Title ヨーロッパ腐蛆病菌(Melissococcus plutonius)の多様性および新たな検出法に関する研究 ( 本文(Fulltext) ) Author(s) 荒井, 理恵 Report No.(Doctoral Degree) 博士(獣医学) 甲第411号 Issue Date 2014-03-13 Type 博士論文 Version ETD URL http://hdl.handle.net/20.500.12099/49034 ※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。
ヨーロッパ腐蛆病菌(
Melissococcus plutonius
)の
多様性および新たな検出法に関する研究
2013 年
岐阜大学大学院連合獣医学研究科
(岐阜大学)
荒井理恵
目次
緒言
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・1第一章
M. plutonius
様菌の分類学的位置づけの検討および国内
で 分 離 さ れ た
M. plutonius
株 の 性 状 ・ 分 子 疫 学的 解
析
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・4 序論・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・5 材料および方法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・6 1 供試株 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・6 2 供試培地・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・6 3 培養性状の確認 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・7 4 生化学性状試験・・・・・・・・・・・・・・・・・・・・・・・・・・・・・7 5 グラム染色 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・7 6 ゲノムDNA の抽出 ・・・・・・・・・・・・・・・・・・・・・・・・・・・8 7 16S rRNA 遺伝子解析・・・・・・・・・・・・・・・・・・・・・・・・・・8 8 DNA-DNA ハイブリダイゼーション ・・・・・・・・・・・・・・・・・・9 9 パルスフィールドゲル電気泳動法(PFGE)によるM. plutonius 株およ びM. plutonius 様菌株の遺伝的背景の解析・・・・・・・・・・・・・・10 10 アクセッション番号・・・・・・・・・・・・・・・・・・・・・・・・・12 結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・13 1 M. plutonius および M. plutonius 様菌の性状・・・・・・・・・・・・・13 (1)培養性状・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・13 (2)生化学性状・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・14 (3)コロニー形状およびグラム染色像・・・・・・・・・・・・・・・・・・14 2 M. plutonius 様菌の分類学的位置づけ・・・・・・・・・・・・・・・・・15 3 M. plutonius の遺伝学的多様性・・・・・・・・・・・・・・・・・・・・16考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・17 図表 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・20
第二章
M. plutonius
非典型株の病原性の証明
・・・・・・・・・・・29 序論 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・30 材料および方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・32 1 供試動物と飼育方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・32 2 供試株と感染方法・・・・・・・・・・・・・・・・・・・・・・・・・・・・32 3 感染幼虫および人工餌からのM. plutonius 分離・・・・・・・・・・・・33 4 生存曲線の作製・・・・・・・・・・・・・・・・・・・・・・・・・・・・・33 結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・34 1 ミツバチ幼虫へのM. plutonius 感染の確認と臨床症状 ・・・・・・・34 2 生存率および生存曲線・・・・・・・・・・・・・・・・・・・・・・・・・34 考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・36 図表 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・39第三章
M. plutonius
典型株/非典型株検出・識別用 Duplex PCR
法の開発
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・41 序論 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・42 材料および方法 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・44 1 供試株・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・44 (1)腐蛆からのM. plutonius 分離・・・・・・・・・・・・・・・・・・・・44 (2)健康幼虫からの細菌分離・・・・・・・・・・・・・・・・・・・・・・・44 2 DNA 抽出・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・45 3 特異遺伝子領域の抽出およびプライマー設計・・・・・・・・・・・・・45 4 M. plutonius 特異的 PCR 法の特異性と感度の検証・・・・・・・・・・46 5 野外発症幼虫を用いたM. plutonius 検出法の有用性の検証 ・・・・・46 6 アクセッション番号 ・・・・・・・・・・・・・・・・・・・・・・・・・・47結果 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・48 1 Duplex PCR 法の標的遺伝子とプライマーセット・・・・・・・・・・・48 2 各PCR 法の特異性・・・・・・・・・・・・・・・・・・・・・・・・・・・48 3 Duplex PCR 法の感度・・・・・・・・・・・・・・・・・・・・・・・・・49 4 野外発症幼虫を用いたM. plutonius 分離培養法と Duplex PCR 法の比 較・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・49 考察 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・51 図表 ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・54
