス ー パ ー オ キ シ ド ジ ス ム タ ー ゼお よ び カ タ ラ ー ゼを Synechococcus elongates PCC 7942 で 過 剰 発 現 す る と ,強 光 下 で D1 タ ン パ ク 質 の 新 規 合 成 が 促 進 し ,PSII の 光 阻 害 が 緩 和 す る (Sae-Tang et al. 2016)。 こ れ ら の 研 究 か ら , タ ン パ ク 質 合 成 を 酸 化 傷 害 か ら 防 御 す る こ と が ,PSIIの 光 防 御 に 重 要 で あ る と 考 え ら れ る 。
5-2.光 阻 害 と カ ロ テ ノ イ ド の 関 係
前 述 し た よ う に ,カ ロ テ ノ イ ド は 光 化 学 系 複 合 体 の 形 成 や 保 護 に 重 要 で あ る 。β- カ ロ テ ン を 蓄 積 で き な い Synechocystis は,機 能 的 な PSII 複 合 体 を 形 成 で き ず ,光 独 立 栄 養 条 件 で は 生 育 で き な い (Sozer et al. 2010)。Synechocystis で は ,エ キ ネ ノ ン と ゼ ア キ サ ン チ ン は ,そ れ ぞ れ β-カ ロ テ ン の ケ ト 化,β-カ ロ テ ン へ の 水 酸 基 の 付 加 に よ り 合 成 さ れ る (Takaichi and Mochimaru 2007)。こ の 合 成 は そ れ ぞ れ ,β-カ ロ テ ン ケ ト 化 酵 素(CrtO),β-カ ロ テ ン 水 酸 化 酵 素(CrtR)に よ り 触 媒 さ れ る 。CrtO 欠 損 株(crtO)で は エ キ ネ ノ ン が 検 出 限 界 以 下 ま で 減 少 し,CrtR 欠 損 株(crtR)で は ゼ ア キ サ ン チ ン が 検 出 限 界 以 下 ま で 減 少 す る (Kusama et al. 2015, Schafer et al.
2005)。強 光 下 で 培 養 す る と,crtO,crtR,crtOcrtR 二 重 欠 損 株 の い ず れ に お い て も 光 合 成 活 性 が 低 下 す る が ,特 に crtR,crtOcrtR に お け る 活 性 低 下 は 著 し い (Schafer et al. 2005).。近 年 , こ の よ う な カ ロ テ ノ イ ド 合 成 酵 素 欠 損 株 の 光 合 成 活 性 低 下 は , PSII の 光 阻 害 に 起 因 す る こ と が 明 ら か に さ れ て い る (Kusama et al. 2015)。 さ ら に , PSII の 光 阻 害 は , 光 損 傷 の 促 進 で は な く 修 復 能 力 の 低 下 が 原 因 と な っ て い る こ と や , 強 光 下 で D1 タ ン パ ク 質 の 新 規 合 成 が 低 下 す る こ と が わ か っ て い る 。 ま た crtOcrtR で は ,強 光 下 で の 一 重 項 酸 素 の 発 生 が 著 し く 増 加 し た こ と か ら ,カ ロ テ ノ イ ド は 一 重 項 酸 素 の 発 生 を 抑 制 し , タ ン パ ク 質 合 成 の 酸 化 傷 害 を 防 い で PSII の 光 阻 害 を 緩 和 し て い る こ と が 考 え ら れ る (Kusama et al. 2015)。
OCP に よ る 光 エ ネ ル ギ ー 散 逸 も , カ ロ テ ノ イ ド に よ る 光 化 学 系 の 保 護 の 一 例 と し て 挙 げ ら れ る 。OCP は 結 合 す る カ ロ テ ノ イ ド を 介 し て , 集 光 性 複 合 体 フ ィ コ ビ リ ソ ー ム に お い て 過 剰 エ ネ ル ギ ー を 散 逸 す る (El Bissati et al. 2000, Wilson et al.
2006)。OCPを 欠 損 し た Synechocystis 変 異 株 で は 光 エ ネ ル ギ ー の 散 逸 活 性 が 減 少 し て お り (Wilson et al. 2006), 強 光 下 で 誘 導 さ れ る NPQ が 低 下 す る (Kusama et al.
