タバコ(Nicotiana tabacum L.)における翻訳開始
因子遺伝子を利用したウイルス病抵抗性育種に関す
る研究
著者
宇田川 久史
学位授与機関
Tohoku University
学位授与番号
11301甲第19318号
URL
http://hdl.handle.net/10097/00127863
博士論文要約
タバコ(
Nicotiana tabacum L.)における翻訳開始因子遺伝子を
利用したウイルス病抵抗性育種に関する研究
東北大学大学院農学研究科
応用生命科学専攻
宇田川 久史
指導教員
北柴 大泰 教授
第1 章 序論
タバコ(Nicotiana tabacum L.)は世界 120 か国以上で栽培されている工芸作物であ り(FAOSTAT; http://www.fao.org/faostat)、栽培地域ごとに様々な品種が開発されてい る。他作物同様、タバコにおいても、病害抵抗性は収量・品質と並ぶ重要な育種目標
である。タバコの重要病害の一つであるtobacco bushy top disease(TBTD)は、感染個
体に生育遅延やmottling、わき芽の異常伸長を引き起こすタバコのウイルス病である。
本病の原因ウイルスとして、Tobacco bushy top virus(TBTV; Umbravirus 属)および
Ethiopian tobacco bushy top virus(ETBTV; Umbravirus 属)が知られている。これまで、 N. tabacum における本病に対する抵抗性遺伝資源は見つかっていない。また、別の重
要病害に、Potato virus Y(PVY; Potyvirus 属)を原因ウイルスとする黄斑えそ病があ
る。PVY に感染したタバコは、葉脈えそやモザイク症状を示す。タバコ系統‘Virgin A Mutant’(VAM)に由来する劣性対立遺伝子座 va がタバコの PVY 抵抗性育種に広く利
用されているが、va 導入品種においても本病に罹病することがあり、さらに強く、安
定した抵抗性が求められている。
近年、宿主の翻訳開始因子(eukaryotic translation initiation factor; eIF)の欠損による ウイルス抵抗性獲得の事例が多く報告されている(Bastet et al. 2017)。しかしながら、 TBTD の原因ウイルスと eIF 遺伝子ファミリーとの関係や、それらの改変による抵抗 性系統作出の可能性は不明である。PVY については、タバコ Va 座に eIF 遺伝子ファミ
リーの一つeIF4E 群の遺伝子が座乗しており、va 座による PVY 抵抗性はこの遺伝子
の欠失に起因することが明らかになっているが(Noguchi et al. 1999; Julio et al. 2015)、 PVY 抵抗性の不安定性の原因解明には至っていない。
本研究では、eIF4E 群に注目した分子遺伝学的な研究により、これらの遺伝子と
TBTD および黄斑えそ病に対する抵抗性の関係を明らかにすることを目的とした。
第2 章 Tobacco bushy top disease(TBTD)抵抗性に関与する eIF4E 遺伝子の探索と
同定 タバコ主要産国の一つであるアフリカのマラウイ国でTBTD が発生し、大きな問題 となっている。そこで本章では、マラウイ国で見出されたTBTD の原因ウイルスを同 定し、eIF 遺伝子ファミリーと TBTD 抵抗性との関係を調査した。 第1 節 マラウイにおける TBTD の原因ウイルスの同定とゲノム配列決定 TBTV および ETBTV の塩基配列を基に設計したプライマー(Fig. 1)を用いてマラ ウイ由来のTBTD 罹病葉の RT-PCR を行い、増幅産物(‘Seq-A’とする。Fig. 1)の塩基 配列を決定した。プライマー領域を除く1,860 nt の BLAST 検索の結果、Seq-A は ETBTV と最大の相同性(87.0%)を示した。この ETBTV 様ウイルスの全ゲノム塩基 配列を取得するためRNA-seq 法により TBTD 罹病葉由来の塩基配列 3.8Gb を解読し た。タバコゲノム塩基配列へのマッピングによりタバコ由来の配列を取り除き、マッ ピングされなかったリードのアセンブルにより、24 のコンティグ(contig)を得た。
