日本サンゴ礁学会誌 第 17 巻,1-17(2015)
四国西南海域の高緯度生息地におけるハナヤサイサンゴ科
2 種の幼生放出期および日周幼生放出パターン
宮本麻衣
1*・横地洋之
2 1国立研究開発法人水産総合研究センター国際水産資源研究所 〒236-8684 神奈川県横浜市金沢区福浦 2-12-4 2東海大学海洋学部水産学科生物生産学専攻 〒424-8610 静岡県静岡市清水区折戸 3-20-1 要旨 高緯度生息地における幼生保育型イシサンゴ類の幼生放出の特徴と群集維持戦略を明らかにするため,ハナヤサイサンゴ Pocillopora damicornis とショウガサンゴ Stylophora pistillata について,高知県 大月町西泊において 2006 年から 2007 年の 2 年間,飼育下における幼生放出観察と自然群体の解剖観察か ら幼生放出の時期と回数を調査した。また,野外における 2 種の親群体および新規加入群体の分布調査か ら幼生の分散を調査した。ハナヤサイサンゴの幼生放出は 1 年に 2 回,6 月と 7 月のそれぞれの月で 15 日間前後行われることが確認された。新月付近に幼生放出のピークがあり,月齢への同調性が明らかと なった。幼生放出は終日観察され,放出数を昼夜で比較すると 2006 年は夜間が有意に多かったが,2007 年は有意な差は見られなかった。ショウガサンゴでは,6 月下旬から 8 月の約 2ヵ月半の間に,25 日間程 度の幼生放出を 2 回行うことが確認された。上弦から新月に幼生放出のピークがあり,月齢への緩やかな 同調性が示唆された。幼生放出は終日観察されたが,放出数は 2006 年,2007 年ともに夜間の方が有意に 多かった。2 種ともに,周年にわたって幼生放出が行われる低緯度サンゴ礁域に比べて年間の幼生放出回 数は少ないが,1 回の幼生放出期の日数は長い傾向が見られた。分布調査においては,ハナヤサイサンゴ およびショウガサンゴの新規加入群体は親群体近傍で有意に多かった。このような幼生放出パターンと幼 生の分散は,繁殖可能な季節が短い高緯度海域において,幼生の放出回数をできるだけ増やし,放出され た幼生は親群体近傍に加入することで群集を維持しようとする 2 種の繁殖戦略を反映するものと考えられ る。 キーワード 幼生放出,高緯度生息地,ハナヤサイサンゴ,ショウガサンゴ,月齢同調性
はじめに
世界的に知られるサンゴ礁海域であるグレートバ リアリーフでは,1980 年代前半より造礁サンゴ類 の 産 卵 に つ い て の 研 究 が 盛 ん に 行 わ れ て い る (Babcock et al. 1986; Harrison et al. 1984; Willis et al. 1985)。日本国内においても 1980 年代後半から, サンゴ礁海域および非サンゴ礁海域において造礁サ ンゴ類の産卵に関する多くの研究がなされてきた (Heyward et al. 1987; Hayashibara et al. 1993;Woesik 1995; Yeemin 1991; Nozawa et al. 2006; 目 崎ら 2007)。造礁サンゴ類の産卵生態情報は,その 海域における新規加入や定着量を予測する基礎資料 となり,サンゴ礁の保全に向けて群集動態を理解す る上で欠かせない知見である(目崎ら 2007)。 サンゴ礁域から離れた高緯度に位置する非サンゴ 礁海域における造礁サンゴ類の産卵生態研究とし て,わが国では和歌山県串本(33°29′N, 135°45′E) での長期にわたる観察により計 21 種の産卵が確認さ れている(御前 1989, 1990, 1995-2001, 2003-2006)。 熊本県天草(32°10′N, 130°02′E)では水槽観察に よって計 22 種の産卵が観察され(Yeemin 1991), その後の野外観察で新たに 7 種の産卵が報告された (Nozawa et al. 2006)。本研究を行った四国西南海 域では,高知県大月町尻貝(van Woesik 1995)と 西泊地先(目崎ら 2007; Nozawa 2012)の造礁サン ゴ類の産卵パターンについて報告がある。これらの 研究はいずれも放卵放精型の造礁サンゴ類を対象と * 連絡著者 E-mail: [email protected] 担当編集者 : 波利井佐紀(生態分野) 原著論文
するものであり,分散能力が限られる幼生放出型造 礁サンゴ類の幼生放出パターンは,放卵放精型の種 とは異なる特徴をもつことが予想されるが,幼生放 出型の種を扱った研究は少ない(Harii et al. 2001, Thamrin et al. 2001, Tioho et al. 2001)。 幼生放出型の造礁サンゴ類の幼生放出パターンに ついては,ハナヤサイサンゴ Pocillopora damicor-nis の月周期への同調性に関する研究がグレートバ リアリーフ(Harriott 1983, Tanner 1996),ハワイ (Richmond and Jokiel 1984, Jokiel 1985),台湾 (Fan et al. 2002),フィリピン(Villanueva et al. 2008),ジャワ島(Suharsono et al. 2008),パラオ (Atoda 1947a)などサンゴ礁海域の各地から報告 されており,その内の Tanner(1996)では同時に ショウガサンゴ Stylophora pistillata とトゲサンゴ Seriatopora hystrix についても同様の研究が行われ て い る。 台 湾 か ら は ト ゲ サ ン ゴ と ハ ナ サ ン ゴ Euphyllia glabrescens,フィリピンからはトゲサン ゴ,フトトゲサンゴ S. aliendrum,S. guttatus,イ ボハダハナヤサイサンゴ P. verrucosa などハナヤ サイサンゴ以外の幼生放出型サンゴ類の幼生放出の 日周期に関する研究も報告されている。また,沖縄 からはハナヤサイサンゴの配偶子形成と幼生発生の タイミングについての組織学的研究(Permata et al. 2000)や幼生の大きさと寿命の関係に関する研究が 報告されている(Isomura and Nishihira 2001)。 ショウガサンゴでは,パラオ(Atoda 1947b),沖 縄(山里 1991),エイラート(Rinkevich and Loya 1979a, b)から,月周期と生殖期間の関係について 報告されている。しかし,高緯度の非サンゴ礁域に おけるハナヤサイサンゴ科の生殖パターンに関して は,天草のハナヤサイサンゴについて Tioho et al.(2001)が報告しているのみに過ぎない。また近 年では,ハナヤサイサンゴの種分類やハナヤサイサ ンゴを含む同属サンゴ類の産卵生態について再検討 され,従来幼生放出型とみなされてきたハナヤサイ サンゴや P. acuta,P. meandrina でも生息する海 域によって,放卵放精型または両方の産卵パターン をもつ種があることが報告されており(Schmidt-Roach et al. 2012, 2013, 2014a, b),それぞれの生息 地における産卵生態の詳細な観察も必要である。そ こで本研究では,四国南西海域において幼生放出型 とされているハナヤサイサンゴとショウガサンゴを 対象として,飼育下における幼生放出観察と自然条 件下の群体の解剖観察を行った。また,野外におい てハナヤサイサンゴとショウガサンゴの親群体およ び新規加入群体の分布調査を行った。得られた結果 から,幼生放出期の長さ,頻度,群体間の同調性と 環境要因との関係,幼生の分散について検討した。 その後,他海域における幼生放出期間や環境要因と の関係に関する既往の知見と比較することによっ て,高緯度海域におけるハナヤサイサンゴ科 2 種の 幼生放出の特徴と群集維持における戦略を考察した。