結論
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・64謝辞
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・65引用文献
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・661
緒言
ヨーロッパ腐蛆病(European foulbrood; EFB)はミツバチの細菌感染症で あり,欧米諸国では19 世紀終わりから知られる古典的な疾病である。主に 4~5 日齢の無蓋幼虫が発症し,発症幼虫は黄色~茶色に変色,水様性となり,死に 至る。死亡幼虫はPaenibacillus alvei や Enterococcus faecalis 等の二次感染 菌の作用によって変性し,酸臭を発することもある。現在,本病は,アジア・ オセアニア・アフリカ地域を含む世界中で発生が認められ,養蜂業に大きな経 済的損失を与える疾病の一つとして認識されている。 原因菌のヨーロッパ腐蛆病菌(Melissococcus plutonius)はグラム陽性紡錘 形球菌であり,顕微鏡下の観察に基づき,White によって 1912 年にその存在 が初めて報告された(43)。しかし,M. plutonius は発育に特殊な培養条件を 必要とするため,長い間,その細菌学的性状は未知のままであった。その後, 1957 年に Bailey が本菌の培養に成功したことによって,詳細な性状が初めて 明らかとなった(4)。Bailey と Collins の報告によると,M. plutonius の発育 には嫌気性または微好気性環境と二酸化炭素が必要とされている。さらに,発 育に必要な培地中のNa/K 比は 1 未満,すなわち,ナトリウム濃度よりカリウ ム濃度の方が高い環境下でのみ発育すると報告されている(9)。そのため,培 養時には培地へのカリウム塩添加が必要であり,カリウム塩無添加の Brain Heart Infusion (BHI)培地には発育しないことが知られていた。実際,世界 各地のEFB 症例から分離されるほぼ全てのM. plutonius 株の発育には嫌気ま たは微好気条件と培地へのカリウム塩添加が必要であり,また,分離株の形態 や抗原性も均質であった(1, 10, 11)。さらに,Djordjevic らが,オーストラリ ア各地で分離されたM. plutonius 株とイギリスで分離された M. plutonius 株
2
の遺伝学的な類似性を調べたところ,これらの株は,分離地域にかかわらず, 互いに類似した遺伝的背景を持つことが示された(15)。そのため,M. plutonius は遺伝的にも性状的にも類似した,均一な株によって構成されている菌種だと これまで考えられてきた。
EFB は日本では,Paenibacillus larvae を原因とするアメリカ腐蛆病ととも に,家畜伝染病予防法(昭和26 年 5 月 31 日法律第 166 号)において法定伝染 病に指定されており,1981 年にその初発例(17)が報告されて以降,全国的 に散発的な発生が認められている。家畜伝染病の標準的診断手法がまとめられ ている病性鑑定指針(33)には,過去の報告(9)に従い,「M. plutonius は BHI 培地に発育しない」ことが明記されているが,我が国では EFB を疑う幼 虫から培地へのカリウム塩添加を必要としない,すなわち,BHI 培地に発育す る M. plutonius 様菌が分離される事例がしばしばある。しかし,これら M. plutonius 様菌が分類学的に M. plutonius と同一であるのか,また,本当に EFB を引き起こしているのかについては十分な検討がされていなかった。 ミツバチは蜂蜜の生産だけでなく,農作物生産における花粉媒介という重要 な役割を担っている。花粉媒介によるミツバチの経済効果は,アメリカ合衆国 内においてだけでも146 億ドル(約 1 兆 4600 億円)に上ると推定されている (31)。また,日本国内においても 3000 億円を上回る経済効果がミツバチによ る花粉媒介によってもたらされていると試算されている(32)。そのため,EFB を始めとするミツバチの疾病は養蜂業のみならず,農業全般に大きな損害を与 える重要な問題である。そのような重要性にもかかわらず,EFB の発病メカニ ズムは未だ全く分かっていない。その解明のためには実験的に EFB を再現す る手法が必要となるが,M. plutonius は人工培地で継代培養を繰り返すことに より急速にその病原性が低下することが報告されている(3, 30)。そのため,
3 人工培養菌を用いた感染実験によりEFB を再現することは非常に困難であり, このことが本病の病態解明の大きな妨げとなっている。また,原因菌である M. plutonius 自体に関する報告も少なく,基本性状に関する情報はこの 20 年 弱全く更新されていない。さらに,研究に用いられている株もヨーロッパ,南 北アメリカおよびオーストラリアで分離された株がほとんどであり,日本を含 むアジア分離株についての情報は皆無に等しい。このような情報不足のため, 典型的なM. plutonius 株と異なる性状を示す M. plutonius 様菌株が見過ごさ れ,より大きな経済的損失を被ってきた可能性がある。 そこで本研究では,EFB の正確な診断・予防の一助とすることを目的として, 以下の3つの実験を実施した。第一章では,これまで均一だと考えられてきた M. plutonius の多様性を調べるため,国内で分離される M. plutonius 様菌の 分類学的位置づけの検討と国内分離M. plutonius 株の性状・分子疫学的解析を 行った。第二章では,M. plutonius 様菌のミツバチに対する病原性を確認する ため,セイヨウミツバチ幼虫を用いて人工培養菌による感染実験を行った。第 三章では,EFB の診断・疫学調査に活用することを目的に,M. plutonius と M. plutonius 様菌を EFB 発症幼虫から直接検出し,かつ両者を識別するため のDuplex PCR 法の開発を試みた。
4
第一章
M. plutonius
様菌の分類学的位置づけの検討および国内で
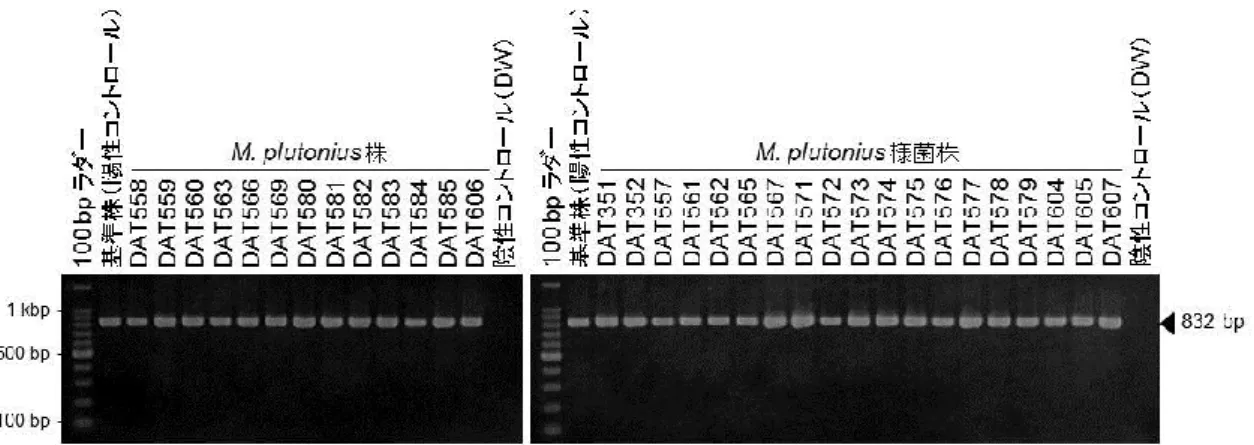
