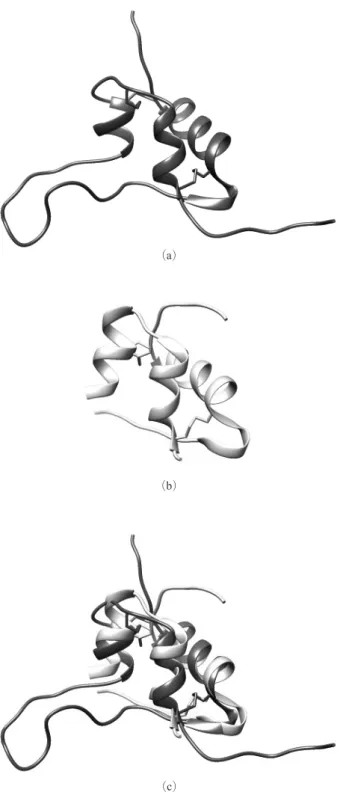
1. は じ め に 血糖値を下げる働きを持つホルモンとして有名なインス リンは残基数 51,原子数は約 690 の二本鎖のタンパク質 であり,3 つのジスルフィド結合を持つホルモンタンパク 質である.本研究で研究対象とするインスリン様成長因子 1(IGF-1)は残基数 70,原子数は約 1050 の一本鎖のタン パク質であり,インスリンと同様 3 つのジスルフィド結合 を持つホルモンタンパク質である.その名の通りインスリ ンとよく似た構造のタンパク質であるが,生体内で果たす 役割は大きく異なる. IGF-1 とインスリンは①配列,②立体構造の 2 点でよく 似ている. ①配列の相似性 IGF-1 分子は残基番号 1 番~29 番の B 領域,30 番~41 番のC 領域,42 番~62 番の A 領域,63 番~70 番の D 領 域の 4 つの領域に分かれている.一方でインスリンは 21 残基のA 鎖と 30 残基の B 鎖という 2 本の鎖からなる. IGF-1 の B 領域はインスリンの B 鎖に,IGF-1 の A 領域は インスリンのA 鎖に対応している.IGF-1 とインスリンの シーケンスアラインメントをとると,B 領域/ B 鎖では 29 残基中 14 残基が一致し,A 領域/ A 鎖では 21 残基中 11 残基が一致する.IGF-1 にしかない C 領域および D 領 域を除けばおよそ半分の残基が一致することになる. ②立体構造の相似性 IGF-1/インスリンの B 領域/ B 鎖には 1 本の,A 領域 /A 鎖には 2 本の α ヘリックスがある.これらの α ヘリ ックスはヘリックス内部での水素結合およびヘリックス間 のジスルフィド結合によって構造的に安定している.この 構造的に安定な領域において,IGF-1 とインスリンはよく 似た形をしている.IGF-1 の 3 つの α ヘリックスに含まれ る残基とインスリンの配列上対応する残基との間で,主鎖 の各原子のRMSD(平均二乗偏差)が最小になるように 2 つの分子を重ね合わせたところ(Figure 1),主鎖の 116 原 子のRMSD が 1.69Å という比較的小さな値となった. このようにIGF-1 とインスリンはよく似た構造を持ちな がら,生体内での機能は全く異なる.IGF-1 は細胞の成長 や分裂・分化などを促進する一方,インスリンは主に血糖 を 抑 制 す る 作 用 を 持 つ. こ の 機 能 の 違 い は,IGF-1 は IGF-1 受容体(IGF-1R)に,インスリンはインスリン受容 体(IR)にそれぞれ特異的に結合する基質特異性に由来す る.構造が似ているのに基質特異性が生じるメカニズムは 未だ明らかにされていない.そこで,これら 2 つの分子間
インスリン様成長因子1とインスリンの電子構造の比較
Comparison of Electronic Structures between Insulin-like Growth Factor 1 and Insulin
佐々木 光
*・平 野 敏 行
**・佐 藤 文 俊
**Hikaru SASAKI, Toshiyuki HIRANO and Fumitoshi SATO
要 旨 インスリン様成長因子(IGF-1)はインスリンとよく似た構造を持つタンパク質分子であり,細胞の成長,分化 などを促進する.一方,インスリンは血糖値を下げる作用をもつタンパク質である.すなわち,インスリンとイン スリン様成長因子は似た構造を持っているものの,それぞれ異なる受容体に結合している.これら 2 つの分子のど のような差異を利用しているかを推測するために,IGF-1 とインスリンの電子構造の比較を行った.その結果,こ れらの間で静電ポテンシャルの分布ならびに電子密度分布の特徴的な部位を確認することができた. AbstractInsulin-like growth factor (IGF-1), whose structure is similar to insulin, promotes growth and differentiation of cells. On the other hand, insulin lowers the level of sugar in the blood. These proteins are similar in structure, but they bind to different receptors. To infer what differences are used to discriminate between them, we compared those electronic structures. As a result, the significant differences were found in the distribution on electrostatic potential and electron density.
