粘膜関連リンパ組織リンパ腫転座 1
(Malt1) のプロテアーゼ活性による炎症反
応の制御とその機序に関する研究
2020 年 3 月
中村 嘉希
岡山大学大学院
環境生命科学研究科
略語
AHA, Autoimmune hemolytic anemia; BAC, bacteria artificial chromosome; BMDC, bone marrow-derived dendritic cell; BCR, B-cell receptor; CARD, caspase-associated recruitment domain; CD, cluster of differentiation; cDNA, complementary DNA; cIAP2, cellular inhibitor of apoptosis protein 2; DNA, deoxyribonucleic acid; DTT, dithiothreitol;
EAE, experimental autoimmune encephalomyelitis; EDTA, ethylenediaminetetraacetic acid; ELISA, Enzyme-linked immunosorbent assay; ES cell, embryonic stem cell; FBS, fetal bovine serum; FITC, fluorescein isothiocyanate; FcγR, Fcγ receptor; FcR, Fc receptor; GM-CSF, Granulocyte macrophage colony-stimulating factor; HRP, horseradish peroxidase; IACUC, Institutional Animal Care and Use Committee; IBD, inflammatory bowel disease; IC, immune complex; IκBα, inhibitor of κB-α; IFN-γ, interferon-γ; Ig, Immunoglobulin; IL, interleukin; ITP, immune thrombocytopenic purpura; mAb, monoclonal antibody; MALT, mucosa-associated lymphoid tissue; Malt1, MALT lymphoma-translocation protein 1; MLN, mesenteric lymph node; NF-κB, nuclear factor- κB; PCR, polymerase chain reaction; PD, protease-deficient; PE, phycoerythrin; PKC, protein kinase C; PMA, phorbol 12-myristate 13-acetate; P/I, PMA/Ionomycin; qPCR, quantitative polymerase chain reaction; qRT-PCR, quantitative-reverse transcription- polymerase chain reaction; Treg cell, regulatory T cell; RNA, ribonucleic acid; SDS- PAGE, sodium dodecyl sulfate-poly-acrylamide gel electrophoresis; SEM, standard error of the mean; SLE, systemic lupus erythematosus; SPF, specific pathogen-free; Syk, spleen tyrosine kinase; TCR, T-cell receptor; TGF-β, transforming growth factor-β; Th cell, helper T cell; Th1/17 cell, IL-17 and IFN-γ double-producing cell, TNF-α, tumor necrosis factor-α; TRAF6, tumor necrosis factor receptor-associated factor 6; WT, wild- type.
1
2
目次
略語 ... 1
第1章 序論 ... 4
第2章 Malt1プロテアーゼ活性の不活化によるTh17 とTh1/17細胞の制御と大腸 炎の抑制効果 ... 11
第1節 緒言 ... 11
第2節 材料および方法 ... 12
第1項 Malt1 PDマウスの作出 ... 12
第2項 動物 ... 14
第3項 T細胞の刺激とウエスタンブロッティング ... 15
第4項 In vitroサイトカイン産生試験 ... 16
第5項 抗CD3抗体誘発in vivoサイトカイン産生試験 ... 17
第6項 Naïve T細胞移入マウス大腸炎モデル ... 17
第7項 リアルタイムqRT-PCR ... 19
第8項 腸間膜リンパ節のCD4+ T細胞における細胞内サイトカイン染色... 20
第9項 統計解析 ... 21
第3節 結果 ... 21
第1項 Malt1 PDマウスにおけるプロテアーゼ活性の不活化検討 ... 21
第2項 Malt1 PDマウス由来T細胞のIL-2産生能 ... 24
第 3 項 Malt1 PDマウスにおける抗CD3 抗体刺激による in vivoサイトカイン産 生試験 ... 24
第4項 Naïve T細胞移入マウス大腸炎モデルの病態におけるMalt1プロテアー ゼ不活化の影響 ... 27 第 5 項 大腸炎モデルの腸間膜リンパ節におけるTh17細胞と Th1/17細胞の比
3
較 ... 28
第6項 大腸炎モデルにおける大腸のサイトカイン発現量の比較 ... 29
第4節 考察 ... 30
第 3 章 Fcγ 受容体シグナルを介した炎症性疾患の病態における Malt1 プロテア ーゼ活性の関与 ... 35
第1節 緒言 ... 35
第2節 材料および方法 ... 36
第1項 動物 ... 36
第2項 骨髄由来樹状細胞 (BMDC) の分化 ... 37
第3項 BMDCに対するFcγR刺激 ... 37
第4項 FcγR刺激によるin vivoサイトカイン発現 ... 38
第5項 In vivo ITPモデル ... 38
第6項 統計解析 ... 39
第3節 結果 ... 39
第1項 Malt1 PDマウス由来骨髄細胞のBMDCへの分化能 ... 39
第2項 BMDCに対するFcγR刺激によるサイトカイン産生 ... 40
第3項 FcγR刺激によるin vivoサイトカイン発現試験 ... 42
第4項 Malt1 PDマウスにおけるFcγRシグナル依存的な血小板減少 ... 42
第4節 考察 ... 43
第4章 総括... 46
参考文献 ... 51
謝辞 ... 61
4
第1章 序論
免疫とは、細菌やウイルスなどの病原体から生体を防御するために備わったシステ ムである。免疫システムには自然免疫と獲得免疫があり、いずれも病原体の排除に重 要な役割を果たしている。自然免疫は、樹状細胞やマクロファージ、そしてナチュラル キラー細胞などの免疫細胞が、細菌やウイルスを非特異的に認識し、貪食作用やサイ トカイン産生により排除する。獲得免疫は、主に CD4 陽性のヘルパーT 細胞 (helper T細胞; Th細胞) と、CD8陽性の細胞傷害性T細胞 (cytotoxic T lymphocytes; CTL)、 並びにB細胞が重要な役割を果たす。Th細胞とCTLは、様々な外来性の抗原に対 応するため、多様なT細胞受容体 (T-cell receptor; TCR) を発現しており、リンパ節に おいて樹状細胞などの抗原提示細胞から各々の TCR に特異的な抗原の提示を受け、
活性化される。Th 細胞は、活性化するとインターフェロン-γ (interferon-γ; IFN-γ)、イン ターロイキン-4 (interleukin-4; IL-4)、IL-17などのサイトカインを産生して自然免疫細胞 を活性化させたり、B細胞と相互作用して抗体産生を誘導したりする。CTLもTh細胞 と同様に抗原提示を受けて活性化すると、IFN-γ や腫瘍壊死因子-α (tumor necrosis
factor-α; TNF-α) などのサイトカインや、パーフォリンやグランザイムなどの細胞傷害因
子を産生し、その抗原を排除する。B細胞は、T細胞と同様に多様な種類のB細胞受
容体 (B-cell receptor; BCR) を発現しており、様々な病原体に対して反応することを
可能にしている。B 細胞は Th細胞の働きなどにより形質細胞へと分化すると、抗原に 特異的な抗体を産生し、抗原を排除する。こうした獲得免疫はヒトにとって非常に重要 なシステムであり、獲得免疫が働かないと X 連鎖無ガンマグロブリン血症などの免疫 不全を引き起こす (以上、[1]を参照)。
しかしながら、本来外来性の抗原に対して反応するシステムが、自己の細胞や組織 を抗原として認識し、過剰な炎症反応を起こすと、自己免疫疾患を発症する。自己免
5
疫疾患は、臓器特異的、或いは全身で炎症を起こし、多様な病態を示す。例えば、炎 症性腸疾患 (inflammatory bowel disease; IBD) の患者では、活性化T細胞が大腸と 小腸で過剰な炎症反応を起こして、慢性的な下痢や腹痛を引き起こす。多発性硬化 症の患者では、脳や脊髄といった中枢組織で B 細胞やT 細胞が炎症反応を起こし、
四肢の麻痺や失明などの様々な神経症状をもたらす。血液で起こる自己免疫疾患と しては免疫性血小板減少性紫斑病 (immune thrombocytopenic purpura; ITP) や、自 己免疫性溶血性貧血 (Autoimmune hemolytic anemia; AHA) が知られ、それぞれ血 小板や赤血球の自己抗体が産生され、自己抗体を介した自然免疫細胞の活性化、
並びに貪食作用により血小板や赤血球が減少する。
このように、自己免疫疾患は様々な臓器や全身で多様な病態を示すが、その病因は 大きく分けて二種類に特徴づけられる。1) 細胞依存的な免疫反応と、2) 自己抗体依 存的な免疫反応である。細胞依存的な免疫反応の病態生理は完全には解明されて いないが、Th細胞が重要な役割を果たしていると考えられている。Th細胞は、活性 化していないナイーブな状態のnaïve Th細胞から、抗原提示によるTCRの刺激と共 に自然免疫細胞から産生されるIL-12の刺激を受けてIFN-γ産生性のTh1細胞に 分化する。同様にして、TCRとIL-4などの刺激によりIL-4やIL-13産生性のTh2細 胞に、さらにTCRとIL-23、IL-6、トランスフォーミング増殖因子-β (transforming growth factor-β; TGF-β) の刺激を受けてIL-17産生性のTh17細胞へと分化する (Figure 1) [2]。また、近年はTCR とTGF-βの刺激で制御性T細胞 (regulatory T cell; Treg cell) へと分化することも確認されており、Th1/Th2/Th17細胞の過剰な免疫 反応を抑制すると考えられている (Figure 1) [3]。従来は過剰なTh1細胞の活性化が 自己免疫疾患に、Th2細胞の活性化がアレルギーの発症に関与すると考えられてき たが、近年Th17細胞が自己免疫疾患における炎症反応に重要な役割を果たすこと が示唆され、注目されている。例えば、クローン病や潰瘍性大腸炎に代表されるIBD
6
患者の病変部位において、Th17細胞の浸潤が健常者と比較して多く認められている [4]。また多発性硬化症の動物モデルである実験的自己免疫性脳脊髄炎
(experimental autoimmune encephalomyelitis; EAE) モデルにおいては、Th17細胞が EAEの発症に重要な役割を果たしていることが判明している [5]。また皮膚角化疾患 である乾癬患者においても、Th17細胞が産生するIL-17A、IL-17F、IL-22といった複 数の炎症性サイトカインが角化細胞の増殖を促進し、発疹を引き起こしている [6]。こ のように、細胞依存的な自己免疫疾患においては、Th17細胞などのTh細胞が、過 剰な炎症反応の惹起に重要な役割を果たしていると考えられている。
Figure 1. Differentiation of helper T cells [2]
Naïve CD4+CD62L+ T cells are activated by antigen stimulation via T cell receptor (TCR). Continuous antigen and specific cytokine stimulation induce the differentiation to IFN-γ-producing Th1 cells, IL-4 and IL-13-producing Th2 cells, IL-17-producing Th17 cells, and IL-10-producing Treg cells.