これらの中に、ETBTV の全塩基配列と 87.7%の相同性を示す 4,186 nt の contig が存在
した。この配列からは4 つの主要な ORF(Fig. 2)が予測され、その位置や推測される
タンパク質の機能はいずれも他のUmbravirus 属のウイルスと共通していた。RNA 抽
出に用いた罹病葉を接種源にしてウイルスを継代した後代タバコにおいても同様の病
徴と同一の塩基配列が得られた。以上から、本塩基配列がTBTD の原因ウイルス
TBTV-MW(tobacco bushy top virus Malawi)のゲノム塩基配列であるとした。分子系統 解析の結果、TBTV-MW は他の Umbravirus 属のウイルスの中で ETBTV と最も近縁で あった(Fig. 3)。 第2 節 eIF4E の発現抑制および欠損による TBTV-MW がもたらす TBTD に対する 抵抗性の調査 1) RNAi 発現抑制による抵抗性の調査 タバコはS ゲノムを持つ N. sylvestris と T ゲノムを持つ N. tomentosiformis に由来す る異質倍数体種であり、eIF4E 群については、eIF4E1、eIF4E2、eIF(iso)4E が S ゲノム
型(以降S 型)と T ゲノム型(T 型)でそれぞれ存在する(Sierro et al. 2014)。eIF4E1、
eIF4E2、eIF(iso)4E のそれぞれについて、S 型と T 型を同時に発現抑制した‘Petit
Havana SR1’(以降‘SR1’)由来 RNAi 発現抑制系統(T2)を供試し、TBTV-MW の感受
性に関与するeIF4E を探索した。各系統における RT-qPCR の結果(Fig. 4)、RNAi カ
セットを有する系統では、対象とするS 型、T 型遺伝子の発現量が顕著に低下してい
た。続いて、TBTV-MW を汁液接種し、病徴を観察した(Fig. 5)。eIF4E1-RNAi 系統 では、RNAi カセットを有する(+)、有さない(-)系統間で発病率に差は見られな かった。eIF4E2-RNAi 系統では、RNAi(+)の発病率は RNAi(-)に比べて低い傾
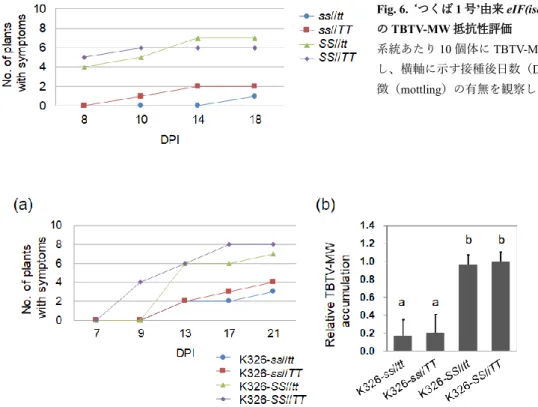
向にあったが、系統間でバラつきが見られた。これらに対して、eIF(iso)4E-RNAi (+)系統では、接種後18 日目の病徴観察においても、供試した 3 系統いずれも発病 率は10%以下であり、RNAi(-)系統および‘SR1’と比較して大きく低下していた。 以上から、eIF(iso)4E 遺伝子が TBTV-MW の感受性に大きく関与することが明らかに なった。 2)突然変異体による抵抗性の調査 eIF(iso)4E 遺伝子と TBTV-MW の感受性との関係について、変異体を用いてさらに 解析した。eIF(iso)4E-S あるいは eIF(iso)4E-T の第一エキソンにナンセンス突然変異が 生じた‘つくば 1 号’由来変異体をそれぞれ選抜し、両変異体の交配と自殖により二重 変異体を作出した。以降、eIF(iso)4E-S と eIF(iso)4E-T の変異型対立遺伝子をそれぞれ ‘s’および‘t’、野生型を‘S’および‘T’と表記する。TBTV-MW を接種した結果(Fig. 6)、 発病個体数は遺伝子型ss//tt を持つ系統と ss//TT 系統で少なく、接種 18 日経過後も 10 個体中2個体以下であった。一方、SS//tt 系統の発病個体数は対照と同程度で、接種後 18 日目には 7 個体が発病した。遺伝的背景の大部分をタバコ品種‘K326’に置換した変 異体系統(‘K326-ss//tt’のように表記)を用いた接種試験においても、K326-ss//tt 系統