5 序論 M. plutonius は一属一種の菌である。世界各国の分離株はいずれも類似の発 育・生化学性状を示し,また,血清学的試験,分子疫学的解析からも株間の多 様性が非常に少ない菌種だと考えられている(1, 10, 11, 15)。 ところが,日本の EFB 野外症例では,培地へのカリウム塩添加を必要とす る典型的な性状を示すM. plutonius が分離される一方で,培地へのカリウム塩 添加を必要としない,すなわち,BHI 培地に発育するM. plutonius 様菌がし ばしば分離されてきている。これらM. plutonius 様菌は,その培養性状は M. plutonius と大きく異なるものの,コロニーの形状やグラム染色像は M. plutonius と類似しており,M. plutonius の同定を目的とした PCR(23)でも M. plutonius に特異的な増副産物が観察される(図 1-1)。しかし,M. plutonius 様菌の詳細な性状や分類学的位置づけは未決定のままであり,家畜保健衛生所 などのEFB 診断の現場において混乱が生じている。 そこで,本章では,日本各地で分離されたM. plutonius 株と M. plutonius 様菌株を収集し,M. plutonius 様菌の分類学的位置づけの検討を行うとともに, これらの株の性状・分子疫学的解析を行った。
6
材料および方法
1 供試株
計33 株のM. plutonius および M. plutonius 様菌株を用いた(表 1-1)。33 株全てがEFB の臨床症状を示すセイヨウミツバチ(Apis mellifera)幼虫から の分離株であり,16S rRNA 遺伝子を標的としたM. plutonius 特異的 PCR(23) で標的遺伝子領域の増幅が確認された(図1-1)。本章では,これらM. plutonius 特異的PCR 陽性株のうち,カリウム塩無添加培地である BHI 培地には発育し ない株を病性鑑定指針(33)に従ってM. plutonius 株とし,一方,BHI 培地 に発育する株をM. plutonius 様菌株として各種試験を行った。33 株中 31 株(M. plutonius 株 12 株および M. plutonius 様菌株 19 株)は日本で 1988~2010 年 に分離された株であり,残り2 株はイギリス(基準株; ATCC 35311)およびパ ラグアイ(DAT569 株)で分離された。基準株は American Type Culture Collection から購入した。
2 供試培地
本研究で用いた培地およびその組成を表 1-2 に示した。Medium 1 は M. plutonius 分離用培地として Bergeyʼs Manual of Systematic Bacteriology (13)に記載されている。Medium 2 ~ Medium 6 は Medium 1 の組成を一 部変更したものである。KSBHI,SBHI,KBHI 培地は BHI 培地(Difco Laboratories, Becton Dickinson, Sparks, MD, USA)を基礎培地とし,リン酸 二水素カリウム(終濃度; 0.15 M)および可溶性デンプン(終濃度; 1%)の両 方またはいずれか一方を添加した培地である。
7 3 培養性状の確認 培養性状の確認のために,各供試株を表1-2 に示す培地を用いて,好気,5% 炭酸ガスおよび嫌気(アネロパックシステム; 三菱ガス化学(株), 東京, 日本) 条件下で37℃・1 週間培養した。KSBHI 培地で嫌気培養した基準株の発育の 程度を+と評価し,これに比較して,より旺盛な発育を+V,より微弱な発育を +Wと評価した。全く発育が認められないもの,および極めて微弱な発育しか 認められないものは-と評価した。 4 生化学性状試験 カタラーゼ試験およびオキシダーゼ試験は定法により行った。その他の生化 学性状試験はAPI20A(bioMerieux, Marcy l'Etoile, France),ラピッド ID32A (bioMerieux)および ID テスト・HN-20 ラピッド「ニッスイ」(日水製薬(株), 東京, 日本)を用いた。いずれの試験も供試菌として,KSBHI 培地で 48 時間 嫌気培養した菌を用いた。 さらに,D-セロビオース,ラクトース,D-ラフィノースおよび D-キシロー スからの酸産生は,糖分解試験用培地(表1-2)を用いて確認試験を実施した。 KSBHI 培地で 48 時間嫌気培養した供試菌を集菌し,概ねマクファーランド 4 の濁度になるよう滅菌蒸留水に浮遊させた。この菌液0.1 ml を 5 ml の試験用 培地が入った試験管管底に接種,37℃・2 週間嫌気培養後,培地の色調変化に より判定を行った。 5 グラム染色 Medium 1 および KSBHI 培地で 37℃・嫌気培養した菌株を,グラム鑑別用 染色液フェイバーG「ニッスイ」(日水製薬(株))を用い,グラム染色した。
8
染色像の観察は光学顕微鏡下(倍率1,000 倍)で行った。
6 ゲノムDNA の抽出
KSBHI 培地で嫌気培養した各菌株を集菌し,5 mg/ml lysozyme および 40 U/ml mutanolysin を含む 500 µl の 5 × TE[50 mM Tris-HCl (pH 8.0), 5 mM EDTA (pH 8.0)] に浮遊した。37℃・1 時間インキュベートして細胞壁成 分を分解した後,10% sodium dodecyl sulphate(SDS)を 20 µl 添加して菌体 を完全に溶菌させ,等量のフェノールで 1 回,フェノール-クロロホルム-イソ アミルアルコール(25:24:1)(PCI)で 3 回以上,さらにクロロホルムで 1 回 以上処理することで,タンパク質や脂質などの夾雑物を取り除き,DNA を抽 出した。抽出したDNA をエタノールで沈殿させた後,70% エタノールで数回 洗浄し,TE[10 mM Tris-HCl (pH 8.0), 1 mM EDTA (pH 8.0)] 100 µl に溶 解した。DNA-DNA ハイブリダーゼーションに用いる DNA は,さらに,50 µg/ml RNase で 37℃・1 時間,200 µg/ml プロテナーゼKで 37℃・1 時間処 理した後,PCI およびクロロホルムで再抽出し,エタノール沈殿を行った。必 要に応じ,菌量および各溶液の容量を増やして抽出を行った。 7 16S rRNA 遺伝子解析 各 菌 株 の 16S rRNA 遺 伝 子 を , 既 報 の プ ラ イ マ ー セ ッ ト F1 ( 5'-GAGTTTGATCCTGGCTCAG-3' ) お よ び R13 (5'-AGAAAGGAGGTGATCCAGCC-3')(16)とTaKaRa Ex Taq(タカラバ イオ(株), 大津, 日本)を用いてゲノム DNA から PCR 法で増幅した。増幅 産物の塩基配列はBigDye terminator v3.1 cycle sequencing kit(アプライド バイオシステムズ, 東京, 日本)および 3130xl Genetic Analyzer(アプライド
9
