論文 内 容の 要旨
ヒ ア ル ロ ン 酸(HA)は , N-ア セ チ ル グ ル コ サ ミ ン (NAG)と グ ル ク ロ ン 酸 (GlcA)の
二 糖 が 交 互 に 結 合 し た 高 分 子 多 糖 で, 哺 乳 動 物 の 結 合 組 織 に 広 く 存 在 す る グ リ コ サ ミ ノ グ リ カ ン の ひ と つ で あ る. その分子量は 107Daに も 及 び, 構成単糖のもつ水 和 能 に 加 え, オ ー バ ー ラ ッ プ し た 三 次 元 ネ ッ ト ワ ー ク が 水 分 子 を 束 縛 す る こ と に よ り , 高 い 水 和 体 積 を 獲 得 す る . HAは 水 和 ゲ ル と し て 組 織 の 水 分 保 持 を 担 い , ま た 組 織 に 機 械 的 特 性 を 賦 与 す る . 皮 膚 真 皮 に 存 在 す る HAは , 細 胞 外 マ ト リ ッ ク ス を 構 築 す る 分 子 と し て 機 能 す る 一 方 で , 表 皮 の HAは 重 層 し た 表 皮 細 胞 の 細 胞 間 隙 に 存 在 し て 細 胞 間 ス ペ ー ス を 維 持 し, 細 胞 の 増 殖 や 分 化 に 寄 与 す る. 皮 膚 のHAは生 体 HAの 約 50% を 占 め る が , 加 齢 に よ り 減 少 す る こ と が 知 ら れ て い る . 細 胞 の 増 殖 ・ 分 化 と い っ た 基 本 的 な 細 胞 機 能 に 関 与 す る HAの 減 少 は , 皮 膚 機 能 の 低 下 に 関 わ る と 考 え ら れ , HA代 謝 制 御 剤 は 抗 老 化 機 能 性 素 材 と し て 有 望 で あ る . 皮 膚HA の半減期は 0.5-1.5 日と短く, そのターンオーバーを担う HA 合成系は, 厳 密 に 制 御 さ れ て い る と 推 測 さ れ る. HA 合成酵素(HAS)には 3 種のアイソザイムが
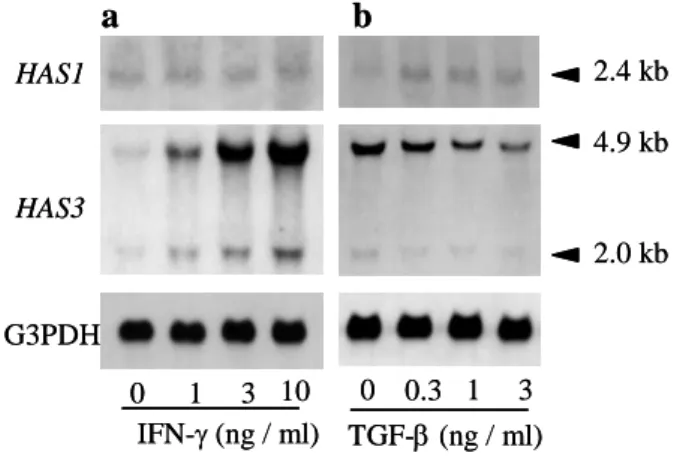
子 の 発 現 に つ い て 検 討 し た と こ ろ , HA 合 成 変 動 と 一 致 し た 発 現 挙 動 を 示 し た の は , 表 皮 細 胞 で ド ミ ナ ン ト に 発 現 す る HAS3 mRNA で あ っ た (Fig. 1). 表 皮 ヒ ア ル ロ ン 酸 合 成 を 促 進 さ せ る レ チ ノ イ ン 酸 の 刺 激 で も HAS3 遺 伝 子 の 顕 著 な 発 現 誘 導 が 観 察 さ れ た . こ れ ら の 結 果 か ら 表 皮 HA の 合 成 は , 3 種 の HAS 遺 伝 子 の う ち , 主 に HAS3 遺 伝 子 の 発 現 を 介 し て 制 御 さ れ る と 考 え ら れ た . ヒ ト 真 皮 線 維 芽 細 胞 に お い て HA 合 成 を 調 節 す る 合 成 酵 素 遺 伝 子 は , 主 に HAS2 で あ る こ と か ら , 表 皮 と 真 皮 で は HAS の 発 現 パ タ ー ン が 異 な る と 推 測 さ れ る . 