とK326-ss//TT 系統で発病個体が少なく、K326-SS//tt 系統および K326-SS//TT 系統では 多くの個体で発病が見られた(Fig. 7a)。RT-qPCR による TBTV-MW 量の半定量的分析 の結果、K326-ss//tt 系統および K326-ss//TT 系統では K326-SS//tt 系統および対照に比べ てTBTV-MW 量が有意に減少していた(Fig. 7b)。自然環境下において、Umbravirus 属 ウイルスはアブラムシを介して感染する。そこで虫媒接種試験を実施した結果、K326-ss//tt 系統および K326-ss//TT 系統の発病率は、接種 25 日後にそれぞれ 0%、26.7%とな り、K326-SS//TT 系統の 73%に比べて顕著に低い発病率を示した(Fig 8a)。接種 25 日 後におけるK326-ss//tt 系統および K326-ss//TT 系統の TBTV-MW 量は K326-SS//TT 系統 と比べて低い値を示した(Fig. 8b)。 本研究から、TBTV-MW は eIF(iso)4E-S の関与する翻訳機構を用いてタバコへの感染 を進行させることが示唆された。さらに、TBTV-MW への抵抗性は eIF(iso)4E-S および eIF(iso)4E-T の変異を組み合わせることで、強くかつ安定することが示された。 第3 章 eIF4E1-S および eIF(iso)4E-T の欠損による黄斑えそ病に対する抵抗性の調査 va 対立遺伝子による PVY 抵抗性の不安定性には、抵抗性を打破して出現する変異
型のPVY(Resistance-Breaking PVY; RB-PVY)が関係している可能性が考えられてい
る(Masuta et al. 1999)。また、RB-PVY は自然条件下においても存在するが(Lacroix et al. 2010; Janzac et al. 2014)、RB-PVY 抵抗性品種はこれまで開発されていない。黄斑
えそ病に対する抵抗性育種の進展には、va 座存在下における PVY 抵抗性の不安定性
の理解と解決、そしてRB-PVY に対する抵抗性品種の開発が必要である。
第一節 va 座存在下における PVY 抵抗性の不安定性の調査
PVY がタバコへ感染する際、PVY の持つ viral genome-associated protein (VPg)とタバ
コのeIF4E1-S が相互作用する。一方、RB-PVY は eIF(iso)4E-T と相互作用しており、
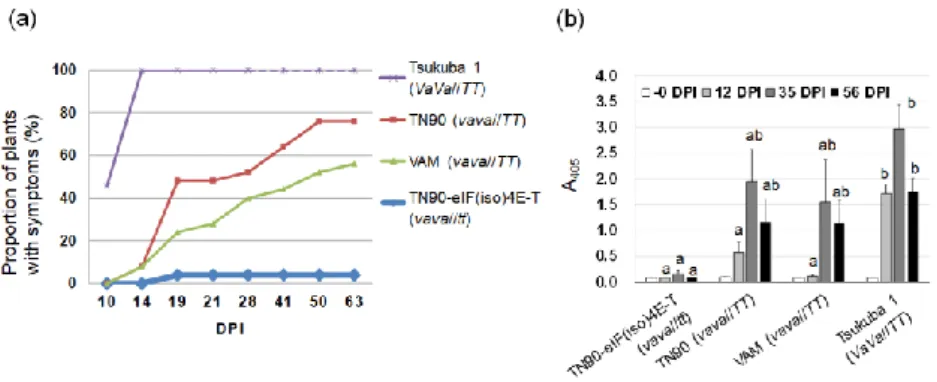
eIF(iso)4E-T の変異によって RB-PVY の感受性が低下する(Takakura et al. 2018)。そこ で、va 座存在下における変異型 eIF(iso)4E-T の PVY 抵抗性への関与について調査し た。 材料には、Va 座に欠失をもつ‘TN90’(vava//TT)、第二章と同様の eIF(iso)4E-T ナン センス突然変異(遺伝子座を‘t’と表記)を‘TN90’に導入した TN90-eIF(iso)4E-T 突然変 異体(vava//tt)、‘つくば 1 号’(VaVa//TT)を用いた。また、va に加えて、未同定の因子 により‘TN90’より強い PVY 抵抗性を示すとされる‘VAM’(vava//TT)を加えた。PVY 接種試験の結果(Fig. 9a)、接種後 14 日目における発病率が‘つくば 1 号’では 100%とな ったのに対して、va を持つ他の 3 系統では 0〜10%と低かった。しかしながら、 ‘TN90’および‘VAM’ではその後徐々に発病率が増加し、接種後 63 日目にはそれぞれ 76%、56%に達した。一方、TN90-eIF(iso)4E-T 変異体では 63 日目においても発病率は わずか4%であった。ELISA によるウイルス量測定においても(Fig. 9b)、‘つくば 1 号’で最もウイルス量が多く、次いで‘TN90’、‘VAM’であった。TN90-eIF(iso)4E-T 突然 変異体のウイルス量は顕著に低く推移した。
PVY 接種源および‘つくば 1 号’と‘TN90’の罹病葉中に含まれる PVY の VPg につい て、部分塩基配列を決定した。推定アミノ酸配列を比較した結果(Fig. 10)、接種源お よび‘つくば 1 号’のアミノ酸配列は同一であった。一方‘TN90’では、供試した 4 個体
全てでRB-PVY への変化を示唆するアミノ酸置換(K105E が 3 個体、V108I が 1 個
体)が生じていた(Masuta et al. 1999; Janzac et al. 2014)。