2015)。OCPを 欠 損 す る と PSII の 修 復 能 力 が 低 下 し , 光 阻 害 が 促 進 す る (Kusama et al. 2015)。 ま た OCP欠 損 株 で は , 強 光 下 で 一 重 項 酸 素 の 発 生 量 が 増 加 し
(Kusama et al. 2015, Sedoud et al. 2014),OCP 過 剰 発 現 株 で は , 光 エ ネ ル ギ ー の 散 逸 活 性 が 大 幅 に 増 加 す る こ と が 報 告 さ れ て い る (Wilson et al. 2008)。OCP 過 剰 発 現 株 で は NPQ の 増 加 や PSII の 光 阻 害 の 緩 和 ,D1 タ ン パ ク 質 の 新 規 合 成 の 促 進 が 見 ら れ て い る ( 投 稿 準 備 中 ) 。 し た が っ て ,OCP に よ る 過 剰 エ ネ ル ギ ー の 熱 放 散
H. Takahashi-8
お よ び 一 重 項 酸 素 の 発 生 抑 制 は , タ ン パ ク 質 合 成 を 酸 化 傷 害 か ら 保 護 し ,PSII の 修 復 を 促 進 し て PSII の 光 防 御 に 寄 与 す る こ と が 考 え ら れ る 。
6. お わ り に
カ ロ テ ノ イ ド は 集 光 色 素 と し て 機 能 し て い る だ け で な く ,光 合 成 を 行 う 上 で 避 け る こ と の で き な い 光 酸 化 ス ト レ ス の 軽 減 に も 役 立 っ て い る( 図 2)。カ ロ テ ノ イ ド に よ る 三 重 項 ク ロ ロ フ ィ ル や 一 重 項 酸 素 の 励 起 状 態 の 緩 和 は ,光 化 学 系 ,特 に PSII の 光 防 御 に 寄 与 す る こ と が 知 ら れ て い る 。こ れ ま で カ ロ テ ノ イ ド が 一 重 項 酸 素 の 発 生 を 抑 制 し て PSII の 光 損 傷 を 防 ぐ と 考 え ら れ て き た が , 近 年 , カ ロ テ ノ イ ド が タ ン パ ク 質 合 成 を 一 重 項 酸 素 に よ る 酸 化 傷 害 か ら 保 護 し , 強 光 下 で PSII の 修 復 を 促 進 し て 光 阻 害 を 緩 和 す る こ と が 示 唆 さ れ て い る 。今 後 ,そ の 保 護 作 用 の 詳 細 な メ カ ニ ズ ム 解 明 が 期 待 さ れ る 。
謝 辞
本 研 究 は JST未 来 社 会 創 事 業「 ゲ ー ム チ ェ ン ジ ン グ テ ク ノ ロ ジ ー に よ る 低 炭 素 社 会 の 実 現 :B21 大 規 模 生 産 に 向 け て 環 境 に ロ バ ス ト な 微 細 藻 類 の 開 発 」, お よ び JSPS 科 研 費 JP18K06276 (Y.N.), JP18K06275 (H.T.)の 助 成 を 受 け た も の で す 。
図 2. 光 合 成 に お け る カ ロ テ ノ イ ド の 機 能
カ ロ テ ノ イ ド は , ク ロ ロ フ ィ ル の 補 助 色 素 と し て 集 光 機 能 を 担 う 。 加 え て , 三 重 項 ク ロ ロ フ ィ ル や 一 重 項 酸 素 の 励 起 状 態 の 解 消 に よ る エ ネ ル ギ ー 散 逸 や , 過 酸 化 物 の 発 生 抑 制 に 寄 与 す る 。 近 年 の 研 究 か ら , カ ロ テ ノ イ ド に よ る 活 性 酸 素 の 発 生 抑 制 が タ ン パ ク 質 合 成 の 酸 化 傷 害 を 防 御 し ,PSII の 光 阻 害 を 緩 和 す る 役 割 を 担 っ て い る こ と が 示 唆 さ れ て い る 。
H. Takahashi-9 引 用 文 献
Amunts A, Drory O, & Nelson N. 2007. The structure of a plant photosystem I supercomplex at 3.4 Å resolution. Nature 447: 58-63.
Ben-Shem A, Frolow F, & Nelson N. 2003. Crystal structure of plant photosystem I. Nature 426: 630-635.
Boulay C, Wilson A, D'Haene S, & Kirilovsky D. 2010. Identification of a protein required for recovery of full antenna capacity in OCP -related photoprotective mechanism in cyanobacteria. Proc. Natl. Acad. Sci. USA 107: 11620-11625.
Demmig-Adams B & Adams WI. 1992. Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant. Phys. 43: 599-626.
Ejima K, Kawaharada T, Inoue S, Kojima K, & Nishiyama Y. 2012. A change in the sensitivity of elongation factor G to oxidation protect s photosystem II from photoinhibition in Synechocystis sp. PCC 6803. FEBS lett. 586: 778-783.
El Bissati K, Delphin E, Murata N, Etienne A, & Kirilovsky D. 2000. Photosystem II fluorescence quenching in the cyanobacterium Synechocystis PCC 6803: involvement of two different mechanisms. Biochim. Biophys. Acta 1457: 229-242.
El-Agamey A, Lowe GM, McGarvey DJ, Mortensen A, Phillip DM, Truscott TG, & Young AJ. 2004. Carotenoid radical chemistry and antioxidant/pro -oxidant properties. Arch.
