Copyright 近畿作物・育種研究会(The Society of Crop Science and Breeding in Kinki, Japan) 21
Introduction
Soybeans with black seed coats have been widely utilized as food and as material for Oriental medicine because of the phenolic compounds present in their seed coat (Choung et al. 2001). Proanthocyanidins (PAs), the polymers of flavan-3-ols, are also a kind of phenolic compound, otherwise known as condensed tannins. Various colored soybean seed coats contain PA, and black seed coats have higher PA levels than other colored seed coats (Takahata et al. 2001). PAs have been shown to have some positive effects on human health, such as anti-inflammatory, anti-infectious, anti-carcinogenic and cardio-protective properties (Wang et al. 2011).
Previous reports have described the genetic diversity of PA in black soybean cultivars. Kim et al. (2006) showed a strong correlation between the seed coat color and the phenolic compounds. Slavine et al. (2009) compared the total phenolic content in 18 soybeans with different colored seed coats and concluded that the soybeans with black seed coats have a higher phenolic content than do other colored soybeans. Zhang et al. (2011) examined the phenolic content of 60 Chinese black soybean seed coats using HPLC and classified them into four groups using a hierarchical clustering analysis.
We found wide genetic variations in the PA content using 227 black soybean cultivars. A population containing 177 lines was developed from a cross between a high PA variety, ‘Peking’, and a low PA variety, ‘Tambaguro’. The objective of the present study was to identify the genetic regions associated with PA content in the seeds. ‘Peking’ is a landrace originating in China and has a small seed, whereas ‘Tambaguro’ originated in the
Kinki region and has a large seed. We also examined seed size and flowering time in order to investigate the relationships between these traits and PA content, and discussed the potential for breeding ‘Tambaguro’ that has a high PA content.
Materials and methods
Plant materials
A population was generated by crossing a high PA content variety (‘Peking’) with a low PA content variety (‘Tambaguro’). In total, 177 F3 and F4 segregation lines (hereafter called PTn lines) were used in this study. They were planted together with their parental varieties in the fields at the experimental farm at Kyoto University, Kyoto, Japan. This field experiment was conducted in 2009 (F3) and 2010 (F4). Each plot contained five plants. The flowering date (R1) and beginning pod date (R3) were observed for all PTn-lines in 2009. After harvesting, the seed size was also measured for all PTn-lines in 2009.
Measurement of proanthocyanidin content
Extraction of PA from the soybean seed coat was performed according to the method of Xu and Chang (2008). Briefly, 20 mg of seed coats, peeled from the seeds, were immersed in acetone/ water/acetic acid (70:29.5:0.5, v/v/v). The samples underwent extraction twice, first with 1 ml for 16 hours and then with 0.5 ml for 6 hours, at room temperature under dark conditions. The combined extract was stored at 4℃ in the dark until use. The extractions were conducted in triplicate for each individual line. An aliquot (100μl) of the extraction sample was added to 6 ml of 0.4% (w/v) vanillin in methanol and 3 ml of 4% (v/v) hydrochloric acid, and then allowed to stand for 15 min (Broadhurs and Jones, 1978). The absorbance was measured at 500 nm against methanol as a blank using a spectrophotometer
Identification of genomic regions involved in proanthocyanidin content of
black seed-coated soybean
Phommalath Siviengkhek, Masayoshi Teraishi, Takuji Tsukiyama and Yutaka Okumoto Graduate school of Agriculture, Kyoto University(Kitashirakawa, Sakyo, Kyoto 606 − 8502, Japan) Summary: Proanthocyanidin (PA), contained in the seed coat of black soybeans, has positive effects on human health. There were wide genetic variations in the PA content in the black soybean seed coat. In this study, chromosomal regions associated with the PA content were determined using the segregating population developed from a cross between a low PA variety, ‘Tambaguro’, and a high PA variety, ‘Peking’. The PA content was not related to flowering time or seed size. Single marker analysis with 150 polymorphic SSR (Single Sequence Repeat) markers revealed that Satt 592, located on Chromosome 10, was detected in both years of the experiment, while other markers were detected only in one year. These results indicate that a primary QTL (Quantitative Trait Locus) was located close to Satt 592 and that it was stable against environmental conditions.
Key words: Soybean, proanthocyanidin, QTL analysis
Acccepted : January 8, 2013
Corresponding author:Yutaka Okumoto([email protected])
J. Crop Res. 58:21 − 24(2013)
22
(BioSpec 1600, Shimadu, Japan). The amount of PA content was expressed as mg of (+)− catechin equivalents (mg CAE/g sample) using the (+)− catechin calibration curve. The linear range of the calibration curve was 10 μg/ml to 1000 μg/ml. All measurements were performed in triplicate.