一方で、自己抗体依存的な免疫応答を介した自己免疫疾患の発症には、以下の 三つの機序が知られている (Figure 2)。1) 自己の細胞の表面抗原や基底膜に対する 抗体が産生され、マクロファージや補体が活性化することで、細胞傷害や組織損傷を
7
引き起こすもの (ITP、AHA や、グッドパスチャー症候群など)。2) 自己の細胞と、そ れに対する抗体が免疫複合体 (Immune complex) を形成し、好中球などの免疫細胞 を活性化して組織損傷を引き起こすもの (全身性エリテマトーデス (systemic lupus
erythematosus; SLE) など)。3) 自己抗体そのものが直接的に自己抗原に作用するも
の(重症筋無力症、バセドウ病など)。これらのうち、3) 以外は、免疫グロブリン G (IgG) 抗体のFc鎖が自然免疫細胞のFcγ受容体 (Fcγ receptor; FcγR) と相互作用 し、FcγR シグナルが活性化することで、サイトカインの産生と、貪食作用を促進してい る。
Figure 2. Pathophysiology of autoantibody-dependent immune reaction [1]
こうした自己免疫疾患の治療には、ステロイド製剤や、免疫抑制剤 (チオプリン製 剤やカルシニューリン阻害剤等) が広く使われている。これらの薬剤は様々な自己免 疫疾患で有効であるが、感染症や骨粗しょう症、白血球減少症や腎毒性など様々な 副作用を引き起こすため、長期投与が困難である [7-9]。近年は、これら低分子の薬 剤に加えて、抗 TNF-α 抗体、抗 α4β7 抗体などの抗体医薬が上市され、ステロイド製 剤や免疫抑制剤が無効な重篤な患者においても有効性が認められた [10-12]。しか しながら、なお無効な症例が存在し、インフュージョンリアクションなどの副作用が見ら れることもある [13-15]。このように、抗体医薬などの新薬が登場してもなお、経口投与 可能で、かつ新たな作用機序の薬剤は待ち望まれている。
そこで筆者は、粘膜関連リンパ組織リンパ腫転座 1 (Mucosa-associated lymphoid
8
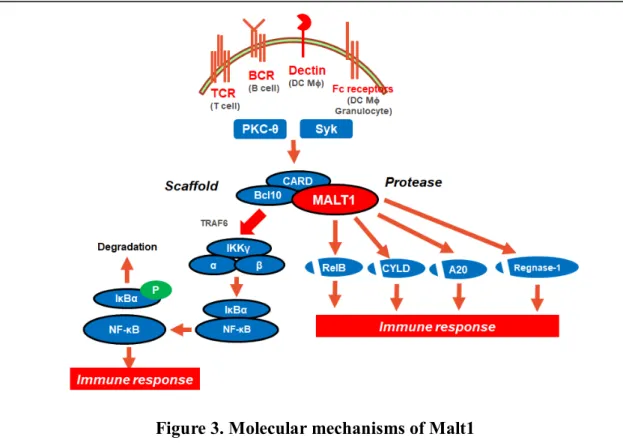
tissue lymphoma translocation 1; Malt1) の分子メカニズムに着目した。本分子は、粘 膜関連リンパ組織リンパ腫 (MALT リンパ腫) の患者における染色体異常として発見 された。本分子の一部が転座してcellular inhibitor of apoptosis protein 2 (cIAP2) と融 合遺伝子を形成し、MALTリンパ腫の発症に重要な役割を果たすことから、MALTリン パ腫転座1 (MALT lymphoma translocation 1) と名付けられた [16]。Malt1は、T 細 胞、B 細胞、並びに自然免疫細胞に発現し、その活性化に寄与する分子である
(Figure 3)。まず、自己の抗原や自己抗体が、それぞれの受容体 (TCR や BCR、FcR
など) に結合すると、T 細胞においてはプロテインキナーゼ C-θ (protein kinase C-θ, PKC-θ) が、B 細 胞 と自 然 免 疫 細 胞 にお い ては脾 臓 チロシ ンキナ ー ゼ (spleen tyrosine kinase; Syk) がそれぞれ活性化される [17, 18]。続いて、PKC-θ と Syk は、
CARD9やCARD11などのcaspase activation and recruitment domain (CARD) を含 むタンパクをリン酸化し、Malt1とBcl10と共に三量体 (CBM複合体) を形成させる。
CBM複合体が形成されると、さらにTRAF6と、それに続くIKK複合体の活性化を誘 導する [19]。IKK 複合体が活性化すると、NF-κB と結合している inhibitory κBα
(IκBα) をリン酸化し、プロテアソームによる分解を誘導する。その結果単量体となった
NF-κB は核内に移行し、転写因子としてサイトカインの産生誘導や、細胞の活性化・
増殖を引き起こす。このように、Malt1 は NF-κB の活性化に必須の分子で、その結果 生じる炎症反応に重要な役割を果たしている。実際に、Malt1ノックアウトマウスにおい ては、EAEモデルの発症が抑制され [20, 21]、TCR刺激によるIL-2産生の抑制や、
BCRシグナルの抑制・並びに免疫グロブリンE (IgE) の受容体であるFcε受容体シグ ナルの抑制も認められている [22, 23]。このように、Malt1の機能を抑制することは、自 己免疫疾患に高い効果があると考えられるが、Malt1 は CBM 複合体を形成して機能 を発揮する、いわば足場タンパクとして働くため、そのタンパク質同士の相互作用を低 分子化合物で阻害することは極めて困難であった。また抗体は細胞膜を通過できない
9
ため、CBM複合体の形成を抗体医薬で抑制することも不可能であった。
Figure 3. Molecular mechanisms of Malt1
そこで、異なるアプローチとして Malt1 がシステインプロテアーゼとして働くことに着
目した。Malt1 はパラカスパーゼとしても知られるが、これはカスパーゼ様タンパク質フ
ァミリーの一つとして2000年に同定された際に名付けられたものである (Figure 4) [24]。 パラカスパーゼは動物および粘菌に存在するが、哺乳類のパラカスパーゼは、現在 Malt1以外見つかっていない [25]。
10
Figure 4. Structure of Malt1 [26]
MALT1 protein contains an N‑terminal death domain, followed by two immunoglobulin-like domains (Ig), a caspase-like domain and a C‑terminal region that contains another immunoglobulin-like domain.