表 皮 細 胞 の HA 合 成 を 抑 制 し た TGF-βは , 線 維 芽 細 胞 で は 主 に HAS2 mRNA 発 現 誘 導 を 介 し て HA 合 成 を 促 進 す る . TGF-βが, 表 皮 細 胞 の 増 殖 を 抑 制 し, 逆 に 線 維 芽 細 胞 の 増 殖 を 促 進 す る サ イ ト カ イ ン で あ る こ と を 考 え あ わ せ る と , TGF-β に よ る ΗΑ合 成 制 御 は , 細 胞 増 殖 時 に ス ペ ー ス を 確 保 す る と い う HA の 役 割 を 支 持 す る 応 答 で あ る と 考 え ら れ た . ヒ ト 表 皮 細 胞 に よ り 合 成 さ れ る HA分 子 量 サ イ ズ は , サ イ ト カ イ ン に よ る 刺 激 の 有 無 に 関 わ ら ず , 高 分 子 量 ( >1×106 Da) で あ っ た . し た が っ て , HAS3 に よ り 合 成 さ れ る HAの 分 子 量 サ イ ズ は , 高 分 子 で あ る と 考 え ら れ た . 本 研 究 に よ り, 表皮細胞における HA の合成制御には, 3 種の HAS 遺伝子のうち 主 に HAS3 遺 伝 子 の 発 現 が 重 要 な 役 割 を 担 っ て い る こ と が 明 ら か に な っ た . 表 皮 に お け る 速 い HA の タ ー ン オ ー バ ー を 勘 案 す る と , 合 成 機 構 だ け で な く 分 解 機 構 の 解 明 も 必 要 で あ る. 第 2 章 レ チ ノ イ ン 酸 レ セ プ タ ー を 介 し た HA 合 成 制 御 レ チ ノ イ ン 酸 (RA)は ビ タ ミ ン A の 活 性 本 体 で , 細 胞 増 殖 お よ び 分 化 の 制 御 に 重 要 な 役 割 を 演 ず る が , そ の 前 駆 体 はβ−カ ロ テ ン や β−ク リ プ ト キ サ ン チ ン に 代 表 さ れ る 食 餌 性 の プ ロ ビ タ ミ ン A カ ロ テ ノ イ ド で あ る . β-カロ テ ン が RA と 同 様 に , HAS3 mRNA の 発 現 を 誘 導 す る こ と, 加えてその HA 合成誘導が レチノイン酸レセ プ タ ー(RAR)拮抗薬である LE540 により阻害されたことにより, 表皮細胞における プ ロ ビ タ ミ ン A か ら RA に 至 る 代 謝 経 路 の 存 在 が 示 さ れ た . 一 方, 哺乳類ではレチ ノ イ ド に 代 謝 さ れ な い ル テ イ ン, ゼ ア キ サ ン チ ン お よ び ア ス タ キ サ ン チ ン な ど の ノ ン プ ロ ビ タ ミ ン A カロテノイドにも HAS3 mRNA 発現誘導活性およびその発現 0 1 3 10 IFN-γ (ng / ml) TGF-β (ng / ml) 0 0.3 1 3 HAS1 HAS3 G3PDH b a 2.4 kb 4.9 kb 2.0 kb
Fig. 1 The HAS3 transcript level of human keratinocytes was upregulated by IFN-γ but downregulated by TGF-β in a dose-dependent manner. Human keratinocytes were cultured in
various concentrations of IFN-γ (a) or TGF-β (b).