材料と方法
調査地 飼育群体と解剖観察用の群体片の採集は高知県大 月町西泊(32゜46.6′N,132°43.9′E 以下,西泊)で 行った(図 1)。採集地点はハナヤサイサンゴとショ ウガサンゴが岩盤上に群生する場所である。飼育観 察用の群体と組織観察用の群体片は,スキューバダ イビングで水深 2~5 m の岩礁から採集した。水温 は調査地に黒潮生物研究所が設置した水温ロガーの 計 測 値 を 使 用 し た。 水 温 ロ ガ ー は 海 岸 か ら 約 100 m,水深約 6 m の海底に敷設された取水口付近 に設置した。なお水温ロガーは 1 時間毎の水温を記 録するように設定した。分布調査は西泊に加えて高 知県土佐清水市竜串(32°46′N, 132°43′E 以下,竜 串)で行った(図 1)。分布調査を行った西泊の地 点は前述の採集場所とは航路を挟んで約 300 m 離 れた場所であり,竜串の地点は横に広がる岩礁上に ハナヤサイサンゴとショウガサンゴが点在する場所 である。 飼育観察 ハナヤサイサンゴでは,2006 年は直径 8~13.5 cm の 10 群 体 を 飼 育 観 察 に 用 い た。Tioho et al. (2001)による 1997 年から 1998 年の幼生放出観察 によると,西泊とほぼ同緯度に位置する天草での幼 生放出は 2 年続けて 7 月の満月に同調して起こり, 1997 年に確認した放出日数は 9 日間であったこと が報告されている。このことから本調査地でも 7 月 の満月前後に幼生放出が起こると予測し,約 1ヵ月 前の 2006 年 6 月 14 日(月齢 17)から 10 群体(直 径 8~13.5 cm)の 飼育観察を始めた。2007 年は, 2006 年に幼生放出が始まった 6 月 23 日の月齢 26ハナヤサイサンゴ科 2 種の幼生放出期 3 と一致する 6 月 12 日に幼生放出が始まると予測し, 2006 年とは異なる 5 群体(直径 6~11 cm)を 6 月 1 日(月齢 15)に採集し,飼育観察を開始した。 ショウガサンゴでは,2006 年 6 月 14 日から 9 群 体(直径 10~21 cm)を採集して飼育観察を始めた。 2007 年は,2006 年に幼生放出が始まった 6 月 26 日 の月齢 0(新月)と一致する 6 月 15 日から幼生放 出が始まると予測し,6 月 1 日に採集し 7 月 31 日 まで(以下,1 回目)と 8 月 1 日に採集し 9 月 10 日まで(以下,2 回目)にそれぞれ異なる 5 群体(1 回目 8~13 cm,2 回目 9.5~18 cm)について飼育観 察を行った。 採集した群体は個別の容器に入れて飼育した。ハ ナヤサイサンゴは高さ 13 cm,底面 22×25 cm の四 角いプラスチックバケツ,ショウガサンゴは高さ 20 cm,底面の直径 19 cm,開口部の直径 28 cm の 円形のプラスチックバケツに,L 字の塩ビ管で排水 口を取り付けたものを使用した。排水口の先には高 さ 10 cm 直径 5 cm の円筒形のプラスチック容器を 幼生採捕器として取り付けた。幼生採捕器は側面に 4 cm 四方の窓を対面 2 か所にあけ,目合 100 µm の プランクトンネットを張り付けた。飼育水槽は調査 地の海岸から約 100 m 沖合に敷設された取水口か らの汲み上げ自然海水による流水式とし,バルブで 流量 800~1000 ml/分に調節した。注水管には 50 µm のフィルターを取り付け,海水と一緒に外部から幼 生が混入することを防ぐとともに,細かい砂などの 堆積物を除去した。水槽は太陽光のあたるサンルー ムに設置し,遮光ネットで周りを囲い日射による水 温上昇を防いだ。 観察開始後は 1 日に 1 回,朝 6 時に幼生放出の有 無を確認した。最初の幼生放出を確認した後は,1 日 2 回,6 時と 18 時に幼生採捕器に捕えられた幼 生を計数した。幼生採捕器はそれぞれの種について 20 個ずつ用意し,取り替えながら設置して幼生の 混入や取り逃しを防いだ。採捕した幼生を計測した 後,洗浄し幼生が残っていないことを確認してから 再度使用した。また 6 時から 18 時を日中,18 時か ら 6 時を夜間とし,放出された幼生の数を比較し た。幼生放出が連続して 10 日以上見られなくなっ た場合に幼生放出は終了したと判断し,観察を終了 した。 解剖観察 調査地のハナヤサイサンゴとショウガサンゴ各 10 群体にタグをつけて識別し,毎回各群体から枝 先 3~4 cm の群体片を 2~3 個採取した。採取は, ショウガサンゴでは 2006 年 6 月 9 日から,ハナヤ サイサンゴは 2006 年 6 月 19 日から開始し,両種と も 2007 年 9 月 12 日まで行った。採取間隔は基本的 に月 1 回としたが,水槽で飼育観察中の群体から幼 生が放出されている期間は,2006 年は月齢周期に 合わせて週 1 回,2007 年は 2 週間に 1 回とした。 2006 年に採取対象としていた群体を 2007 年にも用 いたが,これ以上の群体片採取が困難と判断された ハナヤサイサンゴ 3 群体は新たな群体と交換した。 ショウガサンゴでは 2007 年 7 月 17 日のサンプリン グ前と 7 月末の台風後にそれぞれ 1 群体が流失した ので,その後は 8 群体を調査対象とした。 採取した各群体片は, 10% 中性緩衝ホルマリンで 24 時間以上固定した後,25% 蟻酸と 10% アミノ酸 を 1 対 1 の割合で混合した液に 24 時間以上浸漬し て脱灰したものを,10% 中性緩衝ホルマリンで保 存した。そして,軟体部をポリプの裏側から双眼実 体顕微鏡で観察し,採集した小片の全ポリプ数,お よびその中で幼生を保有しているポリプと卵母細胞 図 1 高知県大月町西泊および土佐清水市竜串の 調査地点
Fig. 1 Map of the study site at Nishidomari, Otsukiand and Tatsukushi, Tosashimizu in south-west Kochi, Japan.