*東京大学大学院工学系研究科
**東京大学生産技術研究所 機械・生体系部門
の相違を電子レベルで明確にするために,本研究では密度 汎関数法(DFT)に基づく IGF-1 の正準 Kohn-Sham 分子軌 道を計算し,IGF-1 とインスリンの電子構造を比較して基 質特異性をもたらすような電子構造の違いを見出すことを 目指した. 2. 計 算 方 法 2.1 計算構造の作成 これまで発表されたIGF-1 の立体構造のうち,残基やプ ロトンの情報が欠落していない構造で,残基配列が天然の IGF-1 から改変されておらず,ひずみも小さい立体構造で ある 1PMX1)のmodel 1 を採用した.インスリンの立体構 造もIGF-1 と同様の基準で選び,2MVC2)のmodel 1 を採 用した. 緩衝溶液モデルとして正電荷を帯びた解離基の近傍には -1 の電荷を,正電荷を帯びた解離基の近傍には +1 の電荷 を置いた.IGF-1 の計算モデルには Arg 6 個,Lys 3 個の合
計 10 個の正電荷を帯びた解離基があり,またAsp 4 個, Glu 4 個および C 末端の合計 9 個の負電荷を帯びた解離基 がある.これらのうち塩橋を形成しているとみなせる Asp53 と Arg56 を除く,正電荷の解離基 9 個と負電荷の解 離基 8 個の近傍にそれぞれ -1 と +1 の点電荷を置いた.同 様に,インスリンの計算モデルにも -1 の点電荷 3 個と +1 の点電荷 5 個を付加した. 2.2 計算シナリオ 正準分子軌道計算の固有値方程式を解くためには自己無 撞着場(SCF)計算を繰り返してエネルギー等の値を収束 させる必要がある.このSCF 計算を収束させるためには 質の高い初期値が必要であり,本研究ではその初期値を求 めるためにQCLO法3)を用いた. QCLO 法は,タンパク質分子全体の電子状態を求めるた めに,分子の一部を切り取って作ったより小さな分子(こ れをフレーム分子と呼ぶ)の電子状態を先に求め,これを 使ってより大きな分子の電子状態計算の初期値を作成する. この拡張操作(ステップ)を数回経てタンパク質分子全体 の計算を達成する.このような計算の流れのことを「計算 シナリオ」と呼ぶ. IGF-1 の電子構造計算は,以下に示す 4 つのステップか らなる計算シナリオに沿って行った.ステップ 1 では 1 残 基ごとにフレーム分子を作って計算した.ジスルフィド結 合で結合した 3 対 6 つのシステイン残基(Cys6 - Cys48, Cys18 - Cys61, Cys47 - Cys52)および塩橋と判断される 1 対の解離基(Asp53 - Arg56)については,それぞれの対 の相互作用が大きく,1 つのフレーム分子として計算する 方が良い.そこで,これらアミノ酸残基対は 1 つのフレー ム分子を構成するように計算シナリオを作成した.ステッ プ 2 では 5~10 残基の連続する残基を 1 つのフレーム分子 とした.ただしジスルフィド結合を形成する 3 対 6 つのシ ステイン残基および塩橋の 1 対のアミノ酸残基のいずれか が含まれるフレーム分子には,対となる残基を付加した. ステップ 3 ではIGF-1 の A,B 領域(それぞれインスリン のA 鎖および B 鎖に対応)および C 領域のそれぞれを 1 (a) (b) (c)
Figure 1 (a) Structure of IGF-1, (b) Structure of insulin, and (c) Superposition of IGF-1 and insulin structures
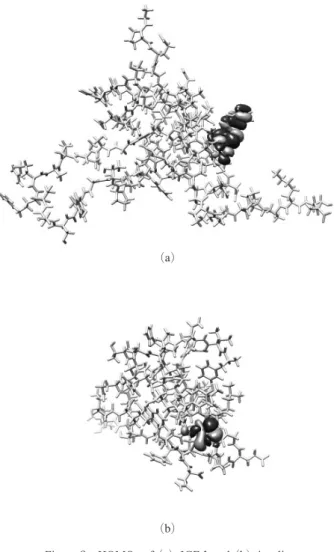