パラカスパーゼ Malt1 がシステインプロテアーゼとして働くことは 2008 年に初めて 報告され [27, 28]、その後RelB、CYLD、A20、Regnase-1、RoquinなどがMalt1の基 質として働くことが示された [27-31]。これらの基質は炎症反応に対して負に働くと考 えられているため、Malt1 のプロテアーゼ活性によりこれらの基質が分解されると、NF- κBの活性化が亢進すると考えられている。そこで筆者は、Malt1の基質結合部位に結 合する低分子阻害剤を用いて、Malt1のプロテアーゼ活性を抑制し、炎症反応を制御 することができるのではないかと考えた。しかしながら、こうした Malt1 のプロテアーゼ 活性が、生体内でどの程度炎症反応に関与するかは定かではなかった。Malt1 ノック アウトマウスでは、Malt1 のプロテアーゼ活性のみならず、足場タンパクとしての機能も 失うため、Malt1のプロテアーゼ活性そのものの炎症反応への寄与は不明であった。
そこで本博士論文において、システインプロテアーゼの活性中心であるシステイン をアラニンに変異させてMalt1の活性を欠損させたマウス (Malt1 protease-deficientマ
ウス; Malt1 PDマウス) を利用し、第2章で大腸炎の病態におけるMalt1の寄与、第
3 章で免疫性血小板減少性紫斑病の病態における Malt1 の寄与について解析を行 った。
11
第2章 Malt1プロテアーゼ活性の不活化によるTh17とTh1/17細胞の
制御と大腸炎の抑制効果
第1節 緒言
クローン病と潰瘍性大腸炎に代表される炎症性腸疾患 (IBD) とは、慢性的な下痢 と、それに伴う腹痛が生活の質を著しく低下させる自己免疫疾患である。重度のクロー ン病患者では、下痢に伴って脱肛や瘻孔が起こり、大腸摘出や人工肛門などの外科 治療が必要になる患者も存在する。IBDの患者に対しては、ステロイド製剤や免疫抑 制剤が処方されるが、副作用の懸念があるため、長期の服用は難しい [32]。これら単 剤で臨床的寛解が達成できなかった患者に対しては、二次治療として抗TNF-α抗体 や抗α4β7抗体の使用が認められている [33-35]。これらの抗体医薬はIBDのほとん どの患者に有効であるが、一部の患者には耐性があり、薬効を示さない [14, 15]。こ のような患者に対して、革新的な作用機序をもった新たな治療薬が待ち望まれてい る。
IBDの病因には、古くからCD4陽性ヘルパーT細胞 (Th細胞) が重要な役割を 果たすと考えられてきた。第一章で述べたように、Th細胞にはいくつかのサブタイプ があり、IFN-γ産生性Th1細胞、IL-4産生性Th2細胞、そしてIL-17産生性Th17細 胞が良く知られている。これらのバランスが破綻すると、過剰な炎症反応が起こり、IBD などの自己免疫疾患が発症すると考えられている。その中で、本論文ではIL-23/Th17 連関 (IL-23/Th17 axis) とIBDの病態形成に着目した。抗IL-23抗体は、抗TNF-α 抗体ナイーブ(未治療)のクローン病患者においても、抗TNF-α抗体に耐性のある患 者においても、感染症などの重篤な副作用もなく有意な薬効を示した [36]。加えて
IL-23はTh17細胞の分化並びに活性化を誘導することが知られているため [37,
38]、IL-23/Th17 axisをターゲットとする治療戦略はIBDにおいて有望なアプローチ
12
であると考えた。また、IL-17とIFN-γをいずれも産生するTh1/17細胞もまた、多発性 硬化症やIBDの病因に関与することが示唆されていることから [39-42]、Th17細胞と
Th1/17細胞の制御は、IBDの治療に有望であると考えた。
粘膜関連リンパ組織リンパ腫転座1 (Malt1) は、T細胞を含むリンパ球の活性化に 重要な役割を果たす分子である [22]。抗原がT細胞受容体 (TCR) を刺激すると、
Malt1とBcl10はCARD11と結合して三量体を形成し、 続いてNF-κBの活性化を
誘導する [43]。こうした足場タンパクとしての機能に加えて、Malt1はシステインプロテ
アーゼとして働き、RelB [27] や、CYLD [29]、そしてA20 [28] といったタンパク質を 分解する。これらの基質は免疫反応に対して負に働くため、Malt1依存的なこれらの 基質の分解は、T細胞の活性化を亢進すると考えられている。Malt1分子は様々な免 疫反応に重要な役割を果たすと考えられているが、Malt1のシステインプロテアーゼと しての機能が、生体内でどの程度大腸の炎症反応に寄与するか、明らかではなかっ た。
本章において、Malt1プロテアーゼ活性欠損マウス (Malt1 PDマウス) を用いて、
T細胞依存的な大腸炎の発症に対するMalt1のプロテアーゼ活性の関与と、その作 用機序を解析した。
第2節 材料および方法
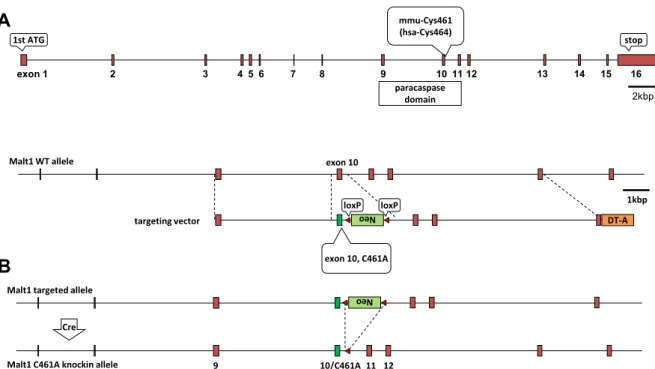
第1項 Malt1 PDマウスの作出
Malt1 PD マウスは、BALB/c 由来の遺伝子組換えマウスで、武田薬品工業株式会
社 (日本、神奈川) にて構築された [44]。まず、Malt1 の活性中心であるシステイン (Cys461) をアラニンに変異させたMalt1 Cys461Ala (C461A) と、ネオマイシン耐性遺 伝子を搭載したターゲットベクターを細菌人工染色体 (bacteria artificial chromosome;
13
BAC) で構築した。BAC ベクターを BALB/c マウス由来の胚性幹細胞 (embryonic
stem cell; ES cell) に導入し、ネオマイシン耐性株の中から定量 polymerase chain reaction (qPCR) で相同組換え株をスクリーニングした (Figure 5A)。続いてCreベクタ ーを一過的に発現させて、ネオマイシン耐性遺伝子を除去した (Figure 5B)。この ES
細胞を C57BL/6 マウス由来の胚盤胞に挿入してキメラマウスを作出し、キメラマウスを
雌性BALB/cマウスと掛け合わせることでヘテロ接合体のMalt1 PDマウスを作出した
(Malt PD/WTマウス)。このMalt PD/WTマウスを掛け合わせて、ホモ接合体のMalt1 PDマ ウス (Malt1PD/PD マウス) と野生型マウス (Malt1WT/WT マウス) を作出した。野生型
(wild-type; WT) マウスと、ホモ接合体の Malt1 PD マウスを見分けるため、以下に示
すプライマーを用いてジェノタイピングを実施した。
Forward: 5'-TAAGGCAATTCTGTACGGTGGTAAGAACC-3' Reverse: 5'-GGGAGGAGTAATCACTACTTCTGACCCTTG-3'
WTマウスでは、約300 bpのバンドが、ホモ接合体のMalt1 PDマウスでは390 bpの バンドが認められた。一方、ヘテロ接合体のマウスでは、2つのバンドが確認できた。こ のようにして得られたWTマウスとホモ接合体のMalt1 PDマウスを本研究に用いた。
14
Figure 5. Development Malt1 protease-deficient (PD) allele
A. Malt1 genomic structure (48kbp) (upper). The BAC vector harboring targeting vector with neomycin-resistance cassette was introduced into BALB/c embryonic stem (ES) cells (lower). B. The Cre vector was transiently transfected in the resulting targeted allele to delete the neomycin-resistance cassette.