を保有しているポリプの数をそれぞれ計数した。調 査対象とした群体の中で幼生を保有していた群体と 卵母細胞を保有していた群体の割合を求めた。幼生 を保有していた群体片の全観察ポリプに対する幼生 保有ポリプと卵母細胞保有ポリプの各割合を群体ご とに求め,観察日ごとに平均を算出した。 解剖観察の参考として,顕微鏡下で計数するもの が幼生または卵母細胞であることを確認するために 2 種それぞれの 2006 年 6 月から 2007 年 5 月の各月 1 群体の群体片について組織切片を作成し,観察し た。軟体部をブアンホーランド液で再固定した後, 水洗し,50% から 100% のエタノールに順次移し入 れて脱水した。脱水後,定法によりパラフィン包埋 し,厚さ 6 µm の連続切片を作成して,ヘマトキシ レン・エオジン染色を施した。作成した標本を光学 顕微鏡で観察し,Harriott(1983)にならい幼生と 卵母細胞を確認した後,解剖した群体片と照合した。 分布調査 2006 年 10 月に西泊と竜串の 2 調査点において, 任意に設置した 5×5 m の調査区を,400 の小区画 (0.25×0.25 m)に分けて調査した。調査区内のハナ ヤサイサンゴとショウガサンゴの親群体の種類と群 体数を記録した。同時に幼生加入に由来すると思わ れる小型(1 cm 以上 5 cm 以下)の新規加入群体の 有無と群体数を小区画ごとに種類別に記録した。小 区画内の底質を記録し,幼生の着底が可能と思われ る岩盤の基底をもつ小区画を有効区画とした。西泊 と竜串の調査区について,有効区画数と新規加入群 体が観察された区画数を種別に比較した。 統計処理 飼育観察を行った群体の群体サイズと総幼生放出 数の直線相関の有意性は,線形回帰分析を行った 後,F 検定を用いて評価した。飼育観察を行った群 体から放出された幼生数の日中と夜間での比較は, Mann-Whitney U 検定を用い,分布調査における西 泊と竜串の 2 調査点での有効区画中の新規加入群体 が観察された区画数の比較には,Fisher の正確確 率検定を用いた。すべて有意水準は 5% とした。
結果
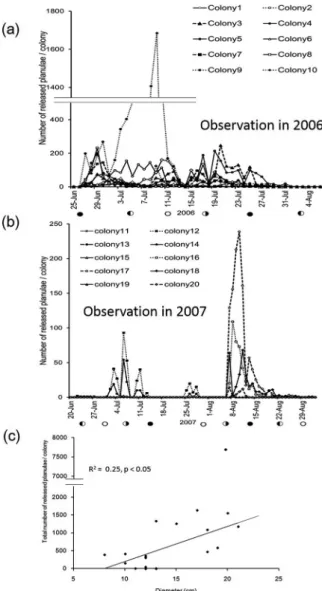
ハナヤサイサンゴの飼育観察 2006 年の幼生放出期間の野外日平均水温は 23.2 ~25.4℃で,幼生放出数のピークでは 25.1℃であっ た。2007 年は,2 回の幼生放出期間の日平均水温は それぞれ 24.2~25.2℃,22.3~24.7℃でありピーク時 はそれぞれ 24.6℃と 22.8℃であった(表 1,図 2)。 2006 年は 6 月 24 日(月齢 27)~7 月 9 日(月齢 13)の 16 日間に幼生放出が観察された(図 3a)。6 月 24 日には 10 群体すべてが幼生を放出した。幼生 放出日数は最も長い群体で連続した 16 日間であり, 最も短い群体で 11 日間であった。2006 年に飼育下 で観察された幼生放出はこの 1 回のみであり,10 群体の幼生放出日数の平均は 12.5 日間(SE=0.8) であった。幼生放出のピークは 6 月 24~26 日に認 められ,24 日に 1 群体(図 3a,△),25 日に 8 群体, 26 日に 1 群体(図 3a,□)がそれぞれ調査期間中 の 1 日あたり最多幼生放出数を記録した。幼生の総 放出数は,最も多い群体では 4318 で,その群体サ イズは直径 11 cm と 10 群体中 3 番目に小さな群体 であった。最も少ない群体は直径 12 cm の群体で総 幼生放出数は 302 であり,10 群体の総幼生放出数 の平均は 1128(SE=385.9)であった。 2007 年は,観察期間中に 2 回の幼生放出期が観 察された。1 回目は 6 月 12(月齢 26)~21 日(月 齢 6) の 10 日 間,2 回 目 は 7 月 5( 月 齢 20) ~19 日(月齢 4)の 15 日間で,それぞれ 2 群体が幼生 放出を行った(図 3b)。1 回目の幼生放出日数は 2 群体とも 10 日間であったが(図 3b,□▲),2 回 図 2 西泊における観測水温の変化。 Fig. 2 Fluctuation of daily water temperature at Nishidomari from June 2006 to August 2007.ハナヤサイサンゴ科 2 種の幼生放出期 5 目は 15 日間と 8 日間であった(図 3b,▲○)。4 群体の 1 回あたりの幼生放出日数は平均 10.8 日間 (SE=1.5)であった。幼生放出数のピークは,1 回 目は 2 群体とも 6 月 18 日に観察されたが,2 回目 は 7 月 17 日に 1 群体でのみ顕著なピークが見られ た(図 3b)。幼生の総放出数は,1 回目の最も多い 群体で 1153 であり,その群体サイズは直径 7 cm で 5 群体中 2 番目に小さな群体であった(図 2b,□)。 最も少ない群体では 104 で直径 10 cm の 2 番目に 大きな群体であった。2 回目に顕著なピークを示し た群体は 1 回目にも幼生放出を行った群体であり, 2 回目の総放出数は 2200 であった(図 3b,▲)。 幼生放出を行ったのべ 4 群体での総幼生放出数の平 均は 714(SE=427.3)であった。2006 年と 2007 年 に観察した 15 群体について,ハナヤサイサンゴに おいては群体サイズ(直径)と総幼生放出量の間に 有意な相関は認められなかった(図 3c,R2=0.066 p>0.05,F 検定)。 2006 年と 2007 年ともに幼生放出の時間帯は特に 限定しておらず,日中と夜間のどちらでも幼生放出 が確認されたが,2006 年は 18 時~6 時の夜間の方 が 6 時~18 時の日中に比べて有意に多かった(図 4)。2007 年は日中の方が夜間に比べて幼生放出数 が多い傾向が見られたが,有意な差ではなかった。 また,2006 年に 1 回,2007 年に 2 回観察された幼 生放出のピークは,ともに新月付近であった。 ハナヤサイサンゴの解剖観察 解剖観察では,幼生は胃腔内で観察され,二重膜 図 3 飼育観察したハナヤサイサンゴの群体別幼 生放出数(a. b)および群体サイズによる幼生放 出数(c)。2006 年は 10 群体(a),2007 年は 5 群 体(b)を飼育観察した。各記号は個々の群体を 表す。 Fig. 3 Daily number of released planulae from P.damicornis colonies kept in tanks, ten colonies in 2006 (a) and five colonies in 2007 (b) were observed. Each symbol indicates individual colony. (c) Rerationship between colony diameter and total number of planulae released (R2=0.066, p>0.05) 図 4 日中と夜間のハナヤサイサンゴの平均幼生 放出数の比較。バーは標準誤差を示す。 Fig. 4 Comparison of the average number of released planulae from P. damicornis during the day and night. Vertical bars demonstrate standard errors of mean. * p<0.05, Mann-Whitney U test
構造,隔膜構造および褐虫藻が確認された。卵母細 胞は隔膜内で観察され,幼生で確認されたような二 重膜構造などは見られなかった。 2006 年には, 6 月 19 日~7 月 18 日に採取した 5 回分の群体片の解剖観察でそれぞれ 5~10 群体で幼 生の保有を確認した(図 5a)。幼生を保有していた ポリプの割合は,6 月 19 日の 7 群体で平均 8.9% で あったが,26 日には 10 群体で平均 7.2%,7 月 4 日 には 5 群体で平均 2.0% へと一旦減少したが,11 日 には 5 群体で平均 12.7% へと増加に転じ,18 日は 9 群体で平均 11.2% となった(図 5b)。6 月 19 日以 前は採集を行っていないため幼生の保有は不明であ り,7 月 18 日~8 月 20 日は海況が悪かったため群 体片の採取ができなかった。8 月 20 日以降に採集 した群体片では幼生は確認されなかった。 2007 年は 6 月 2 日~7 月 17 日に採取した 4 回分 の群体片の解剖観察でそれぞれ 1~9 群体で幼生の 保有を確認した(図 5a)。6 月 2 日は 1 群体でのみ 幼生保有が確認され,幼生を保有していたポリプの 割合は平均 0.2% であった。15 日は 1 群体で 0.4%, 7 月 1 日は 9 群体で平均 9.0%,17 日は 6 群体で平 均 2.7% のポリプが幼生を保有していた(図 5b)。5 月 7 日以前と 8 月 6 日以降の群体片では幼生は見ら れなかった。野外における水温は,両年ともに幼生 が確認される約 1ヶ月前の 5 月初めから 20℃を上回 るようになり,幼生が確認された 6 月中旬には 20℃以上の水温で安定していた(図 2)。両年とも 1 つのポリプが保有していた幼生の数は 2~4 個で あった。 卵母細胞は 1 年を通してほとんどの群体片で保有 が確認された。卵母細胞を保有しているポリプの割 合は,幼生の保有が確認されるようになる約 3ヶ月 前から徐々に高くなり始め,およそ 1ヵ月前にピー クを迎えた後,幼生保有期の終盤に急激に低下し 0.2~15% 程度になった後,低水準で卵母細胞を保 有する傾向が見られた(図 5c)。卵母細胞の保有数 は 1 ポリプにつき 2~4 個であった。組織切片観察 では,幼生は隔膜内および胃腔内で観察された(図 6)。ポリプ内の幼生には隔膜構造が認められ,共生 藻を保有していた。 ショウガサンゴの飼育観察 2006 年の幼生放出期間の日平均水温は 23.1~ 28.7℃で,幼生放出数のピーク時は 26.8℃であった。 2007 年の幼生放出期間の日平均水温は 22.9~25.4℃, 24.8~29.4℃であり,ピーク時はそれぞれ 24.3℃, 26.7℃であった(表 1,図 2)。また,観察期間中の 図 5 解剖観察におけるハナヤサイサンゴの幼生 および卵母細胞を保有していた群体の割合(a) と幼生が観察されたポリプの割合(b)および卵 母細胞が観察されたポリプの割合(c)。(b)およ び(c)の図の縦線は標準偏差を示す。卵母細胞 と幼生の項目には両方を保有している群体または ポリプが含まれ,幼生を保有している群体はすべ て卵母細胞を保有していた。 Fig. 5 Percentage of colonies that held oocytes or planulae (a), percentage of polyps that held planulae (b) and percentage of polyps that held oocytes (c) of P. damicornis counted oocytes and planulae. Vertical bars in (b) and (c) indicate standard errors of mean. The colonies or polyps holding both oocytes and planulae are separately counted as those with oocytes and as those with planulae.