バイオシステムズ)を用いて決定し,決定した塩基配列の解析は Sequencher Ver. 4.8(Gene Codes Corporation, Ann Arbor, MI, USA),CLUSTALW Ver. 1.83 ( http://clustalw.ddbj.nig.ac.jp/top-e.html ) お よ び BLAST (http://www.ncbi.nlm.nih.gov/BLAST)を用いて行った。 8 DNA-DNA ハイブリダイゼーション DNA-DNA ハイブリダーゼーションは,Ezaki らの方法(18)および Kusunoki らの方法(27)に準じ,マイクロプレート法により行った。 供試菌株には16S rRNA 遺伝子解析結果から,M. plutonius 株 3 株(基準株, DAT585 株, DAT606 株),M. plutonius 様菌株 4 株(DAT351 株, DAT561 株, DAT565 株, DAT607 株)を代表株として用いた。さらに,Melissococcus 属と 分類学上最も近縁な属であるEnterococcus 属に属する,Enterococcus faecalis (基準株; NCTC 775)を対照として用いた。 先ず,200 µg/ml の供試菌株ゲノム DNA 液および子牛胸腺 DNA 液(陰性コ ントロール)を10 分煮沸後,氷冷し,一本鎖 DNA とした。この一本鎖 DNA 液を0.1 M MgCl2添加リン酸緩衝生理食塩水(PBS)で 10 倍希釈し,その 100 µl をマイクロプレートウェルに添加して 30℃・16 時間インキュベートするこ とにより,各ウェルの底面に一本鎖 DNA を固相化させた。固相化後,残りの DNA 液を除去し,ウェルを乾燥させた。 固相化一本鎖 DNA と反応させる熱変性一本鎖 DNA は,メーカーの推奨す る方法に従い,フォトビオチン(Vector Laboratories, Inc., Burlingame, CA, USA)で標識した。ハイブリダイゼーション液は,標識一本鎖 DNA を 3 µg/ml の 濃 度 に な る よ う に プ レ ハ イ ブ リ ダ イ ゼ ー シ ョ ン 液 (2 × SSC, 2 × Denhardt’s solution, 50% ホルムアミド, 40 µg/ml 熱変性一本鎖子牛胸腺
10 DNA)に添加して作製した。 標識一本鎖 DNA のウェル底面への非特異的な吸着を防ぐため,各ウェルを 200 µl のプレハイブリダイゼーション液で 37℃・30 分間インキュベートした。 プレハイブリダイゼーション液を除去後,ハイブリダイゼーション液 100 µl を添加し,37℃・16 時間インキュベートすることで,ウェルに固相化させた一 本鎖DNA の相同配列に標識一本鎖 DNA が結合した二本鎖 DNA を形成させた。 各ウェルを1 × SSC で 3 回洗浄して二本鎖 DNA を形成していない標識一本 鎖DNA を除去した後,ヒストファイン((株)ニチレイバイオサイエンス, 東 京, 日本) を 1% 牛血清アルブミン含有 PBS で 10 倍希釈して作製したスト レプトアビジン-アルカリフォスファターゼ液 100 µl を添加し,37℃・30 分間 インキュベートすることで,形成した二本鎖 DNA 中の標識フォトビオチンに アルカリフォスファターゼを結合させた。各ウェルを1 × SSC で 3 回洗浄後, アルカリフォスファターゼの基質となる p-ニトロフェニルリン酸溶液(KPL, Gaithersburg, MD, USA)を 100 µl 添加し,室温・30 分間反応させた。反応 終了後 30 分以内に,マイクロプレートリーダーARVO MX(PerkinElmer, Waltham, MA, USA)を用い,波長 405 nm における各ウェルの吸光度を測定 し,固相化一本鎖 DNA と二本鎖を形成した標識一本鎖 DNA の相対量を算出 することで,ゲノム DNA の相同性とした。相同性算出の際,各ウェルの吸光 度の値から,陰性コントロールの吸光度の値を減じ,値の補正を行った。また, 固相化一本鎖DNA と標識一本鎖 DNA が同一株由来であるウェルの値を 100% として,各ウェルの相対値(%)を算出した。 9 パルスフィールドゲル電気泳動法(PFGE)によるM. plutonius 株および M. plutonius 様菌株の遺伝的背景の解析
11
M. plutonius および M. plutonius 様菌の遺伝的背景の解析は,各株のゲノ ムDNA を制限酵素で消化し,生じた DNA 断片の大きさと数を PFGE で比較 することによって行った。
PFGE に用いる制限酵素で消化された各供試菌株のゲノム DNA を含有する サンプルプラグは下記のように作製した。先ず,KSBHI 培地で嫌気培養した 菌を集菌し,Tris-saline buffer[10 mM Tris-HCl (pH 8.0), 1 M NaCl]で洗 浄後,EDTA-sarcosine buffer (6 mM Tris-HCl, 1 mM NaCl, 100 mM EDTA, 1% sodium N-lauroylsarcosine; pH 7.6)に浮遊した。調整した菌浮遊液を等量 の1.0% SeaKem Gold agarose(Lonza Rockland, Rockland, ME, USA)含有 EDTA-sarcosine buffer と混合し,0.7-mm sample plug caster(Bio-Rad Laboratories, Hercules, CA, USA)内で固化させることによって各菌株を寒天 に包埋したサンプルプラグを作製した。次に,サンプルプラグを lysis buffer [0.5 M EDTA (pH 8.0), 2.5 mg/ml lysozyme, 10 U/ml mutanolysin] で 37℃・3 時間,プロテナーゼ K 液[0.5 M EDTA (pH 8.0), 1% sodium N-lauroylsarcosine, 1 mg/ml proteinase K]で 50℃・18 時間インキュベート することによってプラグ中の菌を溶菌し,ゲノム DNA を露出させた。サンプ ルプラグは1 mM Pefabloc SC(Roche Applied Science, Basel, Switzerland) 含有TE で 2 回,50℃・30 分間処理することによってプロテナーゼ K を失活 させ,さらに,TE で 4℃・3 回洗浄して Pefabloc SC を除去した。その後,各 プラグをメーカーが推奨する制限酵素緩衝液で4℃・30 分間インキュベートし, プラグ内のゲノム DNA を 20 U の制限酵素SmaⅠ(タカラバイオ(株))で 30℃・18 時間消化した。
制限酵素処理の結果生じたDNA 断片は,1.0% SeaKem Gold agarose gel およびCHEF-DR Ⅱ system(Bio-Rad Laboratories)を用いた PFGE により
12
解析した。泳動条件は15℃・20 時間,電界強度 6 V/cm,電場交替間隔 2.9-17.3 秒とし,泳動用緩衝液は50 µM チオ尿素含有 0.5 × TBE buffer(44.5 mM Tris, 44.5 mM boric acid, 1 mM EDTA; pH 8.0)を用いた。泳動終了後,ゲル をエチジウムブロマイドで染色し,UV 光下で撮影後,画像を TIFF ファイル として記録した。可視化された DNA 断片パターンは BioNumerics software version 5.1(Applied Maths, Sint-Martens-Latem, Belgium)を用いた補正お よび目視による確認と手動による修正の後,解析した。類似度の計算は DICE 法により行い,クラスター分析はUPGMA(Unweighted Pair Group Method with Aristmetic mean)を用いて,行った。なお,各断片の位置補正のための Tolerance 値は 1%に,各検体間の移動差補正のための Optimization 値は 1% に設定した。