Biochem. Biophys. 430: 37-48.
Ferreira KN, Iverson TM, Maghlaoui K, Barber J, & Iwata S. 2004. Architecture of the photosynthetic oxygen-evolving center. Science 303: 1831-1838.
Harris D, Tal O, Jallet D, Wilson A, Kirilovsky D, & Adir N. 2016. Orange carotenoid protein burrows into the phycobilisome to provide photoprotection. Proc. Natl. Acad. Sci.
USA 113: E1655-1662.
Hideg E, Spetea C, & Vass I. 1994. Singlet oxygen production in thylakoid membranes during photoinhibition as detected by EPR spectroscopy. Photosynth Res. 39: 191-199.
Holt TK & Krogmann DW. 1981. A carotenoid-protein from cyanobacteria. Biochim.
Biophys. Acta 637: 408–414.
Hudson BS, Kohler BE, & Schulten K. 1982. Linear polyene electronic -structure and potential surfaces. Excited States 6: 1–95.
Jarvi S, Suorsa M, & Aro EM. 2015. Photosystem II repair in plant chloroplasts--regulation, assisting proteins and shared components with photosystem II biogenesis. Biochim.
Biophys. Acta 1847: 900-909.
Jimbo H, Yutthanasirikul R, Nagano T, Hisabori T, Hihara Y, & Nishiyama Y. 2018.
Oxidation of translation factor EF-Tu Inhibits the repair of photosystem II. Plant Physiol.
in press:
Jordan P, Fromme P, Witt HT, Klukas O, Saenger W, & Krauss N. 2001. Three -dimensional
H. Takahashi-10
structure of cyanobacterial photosystem I at 2. 5 Å resolution. Nature 411: 909-917.
Kamiya N & Shen JR. 2003. Crystal structure of oxygen -evolving photosystem II from Thermosynechococcus vulcanus at 3.7-A resolution. Proc. Natl. Acad. Sci. USA 100: 98- 103.
Keren N, Berg A, van Kan PJ, Levanon H, & Ohad I. 1997. Mechanism of photosystem II photoinactivation and D1 protein degradation at low light: the role of back electron flow.
Proc. Natl. Acad. Sci. USA 94: 1579-1584.
Kerfeld CA, Sawaya MR, Brahmandam V, Cascio D, Ho KK, Trevithick -Sutton CC, Krogmann DW, & Yeates TO. 2003. The crystal structure of a cyanobacterial water - soluble carotenoid binding protein. Structure 11: 55-65.
Kirilovsky D & Kerfeld CA. 2012. The orange carotenoid protein in photoprotection of photosystem II in cyanobacteria. Biochim. Biophys. Acta. 1817: 158-166.
Kojima K, Motohashi K, Morota T, Oshita M, Hisabori T, Hayashi H, & Ni shiyama Y.
2009. Regulation of translation by the redox state of elongation factor G in the cyanobacterium Synechocystis sp. PCC 6803. J. Biol. Chem. 284: 18685-18691.
Kojima K, Oshita M, Nanjo Y, Kasai K, Tozawa Y, Hayashi H, & Nishiyama Y. 2007.
Oxidation of elongation factor G inhibits the synthesis of the D1 protein of photosystem II. Mol. Microbiol. 65: 936-947.
Kurisu G, Zhang H, Smith JL, & Cramer WA. 2003. Structure of the cytochrome b6f complex of oxygenic photosynthesis: tuning the cavity. Science 302: 1009-1014.
Kusama Y, Inoue S, Jimbo H, Takaichi S, Sonoike K, Hihara Y, & Nishiyama Y. 2015.
Zeaxanthin and echinenone protect the repair of photosys tem II from inhibition by singlet oxygen in Synechocystis sp. PCC 6803. Plant Cell Physiol. 56: 906-916.
Leverenz RL, Sutter M, Wilson A, Gupta S, Thurotte A, Bourcier de Carbon C, Petzold CJ, Ralston C, Perreau F, Kirilovsky D, & Kerfeld CA. 2015. A 12 Å carotenoid translocation in a photoswitch associated with cyanobacterial photoprotection. Science 348: 1463-1466.
Liu Z, Yan H, Wang K, Kuang T, Zhang J, Gui L, An X, & Chang W. 2004. Crystal structure of spinach major light-harvesting complex at 2.72 A resolution. Nature 428: 287-292.
Loll B, Kern J, Saenger W, Zouni A, & Biesiadka J. 2005. Towards complete cofactor arrangement in the 3.0 Å resolution structure of photosystem II. Nature 438: 1040-1044.
Melnicki MR, Leverenz RL, Sutter M, Lopez-Igual R, Wilson A, Pawlowski EG, Perreau F, Kirilovsky D, & Kerfeld CA. 2016. Structure, diversity, and evolution of a new family of soluble carotenoid-binding proteins in Cyanobacteria. Mol. Plant 9: 1379-1394.