つのフレーム分子として計算を行った.なお,D 領域の計 算はステップ 2 で済んでいる.また,ジスルフィド結合お よび塩橋についてはステップ 2 と同様に取り扱った.最後 にステップ 3 の結果を用いて,最終ステップのステップ 4 ではIGF-1 分子全体の正準分子軌道の計算を行った.イン スリンについても同様の計算シナリオを作成した. 2.4 計算法と計算環境 タンパク質の密度汎関数計算にはProteinDF4),QCLO 法 計算にはQCLObot 5)を用いた.交換相関汎関数にはB3LYP6), 基底関数はdouble-zeta valence (DZV) を採用した.なお, インスリンのステップ 4 の計算時には,空軌道群を 0.1 a.u. 持ち上げるレベルシフト法7)を適用した.その後,レ ベルシフトを外して計算が収束することを確認した.計算 にはAMD Opteron Processor 6174 (12 コア) 4 ソケット× 4 ノード,メモリ 128 GB/ ノードを用いた. 3. 結 果 お よ び 考 察 3.1 正準 Kohn-Sham 分子軌道計算の達成 IGF-1 のステップ 4 について,SCF 計算における各分子 軌道のエネルギー準位および全エネルギーの推移を示した (Figure 2). Figure 2 は SCF 計算の各回転における IGF-1 の軌道エネ ルギー準位の変遷を示したものであり,縦軸がエネルギー (eV),横軸が SCF 回転数である.各 SCF 回転数における 一つ一つの軌道エネルギー値は横線で示しており,-5 eV 付近に観られる線は最高被占有軌道(HOMO),その上に ある軌道は最低空軌道(LUMO)である.Figure 2 から明 らかなとおり,SCF1 回転目から HOMO-LUMO 間に十分 なギャップを保持したままIGF-1 の正準分子軌道計算が極 めて安全に収束している様子が観測された. 3.2 分子軌道の比較 電子構造計算によって求められたIGF-1 およびインスリ ン分子の分子軌道を比較した.Figure 3, 4 に HOMO およ びLUMO を示した.分子軌道はいずれも等値面表示(0.01)
で示した.Figure 3(a)は IGF-1 の分子骨格の上に描いた IGF-1 の HOMO である.IGF-1 における HOMO は Phe16
およびVal17 のあたりの寄与が大きかった.(b)はインス
リンの分子骨格の上に描いたインスリンのHOMO である.
インスリンのHOMO は GluA17 の側鎖近辺の寄与が大き
かった.
Figure 4(a), (b)はそれぞれ IGF-1 およびインスリン のLUMO を示した.IGF-1 の LUMO は Leu10,Ile43 を中 心とした,分子の内側の疎水性残基が集まる領域の寄与が
大きかったのに対し,インスリンのLUMO は GlyA1 付近
の寄与が大きかった. Figure 2 Orbital energy distribution of IGF-1 vs. the number of SCF
(a)
(b)
IGF-1 およびインスリンにおけるフロンティア軌道の近 傍 10 個の軌道について比較したところ,以下の知見を得た.
・ Cys18/B19-Cys61/A20 のジスルフィド結合の付近に分布
する軌道はIGF-1 とインスリン両方の HOMO 軌道付近
の占有軌道に複数存在していた(IGF-1 の HOMO - 8 と HOMO - 10, イ ン ス リ ン の HOMO - 3, HOMO - 4 と HOMO - 8).