10 アクセッション番号
決定したM. plutonius および M. plutonius 様菌の 16S rRNA 遺伝子配列は, DDBJ/GenBank/EMBL に登録および公開している。アクセッション番号は表 1-1 に示すとおりである。
13 結果 1 M. plutonius および M. plutonius 様菌の性状 (1)培養性状 M. plutonius の性状に関する過去の報告(1, 4, 9, 11)と同様に,本研究 に供試した,基準株を含む全ての M. plutonius 株は,特定の培養条件下で しか良好な発育を示さなかった(表1-3)。すなわち,リン酸二水素カリウム を添加した培地(Medium 1, KBHI 培地, KSBHI 培地)では,これらの株 は嫌気条件下で良く発育したが ,リン酸 二水素 カリウム無添加の培地 (Medium 2, SBHI 培地, BHI 培地)やリン酸二水素カリウムの替わりにリ ン酸二水素ナトリウムを添加した培地(Medium 6)では,全く発育しない か,または弱い発育を示すのみであった。基準株はリン酸二水素カリウム以 外のカリウム塩を加えた培地(Medium 3, Medium 4, Medium 5)でも嫌気 条件下で発育が認められたが,野外分離株は同培養条件では全く発育しない か,またはわずかな発育が認められるのみであった。さらに,好気条件下で は,供試した全ての M. plutonius 株は今回用いたいずれの培地にも発育し なかった。M. plutonius 分離培養に汎用される Medium 1 および KSBHI 培地には共通して可溶性デンプンが含まれているが,M. plutonius 株の発育 に必須な成分ではなかった。また,可溶性デンプンは M. plutonius の発育 を促進させることが報告されているが(8),嫌気培養時では明らかな発育促 進効果を示さず,リン酸二水素カリウム存在下の炭酸ガス培養時にのみ,発 育を促進させる傾向を示した。 一方,発育にリン酸二水素カリウムを要求するM. plutonius 株とは異なり, M. plutonius 様菌株は幅広い培養条件下で良好な発育が認められた(表 1-3)。
14
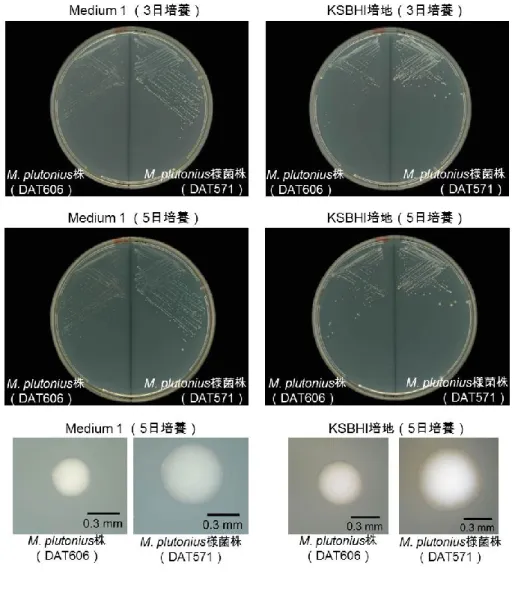
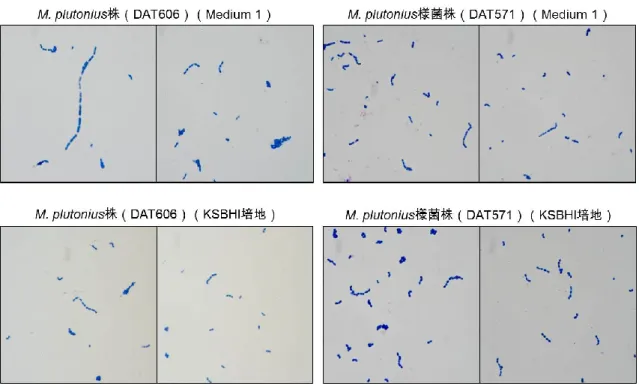
これらの株は嫌気条件下では,カリウム塩添加培地(Medium 1, Medium 3, Medium 4, Medium 5, KBHI 培地, KSBHI 培地)だけでなく,カリウム塩 無添加培地(Medium 2, SBHI 培地, BHI 培地)やリン酸二水素ナトリウム 添加培地(Medium 6)でも良く発育した。さらに,好気培養や炭酸ガス培 養でも,様々な培地で発育が認められた。培地への可溶性デンプン添加は, M. plutonius 様菌株の発育にも必須ではなく,嫌気培養時には発育促進効果 も示さなかったが,リン酸二水素カリウム存在下では炭酸ガスおよび好気培 養時にその発育を促進させる傾向を示した。 (2)生化学性状 培養性状と同様に,生化学性状もM. plutonius 株と M. plutonius 様菌株 では異なっていた(表 1-4)。M. plutonius 株は,グルコース,フルクトー ス,D-マンノースから酸を産生したが,その他の糖からの酸産生は認められ なかった。一方,M. plutonius 様菌株は全ての株で,これら3つの糖に加え て,L-アラビノース,D-セロビオース,サリシンからも酸を産生し,さらに, DAT557 株および DAT565 株の 2 株では,ラクトースからの弱い酸産生も 認められた。加えて,全ての M. plutonius 様菌株がエスクリン加水分解能 およびβ-グルコシダーゼ活性を有していた。また,ホスファターゼおよび β-ガラクトシダーゼ活性は,M. plutonius 株に比較して,M. plutonius 様 菌株で明らかに強かった。 (3)コロニー形状およびグラム染色像 培養性状・生化学性状が大きく異なるにもかかわらず,M. plutonius 株と M. plutonius 様菌株は形態学的には類似していた。リン酸二水素カリウム添
15 加培地(Medium 1, KSBHI 培地)で嫌気培養した場合,両者とも 37℃で数 日培養後,目視可能なコロニーを形成した。コロニーは正円,白色,半透明 で,表面は滑らかであり,大きさは直径1 mm 程度であった(図 1-2)。菌体 のグラム染色像はグラム陽性紡錘形球菌を示し,様々な長さで連鎖すること があり,時に桿菌様の形態を示した(図1-3)。以上のように両菌株は,形態 によって区別することは不可能であったが,M. plutonius 株に比較して, M. plutonius 様菌株はやや大きいコロニーを形成する傾向を示した。また, M. plutonius 株は KSBHI 培地上で,時にコロニーのサイズが大きくなった 変異体を生じ,一見コンタミネーションが起きたと思われるような様相を示 すことがあった。一方,この様な変異体は M. plutonius 様菌株では観察さ れなかった。 2 M. plutonius 様菌の分類学的位置づけ M. plutonius 様菌株の分類学的位置づけを決定するために,全供試株の 16S rRNA 遺伝子の部分塩基配列(1,510 bp)を決定した。M. plutonius および M. plutonius 様菌の全ての野外分離株の配列は,M. plutonius 基準株の配列と 99.8 %以上の高い相同性を示し,M. plutonius 様菌株は M. plutonius と同一 菌種であることが強く示唆された。この結果を確認するために,16S rRNA 遺 伝子配列に基づいて選択したM. plutonius および M. plutonius 様菌の代表株 をDNA-DNA ハイブリダイゼーションに供試した。表 1-5 に示すように,供試 した全ての M. plutonius 様菌株は,基準株を含む全ての M. plutonius 株と 80 %以上のゲノム DNA の相同性を示した。一方,E. faecalis 基準株とは約 10 % の相同性しか示さなかった。ある2 つの菌株のゲノム DNA が,DNA-DNA ハ イブリダイゼーションにおいて70%以上の相同性を示した場合,その 2 株は同