三 室 守, 高 市 真 一, & 富 田 純 史 2006. カ ロ テ ノ イ ド ー そ の 多 様 性 と 生 理 活 性 ー. 裳 華 房, 東 京
Minagawa J & Takahashi Y. 2004. Structure, function and assembly of photosystem II and its light-harvesting proteins. Photosynth. Res. 82: 241-263.
Murata N & Nishiyama Y. 2018. ATP is a driving force in the repair of photosystem II
H. Takahashi-11 during photoinhibition. Plant Cell Environ. 41: 285-299.
Nagano T, Kojima K, Hisabori T, Hayashi H, Morita EH, Kanamori T, Miyagi T, Ueda T,
& Nishiyama Y. 2012. Elongation factor G is a critical target during oxidative damage to the translation system of Escherichia coli. J. Biol. Chem. 287: 28697-28704.
Nelson N & Yocum CF. 2006. Structure and function of photosystem I and II. Annu. Rev.
Plant Biol. 57: 521-565.
Nield J, Kruse O, Ruprecht J, da Fonseca P, Buchel C, & Barber J. 2000. Three - dimensional structure of Chlamydomonas reinhardtii and Synechococcus elongatus photosystem II complexes allows for comparison of their oxygen -evolving complex organization. J. Biol. Chem. 275: 27940-27946.
Nishiyama Y, Allakhverdiev SI, Yamamoto H, Hayashi H, & Murata N. 2004. Singlet oxygen inhibits the repair of photosystem II by suppressing the translation elongation of the D1 protein in Synechocystis sp. PCC 6803. Biochemistry 43: 11321-11330.
Nishiyama Y & Murata N. 2014. Revised scheme for the mechanism of photoinhibition and its application to enhance the abiotic stress tolerance of the photosynthetic machinery.
Appl Microbiol Biotechnol. 98: 8777-8796.
Nishiyama Y, Yamamoto H, Allakhverdiev SI, Inaba M, Yokota A, & Murata N. 2001.
Oxidative stress inhibits the repair of photodamage to the photosynthetic machinery.
EMBO J. 20: 5587-5594.
Niyogi KK & Truong TB. 2013. Evolution of flexible non -photochemical quenching mechanisms that regulate light harvesting in oxygenic photosynthesis. Curr. Opin. Plant Biol. 16: 307-314.
Ohnishi N, Allakhverdiev SI, Takahashi S, Higashi S, Watanabe M, Nishiyama Y, &
Murata N. 2005. Two-step mechanism of photodamage to photosystem II: Step 1 occurs at the oxygen-evolving complex and step 2 occurs at the photochemical reaction center.
Biochemistry 44: 8494-8499.
Punginelli C, Wilson A, Routaboul JM, & Kirilovsky D. 2009. Influence of zeaxanthin and echinenone binding on the activity of the orange carotenoid protein. Biochim. Biophys.
Acta 1787: 280-288.
Qin XC, Suga M, Kuang TY, & Shen JR. 2015. Structural basis for energy transfer pathways in the plant PSI-LHCI supercomplex. Science 348: 989-995.
Sae-Tang P, Hihara Y, Yumoto I, Orikasa Y, Okuyama H, & Nishiyama Y. 2016.
Overexpressed superoxide dismutase and catalase act synergist ically to protect the repair of PSII during photoinhibition in Synechococcus elongatus PCC 7942. Plant Cell Physiol.
57: 1899-1907.
Schafer L, Vioque A, & Sandmann G. 2005. Functional in situ evaluation of photosynthesis-protecting carotenoids in mutants of the cyanobacterium Synechocystis PCC6803. J. Photochem. Photobiol. B: Biol. 78: 195-201.
H. Takahashi-12
Sedoud A, Lopez-Igual R, Ur Rehman A, Wilson A, Perreau F, Boulay C, Vass I, Krieger - Liszkay A, & Kirilovsky D. 2014. The cyanobacterial photoactive orange carotenoi d protein Is an excellent singlet oxygen quencher. Plant Cell 26: 1781-1791.
Sozer O, Komenda J, Ughy B, Domonkos I, Laczko -Dobos H, Malec P, Gombos Z, & Kis M. 2010. Involvement of carotenoids in the synthesis and assembly of protein subunits of photosynthetic reaction centers of Synechocystis sp PCC 6803. Plant Cell Physiol. 51:
823-835.
Stroebel D, Choquet Y, Popot JL, & Picot D. 2003. An atypical haem in the cytochrome b6f complex. Nature 426: 413-418.
Takaichi S & Mochimaru M. 2007. Carotenoids and car otenogenesis in cyanobacteria:
unique ketocarotenoids and carotenoid glycosides. Cell Mol. Life Sci. 64: 2607-2619.