以上の結果から、va 座を持つ品種における PVY 抵抗性の不安定性は、RB-PVY と同
じVPg を持つ変異型 PVY の出現が原因であり、va 座と変異型 eIF(iso)4E-T の両方を
持つことで、突然変異型のPVY の増殖が抑えられると考えられた。
第2 節 eIF(iso)4E-T の欠損を利用した RB-PVY に対する抵抗性の調査
変異型eIF(iso)4E-T による RB-PVY 抵抗性が報告されているが(Takakura et al. 2018)、その抵抗性の強度は詳細に調査されていない。また、RB-PVY は eIF4E1-S と も相互作用していることから(Takakura et al. 2018)、vava//tt 系統と VaVa//tt 系統では抵
抗性の強度に違いが出る可能性が考えられる。そこで、両系統におけるRB-PVY 抵抗
性の強度を調査した。
‘つくば 1 号’、‘TN90’、eIF(iso)4E-T 突然変異体(VaVa//tt)および TN90-eIF(iso)4E-T 突
然変異体を用いてRB-PVY 抵抗性を評価した(Fig. 11a)。接種後 10 日目において、
‘つくば 1 号’および‘TN90’では全個体で発病が見られた一方、eIF(iso)4E-T 突然変異体 では全く発病が観察されなかった。しかしながら、接種後19 日目には eIF(iso)4E-T 変 異体でも発病個体が増加したことから、その抵抗性は完全でないことが明らかになっ た。これらに対して、TN90-eIF(iso)4E-T 変異体では接種後 28 日目においても全く発病 が見られなかった。接種後17 日目における各系統の RB-PVY 量を ELISA で測定した 結果(Fig. 11b)、‘つくば 1 号’および‘TN90’でウイルス量が多く、次いで eIF(iso)4E-T 変異体であった。TN90-eIF(iso)4E-T 変異体ではウイルス量は顕著に低い値となった。
以上から、変異型eIF(iso)4E-T のみでは RB-PVY 抵抗性は安定しないが、va 座と組み
合わせることで強く、安定した抵抗性となることが示された。 第4 章 総合考察 本研究では、TBTD および黄斑えそ病に対する病害抵抗性育種の進展を目指し、 eIF4E 群遺伝子とこれらウイルス病に対する抵抗性の関係を調査した。マラウイ由来 のウイルスTBTV-MW のゲノム解読の結果、TBTV-MW は Umbravirus 属の ETBTV と 近縁であることが明らかとなった。TBTV-MW に対する感受性は eIF(iso)4E-S のナンセ ンス変異によって低下すること、eIF(iso)4E-S と eIF(iso)4E-T の二重変異体は eIF(iso)4E-S 変異体よりも強い抵抗性となることが示された。これまで TBTD に抵抗性 を示すタバコ遺伝資源は知られておらず、本研究によってTBTD 抵抗性育種が実現す る可能性が高まった。また、これまでUmbravirus 属ウイルスに対する抵抗性に eIF(iso)4E が関与するという報告はない。本研究の知見は他の Umbravirus 属ウイルス に対する抵抗性研究に有効な情報となると考えられた。
一方、黄斑えそ病に対しては、va と変異型 eIF(iso)4E-T の組合せにより PVY 抵抗性 が向上した。va による抵抗性の不安定性は、抵抗性を打破する突然変異 PVY の出現 が原因であり、変異型eIF(iso)4E-T がその増殖を抑制することで抵抗性が安定化する と考えられた。vava//tt 遺伝子型は RB-PVY に対しても安定した抵抗性を示したことか ら、vava//tt 遺伝子型の利用は黄斑えそ病に対する抵抗性が安定した品種の開発に貢献 することが期待される。 本研究において作出した抵抗性系統や利用した遺伝子は、対象としたウイルス病抵 抗性の育種に有用と考えられる。各栽培地域における病害発生状況を把握し、必要な eIF4E 群遺伝子の変異を組み合わせて育種を進めることで、より安定的なタバコ栽培 が可能となる。
Fig. 1. TBTD 罹病葉からのウイルス断片の増幅
TBTV と ETBTV の配列を基に設計した縮重プライマー(図中赤矢印)を用いて TBTD 罹病葉から RT-PCR を行い、増