Genotyping
Total genomic DNA was extracted from seed powder from each F2:3 and F3:4 line and parent using the method described by Kamiya and Kiguchi (2003). We estimated genotypes of 150 single sequence repeat (SSR) markers that produced clear bands. The PCR mixture (10 μl) contained 30 ng of genomic DNA, 5 pmol of primers, 2 nmol of dNTPs, 1 μl of 10x PCR buffer and 1 U of Taq polymerase. PCR conditions were as follows: initial denaturation at 95℃ for 5 min followed by 33 cycles of polymerization reaction, each consisting of a denaturation step at 92℃ for 1 min, an annealing step at 47℃ for 1 min, and an extension step at 68℃ for 1 min, with a final extension at 68℃ for 3 min. The PCR products were subjected to electrophoresis on a 12% polyacrylamide gel and stained with ethidium bromide, followed by scanning using an imaging system ChemiDocTM XRS+ (BioRad, Japan).
QTL analysis
QTL (Quantitative trait locus) analysis for PA content was conducted using single marker analysis that was performed using Windows QTL cartographer 2.5 (Wang et al. 2012).
Results and discussion
The PA contents of the 177 F3:4 (2009) and F4:5 (2010) lines analyzed in this study exhibited a wide distribution, from 80.0 to 306.8 μg CAE/mg with an average of 192.2 μg CAE/mg and from 84.4 to 323.1 μg CAE/mg, with an average of 180.8 μg CAE/mg, respectively (Fig.1). However, the PA content of
‘Tambaguro’ and ‘Peking’ in 2009 was 86.5 μg CAE/mg and 234.6 μg CAE/mg, respectively. The segregation pattern was transgressive, indicating that both parents possess independent genetic factors that increased the PA content.
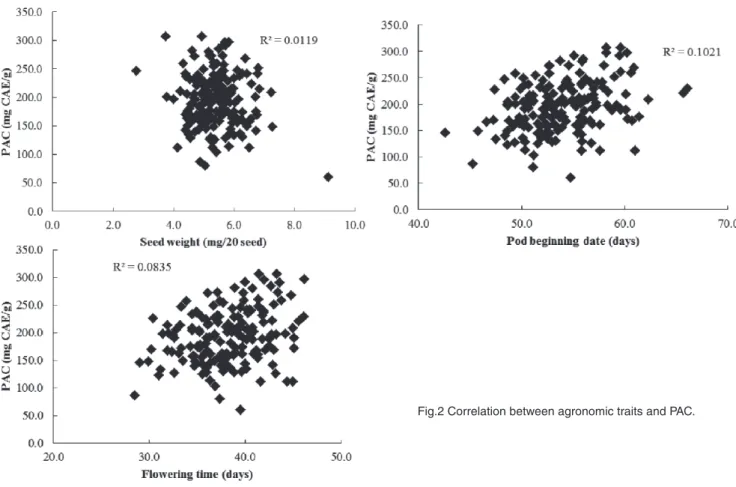
To evaluate the effects of seed size, flowering time and beginning pod time to PA content, the seed weight, flowering date and beginning pod date were measured and the correlation coefficients among them were calculated (Fig.2). There was no relationship between the PA content and the seed size, flowering time or beginning pod time. This result suggests that PA synthesis and accumulation is independent of the difference in seed size and plant life cycle. It also exhibits a potential for using backcrossing to breed high-functional ‘Tambaguro’ varieties that have a high PA content.
To detect chromosomal regions associated with the PA content, single marker analysis was performed using 150 SSR markers. Five and nine markers with significance at P<0.01 were detected in F3:4 and F4:5 populations, respectively (Table 1). Among them, Satt592, which is located on Chromosome 10 (formerly linkage group O), was detected in both populations. An allele derived from ‘Peking’ was desirable for increasing the PA content. This region contained a sole primary stable QTL, which is not effected by environment. The phenotypic variance explained by this QTL was 4.27% and 10.78% in F3:4 and F4:5 generation, respectively. These values were low regardless of the stable QTL. It might be because early generations were used for QTL analysis and the heterosis, derived from remaining heterozygous regions, obscured the QTL effect. Further investigation using fixed recombinant inbred lines carrying few heterozygous regions will be necessary to confirm the efficacy of this QTL. Other markers were detected only in one population, indicating that QTLs close to these markers were unstable and vulnerable.