Tyystjarvi T, Herranen M, & Aro EM. 2001. Regulation of translation elongation in cyanobacteria: membrane targeting of the ribosome nascent -chain complexes controls the synthesis of D1 protein. Mol. Microbiol. 40: 476-484.
Umena Y, Kawakami K, Shen JR, & Kamiya N. 2011. Crystal stru cture of oxygen-evolving photosystem II at a resolution of 1.9 A. Nature 473: 55-60.
Vass I, Styring S, Hundal T, Koivuniemi A, Aro E, & Andersson B. 1992. Reversible and irreversible intermediates during photoinhibition of photosystem II: stable reduced Q A species promote chlorophyll triplet formation. Proc. Natl. Acad. Sci. USA 89: 1408-1412.
Wilson A, Ajlani G, Verbavatz JM, Vass I, Kerfeld CA, & Kirilovsky D. 2006. A soluble carotenoid protein involved in phycobilisome -related energy dissipation in cyanobacteria.
Plant Cell 18: 992-1007.
Wilson A, Kinney JN, Zwart PH, Punginelli C, D'Haene S, Perreau F, Klein MG, Kirilovsky D, & Kerfeld CA. 2010. Structural determinants underlying photoprotection in the photoactive orange carotenoid protein of cyanobacte ria. J. Biol. Chem. 285: 18364- 18375.
Wilson A, Punginelli C, Couturier M, Perreau F, & Kirilovsky D. 2011. Essential role of two tyrosines and two tryptophans on the photoprotection activity of the orange carotenoid protein. Biochim. Biophys. Acta 1807: 293-301.
Wilson A, Punginelli C, Gall A, Bonetti C, Alexandre M, Routaboul JM, Kerfeld CA, van Grondelle R, Robert B, Kennis JT, & Kirilovsky D. 2008. A photoactive carotenoid protein acting as light intensity sensor. Proc. Natl. Acad. Sci. USA 105: 12075-12080.
Yamamoto HY. 1979. Biochemistry of the violaxanthin cycle in higher -plants. Pure Appl.
Chem. 51: 639-648.
Yamamoto HY, Nakayama TO, & Chichester CO. 1962. Studies on the light and dark interconversions of leaf xanthophylls. Arch. Biochem. Biophys. 97: 168-173.
Yutthanasirikul R, Nagano T, Jimbo H, Hihara Y, Kanamori T, Ueda T, Haruyama T, Konno H, Yoshida K, Hisabori T, & Nishiyama Y. 2016. Oxidation of a cysteine residue in
H. Takahashi-13
elongation factor EF-Tu reversibly inhibits translation in the cyanobacterium Synechocystis sp. PCC 6803. J. Biol. Chem. 291: 5860-5870.
S. Takaichi-1
光合成細菌,シアノバクテリア,藻類,植物における 多様なカロテノイド分子種と生合成経路の分布
高市 真一
東京農業大学 生命科学部 分子微生物学科