幅断片の塩基配列を決定した。その後、特異的プライマー(図中黄矢印)で塩基配列を決定した。図中RdRp は RNA
dependent RNA polymerase のコーディング領域を示す。
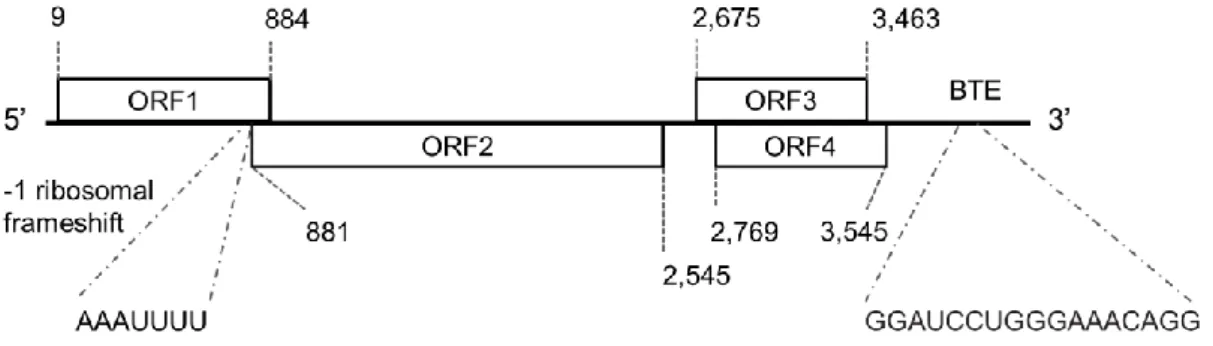
Fig. 2. 推定された原因ウイルスのゲノム構造
水平方向の横線が原因ウイルスのRNA ゲノムの全長(4,186 nt)を示す。ORF1 と ORF2 は-1 ribosomal frameshift によ
り一つの融合タンパク質(RdRp)を、ORF3 と ORF4 はそれぞれ cell to cell movement protein と long-distance movement protein をコードすると推測された。3′ UTR には cap-independent translation elements(3′-CITEs)の一つであり、TBTV で 確認されているBarley yellow dwarf virus (BYDV)-like translation elements(BTE)の保存配列(17 nt)が存在した。
Fig. 3. TBTV-MW および他の Umbravirus 属のウイルスにおける分子系統解析
TBTV-MW、Ethiopian tobacco bushy top virus (ETBTV)、Groundnut rosette virus (GRV)、 Tobacco bushy top virus (TBTV)、
Opium poppy mosaic virus (OPMV)、Pea enation mosaic virus 2 (PEMV-2)、Ixeridium yellow mottle virus 2 (IxYMV-2)、Carrot mottle mimic virus (CMoMV)、Carrot mottle virus (CMoV)、Sclerotinia sclerotiorum umbra-like virus 1 (SsULV1)のゲノム配列
を用いて分子系統樹を作成した。括弧内は各ウイルスのGenBank accession number を示す。各枝の分岐点における数字
Fig. 4. RNAi 系統におけるターゲット eIF4E の発現量 各遺伝子の相対的な転写量をRT-qPCR で測定した。‘SR1’の平均値を 1 としたときの各系統(T2世代)の平均値と標準 誤差を示す(n≧4)。系統番号(#)は各 T2系統が由来するT0個体の番号を示す。ターゲット遺伝子が同じで、かつ (+)系統と(-)系統で系統番号が同じ場合は、共通のT0個体に由来するT2系統である。eIF4E2-RNAi において、 #2 は(-)系統のみ、#7 は(+)系統のみを供試した。RNAi カセット中に含まれる NPT II 遺伝子の PCR を事前に行 い、カセットを有する(+)および有さない(-)ことを確認した上で測定を行った。 Fig. 5. 汁液接種による RNAi 系統の TBTV-MW 抵抗性評価
接種後10 日および 18 日(Days Post-Inoculation; DPI)における病徴(mottling)の有無を観察した。系統ごとに、全接
種個体数(図中括弧)に対する発病個体数を比率で示した。系統番号(#)は各T2系統が由来するT0個体の番号を示 す。ターゲット遺伝子が同じで、かつ(+)系統と(-)系統で系統番号が同じ場合は、共通のT0個体に由来するT2 系統である。eIF4E2-RNAi において、#2 は(-)系統のみ、#7 は(+)系統のみを供試した。 0 20 40 60 80 100 10 DPI 18 DPI P ro p o rti o n o f p la n ts w ith s y m p to m s ( % ) eIF4E1-RNAi lines - #3 (5) #4 (5) #5 (5) #3 (5) #4 (5) #5 (4) eIF4E2-RNAi lines #1 (5) #2 (5) #8 (5) #1 (5) #7 (5) #8 (5) eIF(iso)4E-RNAi lines #1 (9) #7 (10) #15 (9) #1 (10) #7 (10) #15 (10) SR1 (10) - + - + RNAi cassette +