16 一菌種に属することが菌種同定の基準として提唱されている(42)。従って, M. plutonius 様菌株は,分類学的には M. plutonius と同一菌種であり,日本 を含む世界各国で分離される典型的な性状の M. plutonius 株(M. plutonius 典型株)とは異なる,非典型的な性状を示す株(M. plutonius 非典型株)であ ることが確認された。 興味深いことに,基準株を含む全ての典型株の 16S rRNA 遺伝子配列は, DAT585 株を除き,その分離地域や分離年にかかわらず同一であった。一方, 全ての非典型株の配列は,基準株の配列と 2 ~ 3 塩基の違いが認められ,同 一地域・同一年代で分離された非典型株同士でも数塩基の配列の違いが確認さ れた。 3 M. plutonius の遺伝学的多様性 PFGE の結果,基準株を含む典型株は互いに類似した DNA 断片の泳動パタ ーン(PFEG プロファイル)を示し(相同性 83.77 %以上),単一のクラスター を形成した(図1-4)。イギリスで分離された基準株と日本で分離された 2 株は 全く同一のプロファイルを示し,パラグアイで分離されたDAT569 株と日本分 離株9 株は前者とは異なる同一のプロファイルを示した。一方,非典型株も互 いに類似したPFEG プロファイルを示した(相同性 78.81 %以上)が,典型株 とは別の単一のクラスターを形成した。典型株と非典型株のPFGE プロファイ ルは明らかに異なり,これらは培養性状や生化学性状だけでなく,遺伝的背景 も異なることが明らかとなった。
17 考察 日本で分離されたBHI 培地にも発育可能なM. plutonius 様菌株は,これま で知られている典型的な性状を示すM. plutonius 株とは培養・生化学性状が大 きく異なるものの,16S rRNA 遺伝子解析および DNA-DNA ハイブリダイゼー ションの結果から,分類学的にはM. plutonius と同一菌種であることが証明さ れた。また,全ての典型的性状を示すM. plutonius 株(典型株)は分離地域・ 国や年代が異なるにもかかわらず,類似のPFGE プロファイルを示し,ほぼ同 一の 16S rRNA 遺伝子配列を有していたのに対し,非典型的性状を示す M. plutonius 株(非典型株)は典型株とは明らかに異なる PFGE プロファイルを 示した。典型株のうち基準株は,継代を繰り返すことによりごく低頻度ではあ るが,カリウム塩無添加培地でも発育可能な変異株が出現することが報告され ている(39)。しかし,この様な変異株の出現は他の野外分離典型株ではほと んど観察されておらず,類似した遺伝的背景を持つ株の集団はほぼ同一の性状 を示している。このことは,典型株と非典型株間における性状の違いは,診断 施設や研究施設における人工培養の過程で起こった遺伝子変異の結果生じたも のではなく,野外に浸潤している株の性状をそのまま反映していることを示唆 している。これまでM. plutonius は株間の多様性が少ない均一な株から構成さ れる菌種であると考えられてきた。しかし,本研究の結果から,本菌は実は多 様性のある菌種であり,表現型および遺伝的背景が異なる少なくとも2グルー プの株,すなわち,基準株と類似した性状および遺伝的背景を有する典型株と 基準株とは明らかに異なる性状および遺伝的背景を有する非典型株が存在する ことが明らかとなった。 性状が大きく異なる典型株と非典型株を比較解析することで,未だ明らかに
18
なっていない M. plutonius の生理機能や発病機構を解明する手がかりが得ら れる可能性がある。実際,既に典型株と非典型株の全ゲノム配列は決定されて おり(34, 35),Takamatsu ら(39)は,このゲノム情報と彼らが新しく開発 したM. plutonius 用プラスミドベクターを利用して,典型株発育におけるカリ ウム要求性はNa+/H+ antiporter 遺伝子および cation-transporting ATPase 遺
伝子における変異が関係することを明らかにしている。EFB や M. plutonius に関して得られている知見は,その発見から一世紀以上が経過しているにもか かわらず,あまりにも少ない。しかし,本研究による非典型株の発見を契機と して,今後のM. plutonius 研究が進展することが期待される。 本研究で用いた非典型株の分離年は1991~2010 年と幅広く,また,分離地 域も日本各地に広がっている。1980 年代の日本の EFB に関する報告例を精査 すると,典型株とは異なる性状を示すM. plutonius 株が記載されているものが 認められ(17, 44),これには国内の初発例に関する報告(17)も含まれる。佐 賀県および岐阜県で分離されたこれらの株は,硝酸塩還元能やアラビノース・ トレハロース等の糖分解能において今回用いた非典型株とも相違が見られるが, その発育にはカリウム塩添加が必要でないと考えられ,また,好気条件下でも 発育可能である等,非典型株と類似した性状も示している(17, 44)。DNA-DNA ハイブリダイゼーション等の遺伝学的検査は行われていないため,これらの菌 株が真にM. plutonius であるかは定かではない。しかし,これらの株は EFB の典型的な臨床症状を示すミツバチ幼虫から分離されており,現在,把握して いる以上に古くから非典型株は日本国内に存在している可能性がある。 典型株とは異なる性状を示すM. plutonius 株は,少数であるが日本以外でも 報告されている(1, 6, 8, 11)。1962 年には,リン酸カリウム無添加培地でも発 育可能なイギリス分離株が報告されており(11),1984 年と 1993 年の文献に
19 は,リン酸カリウム無添加培地に加えて,リン酸ナトリウム添加培地でも発育 可能なブラジル分離株の記載が確認出来る(1, 8)。また,インドのトウヨウミ ツバチ(Apis cerana)からは,市販の一般的な酵母抽出液を添加した培地上で 高濃度炭酸ガスにより発育が阻止されるM. plutonius 株も分離されている(6)。 これらの報告において用いられた基礎培地や各性状の試験項目および試験方法 は,本研究におけるものとは一致しないため,日本の非典型株との単純な比較 は困難である。しかし,ブラジル分離株は,好気条件下ではカリウム塩添加培 地上であっても発育しない点,スクロースやマルトース等を分解可能である点 等は異なるものの,その培養・生化学性状は日本の非典型株のものと似通って いる(1, 8)。さらに最近,改良型 Multilocus Sequence Typing(MLST)法に よる M. plutonius の新しい型別法が開発され,世界各地で分離される M. plutonius 株は,基準株を含む typical group とそれ以外の atypical group に大 別されることが報告された(24)。興味深いことに,この atypical group には, 日本の非典型株(DAT561 株)の他に,イギリス分離株(1 株),アメリカ分離 株(2 株),ブラジル分離株(1 株),オランダ分離株(1 株)が分類されている。 MLST 法による型別はあくまで遺伝学的な型別であり,atypical group に分類 される株の詳細な性状はブラジル分離株以外では明らかでない。しかし,これ らの報告は,日本の非典型株のように非典型的な性状を示すM. plutonius 株は, これまで見過ごされていただけで,実は世界中に広く分布している可能性を示 唆している。atypical group に分類される世界中の分離株を解析することで, M. plutonius の多様性がより詳細に明らかになるかもしれない。