ハナヤサイサンゴ科 2 種の幼生放出期 7 幼生放出数のピークは両年ともに,幼生放出開始時 から水温が 1℃以上,上昇した日の前後で観察され た。 2006 年は 6 月 27 日(月齢 1)~8 月 2 日(月齢 7) までの 37 日間に幼生放出が観察された(図 7a)。6 月 27 日には 10 群体すべてが幼生放出を開始し,ほ とんどの群体は 7 月 26(月齢 0)~30 日(月齢 4) に放出を終了した。この間,全ての群体が毎日幼生 を放出し,放出期間は最も短い群体で 7 月 14 日ま での 18 日間,最も長い群体で 8 月 2 日までの 37 日 間であった。幼生放出期間の平均は 29.5 日(SE= 1.9)であった。各群体における調査期間中の 1 日 あたり最多幼生放出数のピークは,6 月 28~29 日, 7 月 10 日および 7 月 19~20 日にそれぞれ 6 群体, 2 群体と 7 群体で確認され,そのうちの 3 群体は 2 度の顕著なピークを示した(図 7a,●■□)。幼生 の総放出数は,最も多い群体では 7693 で,その群 体サイズは直径 20 cm であった。最も少ない群体は 最も小さな直径 10 cm の群体で,調査期間中の最多 幼生放出数は 88 と,幼生放出数の顕著なピークは 見られなかった。総幼生放出数の 10 群体での平均 は 1737.7(SE=711.6)であった。 2007 年は,1 回目の観察では 5 群体中 2 群体で 6 月 23 日(月齢 8)~7 月 13 日(月齢 28)の 21 日 間と 7 月 25(月齢 10)~28 日(月齢 13)の 4 日間 に幼生放出が観察された(図 7b)。幼生放出日数は 1 群体が 6 月 23 日からの 21 日間と 7 月 25 日から の 4 日間,他方は 6 月 26 日からの 18 日間と 7 月 26 日からの 3 日間であった。2 回目の観察では 5 群 図 6 ハナヤサイサンゴの組織切片観察。隔膜構 造が発達途中の幼生。2006 年 6 月の標本を観察し た。P,幼生;M,隔膜;Z,褐虫藻
Fig. 6 Histological observation of P. damicornis. A planula showing development of mesenteries. The observed tissues were collected in June 2006. P, planula; M, mesentery; Z, zooxanthella 図 7 飼育観察したショウガサンゴの幼生放出数 (a, b)および群体サイズによる幼生放出数 (c)。 2006 年(a),2007 年(b)ともに 10 群体を観察 した。各記号は個々の群体を表す。 Fig. 7 Daily number of released planulae from S. pistillata colonies kept in tanks. Ten colonies were observed in 2006 (a) and 2007 (b). Each symbol indicates individual colony. (c) Reration-ship between colony diameter and total number of released planulae (R2=0.25, p<0.05).
体中 4 群体で 8 月 7 日(月齢 23)~9 月 1 日(月齢 19)までの 26 日間に幼生放出が観察された(図 7b)。幼生放出日数は最も短い群体で 8 月 12 日ま での 6 日間であり,最も長い群体では 9 月 1 日まで の 26 日間であった。幼生放出を行った 6 群体の 1 回あたりの平均幼生放出日数は 17.5 日(SE=2.8) であった。調査期間中の幼生放出数のピークは,1 回目は 2 群体共に 7 月 6 日に見られた。2 回目は幼 生放出を行った 4 群体すべてで 8 月 9~11 日に観察 された。1 回目の 2 群体の総幼生放出数の平均は 252.5(SE=104.5)であり,ピーク時の日間放出数 の平均は 73.5(SE=33.0)であった。2 回目の総幼 生放出数は,最も多い群体の群体サイズは直径 12 cm で幼生数 1090 であった。最も幼生放出数が 少ない群体は直径 9.5 cm で総放出数 42,ピーク時 でも日間放出数 12 と少なかった。4 群体での総幼 生放出数の平均は 456.8(SE=223.6)であった。 2007 年に観察された 2 回の幼生放出期の総幼生放 出数の平均は 388.7(SE=150.3)であった。2006 年と 2007 年に観察した 20 群体について,ショウガ サンゴにおいては群体サイズ(直径)が大きいほど 総幼生放出数が増加する有意な相関が認められた (図 7c,R2=0.25,p<0.05,F 検定)。 両年とも幼生放出時間帯は特に限定されておらず 日中と夜間のどちらも放出が確認されたが,夜間の 方が日中に比べて幼生放出数が有意に多かった(図 8)。また,2006 年に観察された 2 回の幼生放出数 のピークは,1 回目は新月付近に,2 回目は下弦付 近で観察された。2007 年の 2 回のピークはともに 下弦付近に観察された。 ショウガサンゴの解剖観察 ショウガサンゴでも,ハナヤサイサンゴと同様な 特徴を持つ幼生や卵母細胞が,それぞれ胃腔内と隔 膜内で観察された。2006 年は,6 月 26 日~8 月 20 日に採取した 8 回分の群体片で幼生の保有を確認し た(図 9a)。幼生を保有していたポリプの割合は, 6 月 26 日には 6 群体で平均 2.8% であったが,7 月 4 日は 8 群体で平均 4.6%,7 月 11 日は 7 群体で平 均 13.3% へと徐々に増加し,18 日は 9 群体片で平 均 8.2%,25 日は 8 群体で平均 8.0%,8 月 2 日も 8 群体で平均 5.4%,12 日は 6 群体で平均 4.0%,20 日 には 3 群体で平均 1.7% へと減少した(図 9b)。6 月 19 日以前および 8 月 25 日以降の群体片では幼生 保有は検出されなかった。 2007 年は,6 月 15 日~8 月 6 日の 4 回分の群体 片で幼生保有が確認された(図 9a)。6 月 15 日は 1 群体のみ幼生の保有が確認され,幼生を保有してい たポリプの割合は 4.0% であった。7 月 1 日は 6 群 体で平均 2.0%,7 月 17 日は 8 群体で平均 5.0%,8 月 6 日は 5 群体で平均 0.8% のポリプが幼生を保有 していた(図 9b)。野外における水温は,両年とも に幼生が確認される約 1ヶ月前の 6 月初めには 24℃ を上回る日が見られるようになり,7 月に一旦低水 温に転じるが,幼生が確認された 6 月後半には 24℃以上で安定した(図 2)。両年とも 1 つのポリ プが保有していた幼生の数は 2~4 個であった。 ショウガサンゴでもハナヤサイサンゴと同様に 1 年を通してほとんどの群体片で卵母細胞の保有が確 認された。卵母細胞の保有率は,幼生保有期間の約 1ヶ月前から急激に高まりピークを迎え,幼生保有 期の終盤にはハナヤサイサンゴ同様に 1.5~7% 程度 まで急激に低下した後,次の繁殖期までは低水準で 卵母細胞を保有する傾向が見られた(図 9c)。卵母 細胞の保有数は 1 ポリプにつき 2~4 個であった。 また,組織切片観察では,幼生を保有している群体 内において複数ポリプおよび同一ポリプ内で発達途 中の卵母細胞,精巣やさまざまな発達段階の幼生が 図 8 日中と夜間のショウガサンゴの平均幼生放 出数の比較。バーは標準誤差を示す。 Fig. 8 Comparison of the average number of released planulae from S. pistillata during the day and night. Vertical bars demonstrate standard errors of mean. * p<0.05, Mann-Whitney U test
ハナヤサイサンゴ科 2 種の幼生放出期 9 同時に観察されたことから,卵成熟と幼生の発達, 保有が並行することが確認された(図 10)。 分布調査 西泊の 5×5 m 調査区内の 400 の小区画において は,ハナヤサイサンゴとショウガサンゴの親群体は 観察されなかった。新規加入群体はショウガサンゴ が 5 小区画で 5 群体観察され,ハナヤサイサンゴは 確認されなかった(表 3)。調査区の 70% ほどは底 質が岩盤であり,その他は砂礫であった。幼生着底 に適した岩盤を含む有効小区画数は 296 であった。 竜串の調査区においては,ハナヤサイサンゴ 3 群 体,ショウガサンゴ 65 群体の親群体が観察され, 図 9 解剖観察におけるショウガサンゴの幼生お よび卵母細胞を保有していた群体の割合(a)と 幼生が観察されたポリプの割合(b)および卵母 細胞が観察されたポリプの割合(c)。(b)と(c) の縦線は標準誤差を示す。卵母細胞と幼生の項目 には両方を保有している群体またはポリプが含ま れ,幼生を保有している群体はすべて卵母細胞を 保有していた。 Fig. 9 Percentage of colonies that held oocytes and planulae (a), percentage of polyps that held planulae (b) and percentage of polyps that held oocytes (c) of S.pistillata counted oocytes and planulae. Vertical bars in (b) indicate standard errors of mean. The colonies or polyps holding both oocytes and planulae are separately counted as those with oocytes and as those with planulae. 図 10 ショウガサンゴの組織切片観察(A, B)。 隔膜が発達した幼生(A)およびポリプ内の卵母 細胞,精巣,幼生(B)。2006 年 6 月の標本を観 察した。P,幼生;O,卵母細胞;T,精巣;M, 隔膜;Z, 褐虫藻