Fig. 6. ‘つくば 1 号’由来 eIF(iso)4E 突然変異体 のTBTV-MW 抵抗性評価 系統あたり10 個体に TBTV-MW を汁液接種 し、横軸に示す接種後日数(DPI)における病 徴(mottling)の有無を観察した。 Fig. 7. ‘K326’の遺伝的背景を持つ eIF(iso)4E 突然変異体の TBTV-MW 抵抗性評価 (a)系統あたり 10 個体に TBTV-MW を汁液接種し、横軸に示す接種後日数(DPI)における病徴(mottling)の有無を 観察した。(b)抵抗性評価試験とは別に接種を行った系統を用いて 13 DPI における TBTV-MW 量を RT-qPCR で測定し た。K326-SS//TT 系統の平均値を 1 として、各系統の平均値と標準誤差を示した(n≧4)。異なるアルファベットのつい た系統間では統計的有意差が認められた(Tukey’s test, P < 0.05)。 Fig. 8. 虫媒接種試験における eIF(iso)4E 変異体の抵抗性評価 (a)アブラムシを介して K326-ss//TT 系統、K326-ss//tt 系統、K326-SS//TT 系統に TBTV-MW を感染させ、横軸に示す 接種後日数(DPI)における病徴(mottling)の有無を観察した(n=15)。(b)25 日目の観察後、TBTV-MW 量を RT-qPCR で測定した。K326-SS//TT 系統の平均値を 1 として、各系統の平均値と標準誤差を示した(n=15)。異なるアルフ ァベットのついた系統間では統計的有意差が認められた(Tukey’s test, P < 0.05) 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 R e la ti v e T BT V -MW a c c u m u la ti o n N o . of p la n ts w it h s y m p to m s a b ab (b) (a) 0 3 6 9 12 15 14 19 25 DPI K326-sstt K326-ssTT K326-SSTT K326-ss//tt K326-ss//TT K326-SS//TT
Fig. 9. 汁液接種による PVY の抵抗性評価
(a)系統あたり 25 個体(‘つくば 1 号’は 24 個体)に PVY を汁液接種し、横軸に示す接種後日数(DPI)における病徴 (葉脈えそ)の有無を観察した。接種個体数における発病個体数の割合を示した。‘つくば 1 号’では発病個体が 100%
に達したことから28 DPI で観察を終了した。そのため 28 DPI 以降は点線で示した。(b)接種試験に用いた各系統の個
体からランダムに5 個体ずつ選び、接種直前(-0 DPI)および 12 DPI、35 DPI、56 DPI において、PVY 量を ELISA に
より測定した。各系統の平均値と標準誤差を示した。各DPI における系統間で Tukey’s test(P < 0.05)を行い、統計的
有意差が認められた系統間は異なるアルファベットで示した。 Fig. 10. 接種源および接種個体における PVY の推定アミノ酸配列 接種源およびPVY 接種後 35 日目に罹病していた‘つくば 1 号’と‘TN90’各 4 個体において、罹病葉中に含まれる PVY のVPg について塩基配列の一部(VPg の全 188 残基のうち 93 から 120 残基に相当する領域)を解読した。接種源の推 定アミノ酸配列をリファレンスとして、各サンプルにおけるVPg の推定アミノ酸配列を比較した。リファレンスと一 致しているアミノ酸残基は‘-’で表した。 Fig. 11. 汁液接種による RB-PVY の抵抗性評価
(a)系統あたり 10 個体(‘TN90’のみ 15 個体)に RB-PVY を接種し、横軸に示す接種後日数(DPI)における病徴(葉脈 えそ)の有無を観察した。接種個体数における発病個体数の割合を示した。(b)接種試験に用いた各系統の個体からラ
ンダムに3 個体ずつ選び、RB-PVY の汁液接種後 17 日目において、RB-PVY 量を ELISA により測定した。各系統の平