〒156-8502 世田谷区桜丘
Shinichi Takaichi
Diversities of carotenoid species and carotenogenesis pathways among phototrophs; photosynthetic bacteria, cyanobacteria, alga, and land plants
Keywords: carotenoid, carotenogenesis, diversity phototroph
Department of Molecular Microbiology, Faculty of Life Science, Tokyo University of Agriculture Sakuragaoka, Setagaya, Tokyo 156-8502, Japan
DOI: 10.24480/bsj-review.9b3.00135
光合成生物は必ず色素として(バクテリオ)クロロフィルとカロテノイドを持っている。(バクテリ オ)クロロフィルの構造や合成系,光化学系には類似性が見られる。一方,カロテノイドは分類群ご とに組成や合成経路に大きな差異が見られるので,進化・共生に伴い一部の合成経路を捨てて,一部 の合成経路を共生相手とは別の生物から取り込んだと思われる。本総説では,フィトエンからリコペ ンへの合成経路および酵素,異種のリコペン・シクラーゼの分布,分類群ごとのカロテノイド合成を まとめた。
1.はじめに
光合成生物にとっては(バクテリオ)クロロフィルだけでなくカロテノイドも必須の成分である。
カロテノイドは光合成のための光捕集,強光や活性酸素種からクロロフィルや光化学系などの防御・
保護,色素タンパク複合体の形成・安定化,などの機能を担っている。光合成生物の分類と存在する カロテノイド分子種の基本型や合成酵素の一部を表1にまとめた。光化学系II型をもつ光合成細菌と 光化学系I型をもつ光合成細菌から,酸素発生型のシアノバクテリアができ,そのシアノバクテリア が真核生物に一次共生をして灰色藻類・紅藻類・緑藻類の葉緑体になったと考えられている。その後 の二次および三次共生により種々の藻類や陸上植物が出現したと考えられている。さらに光合成の光 化学系を構成するペプチド,(バクテリオ)クロロフィルの構造や生合成経路,三次構造には,進化的 には一部の成分の置き換えはあるが,類似性と連続性が見られる。一方,カロテノイドは生物分類ご とに一部に共通性は見られるが,共生段階などが変わるごとに組成ばかりでなく,生合成経路や酵素 に大きな変化が見られる。従って,進化や共生に伴い一部のカロテノイド合成経路を捨てて,一部の 合成経路を共生相手とは別の生物から取り込んだと思われる。カロテノイドは結合タンパクや機能に 対して柔軟性が高いためだろう。本総説ではカロテノイド合成系の変化や進化について紹介する。た だし,全体像を見るために,一部の例外的な存在は省略した。
S. Takaichi-2 2.フィトエンからリコペンの合成
15-シス-フィトエンから全トランス-リコペン
の合成には4 段階の不飽和化反応が必要である
(図 1)。原核生物はフィトエン不飽和化酵素
phytoene desaturase (CrtI) [注:下記のように複数 の略語をもつことがある]が担っている。紅色細
菌(図 2)とクロロフレクサス(Harada et al.,
unpublished)ではこの酵素の機能が確認されてお り,ヘリオバクテリアには crtI に相同性がある 遺伝子crtNがあるがまだ機能が確認されていな い(表1)。紅色細菌のRhodobacterなどはCrtIが 最後の4 段階目の不飽和化をできないため,リ コペンではなくニューロスポレンが最終産物と なりニューロスポレンからスフェロイデンなど を合成する(図2)。ヘリオバクテリアは鎖状の
C30-ジアポニューロスポレンが最終産物である
(Takaichi et al. 1997)。シアノバクテリアの中では,
例外的に原始的なGloeobacter violaceusのみCrtIを使うが,他のカロテノイド合成酵素は他のシアノバ クテリアと共通性がある(Tsuchiya et al. 2005)。
原核生物においては緑色硫黄細菌とシアノバクテリアのみ,葉緑体と同様にフィトエンからζ-カロ テンまでをフィトエン不飽和化酵素phytoene desaturase (CrtP, Pds)[注:CrtIと同じ酵素名であるが機能 は異なる]が,ζ-カロテンからリコペンまでをζ-カロテン不飽和化酵素 -carotene desaturase (CrtQ, Zds) が各々2段階ずつ不飽和化する(図1)。さらにCrtQの反応に伴い合成されるポリシス型のニューロス ポレンとリコペンを,カロテン異性化酵素carotene isomerase (CrtH, CrtISO)あるいは光照射によりトラ 表1:生物の分類とカロテノイド(高市2006, Takaichi and Mochimaru 2007, Takaichi 2009, Takaichi 2011, Takaichi 2013)
光化学系 光合成生物 カロテノイド型