Fig. 10 Histological observation of S. pistillata (A, B). A: A planula showing development of mesenteries. B: A planula, an oocyte and a testis. The observed tissues were collected in June 2006. P, planula; O, oocyte; T, testis; M, mesentery; Z, zooxanthella
新規加入群体はハナヤサイサンゴが 11 小区画で 11 群体,ショウガサンゴが 27 小区画で 29 群体観察さ れた(表 3)。海底のほぼ 90% が岩盤であり,有効 小区画数は 371 であった。それぞれの調査区で有効 小区画数と 2 種のサンゴの新規加入群体が観察され た小区画数を比較したところ,いずれの種も調査区 内に親群体が多かった竜串での出現小区画数が有意 に多かった(Fisher の正確確率検定,p<0.05)。
考察
ハナヤサイサンゴ 本研究のおもな結果を表 1 と表 2 にまとめた。 Schmidt-Roach et al.(2012)は,ハナヤサイサン ゴおよび P. acuta,P. meandrina などの種類では, 同一種内でも海域によって異なる産卵生態を示すこ とを報告しているが,本海域のハナヤサイサンゴで は放卵放精は観察されず,幼生放出型の産卵パター ンのみを持つことが確認された。2006 年と 2007 年 の結果から,本調査地のハナヤサイサンゴは 6~7 月に,連続した 15 日間前後の幼生放出を行うこと が確認された。2007 年の飼育観察では幼生放出量 のピークが 2 回観察されたことに加えて,2006 年 には飼育下における幼生放出が終了した後にも自然 条件下から採取した群体片で幼生保有が確認された ことから,本調査地のハナヤサイサンゴは 6 月と 7 月に 1 回ずつ,年 2 回の幼生放出期を持つものと考 えられる(表 1,2)。ほぼ同緯度に位置する非サン ゴ礁域の天草では,年 1 回,7 月に 9 日間の幼生放出 を行うことが報告されており(Tioho et al. 2001), 本海域の幼生放出期は天草よりも回数が多くやや長 期に及ぶことが明らかとなった。これは,西泊が位 置する四国西岸が,天草が位置する熊本県牛深と比 較して年平均水温が高く,ハナヤサイサンゴの幼生 放出の下限水温である 20℃以上の期間がやや長い ためと考えられる(環境省・日本サンゴ礁学会 2004)。また,解剖観察の結果から,幼生を保有し ていた群体片のポリプの幼生保有率は幼生放出期間 を通して 0.2~34.0% で推移しており(表 2),群体 内および群体間ともに幼生放出は短期間に集中する のではなく長期にわたることが明らかとなった。さ らに,1 年を通して卵母細胞の保有が観察された。 Harii et al. (2001)によると,東京湾における幼生 表 1 ハナヤサイサンゴとショウガサンゴの飼育観察結果の要約 Table 1 Summary of observation of planulation in captive P. damicornis and S. pistillataハナヤサイサンゴ科 2 種の幼生放出期 11 表 2 ハナヤサイサンゴとショウガサンゴの解剖観察結果の要約 。卵母細胞と幼生のカラムには両方を保有している群体またはポリプが含まれ ,幼生を保有 している群体はすべて卵母細胞を保有していた。 Table 2 Summary of oocytes and planulae count in P. damicornis and S. pistillata. The colonies or polyps holding both oocytes and planulae are included in the oocytes and the planulae columns. All the colon ies which brooded planulae held oocytes. * Range of percentage, ** Average of percentage (range of percentage)
保育型サンゴであるニホンアワサンゴ Alveopora japonica の組織観察では,年間を通して卵母細胞が 確認され,月平均水温が年間最高値を記録した後に 幼生放出期を迎えることが知られており,その卵形 成には 11ヶ月を要し,幼生が観察される 4ヶ月程前 から徐々に卵径が大きくなることが報告されてい る。このことから,本海域のハナヤサイサンゴも幼 生放出が終了する 7 月終わりから次の幼生放出期ま での 11ヶ月前後を,卵形成に費やすことが示唆さ れた。性成熟段階解明のための,より詳細な組織観 察は今後の課題である。 幼生放出と月齢の関係については,本調査地のハ ナヤサイサンゴの幼生放出のピークは新月付近に見 られており,幼生放出は月齢に同調することが明ら かとなった(表 1)。グレートバリアリーフ北部の リザード島では満月付近に幼生放出のピークがあり (Harriott 1983),南部のヘロン島では上弦で組織内 に幼生が見られるようになり新月に幼生放出が起こ ることが報告されている(Tanner 1996)。そのほ かに台湾では新月から上弦に幼生が放出され,ピー クは上弦付近にあることが知られている(Fan et al. 2002)。沖縄では,新月から上弦に同調して幼生 放出が起こり(山里 1991,Permara et al. 2001), 前出の天草では満月に同調して幼生放出が開始され ることが報告されている(Tioho et al. 2001)。この ように,ハナヤサイサンゴは月齢に同調した幼生放 出を行うが,海域によって放出する月齢は異なって いる。 幼生放出の時間帯に関して,本調査地のハナヤサ イサンゴでは日中および夜間のどちらでも幼生放出 が見られたのに対して,天草では早朝から昼前に放 出が限定されており(Tioho et al. 2001),沖縄では 早朝 3 時から 5 時に幼生放出が起こることが報告さ れ て い る(Isomura and Nishihira 2001)。Fan et al. (2006)によると,台湾のハナヤサイサンゴでは 1 日中幼生放出が見られるが,早朝と夕方の 2 回 ピークがあり,明暗サイクルとの関連が示唆されて いる。本研究では, 2006 年は有意に夜間の幼生放 出数が多く,2007 年には有意差は認められなかっ たが日中の方が多い傾向にあった。このことから, 本調査地のハナヤサイサンゴの幼生放出は台湾のハ ナヤサイサンゴに類似して,日に数回のピークを もっている可能性があるが,幼生放出の日周期解明 にはより詳細な調査が必要である。 潮汐との関連性では,本調査地のハナヤサイサン ゴには中潮~大潮に幼生放出のピークが見られた (表 1)。天草では早朝から昼にかけて幼生放出が見 られ,この時間帯は引潮にあたることが報告されて いる(Tioho et al. 2001)。下げ潮時に幼生放出を行 うことは,より遠くへ幼生を分散させる効果を持つ と考えられる。本調査地では幼生放出期間が比較的 長く,しかも日中および夜間のどちらも幼生を放出 していることから,特定の時間の干満を利用して幼 生を分散させる戦略は取っていないと考えられる。 水温との関係では,山里ら(2008)によってハナ ヤサイサンゴは 20℃が幼生放出の下限水温である 表 3 西泊および竜串におけるコドラート調査で観察されたハナヤサイサンゴとショウガサンゴの親群体と 加入群体の数および有効区画数とハナヤサイサンゴ科の加入群体が観察された区画数。
Table 3 Number of parental and recruitment colonies of P. damicornis and S. pistillata observed in 5 m× 5 m quadrats at Nishidomari and Tatsukushi. Number of 0.25×0.25 m grids with new recruitment colonies per total number of grids suitable for coral settlement were compared for each species between the two quadrats.