・ Cys6/B7-Cys48/A7 のジスルフィド結合の付近に分布す
る軌道はIGF-1 とインスリン両方の LUMO 軌道付近の
空 軌 道 に 複 数 存 在 し て い た(IGF-1 の LUMO + 2 と LUMO + 6,インスリンの LUMO + 1, LUMO + 4 と LUMO + 6)
・ Glu58/A17 の側鎖に分布する軌道は IGF-1 とインスリン
両 方 のHOMO 軌 道 付 近 の 占 有 軌 道 に 存 在 し て い た (IGF-1 の HOMO - 6, インスリンの HOMO - 2, HOMO - 1,
HOMO).インスリンの HOMO - 2, HOMO - 1, HOMO
はIGF-1 の HOMO - 6 と比べてエネルギー準位が 1~2
eV 高かった,インスリンの HOMO は IGF-1 のそれより
も 1.3 eV エネルギー準位が高く,LUMO は 1.0 eV 高い.
すなわち,インスリンのHOMO – LUMO ギャップ(1.1
eV)は IGF-1(1.4 eV)に比べて狭かった(0.3 eV). 3.4 静電ポテンシャル分布の比較
Figure 5 は IGF-1((a), (b), (e), (f))とインスリン((c), (d), (g), (h))の静電ポテンシャルを各等電子密度面(0.01) にマッピングしたものである.(a), (b)のペア,(c), (d) のペア,(e), (f)のペアおよび(g), (h)のペアは,そ れぞれ紙面縦方向を軸に分子を 180°回して描いたもので ある.また,(a)~(d)は静電ポテンシャルが負の領域,(e) ~(h)は正の領域をグレイスケールで描いている. IGF-1 とインスリンの共通領域 (A, B 領域) は,IGF-1 では静電ポテンシャルが正の部分と負の部分の両方が観ら れた一方で,インスリンは全体的に負に分布した.また, IGF-1 にしかない C 領域と D 領域は,正の解離基が 2 個ず つあるため正に分布した.共通領域における解離基の数は IGF-1 では正 6 個と負 8 個,インスリンでは正 4 個と負 6 個であり,両者ともに負の解離基が多いが,IGF-1 の共通 領域の静電ポテンシャル分布はC 領域と D 領域の正の解 離基の影響も受けていると考えられる.
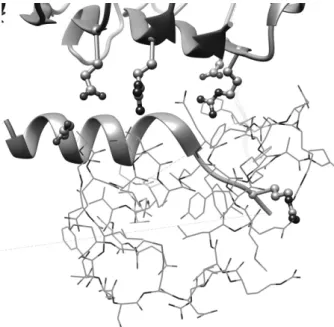
2018 年,Xu らによって IGF-1 と IGF-1 受容体(IGF-1R) の複合体の立体構造(PDB ID: 5U8Q)が報告され,IGF-1 とIGF-1R との相互作用部位が明らかになった8).これに よると,IGF-1 の 17 のアミノ酸残基が IGF-1R の 17 のア ミノ酸残基とコンタクトしている. IGF-1R の 17 個のアミノ酸残基のうち,正の解離基と負 の解離基はそれぞれ 3 個ずつある.Figure 6 は,これらの 解離基の側鎖をボール&スティックモデルで描き,同時に IGF-1 をワイヤーモデルで描き,両者の位置関係を示した ものである.正の解離基の方が負の解離基よりもIGF-1 の 近くに位置していることが見て取れる.このことから,リ ガンドの負の静電ポテンシャルを持つ領域と相補性が高く なる可能性がある. 複合体構造 5U8Q 中の IGF-1 の構造は誘導適合のため IGF-1 単体の構造 1PMX とは若干異なるが,5U8Q の 1GF-1R と 1PMX との間でドッキングシミュレーションを行っ たところ,IGF-1 の Figure 5(b), (f)の方向で中央部に IGF-1R の各アミノ酸残基が配置することが分かった.イ ンスリン 2MVC でも同様の操作を行ったところ,やはり インスリンのFigure 5(d), (h)の方向で IGF-1R の各ア ミノ酸残基がIGF-1 と同様に配置することが分かった (Figure 7). このモデルにおけるリガンド側の相互作用部位の静電ポ テンシャル分布は,IGF-1 では正負両者の部位がある一方 でインスリンでは全体的に負に大きく偏っている.すなわ ち,驚いたことに静電ポテンシャル分布の差からはインス リンの方がIGF-1R とより結合しやすく見える. (a) (b)
(a) (c) (e) (g) (b) (d) (f) (h)
Figure 5 Electrostatic potential distribution of IGF-1((a), (b), (e) and (f)) and insulin((c), (d), (g) and (h)). (a), (b), (c) and (d) are drawn in range of -0.3 ~ 0, and (e), (f), (g) and (h) are drawn in range of 0 ~ 0.3, respectively. (b), (d), (f) and (h) are views from the opposite direction from (a), (c), (e) and (g).