(基本型)
フィトエンから リコペン
リコペン・
シクラーゼ
II型 紅色細菌1 C40-鎖状 CrtI CrtY
クロロフレクサス β,γ-カロテン CrtI2 CrtY2 I型 ヘリオバクテリア C30-鎖状 CrtN (CrtI-type)? non
緑色硫黄細菌 β,γ-カロテン CrtP/Q/H3 CruA/P II+I型
(酸素発生型)
シアノバクテリア β,γ-カロテン CrtP/Q/H + Z-ISO3 CruA/P?
葉緑体 β,α-カロテン CrtP/Q/H + Z-ISO CrtL
1 カロテノイド合成遺伝子はクラスターを形成 2 Harada et al., unpublished, 3 Sugiyama et al., unpublished
図1:フィトエンからリコペンの合成(高市2006, Takaichi and Mochimaru 2007, Takaichi 2009, Takaichi 2011)
S. Takaichi-3
ンス型に変換する。植物においてはポリシス-ζ-カロテンをジシス-ζ-カロテンに変換するζ-カロテン異
性化酵素 -carotene isomerase (Z-ISO)も必要である(Li et al. 2007)。それに相同性のある遺伝子がシアノ
バクテリアにも存在し,最近になって機能確認された。また緑色硫黄細菌のゲノムにはZ-ISOに対応 する遺伝子が見いだされなかった (Sugiyama et al. unpublished)。従ってCrtP/Q型はリコペン合成に合 計3あるいは4種類の酵素を必用とする(表1、図1)。
crtP, crtQ, crtHはcrtIに低い相同性があるので,crtIから変化したのであろう。4種の酵素によるリ
コペン合成経路は,緑色硫黄細菌からシアノバクテリアに,さらに藻類や陸上植物の葉緑体に引き継 がれたと思われる。ただし細菌において1酵素(CrtI)が担っていた反応を,CrtP/Q型では4酵素に分割 し複雑化したが,そのための原動力やメリットがどこにあったのかは不明である。
3.リコペンの環化(β-カロテン,α-カロテン合成)
紅色細菌はリコペンを鎖状のまま修飾してスピリロキサンチンなどを合成する(図2)。多くのヘリ オバクテリアは鎖状カロテンが最終産物である(Takaichi et al. 1997)(表1)。
リコペン・シクラーゼlycopene cyclaseとして相同性のない3種類の酵素が知られている。多くの細 菌CrtYと葉緑体CrtLに見られる型,緑色硫黄細菌と一部のシアノバクテリアに見られるCruA/CruP 型,一部の細菌(CrtYc+CrtYd)とアーキア(CrtYcd)と菌類(CrtYB)には類似した別の型が存在している。
CrtYとCrtLにはある程度の相同性が見られるので我々は同一グループと考えているが,Bryantらは 2つに分けている(Maresca et al. 2007)。リコペンの片方を環化したγ-カロテンしかつくれないCrtYm
とCrtLmもあるが,各々CrtYやCrtLと相同性が高い。
紅色細菌の一種である好気性光合成細菌の一部とクロロフレクサスも-カロテンと β-カロテンを CrtYにより合成する(表1)(Harada et al. unpublished)。
図2:紅色光合成細菌のカロテノイドと合成経路(高市2006, Takaichi 2009) 酵素の性質の違いや変化また新たな獲得が合成経路や産物の多様性を生じた。
S. Takaichi-4
藻類や陸上植物では,リコペンの両端をリコペン-β-シクラーゼ (CrtL-b, Lcy-b) がβ末端基にしてβ- カロテンがつくられる。一方,α-カロテンは,基質特異性のために先ずリコペン-ε-シクラーゼ (CrtL-
e, Lcy-e) がε末端基を合成して,次いで反対側をCrtL-bがβ末端基にして合成される(Cunningham et
al. 2001)。2つの酵素は相同性が高く,CrtL-bからCrtL-eができたと考えられている。α-カロテンとそ
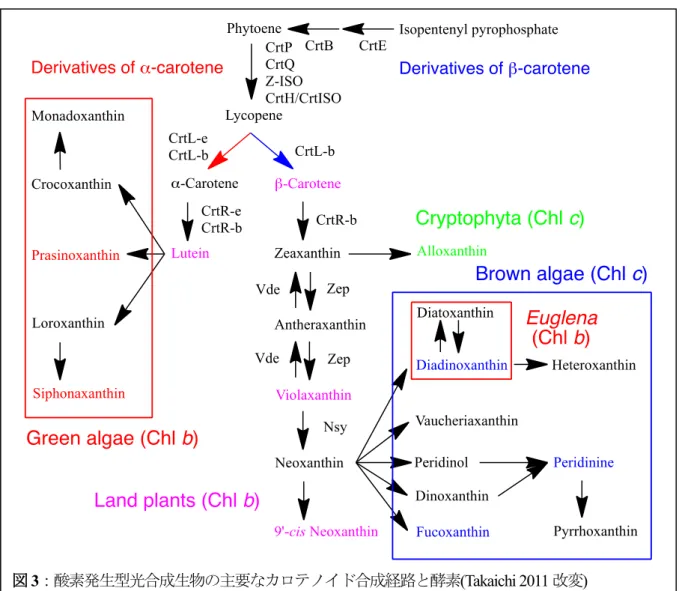
の誘導体は一部の紅藻類,クリプト藻類,緑藻類,陸上植物に限られていて,光合成細菌と2属を除 くシアノバクテリア(Takaichi et al. 2012),褐藻類,ユーグレナには存在しない(図3)。クリプト藻類 Guillardia thetaのα-カロテンはCrtL-b, CrtL-eにより合成される(Konno et al. unpublished)。
シアノバクテリアのリコペン・シクラーゼは未だに充分な機能解析がされていない。最初に Synechococcus sp. PCC 7942 のCrtL (Cunningham et al. 1994),次いで Prochlorococcus marinus のCrtL
(Stickforth et al. 2003)の機能が確認された。近年,別型のリコペン・シクラーゼCruAが緑色硫黄細菌
Chlorobaculum tepidum,CruAとCruPがSynechococcus sp. PCC 7002で機能が確認された(Maresca et al.