ハナヤサイサンゴ科 2 種の幼生放出期 13 ことが報告されている。本海域のハナヤサイサンゴ では,23~25℃で幼生放出の開始が観察され 22.5~ 25.4℃で終了しており,山里らの報告を支持する結 果となった。また,組織観察からも 20℃を上回る 日が見られるようになる,幼生保有期の 3ヶ月程前 から 1ヶ月前にかけて卵母細胞の保有率が増加して おり,この間に受精が行われているものと考えられ る。また,20℃以上の水温が続く 6~7 月にかけて 幼生保有率が急激に増加する傾向が見られ,本海域 でもハナヤサイサンゴの幼生放出の好適水温は 20℃以上であることが示唆された。 ショウガサンゴ 本調査地のショウガサンゴは,飼育観察と解剖観 察の結果から,6 月下旬~8 月に 25 日間前後の幼生 放出を 2 回行うことが確認された。また,幼生を保 有していた群体のポリプの幼生保有率は,幼生放出 期間を通して 0.3~36.0% で推移していた(表 2)。 このことから,ハナヤサイサンゴと同様に群体内お よび群体間ともに幼生放出が短期間に集中するので はなく比較的長期に及ぶことが示唆された。低緯度 のサンゴ礁域であるフィリピンでは,年間を通して 毎月 10 日前後の幼生放出を行うことが報告されて いる(Villanueva et al. 2008)。また沖縄では,4 月 ~6 月の 3ヶ月間に毎月 1 週間程度の幼生放出を行 うとされている(山里 1991)。これらの低緯度海域 と比較すると,本調査地では年間の幼生放出期数は 少ない代わりに,1 回の幼生放出日数が大幅に長い ことが分かる。本調査地のショウガサンゴが幼生を 放出できる季節は水温との関係から夏に限られるた め,1 回の幼生放出期間を長くすることによって, できるだけ多くの幼生を放出することができると考 えられる。さらに,解剖観察からハナヤサイサンゴ と同様に 1 年を通して卵母細胞の保有が観察され, ショウガサンゴでも幼生放出が終了する 8 月終わり から次の幼生放出期までの 10ヶ月前後を卵形成に 費やすことが示唆された。ショウガサンゴにおいて も,性成熟段階の解明にはより詳細な組織観察が必 要である。 本調査海域のショウガサンゴは下弦から新月付近 に幼生放出のピークが見られており(表 1),幼生 放出数の変動は月齢に同調する傾向が認められた。 Fan et al.(2002)によると,台湾のショウガサン ゴは下弦に同調して幼生放出数と幼生放出群体数が 最も多くなる。紅海では新月から満月に同調して幼 生放出が起こり(Rinkevich and Loya 1979b),満 月付近で幼生放出のピークを迎える(Zakai et al. 2006)。また,沖縄では満月から下弦に同調して幼生 放出が起こることが報告されている(山里 1991)。 一方,グレートバリアリーフではリザード島とヘロ ン島ともに月齢への同調性を示さない(Harriot 1983, Tanner 1996)。このように,ショウガサンゴは海 域によって幼生放出の月齢周期への同調性の有無が 異なり,月齢周期に同調した幼生放出を行う場合で も海域によって同調する月齢は異なっている。 幼生放出の時間帯については,本調査地のショウ ガサンゴは日中および夜間のどちらでも幼生を放出 するが,夜間の放出量が日中に比べて多かった。台 湾でもショウガサンゴは 1 日中幼生を放出するが, 午前 5 時から 7 時の早朝に最も多く放出することが 報告されている(Fan et al. 2002)。 本海域のショ ウガサンゴでは小潮から大潮のどの時期にも幼生放 出数のピークが起こっており(表 1),潮汐への同 調性は弱いことが確認された。台湾でも,幼生放出 を起こす刺激として重要なのは明暗サイクルである 可能性が示唆されており,潮汐への同調性は見られ ていない(Fan et al. 2006)。本研究では幼生放出 時刻に関して昼夜比較しか行わなかったが,幼生放 出の日周期解明のためにはより詳細な調査が必要で ある。 山里ら(2008)ではショウガサンゴの幼生放出の 下限水温は 24℃であることが報告されている。本 海域のショウガサンゴでも,水温 24℃以上のタイ ミングで幼生放出が始まり,23.5~27.9℃で終了し た。また,組織観察では 24℃を上回る水温で幼生 保有率が急激に増加している傾向が見られており, この報告を支持する結果となった。さらに,幼生放 出開始時から 1℃以上水温が上昇した日前後で幼生 放出数のピークが観察されたことから,水温が幼生 放出を促す刺激となっている可能性も考えられる。
まとめ
調査地におけるハナヤサイサンゴ科 2 種の幼生放 出の特徴と分布調査の結果から,本海域のハナヤサ イサンゴおよびショウガサンゴの繁殖戦略について 以下にまとめる。本研究からハナヤサイサンゴの幼生放出期は 6~7 月,ショウガサンゴは 6 月下旬~8 月であることが明らかとなった。渡邊ら(2009)が 行った西泊を含む四国西南海域 3 地点での加入量調 査によると,ハナヤサイサンゴ科の定着期は 2007 年が 6~8 月から 9~11 月まで,2008 年は 5~7 月 から 7~9 月であったことが報告されており,幼生 放出の下限水温以上になる夏季にのみ幼生放出が行 われることが確認された。繁殖可能な好適水温の期 間が短い,高緯度に位置する本海域のハナヤサイサ ンゴおよびショウガサンゴは,特定の時間に集中し て幼生を放出するよりも,下限水温以上になる限ら れた繁殖期内の幼生放出期間をできるだけ長くする ことで,多くの幼生が生残できる可能性が高くなる ことが考えられる。低緯度海域の 2 種と比較して幼 生放出期間が長い最も大きな要因としては,前述の 通り繁殖適温である高水温期が短いため,年間の幼 生放出回数が限られていることが考えられるが,そ の他の生態的要因または環境要因についてはより詳 細な研究が必要である。また,分布調査の結果から 親群体が多く生息する場所に新規加入群体が多く, Tioho et al. (2001)によるハナヤサイサンゴ科の幼 生の加入は親群体近傍ほど多いという報告を支持す る結果となった。紅海におけるショウガサンゴの幼 生の生残,着底および分散について行われた研究で は,幼生放出前期の早い時期に放出された幼生は生 残率,着底率のどちらにおいても高かったが,放出 1 日目の夜においては幼生放出期後期の幼生の着底 率が明らかに高かったことから,ショウガサンゴの 幼生の生残や着底は水温に関係していると報告され ている(Amar et al. 2007)。つまり,幼生放出前期 の水温が低いうちは比較的長く生残,分散し着底す るが,幼生放出期後期の水温が高くなる時期に放出 された幼生は,放出後すぐに着底しようとするとい う生態的な違いが見られた。本海域は高緯度海域で あり,水温の高い時期および幼生放出期間がサンゴ 礁海域に比べて非常に短いことから,常に紅海にお ける幼生放出期後期に当たることが考えられ,サン ゴ礁海域よりも親群体近傍への加入が集中するもの と考えられる。このことは,親群体の近傍に加入が 多かった分布調査の結果とも整合する。本海域のハ ナヤサイサンゴ科の繁殖戦略について,幼生放出の 時間帯調査などより詳細な研究が望まれる。
謝辞
施設利用許可及び研究・調査に際し,数々の指導 と助言を頂いた黒潮生物研究財団黒潮生物研究所の 職員諸氏に厚く御礼申し上げる。東海大学海洋科学 博物館学芸課職員の小林弘治氏には組織切片を作成 する際に丁寧なご指導を頂いた。また,渡邊美穂氏 をはじめとする学友諸兄には調査・観察に協力して 頂いた。国立研究開発法人水産総合研究センター国 際水産資源研究所外洋生態系グループの清田雅史博 士および林原 毅博士には論文のとりまとめに際し て有益なアドバイスを頂いた。また本誌 2 名の査読 者および編集委員の方々から頂いたコメントによ り,原稿の改善を図ることができた。重ねてお礼申 し上げる。 なお,この研究は,財団法人黒潮生物研究財団 (現 公益財団法人黒潮生物研究所)の 2006 年度お よび 2007 年度の助成金を受けて行われたものであ る。記して感謝します。引用文献
Amar OK, Chadwick NE, Rinkevich B (2007) Coral planulae as dispersion vehicles: biological prop-erties of larvae released early and late in the season. Mar Ecol Prog Ser 350: 71-78 Atoda K (1947a) The larva and post-larval develop-ment of some reef-building coral, Ⅰ. Pocillopora damicornis cespitosa (Dana). Sci Rep Tohoku Univ 18: 24-47Atoda K (1947b) The larva and post-larval devel-opment of some reef-building coral, Ⅱ. Stylophora pistillata (Esper). Sci Rep Tohoku Univ 18: 48-64 Babcock RC, Bull GD, Harrison PL, Heyward AJ, Oliver JK, Wallace CC, Willis BL (1986) Synchro- nous spawnings of 105 scleractinian coral spe- cies on the Great Barrier Reef. Mar Biol 90: 379-394 Fan TY, Li JJ, Le SX, Fang LS (2002) Lunar peri-odicity of laval release by Pocilloporid corals in Southern Taiwan. Zool Stud 41: 288-294 Fan TY, Lin KH, Kuo FW, Soong K, Liu LL, Fang LS (2006) Diel patterns of larval release by five
ハナヤサイサンゴ科 2 種の幼生放出期 15
brooding scleractinian corals. Mar Ecol Prog Ser 321: 133-142