3.5 マリケン電荷の比較 IGF-1 とインスリンの電子密度分布を比較するため,電 子密度分布を原子電荷の偏りとして反映するマリケン電荷 を比較した.ここでは,アミノ酸残基ごとにマリケン電荷 の和を求め,アミノ酸残基単位の電荷の偏りとして解析し た(Table 1).灰色で網掛した部分は IGF-1 とインスリン との間で配列が保存されている部分である.太字はIGF-1 の相互作用部位である(Figure 7).Figure 7 の結果から, IGF-1 のみに存在する Glu40 を除き,対応するアミノ酸残 基がインスリンの相互作用部位と仮定した.Glu40 を除く 16 アミノ酸残基の内,10/16 がIGF-1 とインスリンで保存 されている同じアミノ酸残基である. まず,保存されている同じアミノ酸残基の間でマリケン 電荷が大きく異なるならば,その違いは分子の電子構造を 反映したものと考えてよい.保存されているアミノ酸残基 のうち,マリケン電荷の残基ごとの合計の差が大きかった ものは,以下の 5 つである. ◦ Arg21 / B22: 0.21 ◦ Glu58 / A17: -0.20 ◦ Val44 / A3: -0.12 ◦ Gly42 / A1: 0.10 ◦ Leu15 / B16: -0.09 ここで,アミノ酸残基名IGF-1 での番号 / インスリンで の番号:マリケン電荷の差である.
このうちArg21 / B22 と Glu58 / A17 に大きな電荷の差 が見られた.インスリンではこれら 2 つの残基が塩橋を形 成している一方で,IGF-1 にはその塩橋がないことが主な 原因であると考えられる.Val44 / A3 と Gly42 / A1 にお けるマリケン電荷の差は,これら近傍にある解離基の違い に起因するものとみられる.すなわち,Val44 / A3 につい ては,インスリンでは近傍に正の解離基であるA 鎖の N 末端がある一方で,IGF-1 ではこれがない.また Gly42 / A1 については,インスリンの方が IGF-1 よりも Gly42 / A1 と負の解離基(Glu45/AspA4)との距離が近かった. Leu15 / B16 についてはマリケン電荷の差を生じさせる原 因を近傍には見つけることができなかった.全体の電子構 造のバランスにより生じた差であることが示唆された.同 じアミノ酸残基でもマリケン電荷に差を観測したことは, タンパク質全体の正準分軌道計算だからこそ得られた結果 であることを強調したい. 続いて,IGF-1R との相互作用部位におけるマリケン電 荷の差に注目した.IGF-R の各残基の電荷は 0 か± 1 のい ずれかで中間の値をとることはなく,またコンタクトして いる 2 つの残基の距離がすべて等しいと近似すると,マリ ケン電荷の差が静電相互作用引力の差に比例するとみなせ るからである.Figure 6 の IGF-1R の正の解離基と相互作 用するIGF-1 および対応するインスリンのアミノ酸残基群 において,それぞれマリケン電荷の和を取ってみると IGF-1 のマリケン電荷の和とインスリンのマリケン電荷の 和の差は -0.091 であった.同様にIGF-1R の負の解離基と 相互作用するIGF-1 およびインスリンのアミノ酸残基群で は,IGF-1 のマリケン電荷の和とインスリンのマリケン電 荷の和の差は -0.015 であった. このようにIGF-1R の解離基が相互作用する相手のリガ ンド部位に着目し,電荷分布差を詳細に検討することで, 実はIGF-1 との静電相互作用引力はインスリンよりも強い ことが示唆された.IGF-1 がインスリンよりも IGF-1R と 強く結合する要因の 1 つだと考えられ,静電ポテンシャル 分布の解析だけでは分からなかったことである.なお,エ ネルギー値などによるさらに詳細な議論には,今後のリガ ンド受容体複合体計算による解析が必要である.