2007)。最近,Arthrospira platensisにおいてもCruAが機能確認された(Sugiyama et al. 2017)。さらに Synechocystis sp. PCC 6803ではCruAの活性発現に結合クロロフィルが必用であると報告された(Xiong
et al. 2017)。crtLあるいはcruA/cruPに相同性のある遺伝子が多くのシアノバクテリアのゲノム上に存
図3:酸素発生型光合成生物の主要なカロテノイド合成経路と酵素(Takaichi 2011改変)
分類群の文字色は同色のカロテノイドを主成分とする。
S. Takaichi-5
在するが,上記を除いては機能確認できていない。cruA/cruPがリコペン・シクラーゼの有力な候補と 思われるが,さらなる機能確認が必要である。
4.紅色細菌のカロテノイドと合成経路の多様性
紅色細菌のカロテノイドにも多様性が見られるが,合成酵素の性質の差異により合成経路に多様性 が生じたと考えられる(図2)(高市2006, Takaichi 2009)。上記のようにCrtIの最終産物の違いがスフ ェロイデン経路とスピリロキサンチン経路を作り出したが,メトキシ基合成は同じヒドロキシニュー ロスポレン合成酵素 hydroxyneurosporene synthase (CrtC),メトキシニューロスポレン不飽和化酵素 methoxyneurosporene desaturase (CrtD), ヒ ド ロ キ シ ニ ュ ー ロ ス ポ レ ン-O-メ チ ル 化 酵 素 hydroxyneurosporene-O-methyl transferase (CrtF)がどちらの経路でも働いている。
5.緑色硫黄細菌とクロロフレクサスのカロテノイドと合成系
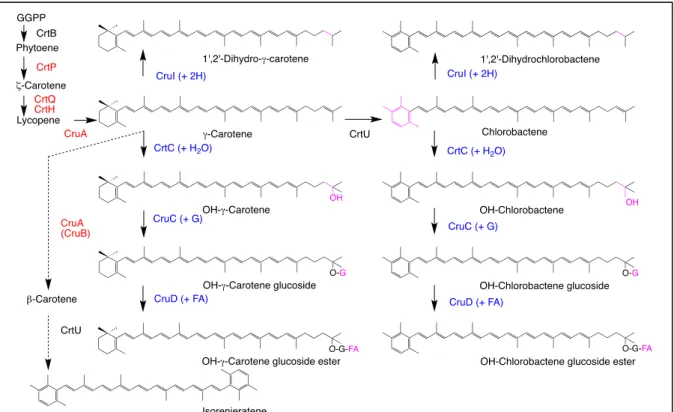
緑色硫黄細菌とクロロフレクサスの合成するカロテノイドとその合成経路はまだ充分には解析され ていない(図4)(高市2006)。緑色硫黄細菌Chlorobacula tepidumはOH-γ-カロテン・グルコシド・エ
ステル,OH-クロロバクテン・グルコシド・エステル,1,2-ジヒドロ-γ-カロテンなどを合成する(Takaichi
et al. 1997)。一部の合成酵素・遺伝子のうちCrtC,グルコース添加酵素glucosyltransferase (CruC),グル
コース脂肪酸添加酵素glucosyl esterase (CruD)は機能解析された(Frigaard et al. 2004)。また一部の種はβ 末端基を芳香環にしたイソレニエラテンをもつ。最近,カロテノイド-1,2-還元酵素 1,2-carotenoid- reductase (CruI)の機能確認がなされた(Canniffe et al. 2018)。
図4.緑色硫黄細菌Chlorobacula tepidumのカロテノイド合成経路(Takaichi et al. 1997, Maresca
et al. 2007, 改変) 赤青字は機能確認された酵素である。クロロフレクサスでもほぼ同じ酵素が働
いている
S. Takaichi-6
一方,クロロフレクサスChloroflexus aurantiacus はOH-γ-カロテン・グルコシド・エステルなどをも つ(Takaichi et al. 1995)。合成酵素としてCrtI, CrtY(表1)が機能しており,さらにCruC, CruDとCrtC の代わりにカロテノイド-1,2-水添加酵素carotenoid-1,2-hydratase (CruF)が機能している(図4)(Harada et al., unpublished)。
6.酸素発生型光合成生物のカロテノイドと合成経路
シアノバクテリア,紅藻類,クリプト藻類,褐藻類,緑藻類,陸上植物など酸素発生型光合成生物 の主なカロテノイド合成経路を図3にまとめた。リコペンからβ-カロテンとα-カロテンが合成され,
中央の経路にはそれらが変化し共通してみられるカロテノイドがあり,さらに左右に枝分かれた誘導 体合成経路があり,これらは生物の系統分類に関係している。
シアノバクテリアは β-カロテン誘導体のみをもち,β-カロテン水酸化酵素 -carotene hydroxylase (CrtR, BHY)や下記のケト化によりエキネノンや水酸化によりノストキサンチンを合成し,さらにシア ノバクテリアに特有なγ-カロテン誘導体のミクソール配糖体を合成する。いくつかの酵素の機能確認 がなされている(図5)。ただしゼアキサンチンから先のエキネノンやノストキサンチンの合成経路は,
葉緑体に引き継がれなかった(図3)。
シアノバクテリアにはケト化酵素として,互いに相同性のない -カロテンケトラーゼ -carotene ketolaseが2種類存在する(CrtO, CrtW) (図5)。Anabaena sp. PCC 7120ではこれらの酵素が働く代謝
図5.シアノバクテリアのカロテノイド合成経路(高市2006, Takaichi and Mochimaru 2007, Takaichi 2011)