第2項 動物
雌性 Malt1 PD マウスと WT マウスは、特定病原体の存在しない空間 (specific
pathogen-free; SPF) で飼育し、6~10 週齢で実験に用いた。雌性 BALB/c マウスは、
日本チャールス・リバー (日本) から購入した。雌性 C.B-17/Icr-scid マウス (SCID マ ウス) は日本クレア (日本) から購入した。SCIDマウスはレシピエントマウスとして用い、
ペーパーチップ上で単飼した。全ての動物実験は人道的ケアの基準に基づいて実施 し、武田薬品工業株式会社のInstitutional Animal Care and Use Committee (IACUC) の承認を得た上で実験を行った (実験番号: 4516、4525、並びに4531)。
exon 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
2kbp
1st ATG stop
paracaspase domain mmu-Cys461 (hsa-Cys464)
Malt1 WT allele
targeting vector
Neo loxP DT-A
Cre
loxP 1kbp
exon 10
exon 10, C461A Malt1 targeted allele
Malt1 C461A knockin allele
Neo
9 10/C461A 11 12
B A
15
第3項 T細胞の刺激とウエスタンブロッティング
Malt1 PDマウスとWTマウス由来の脾臓を破砕し、セルストレーナーに通して単細
胞画分を得た。この細胞に塩化アンモニウム溶液 (免疫生物研究所、日本) を加え て、溶血処理した。得られた脾細胞から、CD4+ T cell isolation kit II (Miltenyi Biotec、 ドイツ) を用いて、CD4+ T細胞をポジティブセレクションにより精製した。精製方法は キットに付属のプロトコルに従った。CD4+ T細胞は100 units/mLのペニシリン、100 μg/mLのストレプトマイシン、並びに10%のFBSを含むRPMI-1640培地に懸濁し、
6-wellプレートに播種した。その後プロテアソーム阻害剤のMG-132 (終濃度5 μM)、
或いは培地を添加し、30分間37℃で静置した。続いて、終濃度20 ng/mLのphorbol 12-myristate 13-acetate (PMA) (和光純薬工業、日本) と、終濃度250 ng/mLのイオノ マイシン (和光純薬工業、日本) を加えて、所定の時間刺激を加えた。ヒトT細胞の 株化細胞であるJurkat細胞を、MG-132存在下でPMAとイオノマイシンで60分間刺 激し、陽性対照とした。その後、プロテアーゼ阻害剤を含むRIPAバッファーを用いて 細胞ペレットを溶解し、ジチオトレイトール (DTT) を用いて還元処理を施した。細胞 のライセートをSDS-PAGEで分離した後、Trans-Blot Turboシステム (Bio-Rad、アメリ カ) を用いて、PDVFメンブレン (Bio-Rad、アメリカ) に転写した。メンブレンをPVDF ブロッキングバッファー (東洋紡、日本) で1時間ブロッキングした後、0.05% Tween
20を含むPBS (PBS-T) で洗浄した。続いて以下に示す一次抗体を用いて、一晩
4℃で振盪し、抗体反応を実施した。一次抗体の希釈液にはCan Get Signal Immunoreaction Enhancer Solution 1 (東洋紡、日本) を用いた。
一次抗体
抗RelB抗体 (クローン;C1E4) (Cell Signaling Technology、アメリカ) 抗CYLD抗体 (クローン; D1A10) (Cell Signaling Technology、アメリカ) 抗Malt1抗体 (クローン;H300) (Santa Cruz Biotechnology、アメリカ)
16
抗IκBα 抗体 (クローン;C-21) (Santa Cruz Biotechnology、アメリカ) 抗β-Actin抗体 (和光純薬工業、日本).
その後PBS-Tで洗浄し、以下に示す二次抗体を用いて、1時間4℃で抗体反応を
実施した。抗体の希釈液にはCan Get Signal Immunoreaction Enhancer Solution 2 (東 洋紡、日本) を用いた。
二次抗体
HRP結合抗マウスIgG (GE Healthcare、アメリカ) HRP結合抗ラビットIgG (GE Healthcare、アメリカ)
PBS-Tで洗浄した後、ECL Prime Western Blotting Detection Reagent (GE
Healthcare、アメリカ) をメンブレンに添加し、LAS-3000 (GE Healthcare、アメリカ) で 化学発光を検出した。
第4項 In vitroサイトカイン産生試験
第3項に示した方法で、Malt1 PDマウスとWTマウスから、CD4+ T細胞を精製し た。CD4+ T細胞は第3項と同様に抗生物質とFBSを含むRPMI-1640培地に懸濁 し、96-wllプレートに播種した。Malt1 PDマウスとWTマウス由来のCD4+ T細胞に、
第3項と同じ条件でPMAとイオノマイシン刺激を加え、24時間37℃で培養した。
TCRに対する刺激として、抗CD3/CD28抗体であるDynabeads Mouse T-activator CD3/CD28 (Life Technologies、アメリカ) をビーズ:細胞の比が1:1になるように加 え、24時間37℃で培養した。その後培養上清中のIL-2濃度をELISAで測定した。
ELISAはMouse IL-2 DuoSet ELISA (R&D Systems、アメリカ) を用いて、付属のプロ トコルに従って実施した。
17
第5項 抗CD3抗体誘発in vivoサイトカイン産生試験
抗CD3抗体 (クローン;145-2C11) (Bio X Cell, アメリカ) と、そのアイソタイプ抗体 (hamster IgG) (BD Biosciences, アメリカ) をそれぞれ5 μg/マウスとなるように、Malt1 PDマウスとWTマウスに腹腔内投与した。3時間後、全血を心臓穿刺により採取し、
ヘパリンナトリウム (味の素、日本) を用いて血漿画分を回収した。血漿中のIL-2、IL- 17とIFN-γの濃度をELISAで測定した。ELISAは全てR&D Systems (アメリカ) 製 のキットを用い、付属のプロトコルに従って実施した。
マウスより全血を採取した後、頚椎脱臼により安楽死させた。大腸を摘出し、生理食 塩水で洗浄した後にRNAlater (Qiagen、ドイツ) 中で 4°Cにて保存した。大腸組織 は第7項で述べる方法で、リアルタイム定量的逆転写PCR (quantitative reverse transcription-PCR; qRT-PCR) に供した。
第6項 Naïve T細胞移入マウス大腸炎モデル
まず、BALB/cマウス、Malt1 PDマウス並びにWTマウスから脾臓を取り出し、
Lympholyte-M Separation Media (CEDARLANE、カナダ) を用いて、2000×g、20分 間の条件で密度勾配遠心を実施し、単核球画分を採取した。この細胞に塩化アンモ ニウム溶液 (免疫生物研究所、日本) を加えて、溶血処理した。CD+CD62L+ naïve T 細胞の精製は、CD4+CD62L+ T Cell Isolation Kit, II (Miltenyi Biotec, ドイツ) を用い た。まず、ビオチンが結合した抗CD8a抗体、抗CD45R抗体、抗CD49b抗体、抗
CD11b抗体、抗Ter-119抗体、抗TCRγ/δ抗体、そして抗CD25抗体を細胞に加え
た。続いて、マグネットビーズが結合した抗ビオチン抗体を加えて、AutoMACS Pro Separator (Miltenyi Biotec、ドイツ) でCD4+CD25- T細胞画分をネガティブセレクショ ンにて精製した。続いて、マグネットビーズが結合した抗CD62L抗体を加えて、
AutoMACS Pro Separatorにてポジティブセレクションを行い、CD4+CD25- CD62L+ T
18
細胞画分を精製した。
このようにして得られたnaïve T細胞を、SCIDマウスの静脈内に移入して、大腸炎 を誘導した。正常群として、細胞を移入しない正常なSCIDマウスも評価した。SCID
マウスにnaïve T細胞を移入して21日後に下痢スコアとマウスの体重変動を評価し
た。下痢スコアは以下のように1-4のスコアで判定した: 1, 正常; 2, 軟便だが形は正
常; 3, 軟便で形が崩れている; 4, 下痢(水状の便がペーパーチップに付着してい
る)。下痢スコアとマウスの体重を測定した後に安楽死処置し、大腸と腸間膜リンパ節 を摘出した。大腸は生理食塩水で洗浄した後、重量を測定し、RNAlater中にて4℃
で保存した。大腸は第7項で述べる方法で、qRT-PCRを実施した。腸間膜リンパ節 は、第8項に示す方法でフローサイトメトリー解析を実施した。
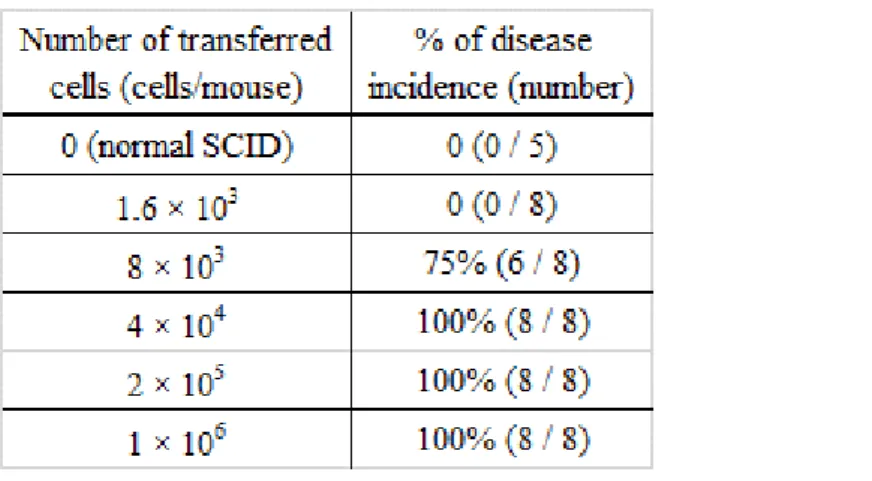
移入する細胞数を決定するための予備実験として、1.6 × 103、8.0 × 103、4.0 × 104、 2.0 × 105、並びに1.0 × 106 個のBALB/cマウス由来naïve T細胞をSCIDマウスの 静脈内に移入した。移入3週間後に下痢スコアと大腸重量を測定したところ、Figure 6 に示すように、下痢と大腸重量は移入細胞数依存的に増加していた。このとき、3週間 の時点での下痢の発症率、即ち下痢スコアが2以上を示した個体の割合をTable 1
に示す。4 × 104 個以上の条件で全例の発症が認められたため、以降の実験は4 ×
104 個の条件で実施した。
19
Figure 6. Transfer of naïve T cells into SCID mice induced colitis associated with diarrhea in a cell number-dependent manner
Diarrhea score and colon weight were determined 3 weeks later in a blind manner. A.