20 表1-1 供試株 分離国・地域 分離年 16S rRNA遺伝子配列の アクセッション番号 基準株(ATCC 35311) イギリス 1982年以前 AB614100 DAT558 日本 関東地方 2008年 AB614071 DAT559 日本 関東地方 2009年 AB614072 DAT560 日本 関東地方 2009年 AB614073 DAT563 日本 関東地方 2009年 AB614076 DAT566 日本 関東地方 2008年 AB614078 DAT569 パラグアイ 1993または1994年 AB614094 DAT580 日本 中国地方 1990年 AB614089 DAT581 日本 中国地方 1991年 AB614090 DAT582 日本 中国地方 1992年 AB614091 DAT583 日本 中国地方 1992年 AB614092 DAT584 日本 中国地方 1995年 AB614093 DAT585 日本 関東地方 1988年 AB614095 DAT606 日本 関東地方 2010年 AB614098 DAT351 日本 中部地方 2004年 AB614069 DAT352 日本 中部地方 2006年 AB614068 DAT557 日本 関東地方 2005年 AB614070 DAT561 日本 関東地方 2009年 AB614074 DAT562 日本 関東地方 2009年 AB614075 DAT565 日本 関東地方 2005年 AB614077 DAT567 日本 東北地方 2009年 AB614079 DAT571 日本 中国地方 1991年 AB614080 DAT572 日本 中国地方 1991年 AB614081 DAT573 日本 中国地方 1991年 AB614082 DAT574 日本 中国地方 1991年 AB614083 DAT575 日本 中国地方 1991年 AB614084 DAT576 日本 中国地方 1991年 AB614085 DAT577 日本 中国地方 1991年 AB614086 DAT578 日本 中国地方 1991年 AB614087 DAT579 日本 中部地方 1997年以前 AB614088 DAT604 日本 関東地方 2003年 AB614096 DAT605 日本 関東地方 2006年 AB614097 DAT607 日本 関東地方 2010年 AB614099 M. plutonius 株名 M. plutonius 様菌 全ての株がEFBの臨床症状を示すセイヨウミツバチ幼虫からの分離株であり,M. plutonius特異的PCR (23)陽性であった。
21 表 1-2 各培地 の 組成 M ed iu m 1 a M ed iu m 2 b M ed iu m 3 a M ed iu m 4 c M ed iu m 5 c M ed iu m 6 d K S B H I 培地 K B H I 培地 S B H I 培地 B H I 培地 糖分解試験 用培地 a 寒天 (Dif co) 15 15 15 15 15 15 15 15 15 15 2 イーストエキス (Dif co) 10 10 10 10 10 10 -10 グルコース (W A K O ) 10 10 10 10 10 10 -可溶性デンプン (Dif co) 10 10 10 10 10 10 10 -10 -10 KH 2 PO 4 e (W A K O ) 13 .6 -20 .4 20 .4 -13 .6 K C l e (W A K O ) -7. 4 -K3 (C6 H5 O7 ) · H2 O e (W A K O ) -10 .8 -K H C O3 e (W A K O ) -10 -N aH 2 PO 4 · 2H 2 O e (W A K O ) -15 .6 -糖 f (W A K O ) -10 g ブロモクレゾールパープル (W A K O ) -0. 03 B ra in H ea rt I n fu si on (D if co) -37 37 37 37 -単位 : g /L c: H C lを用い, p H 6. 6に調整した。 d : N aO H を用い, p H 6. 6 に調整した。 e:カリウム塩およびナトリウム塩は終濃度 0. 03 3 M ( M ed iu m 4 ), 0. 1 M ( M ed iu m 1 , 3 , 5 および 6)または 0. 15 M ( K S B H Iおよび K B H I)となるよう加えた。 f: D-セロビオース,ラクトース, D-ラフィノースまたは D-キシロース g:基礎培地をオートクレーブ滅菌した後,無菌的に糖を加えた。 M ed iu m 1 ~ M ed iu m 6 および糖分解試験用培地は 115 ℃・ 10 分間オートクレーブ滅菌した。その他の培地は 121 ℃・ 15 分間オートクレーブ滅菌した。 a : KOH を用い, pH 6 .6 に調整した。 b : K O H /N aO H ( 1: 1 )混合液を用い, pH 6. 6 に調整した。
22 表1-3 供試株の培養性状 基準株 M. plutonius 株 M. plutonius様菌株 Medium 1 嫌気培養 +v + + 5%炭酸ガス培養 +w – または +w + 好気培養 - – + Medium 2 嫌気培養 +w – または +w + または +v 5%炭酸ガス培養 – – + 好気培養 – – + Medium 3 嫌気培養 + +w + 5%炭酸ガス培養 +w – + 好気培養 – – + Medium 4 嫌気培養 +w – + 5%炭酸ガス培養 – – + 好気培養 – – + Medium 5 嫌気培養 +w – + 5%炭酸ガス培養 – – – 好気培養 – – – Medium 6 嫌気培養 – – +w または + 5%炭酸ガス培養 – – – 好気培養 – – – KSBHI培地 嫌気培養 + + + 5%炭酸ガス培養 – –, +w または + + 好気培養 – – + KBHI培地 嫌気培養 + + + 5%炭酸ガス培養 – – または +w +w または + 好気培養 – – +w または + SBHI培地 嫌気培養 – – + 5%炭酸ガス培養 – – +w または + 好気培養 – – – BHI培地 嫌気培養 – – +v 5%炭酸ガス培養 – – +w または + 好気培養 – – – 結果a 培地 培養条件 a:KSBHI培地で嫌気培養した基準株の発育の程度を+と評価した。この発育の程度と比較 して,より旺盛な発育を+V,より微弱な発育を+Wと評価した。全く発育が認められないも の,および極めて微弱な発育しか認められないものは-と評価した。
23 表1-4 供試株の生化学性状 基準株 M. plutonius 株 M. plutonius様菌株 - - - - - - - - - - - - - - - - - -または± -または±a ± + ± -または± + - - + - - + グルコース + + + フルクトース + + + D-マンノース + + + L-アラビノース - - + D-セロビオース - - + サリシン - - + ラクトース - - -または± その他b - - - ウレアーゼ ゼラチン液化 ホスファターゼ β-ガラクトシダーゼ β-グルコシダーゼ エスクリン加水分解 a:検査回毎にわずかに異なる反応を示した。 b:マンニトール, スクロース, マルトース, D-キシロース, グリセリン, D-メレジトース, D-ラフィノース, D-ソルビトール, L-ラムノースおよびD-トレハロース 結果 糖からの酸産生 項目 カタラーゼ オキシダーゼ 硝酸塩還元 インドール試験