0 1 3 10 IFN-γ (ng / ml) TGF-β (ng / ml) 0 0.3 1 3 HAS1 HAS3 G3PDH b a 2.4 kb 4.9 kb 2.0 kb
Fig. 1 The HAS3 transcript level of human keratinocytes was upregulated by IFN-γ but downregulated by TGF-β in a dose-dependent manner. Human keratinocytes were cultured in
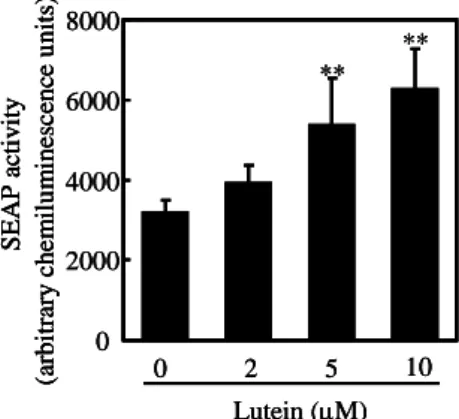
挙 動 と 一 致 す る HA 合 成 の 誘 導 が 観 察 さ れ た . HA 合成に及ぼすルテインの効果は β-カロテンと同様, LE540 で抑制された. ルテインによる刺激後に RARE 依存の転 写 活 性 が 増 大 し た こ と か ら (Fig. 2), ル テ イ ン の 代 謝 物 あ る い は ル テ イ ン 自 体 が RAR の リ ガ ン ド と し て 機 能 し て い る こ と が 示 さ れ た . さ ら に 表 皮 細 胞 で の HA 合 成 に 対 す る ル テ イ ン の 効 果 が , レ チ ナ ー ル 脱 水 素 酵 素 (RALDH)の 阻 害 剤 で あ る シ ト ラ ー ル に よ り 抑 制 さ れ た こ と か ら , ル テ イ ン は プ ロ ビ タ ミ ン A 同 様 , 表 皮 細 胞 内 で ア ル デ ヒ ド 化 合 物 に 変 換 さ れ , 続 い て 酸 化 反 応 を 受 け て RA 類 似 体 に 代 謝 さ れ る こ と で 生 理 活 性 を 発 揮 さ せ る こ と が 示 唆 さ れ た . 第 3 章 細 胞 内 HA 基 質 プ ー ル の 変 動 に よ る HA 合 成 制 御 機 構 表 皮 細 胞 の HA 合 成 を 高 め る 化 合 物 と し て , N-ア セ チ ル グ ル コ サ ミ ン (NAG)を 見 出 し た . 細 胞 内 に は 複 合 糖 質 の リ ソ ソ ー ム 分 解 物 と し て NAG が 存 在 す る . こ の NAG は , NAG キ ナ ー ゼ (NAGK)に よ り NAG6-リ ン 酸 に , 最 終 的 に は 糖 供 与 体 で あ る UDP-NAG に 変 換 さ れ る こ と で , 糖 タ ン パ ク や 糖 脂 質 の 糖 付 加 に 再 利 用 さ れ る . 表 皮 細 胞 で NAGK mRNA の 発 現 を 認 め , さ ら に NAG の 添 加 に よ り 細 胞 内 UDP-NAG 量 が 増 加 し た こ と か ら (Fig. 3), NAG の HA 産 生 促 進 作 用 は , 細 胞 内 基 質 の 増 大 に よ る も の で あ る こ と が 示 さ れ た . 加 え て , HA 生 合 成 過 程 に お い て 基 質 の 合 成 が 律 速 に な っ て い る と 考 え ら れ た . 表 皮 細 胞 に RA と NAG を 同 時 に 作 用 さ せ る こ と に よ り , 相 乗 的 な HA 合 成 促 進 効 果 を 認 め た こ と か ら , HA の も う 一 つ の 糖 供 与 体 で あ る UDP-GlcA の 細 胞 内 プ ー ル は , HA 合 成 の 律 速 に は な っ て い な い こ と が 示 唆 さ れ た . 次 に , NAG の 細 胞 内 取 り 込 み に つ い て も 検 討 し た . NAG の 取 り 込 み は グ ル コ ー ス ト ラ ン ス ポ ー タ ー (GLUT)を 介 す る も の で な く , 単 純 拡 散 に よ り 取 り 込 ま れ る と 予 想 し , 細 胞 内 で NAG へ と 変 換 可 能 な 親 油 性 の NAG グ リ コ シ ド 誘 導 体 を 合 成・評 価 し た . NAG グ リ コ シ ド 誘 導 体 は NAG に 比 し て 低 濃 度 域 で 作 用 を 示 し た . こ の 結 果 は , NAG が 単 純 拡 散 に よ り 細 胞 内 に 取 り 込 ま れ る こ と を 示 唆 し た . こ れ ら の 結 果 か ら , HA の 合 成 調 節 が 転 写 レ ベ ル だ け で な く , 細 胞 や 組 織 に お い 0 2000 4000 6000 8000 Lutein (µM) 0 2 5 10 ** ** S EA P a ctiv ity (a rb it ra ry ch e m il u m in es c en ce uni ts )
Fig. 2 Effect of lutein on RAR activation.
Significantly different from the control value; **P < 0.01 (William’s test). 0 2000 4000 6000 8000 Lutein (µM) 0 2 5 10 ** ** S EA P a ctiv ity (a rb it ra ry ch e m il u m in es c en ce uni ts ) 0 2000 4000 6000 8000 Lutein (µM) 0 2 5 10 ** ** S EA P a ctiv ity (a rb it ra ry ch e m il u m in es c en ce uni ts ) S EA P a ctiv ity (a rb it ra ry ch e m il u m in es c en ce uni ts )
Fig. 2 Effect of lutein on RAR activation.