Diarrhea score. B. Colon weight. Values are presented as the mean ± SEM of 5 (normal SCID mice), or 8 (other groups).
Table 1. Disease incidence of naïve T-cell transferred colitis model
Naïve T cells were intravenously transferred to the SCID mice. Three weeks later, the number of incidence for diarrhea (score; >2) was counted.
第7項 リアルタイムqRT-PCR
大腸の総RNAはRNeasy Mini Kit (Qiagen、ドイツ) とDNaseI (Qiagen、ドイツ) を
20
用 い て、キッ トに付 属 のプロ トコ ル に従 っ て 抽 出 し た。NanoDrop (Thermo Fisher Scientific、アメリカ) を用いてRNA濃度を測定した後、High Capacity cDNA Reverse Transcription Kit (Life Technologies、アメリカ) を用いて逆転写反応を実施し、cDNA を調製した。 続いて、TaqMan Fast Advanced Master Mix (Life Technologies、アメリカ) とTaqMan Gene Expression Assays (Life Technologies、アメリカ) の各プライマー・プロ ーブミックスを用いてqPCRを実施した。qPCR反応にはViiA 7 Real-Time PCR System (Life Technologiesアメリカ) を用いた。TaqMan Gene Expression Assaysのプライマー
とFAM標識TaqManプローブのセットは、以下の各Assay IDのものを使用した。
IL-17A (遺伝子名Il17a): Mm00439618_m1.
IFN-γ (遺伝子名Ifng): Mm00801778_m1.
内 部 標 準 遺 伝 子 は VIC/MGB 標 識 の マ ウ ス β-actin (Actb) コ ントロ ー ル (Life
Technologies、アメリカ) を用いて、⊿⊿Ct 法によりコントロール群と比較した相対発現
量を計算した。
第8項 腸間膜リンパ節のCD4+ T細胞における細胞内サイトカイン染色
第6項で採取した大腸炎モデルの腸間膜リンパ節を、セルストレーナー付きのチュ ーブ上で破砕し、単細胞画分を回収した。この細胞を抗生物質とFBSを含むRPMI- 1640培地に懸濁し、96-wellプレートに播種した。50 ng/mLのPMA、1 μg/mLのイオ ノマイシンと、モネンシンを含むGoldiStop Protein Transport Inhibitor (BD
Biosciences, アメリカ) を加えて、37℃で4時間培養した。FcγRを抗CD16/CD32抗 体 (クローン; 2.4G2) (BD Biosciences、アメリカ) でブロッキングした後、PE標識抗 CD4抗体 (BioLegend、アメリカ) で染色した。染色した細胞は、
Fixation/Permeabilization solution (BD Biosciences、アメリカ) を用いて固定化及び膜 透過処理を実施した。細胞内サイトカイン染色は、FITC標識抗IFN-γ抗体と、Alexa
21
Fluor 647標識抗IL-17A抗体 (BioLegend、アメリカ) を用いて実施した。フローサイ トメトリー解析はBD Accuri C6 Flow Cytometer (BD Biosciences、アメリカ) を用いた。
Th17、Th1/17とTh1細胞の割合は、以下のように定義した。
Th17細胞: CD4陽性細胞中のIL-17+IFN-γ-細胞の割合 Th1/17細胞: CD4陽性細胞中のIL-17+IFN-γ+細胞の割合 Th1細胞: CD4陽性細胞中のIL-17-IFN-γ+細胞の割合
第9項 統計解析
データは全て平均値 ± SEM で表記した。統計解析は SAS System for Windows
(SAS Institute、アメリカ) を用いて実施した。2 群間の差は、以下のように統計処理し
た。まず分散比のF検定を実施し、等分散とみなせる場合、スチューデントのt検定を 実施した。不等分散の場合は、ウェルチのt検定を実施した。下痢スコアはノンパラメト リックなスコアであるため、下痢スコアの比較のみウィルコクソンの順位和検定を実施し た。有意水準は 0.05とし、p値 (probability value) が p<0.05のとき統計学的に有意 とみなした。
第3節 結果
第1項 Malt1 PDマウスにおけるプロテアーゼ活性の不活化検討
Malt1プロテアーゼ活性の生体内での機能を検討するため、武田薬品工業株式会
社において、Malt1 PDマウスを構築した。本マウスは、Malt1の活性中心のシステイン がアラニンに変異しており、Malt1のプロテアーゼ活性が欠損していることが期待され た。まず、Malt1 PDマウスのMalt1タンパクの発現を調べたところ、Malt1 PDマウスと WTマウスで差は認められなかった (Figure 7A)。Malt1のプロテアーゼ活性の有無
22
は、MG-132存在下でPMA/イオノマイシン刺激し、Malt1の基質であるRelBと
CYLDの分解産物を確認することで評価した。MG-132はプロテアソーム阻害剤で、
基質の分解産物がプロテアソームで速やかに分解されるのを防ぐ目的で加えた。
Figure 7Aと7Bに示すように、WTマウス由来のCD4+ T細胞では、PMA/イオノマイ シン刺激によるRelBとCYLDの分解産物が各時点で認められた。一方で、Malt1 PD マウス由来のCD4+ T細胞では、RelBとCYLDの分解産物は見られなかった。
Figure 7Cに示すように、各系統由来のCD4+ T細胞をMG-132非存在下でPMA/イ オノマイシン刺激したところ、いずれの系統においてもIκBαの分解が認められた。
23
Figure 7. Malt1 protease-deficient (PD) mice express inactive form of Malt1 Purified CD4+ T cells from wild-type (WT) and Malt1 PD mice were stimulated with PMA and ionomycin (P/I) for indicated time points. A. Stimulation was conducted with MG-132 and western blot was performed with antibodies against Malt1, RelB and β- Actin. B. Stimulation and western blotting were conducted with MG-132 and antibodies against cylindromatosis (CYLD), respectively. C. Stimulation was conducted without MG-132 and western blotting was performed with antibodies against IκBα and β-Actin.
A 0
WT PD
RelB
β-Actin P/I (min.)
Jurkat 60
Malt1
RelB digested RelB 30
WT PD 60 WT PD
90 WT PD
120 WT PD
CYLD
digested CYLD 0
WT PD
P/I (min.) 30
WT PD 60 WT PD
90 WT PD
120 WT PD
IκBα β-Actin
0 WT PD
P/I (min.) 5
WT PD 15 WT PD
30 WT PD
60 WT PD
B
C
24
第2項 Malt1 PDマウス由来T細胞のIL-2産生能
続いて、in vitroにおけるT細胞活性化に対するMalt1のプロテアーゼ活性の効果
を調べた。Malt1 PDマウスとWTマウスからCD4+ T細胞を精製し、PMA/イオノマイ シン、或いは抗CD3/CD28抗体で刺激した。Figure 8Aと8Bに示すように、いずれの 刺激においてもMalt1 PDマウス由来のCD4+ T細胞では、WTマウスと比較してIL-2 の産生量は有意に低かった。
Figure 8. Impaired IL-2 production from CD4+ T cells isolated from Malt1 protease-deficient (PD) mice
Purified CD4+ T cells from wild-type (WT) and Malt1 PD mice were stimulated with A.
PMA/ionomycin (P/I) or B. anti-CD3/CD28 antibodies for 24 h. Concentrations of IL-2 in supernatants were measured by ELISA. Values are presented as the mean ± SEM of 3.
#p<0.05 (Student's t-test) versus WT cells.
第3項 Malt1 PDマウスにおける抗CD3抗体刺激によるin vivoサイトカイン産生試 験
次に、これらの系統におけるin vivoでのT細胞の活性化に対するMalt1のプロテ アーゼ活性の効果を評価した。抗CD3アゴニスト抗体をWTマウスに腹腔内投与す ると、血漿中IL-2、IL-17及びIFN-γの濃度が上昇した (Figure 9A – C)。 Malt1 PD
25
マウスに同様の処置を施したところ、IL-2とIL-17の産生量はWTマウスと比較して有 意に低かった (Figures 9AとB)。一方で、血漿中のIFN-γの濃度は、WTマウスと比 較して有意な差は認められなかった (Figure 9C)。加えて各マウスの大腸を摘出し、
IL-17AとIFN-γの遺伝子発現量を比較したところ、血漿中サイトカイン濃度の結果と
同様に、Malt1 PDマウスの大腸では、IL-17Aの発現量はWTマウスと比較して有意
に低く、IFN-γの発現量は有意な差が認められなかった (Figure 9DとE)。
26
Figure 9. in vivo impaired cytokine release in Malt1 protease-deficient (PD) mice Anti-CD3 antibody was intraperitoneally injected into wild-type (WT) and Malt1 PD mice, followed by obtaining whole bloods 3 h later. Cytokine concentrations in the plasma were determined by ELISA. Colonic mRNA expressions were measured by quantitative reverse transcription-PCR (qRT-PCR). A. IL-2 concentration. B. IL-17 concentration. C.