Figure 6 Charged residues of IGF-1R in interaction sites between IGF-1 and IGF-1R8) (drawn by the ball & stick model).
1 is drawn by the wire model, and main chain of IGF-1R is drawn by the ribbon model.
Figure 7 Interaction sites between IGF-1 and IGF-1R (left), between insulin and IGF-1R (right). Amino residues of IGF-1R are drawn by the wire model.
領域 B アミノ酸残基 IGF-1 - G P E T L C G A E L V D A L インスリン F V N Q H L C G S H L V E A L マリケン 電荷 IGF-1 - 0.87 0.00 -0.99 0.05 0.02 -0.02 -0.03 0.01 -0.98 -0.02 -0.09 -0.91 0.01 -0.06 インスリン 0.83 0.04 0.00 -0.03 0.04 0.03 -0.08 -0.02 0.00 0.02 -0.01 -0.04 -0.94 -0.01 0.04 差分 - 0.83 0.00 -0.96 0.01 -0.01 0.06 -0.02 0.01 -1.00 -0.01 -0.04 0.04 0.02 -0.09 領域 B アミノ酸残基 IGF-1 Q F V C G D R G F Y F N K P T インスリン Y L V C G E R G F F Y T P K T マリケン 電荷 IGF-1 0.00 0.08 0.04 -0.14 0.03 -0.96 0.96 -0.01 0.00 0.04 -0.04 -0.04 0.96 0.04 0.12 インスリン -0.03 0.00 0.01 -0.12 0.09 -1.00 0.75 -0.01 -0.03 0.07 -0.03 -0.05 0.07 0.95 0.07 差分 0.03 0.08 0.03 -0.02 -0.06 0.04 0.21 -0.01 0.04 -0.03 -0.01 0.01 0.89 -0.92 0.06 領域 C アミノ酸残基 IGF-1 G Y G S S S R R A P Q T インスリン - - - - - - - - - - - - マリケン 電荷 IGF-1 -0.04 -0.09 0.05 0.05 -0.04 0.02 0.99 0.91 0.08 0.01 -0.05 -0.02 インスリン - - - - - - - - - - - - 差分 - - - - - - - - - - - - 領域 A アミノ酸残基 IGF-1 G I V D E C C F R S インスリン G I V E Q C C T S I マリケン 電荷 IGF-1 0.18 -0.02 -0.13 -0.90 -0.98 0.03 0.00 -0.03 0.98 0.02 インスリン 0.08 0.03 -0.01 -0.95 0.04 -0.05 0.06 -0.01 -0.02 0.07 差分 0.10 -0.05 -0.12 0.05 -1.02 0.08 -0.07 -0.03 1.00 -0.05 領域 A アミノ酸残基 IGF-1 C D L R R L E M Y C A インスリン C S L Y Q L E N Y C N マリケン 電荷 IGF-1 0.00 -0.85 -0.02 0.99 0.88 0.03 -0.92 0.03 -0.05 0.07 0.02 Insulin -0.01 -0.03 0.04 0.05 -0.06 -0.03 -0.72 -0.06 -0.08 0.14 -0.86 差分 0.01 -0.82 -0.06 0.94 0.94 0.06 -0.20 0.08 0.03 -0.06 0.88 領域 D アミノ酸残基 IGF-1 P L K P A K S A インスリン - - - - - - - - マリケン 電荷 IGF-1 0.01 -0.05 1.02 0.01 -0.02 0.98 -0.05 -0.82 インスリン - - - - - - - - 差分 - - - - - - - -
* IGF-1 の 29Thr の元の電荷は 0.086,42Gly の元の電荷は 0.014.インスリンの B30Thr の元の電荷は-0.87,A1Gly の元の電荷は 0.82.こ
れら 4 つの残基についてはN 末端および C 末端の効果を取り除くために,末端部を除外してからマリケン電荷の和を取った.