Significantly different from the control value; **P < 0.01 (William’s test).
Fig. 3 Effect of NAG on UDP-NAG pools.
10 mM NAG UDP -N AG ( ng /µ g DN A) 0 50 100 150 + - - Keratinocytes Fibroblasts
Fig. 3 Effect of NAG on UDP-NAG pools.
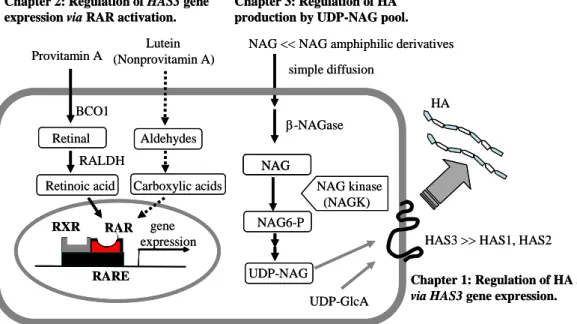
て は 細 胞 内 基 質 レ ベ ル に お い て も 制 御 さ れ る こ と が 示 唆 さ れ た . 以 上, Fig. 4 に示すように, 表皮 HA 代謝に関わる新たな知見について記載した. ま ず , 表 皮 の 転 写 レ ベ ル に お け る HA 合 成 の 制 御 は 主 に HAS3 mRNA が 担 う こ と を 見 出 し た . こ の こ と に よ り , 表 皮 HA 合 成 制 御 剤 開 発 に お い て , HAS3 mRNA 発 現 を 指 標 に し た 薬 剤 探 索 が 可 能 と な る こ と が 期 待 さ れ る . 次 に, RAR 活性化を機序とし, かつ安全性の高い HA 合成制御剤の候補として, プ ロ ビ タ ミ ン A お よ び シ ク ロ ヘ キ セ ン 環 と ポ リ エ ン 構 造 を 有 す る ノ ン プ ロ ビ タ ミ ン A カ ロ テ ノ イ ド を 見 出 し た . 最 後 に , HA 合 成 促 進 剤 と し て 細 胞 内 基 質 プ ー ル を 増 大 さ せ る NAG を 見 出 し , 転 写 レ ベ ル に よ る 制 御 だ け で は な く , 細 胞 内 基 質 プ ー ル へ 作 用 も HA 合 成 制 御 剤 開 発 の 重 要 な ス ト ラ テ ジ ー に な る こ と を 示 し た . 膜 透 過 を 考 慮 し た 両 親 媒 性 NAG 誘 導 体 の 開 発 に よ り , 効 率 的 に HA 合 成 を 亢 進 さ せ る こ と が 可 能 な こ と も 示 し た . 本 研 究 結 果 の 掲 載 1) J. Invest. Dermatol., 118, 43-48 (2002).
2) Biosci. Biotechnol. Biochem., 77, 1282-1286 (2013). 3) Skin Pharmacol. Physiol., 17, 77-83 (2004).
simple diffusion
NAG << NAG amphiphilic derivatives
β-NAGase NAG6-P UDP-NAG NAG kinase (NAGK) NAG UDP-GlcA
HAS3 >> HAS1, HAS2 HA RARE gene expression Provitamin A Retinal Retinoic acid Lutein (Nonprovitamin A) BCO1 RALDH RAR RXR Aldehydes Carboxylic acids
Fig. 4 Regulation of HA synthesis in keratinocyte. Chapter 2: Regulation of HAS3 gene
expression via RAR activation.
Chapter 3: Regulation of HA production by UDP-NAG pool.
Chapter 1: Regulation of HA synthesis
via HAS3 gene expression.
simple diffusion
NAG << NAG amphiphilic derivatives
β-NAGase NAG6-P UDP-NAG NAG kinase (NAGK) NAG NAG UDP-GlcA
HAS3 >> HAS1, HAS2 HA RARE gene expression Provitamin A Retinal Retinoic acid Lutein (Nonprovitamin A) BCO1 RALDH RAR RXR Aldehydes Carboxylic acids
Fig. 4 Regulation of HA synthesis in keratinocyte. Chapter 2: Regulation of HAS3 gene
expression via RAR activation.
Chapter 3: Regulation of HA production by UDP-NAG pool.
Chapter 1: Regulation of HA synthesis