Harii S, Omori M, Yamakawa H, Koike Y (2001) Sexual reproduction and larval settlement of zooxanthellate coral Alveopora japonica Eguchi at high latitudes. Coral Reefs 20: 19-23
Harriot VJ (1983) Reproductive seasonality, settle- ment and post-settlement mortality of Pocillo-pora damicornis (Linnaeus), at Lizard Island, Great Barrier Reef. Coral Reefs 2: 151-157 Harrison PL, Babcock RC, Bull GD, Oliver JK,
Wallace CC, Willis BL (1984) Mass spawning in tropical reef corals. Science 223: 1186-1189 Hayashibara T, Shimoike K, Kimura T, Hosaka S, Heyward A, Harrison P, Kudo K, Omori M (1993) Patterns of coral spawning at Akajima Island, Okinawa, Japan. Mar Ecol Prog Ser 101: 253-262 Heyward A, Yamazato K, Yeemin T, Minei M
(1987) Sexual reproduction of corals in Okinawa. Galaxea 6: 331-343
Isomura N, Nishihira M (2001) Size variation of planulae and its effect on the lifetime of planulae in three pocilloporid corals. Coral Reefs 20: 309-315 Jokiel PL (1985) Lunar periodicity of planula re-lease in the reef coral Pocillopora damicornis in relation to various envioronmental factors. Proc 5th Int Coral Reef Cong 4: 307-312 環境省・日本サンゴ礁学会(2004)日本のサンゴ礁. 環境省,東京 298p Marshall SM, Stephenson TA (1933) The breeding of reef animals. Part I. The corals. Sci Rep Gt. Barrier Reef Exped 3: 219-245 目崎拓真・林 徹・岩瀬文人・中地シュウ・野澤洋 耕・宮本麻衣・富永基之(2007)高知県大月町西 泊におけるイシサンゴ類の産卵パターン.Kuro-shio Biosphere 3: 33-47 御前 洋(1989)串本で観察されたイシサンゴの産 卵について.マリンパビリオ 18 (10): 58-59 御前 洋(1990)串本で観察されたイシサンゴの産 卵について(1990 年).マリンパビリオン 19 (10): 58-59 御前 洋(1995a)イシサンゴの産卵と幼サンゴの 定着状況─ 1994 年の観察より─.マリンパビリ オン 24 (5): 26-27 御前 洋(1995b)イシサンゴの産卵と幼サンゴの 定着状況─ 1995 年の観察より─.マリンパビリ オン 24 (11): 62-63 御前 洋(1996)イシサンゴの産卵と幼サンゴの定 着状況─ 1996 年の観察より─.マリンパビリオ ン 25 (12): 80 御前 洋(1997)イシサンゴの産卵と幼サンゴの定 着状況─ 1997 年の観察より─.マリンパビリオ ン 26 (11): 62-63 御前 洋(1998)イシサンゴの産卵と幼サンゴの定 着状況─ 1998 年の観察より─.マリンパビリオ ン 27 (12): 68 御 前 洋(1999) イ シ サ ン ゴ の 産 卵 に つ い て ─ 1999 年の観察結果─.マリンパビリオン 28 (11): 64-65 御 前 洋(2000) イ シ サ ン ゴ の 産 卵 に つ い て ─ 2000 年の観察結果─.マリンパビリオン 29 (12): 68 御 前 洋(2001) イ シ サ ン ゴ の 産 卵 に つ い て ─ 2001 年の観察結果─.マリンパビリオン 30 (12): 68-69 御 前 洋(2003) イ シ サ ン ゴ の 産 卵 に つ い て ─ 2002 年の観察結果─.マリンパビリオン 32 (4): 26-27 御 前 洋(2004) イ シ サ ン ゴ の 産 卵 に つ い て ─ 2003 年の観察結果─.マリンパビリオン 33 (1): 2-3 御 前 洋(2005) イ シ サ ン ゴ の 産 卵 に つ い て ─ 2004 年の観察結果─.マリンパビリオン 34 (3): 18-19 御前 洋(2006a)イシサンゴの産卵について─ 2005 年の観察結果─.マリンパビリオン 35 (1): 2-3 御前 洋(2006b)イシサンゴの産卵について─ 2006 年の観察結果─.マリンパビリオン 35 (6): 42-43 Nozawa Y (2012) Annual variation in the timing of coral spawning in a high-latitude environment: influence of temperature. Biol Bull 222: 192-202 Nozawa Y, Tokeshi M, Nojima S (2006) Reproduc-tion and recruitment of scleractinian corals in high-latitude coral community, Amakusa, south-western Japan. Mar Biol 149: 1047-1058
Permata WD, Kinzie III RA, Hidaka M (2000) His-tological studies on the origin of planulae of the coral Pocillopora damicornis. Mar Ecol Prog Ser 200: 191-200
Richmond RH, Jokiel PL (1984) Lunar periodicity in larva release in the reef coral Pocillopora damicornis at Enewetak and Hawaii. Bull. Mar Sci 34: 280-287 Rinkevich B, Loya Y (1979a) The reproduction of the Red Sea coral Stylophora Pistillata. I. Gonads and planulae. Mar Ecol Prog Ser 1: 133-144 Rinkevich B, Loya Y (1979b) The reproduction of the Red Sea coral Stylophora pistillata. Ⅱ. Syn-chronization in breeding and seasonality of planula shedding. Mar Ecol Prog Ser 1: 145-152 Schmidt-Roach S, Miller KJ, Woolsey E, Gerlach G, Baird AH (2012) Broadcast spawning by Pocillo-pora species on the Great Barrier Reef. PloS ONE 7 e50847 Schmidt-Roach S, Miller KJ, Andreakis N (2013) Pocillopora aliciae: new species of scleractinian coral (Scleractinia, Pocilloporidae) from subtropi-cal Eastern Australia. Zootaxa 3626 276 Schmidt-Roach S, Johnston E, Fontana S, Jury CP, Forsman Z (2014a) Daytime spawning of Pocillo-pora species in Kaneohe Bay, Hawaii. Galaxea 16: 11-12 Schmidt-Roach S, Miller KJ, Lundgren P, Andreakis N (2014b) With eyes wide open: A revision of species within and closely related to the Pocillo-pora damicornis species complex (Scleractinia, Pocilloporidae) using morphology and genetics. Zool J Linn Soc 170: 1-33