IFN-γ concentration. D. IL-17A mRNA in the colon. E. IFN-γ mRNA in the colon. Values are presented as the mean ± SEM of 5. #p<0.05 (Student's t-test) versus WT mice.
+p<0.05 (Welch's t-test) versus WT mice.
27
第 4項 Naïve T細胞移入マウス大腸炎モデルの病態における Malt1プロテアーゼ
不活化の影響
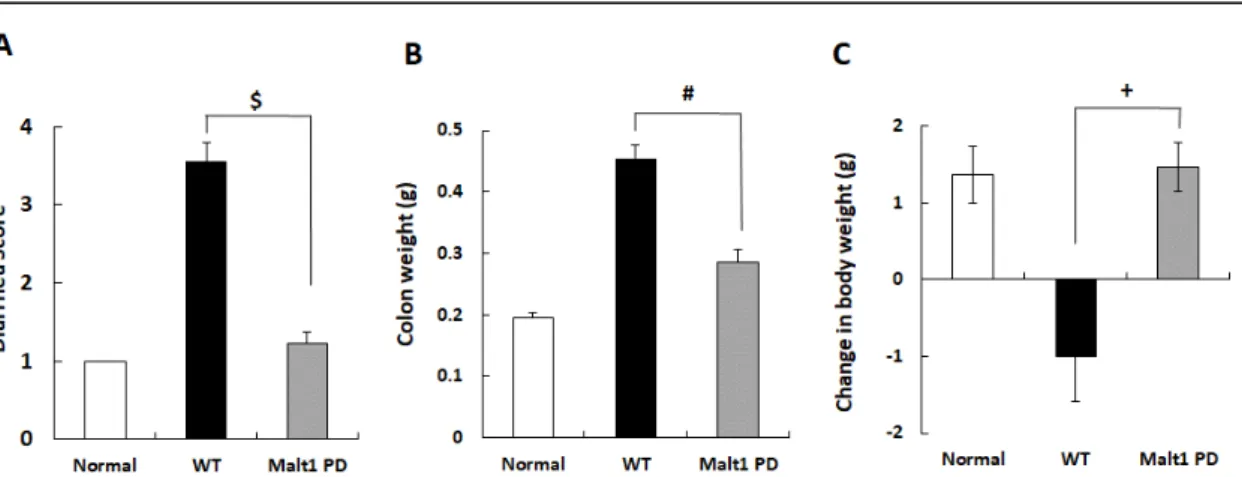
Malt1 PDマウスとWTマウス由来のCD4+CD62L+ naïve T細胞を精製し、4 × 104 個/マウスとなるように、SCIDマウスの静脈内に移入した。3週間後、下痢スコアと大腸 重量、SCIDマウスの体重変化を評価した。Malt1 PDマウス由来のnaïve T細胞を移 入した群では、WTマウス由来の細胞を移入した群と比較して下痢スコアと大腸重量 の増加は有意に低かった (Figure 10AとB)。加えて、Malt1 PDマウス由来のT細胞 を移入した群では、WTマウス由来のT細胞を移入した群で見られた体重の減少が 見られなかった (Figure 10C)。
Figure 10. Naïve T cells from Malt1 protease-deficient (PD) mice induced colitis with less severity
Naïve T cells (4 × 104 cells/mouse) isolated from Malt1 PD and wild-type (WT) mice were intravenously injected into the SCID mice, followed by harvesting the colon 3 weeks later. A. Diarrhea score. B. Colon weight. C. Change in body weight. Values are presented as the mean ± SEM of 4 (normal), or 9 (WT and PD-derived cells-transferred groups).
$p<0.05 (Wilcoxon test), #p<0.05 (Student's t-test), and +p<0.05 (Welch's t-test) versus WT group.
28
第5項 大腸炎モデルの腸間膜リンパ節におけるTh17細胞とTh1/17細胞の比較 大腸炎モデルの腸間膜リンパ節において、CD4+ T 細胞中のTh17、Th1/17とTh1 細胞の割合をフローサイトメトリー解析で調べた。SCIDマウスはT細胞を含まないた め、正常群の腸間膜リンパ節の評価は行わなかった。Figure 11Aに示すように、WT マウス由来のnaïve T細胞を移入して3週間後の腸間膜リンパ節においては、Th17、
Th1/17とTh1細胞の蓄積が認められた。Malt1 PDマウス由来のT細胞を移入した
群では、WTマウス由来細胞移入群と比較して、CD4+ T細胞中のTh17とTh1/17細 胞の割合は有意に低かった (Figure 11BとC)。一方、Th1細胞の割合はほとんど差 がなかった (Figure 11D)。
29
Figure 11. The population of CD4+ Th cells from mesenteric lymph nodes of the mouse receiving Malt1 protease-deficient (PD) and wild-type (WT) T cells The population of Th17, Th1/17 and Th1 cells gated on CD4+ T cells were determined by flow cytometry. A. Representative image of flow cytometry analysis. B. The percentage of Th17 cells. C. The percentage of Th1/17 cells. D. The percentage of Th1 cells. Data are presented as the mean ± SEM of 9 (WT and Malt1 PD groups). +p<0.05 (Welch's t-test) versus WT group.
第6項 大腸炎モデルにおける大腸のサイトカイン発現量の比較
最後に、マウス大腸炎モデルの病変部位である大腸におけるIL-17AとIFN-γの発 現量に対するMalt1プロテアーゼ活性の関与を調べた。Figure 12 Aに示すように、
WTマウス由来のT細胞を移入したマウスでのばらつきが大きかったため有意な差で
30
はなかったものの、Malt1 PDマウス由来のT細胞を移入したマウスにおいて、IL-17A の発現量の低下傾向が認められた。IFN-γの発現量は、Malt1 PDマウス由来のT細 胞を移入したマウスにおいて、WTと比較して僅かな低下が認められるのみであった (Figure 12 B)。
Figure 12. Colonic cytokine expression in the mice receiving Malt1 protease- deficient (PD) and wild-type (WT) T cells
Colonic mRNA expressions were determined by quantitative RT-PCR. A. IL-17A mRNA.
B. IFN-γ mRNA. Values are presented as the mean ±SEM of 3 (normal), or 9 (WT and Malt1 PD T cells groups). +p<0.05 (Welch's t-test) versus WT group.