均値と標準誤差を示した。Tukey’s test(P < 0.05)を行い、統計的有意差が認められた系統間は異なるアルファベット で示した。
引用文献
Bastet, A., C. Robaglia and J.L. Gallois (2017) eIF4E resistance: natural variation should guide gene editing. Trends Plant Sci. 22: 411–419.
Janzac, B., M. Tribodet, C. Lacroix, B. Moury, J.L. Verrier and E. Jacquot (2014) Evolutionary pathways to break down the resistance of allelic versions of the PVY resistance gene va. Plant Dis. 98:1521–1529.
Julio, E., J. Cotucheau, C. Decorps, R. Volpatti, C. Sentenac, T. Candresse and D.F. de Borne (2015) A eukaryotic translation initiation factor 4E (eIF4E) is responsible for the “va” tobacco recessive resistance to potyviruses. Plant Mol. Biol. Rep. 33: 609–623.
Lacroix, C., L. Glais, C. Kerlan, J.L. Verrier and E. Jacquot (2010) Biological characterization of French Potato virus Y (PVY) isolates collected from PVY-susceptible or -resistant tobacco plants possessing the recessive resistance gene va. Plant Pathol. 59: 1133–1143.
Masuta, C., M. Nishimura, H. Morishita and T. Hataya (1999) A single amino acid change in viral genome-associated protein of potato virus y correlates with resistance breaking in ‘virgin a mutant’ tobacco. Phytopathology 89: 118–123.
Noguchi, S., T. Tajima, Y. Yamamoto, T. Ohno and T. Kubo (1999) Deletion of a large genomic segment in tobacco varieties that are resistant to Potato virus Y (PVY). Mol. Gen. Genet. 262: 822– 829.
Sierro, N., J.N. Battey, S. Ouadi, N. Bakaher, L. Bovet, A. Willig, S. Goepfert, M.C. Peitsch and N.V. Ivanov (2014) The tobacco genome sequence and its comparison with those of tomato and potato. Nat. Commun. 8: 3833.
Takakura, Y., H. Udagawa, A. Shinjo and K. Koga (2018) Mutation of a Nicotiana tabacum L. eukaryotic translation-initiation factor gene reduces susceptibility to a resistance-breaking strain of Potato virus Y. Mol. Plant Pathol. 19: 2124–2133.