Suharsono M, Situmorang J, Kamiso HN (2008) Timing of larval release by reef coral Pocillipora damicornis at Panjang Island, Central Java. Marine Research in Indonesia 33 (I): 33-39 Tanner JE (1996) Seasonality and lunar periodicity in the reproduction of Pocilloporid corals. Coral Reefs 15: 59-66 Tioho H, Tokeshi M, Nojima S (2001) Experiment analysis of recruitment in a scleractinian coral at high latitude. Mar Ecol Prog Ser 213: 79-86 Thamrin, Nojima S, Tokeshi M (2001) Experimen-tal analysis of planula release in a scleractinian coral Alveopora japonica. Galaxea, Journal of Coral Reef Studies 3: 25-30
van Woesik R (1995) Coral communities at high latitude are not pseudopopulation: evidence of spawning at 32°N, Japan. Coral Reefs 14: 119-120.
Villanueva RD, Yap HT, Montano MNE (2008) Timing of planulation by pocilloporid coral in the northern Philippines. Mar Ecol Prog Ser 370: 111-119 渡邊美穂・岩瀬文人・横地洋之(2009)四国西南海 域の高緯度サンゴ群集における造礁サンゴ幼生加 入の季節変化.日本サンゴ礁学会誌 11: 73-81 Willis BL, Babcock RC, Harrison PL, Oliver JK, Wallace CC (1985) Patterns in the mass spawn-ing on the Great Barrier Reef from 1981 to 1984. Proc 5th Int Coral Reef Cong 4: 343-348 山里 清(1991)サンゴの生物学.5 サンゴの生殖, 東海大学出版,東京,pp 71-90 山里 清・エリア スワルデイ・サイーダ サルタナ (2008)高緯度における幼生放出型サンゴの生殖 周期.日本サンゴ礁学会誌 10: 1-11 Yeemin T (1991) Ecological studies of scleractinian coral communities above the northern limit of coral reef development in the western Pacific. Ph.D dissertation Kyushu University Fukuoka 101pp Zakai D, Dbinsky Z, Avishai A, Caaras T, Chadwick NE (2006) Lunar periodicity of planula release in the reef-building coral Stylophora pistillata. Mar Ecol Prog Ser 311: 93-102 2015 年 1 月 16 日 受領 2015 年 6 月 4 日 受理 Ⓒ 日本サンゴ礁学会
17
Seasonal and daily planulation patterns of two pocilloporid
corals at a high-latitude habitat in Southwest Kochi, Japan
Mai MIYAMOTO
1* and Hiroyuki YOKOCHI
21 National Research Institute of Far Seas Fisheries, Fisheries Research Agency, 2-12-4 Fukuura, Kanazawa,
Yokohama, Kanagawa 236-8646, Japan
2 Course of Applied Biological Science, Department of Fisheries, School of Marine Science and Technology,
Tokai University, 3-20-1 Orido, Shimizu, Shizuoka 424-8610, Japan * Corresponding author: M. Miyamoto
E-mail: [email protected]
Communicated by Saki Harii (Ecology Editor)
Abstract In order to clarify the characteristics of planulation and reproductive strategies of brooding scleractin-ian corals at high-latitude coral community, seasonal and daily planula release patterns of Pocillopora
damicor-nis and Stylophora pistillata were studied at Nishidomari, Kochi, Japan in 2006 and 2007. Distribution of
re-cruits and adult colonies were also examined. Colonies were collected at the study site and kept in tanks, and the number of daily released planulae was counted. Branch fragments were sampled from tagged colonies in the field, and oocytes and planulae in polyps were observed under a binocular microscope after decalcification. Observation of captive colonies and branch fragments showed that planulation occurred twice each year, either in June and July (P. damicornis) or in July and August (S. pistillata). Brooding of planulae in polyps was ob-served both years during the same periods. The duration of planulation was longer in S. pistillata (approxi-mately 25 days) than in P. damicornis (approxi(approxi-mately 15 days). The planulation of P. damicornis peaked around the new moon, but that of S. pistillata did not show a clear peak synchronized with lunar phase. In both species, planula release occurred all day long during the planulation periods although the number of planulae released was slightly higher at night. Compared to the studies from low latitude coral-reefs areas, both species had less frequent planulation periods per year but longer duration of planulation periods. In the distribution study, re-cruits of P. damicornis and S. pistillata were significantly higher around their parental colonies. These planula-tion and distribuplanula-tion patterns are supposed to reflect the reproductive strategies of these two species in high latitudes; increasing the opportunity of planulation and recruitment around parental colonies to maintain their local population.
Keywords: Planulation, high-latitude habitat, Pocillopora damicornis, Stylophora pistillata, lunar synchroniza-tion
Received: 16 January 2015/Accepted: 4 June 2015 Ⓒ Japanese Coral Reef Society Journal of the Japanese Coral Reef Society 17: 1-17(2015)