第4節 考察
本論文において、T細胞を刺激したときの血中と大腸のIL-17Aの産生に、Malt1の プロテアーゼ活性が重要な役割を果たすことを示した。加えて、Malt1 プロテアーゼの 不活性化が、Th17 細胞と Th1/17 細胞の活性化を制御し、大腸炎の発症を抑制する ことを見出した。
武田薬品工業株式会社にてMalt1 PDマウスの構築に成功したため [44]、本系統
31
のMalt1プロテアーゼ活性の有無をウエスタンブロッティングで調べることにした。
PMAは、T細胞においてはPKC-θを活性化し、Malt1プロテアーゼの活性化と CBM複合体の形成を通じたNF-κB の活性化を誘導する (Figure 3)。Wegenerらの 研究では、Malt1ノックアウトマウス由来の細胞にPMA/イオノマイシン刺激すると、
CBM複合体の形成が阻害され、IκBαの分解能が低下していた [45]。この結果は第 1章で述べたように、CBM複合体の形成によりIκBαの分解が亢進し、NF-κB の活 性化が誘導されることを示している。本研究において、Malt1 PDマウスとWTマウス由 来のCD4+ T細胞を精製し、PMA/イオノマイシン刺激を施したところ、いずれの系統 においてもIκBαの分解が亢進しており (Figure 7C)、Malt1のタンパク量も同程度で あった (Figure 7A)。この結果から、Malt1 PDマウスは正常にCBM 複合体を形成し ていると考えた。一方で、Malt1のプロテアーゼ依存的なRelBとCYLDの分解は Malt1 PDマウス由来のCD4+ T 細胞では認められなかった (Figure 7AとB)。これら の結果から、Malt1ノックアウトマウスとは異なり、Malt1 PDマウスではMalt1の足場タ ンパクとしての機能は保持されておりCBM複合体を正常に形成するが、Malt1のプロ テアーゼとしての機能は欠損していると考えた。この系統を用いて、T細胞の活性化と 大腸の炎症、並びにTh細胞の制御に対するMalt1のプロテアーゼ活性の寄与を評 価した。
まず、Malt1のプロテアーゼ活性の欠損がTCR刺激によるin vitroとin vivoのT 細胞の活性化に影響を与えるか否かを調べた。Figure 8に示したようにMalt1 PDマ ウス由来のCD4+ T細胞では、PMA/イオノマイシン刺激と、抗CD3/CD28刺激による IL-2の産生量がWTマウスと比較して有意に低かった。Malt1 PDマウスに抗CD3抗 体を腹腔内投与したところ、血中におけるIL-2の産生量はWTと比較して低かった (Figure 9A)。IL-2産生はMalt1ノックアウトマウスでは完全に抑制されるが [22]、
Malt1 PDマウスでは部分的かつ有意な低下が認められた。これらの結果から、Malt1
32
のプロテアーゼ活性はNF-κB活性化に対する抑制因子を分解することでNF-κBの 活性化を誘導するが、NF-κBの機能を完全には制御せず、Malt1のCBM複合体と しての機能もNF-κBの活性化に一定程度影響を及ぼすことが示唆された。興味深い
ことに、Malt1 PDマウスを抗CD3抗体で刺激したところ、IL-17の産生量が顕著に減
少していた (Figure 9B)。一方、血漿中のIFN-γ濃度は変化がなかった (Figure 9C)。 加えて、大腸におけるIL-17Aの遺伝子発現量は、WTマウスと比較して有意に低か ったが、IFN-γの発現量は変わらなかった (Figure 9DとE)。このように、Malt1のプロ テアーゼ活性は、末梢血と大腸いずれにおいても、IL-17産生性のT細胞の活性化 を制御しており、TCR刺激による急性炎症に寄与していることが示唆された。
続いて、naïve CD4+CD62L+ T細胞をSCIDマウスに移入することで誘導される大 腸炎の病態に対するMalt1のプロテアーゼ活性の関与を調べた。T細胞移入大腸炎 モデルとしては、CD4+CD45RBhighT細胞移入モデル [46] や、Concanavalin A誘発 活性化CD4+T細胞移入モデル [47] など、いくつかのモデルが報告されているが、
本研究においては、武田薬品工業株式会社において構築された、CD4+CD62L+ T細 胞移入大腸炎モデルを用いた。本モデルはnaïve CD4+CD62L+ T細胞をマグネットビ ーズで精製し、SCIDマウスに移入することで、安定して下痢の発症と大腸重量の増 加が認められるモデルである [48]。本モデルを用いて、Malt1 PDマウス由来のnaïve T細胞をSCIDマウスに移入したところ、下痢スコア、大腸重量の増加、体重減少とい ったいずれの評価指標も、WTマウス由来T細胞移入群と比較して低かった (Figure 10)。このとき、大腸の病理学的所見を武田薬品工業株式会社にて評価したところ、
WTマウス由来naïve T細胞を移入したマウスの大腸では、杯細胞の破壊や炎症性細 胞の浸潤に伴って腸粘膜の肥厚が認められたが、Malt1 PDマウス由来のT細胞移 入群では、わずかな粘膜肥大と細胞の浸潤が誘導された程度で、中等度以上の炎症 症状は見られなかった [49]。これらの結果から、Jaworskiらの研究と同様に [50]、
33
Malt1のプロテアーゼ活性が生体内においても大腸の炎症に重要な役割を果たすこ
とが示された。
さらに、Malt1 プロテアーゼ活性がどのようにして大腸炎の発症に寄与するか、その
作用機序を解析するため、大腸炎モデルにおけるSCIDマウスの腸間膜リンパ節の表 現型を調べた。Figure 11に示したように、Malt1 PDマウス由来のnaïve T細胞を移入 したマウスの腸間膜リンパ節では、WT マウス由来の細胞を移入した群と比較して Th17 細胞の割合が有意に低かった。一方、Th1 細胞の割合は変化がなかった。加え て、大腸での IL-17A の発現量も WT と比較して顕著に低かったが、IFN-γ の発現量 の低下は僅かであった (Figure 12)。これらの結果から、Malt1 のプロテアーゼ活性の 欠損は、大腸における Th17 細胞の分化と活性化に影響を及ぼし、結果として大腸の 炎症反応を低下させることが示唆された。Jaworskiらの研究では、腸間膜リンパ節での Th1 細胞数は低下することが観察されているが [50]、Th1とTh17細胞の割合につい ては報告されていなかった。本研究で見られた所見は、大腸炎の発症時におけるTh1 とTh17 細胞のバランスの制御に Malt1のプロテアーゼ活性が主要な役割を果たすこ とを示唆するものである。Malt1の基質の一つである Regnase-1や Roquinは Th17細 胞への分化を抑制する因子として知られているため [30]、これらの分子が Malt1 のプ ロテアーゼ活性によって分解されることで、Th17細胞への分化が亢進すると考える。
なお、Th17 細胞は IL-17 産生性細胞として定義されるが、IL-17A そのものは大腸
上皮の炎症に対して保護的に働くため、Figure 12で見られたようなIL-17A発現抑制 が大腸炎の発症に多大に関与する可能性は低いと考える [51]。Th17 細胞は、IL- 17A だけでなく、IL-17A の別のサブタイプである IL-17F や granulocyte macrophage colony-stimulating factor (GM-CSF) を産生することが知られている [52, 53]。このこと
から、Malt1 プロテアーゼ活性は IL-17A の産生誘導のみならず Th17 細胞の分化と
活性化を制御し、これらサイトカインの産生を複合的に誘導することで、大腸の炎症反
34
応を亢進させていると考える。
興味深いことに、IFN-γとIL-17をいずれも産生するTh1/17細胞の割合もMalt1 PD マウス由来T細胞移入群では減少していた (Figure 11)。Th1/17細胞はTh17細胞か ら分化すると考えられていることから [41]、Malt1 のプロテアーゼ活性は Th1/17 細胞 の分化に間接的に影響を与えている可能性がある。また Figure 12B で示した、Malt1 PDマウス由来のT細胞を移入したマウスの大腸で認められたIFN-γの遺伝子発現量 の僅かな低下は、Th1/17細胞の減少によるものであると考察する。Th1/17細胞はIBD の病因に関与すると報告されていることから、Malt1 プロテアーゼ活性の抑制による Th17細胞とTh1/17細胞の減少は、IBD患者にとって有益であると考える [40, 41]。
本研究では、Malt1のプロテアーゼ活性が大腸で Th17 と Th1/17 細胞の活性化を 特異的に制御し、大腸炎の発症に重要な役割を果たすことを見出した。興味深いこと に、本研究で用いたnaïve CD4+CD62L+ T細胞移入大腸炎モデルは、IBDの治療薬
である抗 TNF-α 抗体に対して抵抗性をもつ動物モデルであるため [48]、Malt1 のプ
ロテアーゼ活性阻害剤は、抗 TNF-α 抗体に抵抗性のある IBD 患者においても有効 性が期待できると考える。
35
第3章 Fcγ受容体シグナルを介した炎症性疾患の病態におけるMalt1 プロテアーゼ活性の関与
第1節. 緒言
第1章で述べたように、IgG自己抗体は、全身性エリテマトーデス (SLE) [54] や 免疫性血小板減少性紫斑病 (ITP) [55] といった自己免疫疾患の発症に重要な役 割を果たす。自己抗体は、抗原と結合した後、IgGのFc鎖が樹状細胞、マクロファー ジといった自然免疫細胞上のFcγ受容体 (FcγR) に結合し、そのシグナルを活性化 させる。FcγRシグナルの活性化は貪食作用、サイトカイン産生や組織傷害に加えて、
免疫複合体の排除を含む様々な免疫反応を誘導するため、FcγRシグナルは自己抗 体依存的な自己免疫疾患の発症に重要な役割を果たすと考えられている [56]。例え
ば、FcR γ-chainノックアウトマウスでは、FcRシグナルが不活化しているため、関節
炎、腎炎、そして血小板減少症といった様々な自己免疫疾患の発症が抑制される [57, 58]。また、ヒト可溶性FcγR IIBであるvalziflocept (SH652とも呼ばれる) は、自 己抗体のFc鎖をブロッキングして、自己抗体が自然免疫細胞上のFcγRに結合する のを防ぐことで、FcγRシグナルの活性化を抑制する。本薬剤はSLEやITPの患者を 対象とした臨床試験において有効性を示している [59-61]。これらの結果は、FcγRシ グナル経路が、IgG自己抗体依存的な自己免疫疾患の治療における魅力的なターゲ ットであることを示している。
粘膜関連リンパ組織リンパ腫転座1 (Malt1) は、T細胞、B細胞、並びに自然免疫 細胞でシステインプロテアーゼとして働くことは第1章、並びに第2章で述べた。T細 胞と異なり、B細胞や自然免疫細胞においては、Malt1はPKC-θではなくSykを介し て活性化される (Figure 3) [18]。このシグナル経路が活性化されると、Malt1はCBM 複合体としての機能と、システインプロテアーゼとしての機能を通じて細胞を活性化さ