Table 1 Summation of Mulliken charges in residues of IGF-1 and insulin. Shaded areas correspond to residues common to IGF-1 and insulin. Bold figures correspond to residues in binding sites between IGF-1 or insulin and IGF-1R.
4. お わ り に インスリンとアミノ酸残基配列と立体構造の良く似た IGF-1 が基質特異性を現す原因を探るため,IGF-1 および インスリンの密度汎関数法に基づく正準Kohn-Sham 分子 軌道計算を達成し,電子レベルで構造を比較した.その結 果,静電ポテンシャルと電子密度分布の特徴的な部位を確 認することができた.これらの部位はIGF-1 がインスリン よりもIGF-1R と強く結合することに寄与している可能性 がある.このような結果は正準分子軌道で初めて明らかに されたものである. 一方,本研究によってIGF-1 とインスリンの電子レベル で構造の差異が現れるのは一部であり,総体的には似てい ることも明らかとなった.これではインスリンアナログな らばIGF-1R と結合してもおかしくはない.実際,インス リンアナログによる発がん性が問題となっている. インスリンアナログはインスリンの配列を一部改変する ことによって超即効性や遅効性などの性質を持たせた人工 インスリンであり,糖尿病の治療に役立てられている.こ のインスリンアナログの中には恐らくIGF-1R と誤って結 合してしまうために発がん性を持つと疑われているものも 存在する9), 10).その 1 つであるグラルギンはインスリンB 鎖のC 末端にアルギニン 2 個を付加したものである. 本研究により,IGF-1R がインスリンよりも IGF-1 と強 く結合するメカニズムは静電相互作用の差に依存している と考えられた.正の解離基を 2 個付加するグラルギンが IGF-1R と強く結合するようになってしまう可能性は十分 に考えられる.より議論を深めるにはグラルギンや IGF-1R とのドッキングモデルによる電子構造計算が待たれる. (2020 年 3 月 17 日受理) 参 考 文 献
1) M. L. Schaffer et.al., “Complex with a Phage Display-Derived Peptide Provides Insight into the Function of Insulin-like Growth Factor I,” Biochemistry, 42, 31(2003), pp9324-9334
2) K. Křížková et.al., “Structural and Functional Study of the GlnB22-Insulin Mutant Responsible for Maturity-Onset Diabetes of the Young,” PLOS ONE, 9, 11(2014), p. e112883
3) H. Kashiwagi et.al., “Convergence process with quasi-canonical localized orbital in all- electron SCF calculation on proteins,” Molecular Physics, 101, 1-2(2003), pp81-86
4) F. Sato et.al., “Development of a new density functional program for all-electron calculation of proteins,” International Journal of Quantum Chemistry, 63, 1(1997), pp245-246
5) T. Hirano, “ProteinDF software package,” [Online]. Available: http://proteindf.github.io/.
6) A. D. Becke, “Density-functional thermochemistry. III. The role of exact exchange,” The Journal of Chemical Physics., 98, 7(1993) pp5648-5652
7) R.Carbó et.al., “Unconditional convergence in SCF theory: a general level shift technique,” Chemical Physics Letters, 47, 3(1977), pp581-583
8) Xu Y. et.al., “How ligand binds to the type 1 insulin-like growth factor receptor,” Nature Communications, 9, Article number: 821(2018)
9) L. G. Hemkens et.al., “Risk of malignancies in patients with diabetes treated with human insulin or insulin analogues: a cohort study,” Diabetologia, 52, 9(2009), pp1732-1744
10) D. Weinstein et.al., “Insulin analogues display IGF‐ I‐ like mitogenic and anti‐apoptotic activities in cultured cancer cells,” Diabetes Metabolism Research and Reviews, 25, 1(2009), pp41-49