J. Crop Res. 58(2013)
23 In conclusion, we found a stable QTL associated with the PA
content on chromosome 10. There was no observed relationship between the PA content and the seed size or flowering time. Therefore, there is the potential to use backcrossing to breed more bioactive ‘Tambaguro’ with a high PA content. Future research will involve developing residual heterozygous lines
(RHLs) (Yamanaka et al 2005) that carry a heterozygous segment around Satt592 on chromosome 10, and then proceeding to fine mapping.
Identification of genomic regions involved in proanthocyanidin content of black seed-coated soybean
Fig.2 Correlation between agronomic traits and PAC.
24
References
Broadhurst, R. B. and W. T. Jones (1978) Analysis of condensed tannins using acidified vanillin. J. Sci. Food Agric. 29 : 788− 794.
Choung, M., I. Baek, S. Kang, W. Han, D. Shin, H. Moon and K. Kang (2001) Isolation and determination of anthocyanins in seed coats of black soybean (Glycine max (L.) Merr.). J. Agric. Food Chem. 49 : 5848− 5851.
Kamiya, M. and T. Kiguchi (2003) Rapid DNA extraction method from soybean seeds. Breed. Sci. 53 : 277− 279.
Kim, J., W. Jung, S. Chun, C. Yu, K. Ma, J. Gwag and I. Chung (2006) A correlation between the level of phenolic compounds and the antioxidant capacity in cooked-with-rice and vegetable soybean (Glycine max L.) varieties. Euro. Food Res. Tech. 224 : 259− 270.
Slavin, M., W. Kenworthy and L. Yu (2009) Antioxidant properties, phytochemical composition, and antiproliferative activity of Maryland-grown soybeans with colored seed coats. J. Agric. Food Chem. 57 : 11174− 11185.
Takahata, Y., M. Ohnishi-Kameyama,S.Furuta, M. Takahashi and I. Suda (2001) Highly polymerized procyanidins in brown
soybean seed coat with a high radical-scavenging activity. J. Agric. Food Chem. 49 : 5843− 5847.
Wang, S., C. J. Basten and Z. B. Zeng (2012) Windows QTL Cartographer 2.5. Department of Statistics, North Carolina State University, Raleigh, NC. (http://statgen.ncsu.edu/qtlcart/ WQTLCart.htm)
Wang, Y., S. Chung, W. O. Song and O. K. Chun (2011) Estimation of daily proanthocyanidin intake and major food sources in the U.S. diet. J. Nutrition 141 : 447− 452.
Xu, B. and S. K. Chang (2008) Antioxidant capacity of seed coat, dehulled bean, and whole black soybeans in relation to their distributions of total phenolics, phenolic acids, anthocyanins, and isoflavones. J. Agric. Food Chem. 56 : 8365− 8373. Yamanaka, N., S. Watanabe, K. Toda, M. Hayashi, H. Fuchigami,
R. Takahashi and K. Harada (2005) Fine mapping of the FT1 locus for soybean flowering time using a residual heterozygous line derived from a recombinant inbred line. Theor. Appl. Genet. 110 : 634− 639.
Zhang, R. F., F. X. Zhang, M. W. Zhang, Z. C. Wei, C. Y. Yang, Y. Zhang, X. J. Tang, Y. Y. Deng and J. W. Chi (2011) Phenolic composition and antioxidant activity in seed coats of 60 Chinese black soybean (Glycine max L. Merr.) varieties. J. Agric. Food Chem. 59 : 5935− 5944.
J. Crop Res. 58(2013)
黒ダイズのプロアントシアニジン含量に関与するゲノム領域の同定
Phommalath Siviengkhek・寺石政義・築山拓司・奥本 裕
京都大学大学院農学研究科(〒 606 − 8502 京都市左京区北白川追分町) 要旨:プロアントシアニジンは、種皮色が黒のダイズの種皮に多く含まれており、様々な生体調節機能をもつとされて いる。黒ダイズ種皮に含まれるプロアントシアニジン含量には大きな遺伝的変異がある。今回の研究では、低プロアン トシアニジン品種である「丹波黒」と高プロアントシアニジン品種である「ペキン」の交雑後代を用いてプロアントシ アニジン含量に関連する染色体領域を同定した。プロアントシアニジン含量は開花日や種子重と相関がなかった。 150個の SSR マーカーを使った single marker analysis では染色体10に座乗する Satt592 のみが二年とも検出され、 他のマーカーは一年のみしか検出されなかった。主たる QTL は Satt592 の近傍に座乗しており、栽培条件などに対し安 定的であることが示唆された。キーワード:ダイズ・プロアントシアニジン・QTL 解析
作物研究 58:21 − 24(2013) 連絡責任者:奥本 裕([email protected])