36
せる [62]。Malt1のプロテアーゼ活性が生体内でどの程度炎症反応に関与するか調
べるため、Malt1プロテアーゼ活性欠損マウス (Malt1 PDマウス) が様々な研究者に よって構築された。Malt1 PDマウスを用いた他のチームの研究から、Malt1プロテア ーゼはT細胞、B細胞、樹状細胞において、それぞれTCR、BCR、dectin受容体の シグナル伝達に重要な役割を果たすことが示された [50, 63, 64]。Malt1プロテアーゼ 活性の欠損は、実験的自己免疫性脳脊髄炎 (EAE) の発症を抑制することも明らかと なった。しかしながら、自然免疫細胞におけるFcγRシグナルへのMalt1のプロテアー ゼ活性の関与は不明であった。本章では、Malt1のプロテアーゼ活性と、FcγRシグナ ルを通じた自然免疫細胞の活性化の関係をMalt1 PDマウスを用いて明らかにするこ とを目的とした。まず、Malt1 PDマウス由来の骨髄由来樹状細胞 (bone marrow- derived dendritic cells; BMDC) をFcγR刺激したときの炎症性サイトカインの産生量を WTマウス由来の細胞と比較した。さらに、in vivoのITPモデルにおいて、FcγRを介 した自然免疫細胞の活性化に対する Malt1のプロテアーゼ活性の役割を調べた。こ れらの研究の結果、Malt1のプロテアーゼ活性が、FcγR刺激による自然免疫細胞の 活性化に重要な役割を果たし、ITPといった自己抗体を介した自己免疫疾患の病態 生理に深く関与することを明らかにした。
第2節 材料および方法 第1項 動物
第2章で述べたように、武田薬品工業株式会社において構築されたMalt1 PDマウ スとWTマウスを実験に用いた。各マウスはSPF環境下で維持され、6 – 10週齢で実 験に用いた。全ての動物実験は人道的ケアの基準に基づいて実施し、武田薬品工業 株式会社におけるIACUCの承認を得た上で実施した (実験番号: 4516, 10180)。
37
第2項 骨髄由来樹状細胞 (BMDC) の分化
まず、Malt1 PDマウスとWTマウスの両足から、大腿骨と脛骨を摘出し、それぞれ
の骨にRPMI-1640培地を注入して骨髄を採取した。骨髄をセルストレーナー上で破
砕して単細胞画分を採取した。本細胞に塩化アンモニウム溶液 (免疫生物研究所、
日本) を加えて溶血処理した。得られた骨髄由来の細胞を、100 units/mLのペニシリ ンと、100 μg/mLのストレプトマイシン、並びに10%FBSを含むRPMI-1640培地に再 懸濁した。終濃度20 ng/mLのマウスgranulocyte macrophage colony-stimulating factor (GM-CSF) (Peprotech、アメリカ) を加えて、細胞培養100-mmディッシュに播種 し、37℃で培養した。3日後に培地を交換し、さらに3日間培養を継続した。6日目に 接着細胞を2 mMのEDTAを含むPBSでピペッティングして回収し、骨髄由来樹状 細胞 (BMDC) として以下の実験に用いた。FITC標識抗CD11c抗体 (BD
Biosciences、アメリカ) を用いて染色し、BD Accuri C6 Flow Cytometer (BD
Biosciences、アメリカ) でフローサイトメトリー解析を実施して、得られたBMDCの
CD11c陽性率を調べた。
第3項 BMDCに対する FcγR刺激
第2項で得られたBMDCを30 μg/mLの抗CD16/CD32抗体 (クローン; 2.4G2) (Bio X Cell, USA) を予めコートした6-wellプレートに播種し、24時間37℃で培養し た。上清中のTNF-α、IL-6、IL-12p40、並びにIL-23の濃度をELISAで調べた。全て R&D Systems (アメリカ) 社製のELISAキットを用いて、キットに付属のプロトコルに従 って実施した。
38
第4項 FcγR刺激によるin vivoサイトカイン発現
FcγRの活性化を通じたin vivoでのサイトカインの発現量を確認するため、Malt1 PD マウスと WT マウスに 1 匹あたり 3 μg の抗 CD41 (gpIIb/IIIa) 抗体 (クローン; MWReg30) (BD Biosciences、アメリカ) を腹腔内投与した。3時間後、頚椎脱臼により 安楽死処置し、脾臓を摘出した後RNAlater (Qiagen、ドイツ) 中にて4℃で保存した。
脾臓のRNAを、RNeasy Mini Kit (Qiagen、ドイツ) とDNaseI (Qiagen、ドイツ) を用い て、付属のプロトコルに従って抽出した。NanoDrop (Thermo Fisher Scientific、アメリカ) で RNA 濃度を測定した後、High Capacity cDNA Reverse Transcription Kit (Life
Technologies、アメリカ) を用いて逆転写反応を実施し、cDNA を合成した。続いて、
TaqMan Fast Advanced Master Mix (Life Technologies、アメリカ) と TaqMan Gene Expression Assays (Life Technologies、アメリカ) の各プライマー・プローブミックスを用 い て qPCR を 実 施 し た 。qPCR 反 応 に は ViiA 7 Real-Time PCR System (Life Technologiesアメリカ) を用いた。TaqMan Gene Expression AssaysのプライマーとFAM
標識TaqManプローブのセットは、以下の各Assay IDのものを使用した。
TNF-α (遺伝子名Tnf): Mm00443258_m1 IL-6 (遺伝子名Il6): Mm00446190_m1 IL-23p19(遺伝子名Il23a) : Mm00518984_m1
内 部 標 準 遺 伝 子 は VIC/MGB 標 識 の マ ウ ス β-actin (Actb) コ ントロ ー ル (Life
Technologies、アメリカ) を用いて、⊿⊿Ct 法によりコントロール群と比較した相対発現
量を計算した。
第5項 In vivo ITPモデル
血小板減少を起こすため、抗CD41抗体、或いはアイソタイプ抗体であるラット IgG1抗体 (Bio X Cell、アメリカ) を1 μg/マウスとなるようにMalt1 PDマウスとWTマ
39
ウスに腹腔内投与した。翌日、全血を心臓穿刺により採取した。このとき、抗凝固剤と
して0.5M EDTAを加えた。血小板数、リンパ球数、並びに赤血球数を多項目自動血
球分析装置XT-1800i (シスメックス、日本) で計測した。
第6項 統計解析
データは全て平均値 ± SEM で表記した。統計解析は SAS System for Windows
(SAS Institute、アメリカ) を用いて実施した。2 群間の差は、以下のように統計処理し
た。まず分散比のF検定を実施し、等分散とみなせる場合、スチューデントのt検定を 実施した。不等分散の場合は、ウェルチのt検定を実施した。有意水準は0.05とし、p 値 (probability value) が p<0.05のとき統計学的に有意とみなした。
第3節 結果
第1項 Malt1 PDマウス由来骨髄細胞のBMDCへの分化能
まず、FcγR刺激による樹状細胞の活性化に対するMalt1プロテアーゼ活性の役割 を調べるため、それぞれの系統の骨髄由来の細胞から、GM-CSFで刺激して樹状細 胞に分化させた。Malt1 PDマウス由来の骨髄細胞が正常にBMDCに分化したことを 確認するため、マウス樹状細胞のマーカーであるCD11cの発現をフローサイトメトリー で調べたところ、CD11c陽性細胞の割合は、Malt1 PDマウス由来のBMDCとWTマ ウス由来のBMDCの間で差がなかった (Figure 13)。
40
Figure 13. The population of CD11c+ dendritic cells (DC) differentiated from Malt1 protease-deficient (PD) mouse and wild-type (WT) mouse
Bone marrow-derived dendritic cells (BMDC) were differentiated using granulocyte macrophage colony-stimulating factor (GM-CSF). The frequency of CD11c+ cells was determined by flow cytometry with FITC-conjugated anti-CD11c monoclonal antibody.
第2項 BMDCに対する FcγR刺激によるサイトカイン産生
次に、Malt1 PDマウスとWTマウス由来のBMDCを抗FcγR抗体で刺激し、炎症
性サイトカインの産生量を比較した。Figure 14のA – Cに示すように、Malt1 PDマウ ス由来のBMDCから産生されるTNF-α、IL-6とIL-12p40の濃度は、WTマウス由来 のBMDCと比較して、有意に低かった。また、Malt1 PDマウス由来の細胞から産生さ れるIL-23の産生量は低下傾向が認められた (Figure 14 D)。
![Figure 1. Differentiation of helper T cells [2]](https://thumb-ap.123doks.com/thumbv2/123deta/5820209.1034464/7.892.129.770.475.795/figure-differentiation-helper-t-cells.webp)
![Figure 2. Pathophysiology of autoantibody-dependent immune reaction [1]](https://thumb-ap.123doks.com/thumbv2/123deta/5820209.1034464/8.892.134.769.478.679/figure-pathophysiology-autoantibody-dependent-immune-reaction.webp)
![Figure 4. Structure of Malt1 [26]](https://thumb-ap.123doks.com/thumbv2/123deta/5820209.1034464/11.892.177.713.164.285/figure-structure-of-malt.webp)