24 表 1-5 D N A-D N A ハイブリダイゼーション結果 M . p lu ton iu s 基準株 D A T 35 1 D A T 56 1 D A T 56 5 D A T 58 5 D A T 60 6 D A T 60 7 E . f aec al is 基準株 M . p lu ton iu s 基準株 10 0. 0 a 99 .5 94 .2 85 .5 95 .7 97 .7 94 .1 8. 9 D A T 35 1 b 96 .4 10 0. 0 97 .8 86 .4 89 .6 97 .1 94 .4 9. 2 D A T 56 1 b 94 .4 98 .9 10 0. 0 88 .7 82 .1 95 .2 95 .9 8. 5 D A T 56 5 b 10 1. 5 10 9. 2 10 1. 0 10 0. 0 94 .3 99 .9 99 .9 10 .6 D A T 58 5 c 10 1. 7 98 .8 95 .8 85 .9 10 0. 0 10 0. 0 91 .4 9. 0 D A T 60 6 c 10 7. 0 10 7. 2 10 0. 3 87 .9 97 .3 10 0. 0 93 .8 9. 7 D A T 60 7 b 10 4. 9 11 2. 1 10 6. 6 96 .3 96 .4 98 .8 10 0. 0 9. 0 E . f aec al is 基準株 11 .6 12 .9 10 .9 10 .1 11 .2 13 .4 11 .9 10 0. 0 a:相同性( % )。 b: M . p lu to ni us 様菌株 c: M . p lu to ni us 株 ハイブリダイズさせた D N A 固相化 D N A 相同 性は ,固相 化 D N A と 二本 鎖を 形成 し た 標識 D N A に 結合 し た ア ル カ リ フ ォ ス フ ァ タ ーゼ の 活性 を 測り 算出 し て い る 。酵素 反応 の 強さ を 基に し て 算出 し た 値で あ り ,実際 の 塩基 配列 を 比較 し て 算出 し た 値で は な い た め ,時に 100% を 超え る こ と が あ る 。
25
図1-1 M. plutonius 特異的 PCR 法の結果
M. plutonius株およびM. plutonius様菌株から抽出したDNA を用いてM. plutonius
特異的PCR(23)を行った結果,全ての供試株で特異的増幅が認められた。陽性コントロ
26
図1-2 コロニー形状
いずれの株も37℃・嫌気培養を行った。M. plutonius株およびM. plutonius様菌株の
27
図1-3 グラム染色像
37℃・5 日間嫌気培養した供試株をグラム染色した。染色像の観察・撮影は光学顕微鏡
28
図1-4 PFGE デンドログラム
培養・生化学性状が異なるM. plutonius典型株および非典型株は,それぞれ別のクラス
29
第二章
30 序論 ある感染症を引き起こす病原体を特定するためには,感染実験による病原性 の証明が必要である。M. plutonius がまだ培養出来ていなかった時代には,P. alvei や E. faecalis といった腐蛆から良く分離されてくる菌やハチへの病原性 があると考えられるウイルス等の EFB 発症への関与を感染実験により否定す ることで,EFB の一次病原体はM. plutonius であろうことを推測していた(43)。 その後,Bailey(4)が M. plutonius(典型株)の培養に成功してから,M. plutonius そのものを用いて様々な方法で感染実験が試みられてきた(3, 5, 12, 30)。例えば,Bailey は 2 回継代培養したM. plutonius と‘Bacterium eurydice (正式な菌種名ではなく,本菌の分類学的位置は未確定)’を混合培養した菌液 を用いて,ミツバチ幼虫にEFB を実験的に再現させた(3)。‘B. eurydice’は EFB の二次感染菌であり,本菌単独では EFB を引き起こさないことも確認さ れている(3)。また,McKee らは腐蛆から直接抽出したM. plutonius を用い ることでミツバチ幼虫への実験感染を成功させた(30)。これらの長年にわた る様々な研究の蓄積によって,現在では,M. plutonius が EFB の原因菌であ ることが広く認識されている。しかし,人工培養したM. plutonius のみを用い てEFB を再現する試みはほとんど成功しておらず,M. plutonius(典型株)は 人工培地で継代培養を繰り返すことで急速に病原性を失うと考えられている (3, 30)。 第一章の実験により,日本で分離された既知のM. plutonius の性状とは異な る性状を示すM. plutonius 様菌は,分類学的に M. plutonius と同一菌種(M. plutonius 非典型株)であることが証明された。しかし,第一章で用いた非典 型株は全てEFB の症状を示すミツバチ幼虫から分離された株ではあるものの,
31 非典型株が実際にミツバチ幼虫に病原性を発揮し,EFB の原因となり得るか否 かについては未だ実験的に確認されていない。また,非典型株も典型株と同様 に,人工培養条件下で急速に病原性が失われる可能性もある。そこで,本章で は,セイヨウミツバチ幼虫を用いて人工培養菌による感染実験を試み,非典型 株のミツバチ幼虫に対する病原性の有無を検証した。
32 材料および方法 1 供試動物と飼育方法 実験に用いる孵化後 24 時間以内のセイヨウミツバチ幼虫は,独立行政法人 農業・食品産業技術総合研究機構畜産草地研究所で飼育されている健康蜂群か ら下記の方法に従って採取し,Aupinel らの方法に準じて飼育した(2)。 先ず,女王蜂を巣箱の中に設置したケージ内で隔離飼育し,産卵させた。産 卵から3 日後に,ケージ内で孵化した幼虫を面相筆で拾い上げ,35 mm 径細胞 培養用プレートに薄く敷いた約300 µl の人工餌(50% ローヤルゼリー,37% 滅 菌蒸留水,6% D-グルコース,6% D-フルクトース,1% イーストエキス)上 に浮遊させた。幼虫を飼育するプレートは,温度 35℃,相対湿度 90%のイン キュベーター内で維持した。 なお,当該研究所の蜂群においてEFB の発生はこれまで認められておらず, M. plutonius を摂取させなかった対照群の幼虫からは M. plutonius は分離さ れていない。 2 供試株と感染方法
感染実験には,典型株3 株(DAT583 株,DAT585 株,DAT606 株)および 非典型株3 株(DAT351 株,DAT561 株,DAT573 株)の計 6 株を用いた。こ れら菌株は実験に用いるまで人工培地で約6 回継代培養されていた。感染用人 工餌作製のために,供試株をKSBHI 培地で 35℃・1 週間嫌気培養し,回収し た菌を滅菌生理食塩水で2 回洗浄後,約 1×107 cfu/ml となるように滅菌生理
食塩水に浮遊した。この菌液と人工餌を 1:1 の割合で混ぜ,約 5×106 cfu/ml
33 採取した幼虫を実験群6 群・対照群 1 群(各群 24 匹以上)に分け,各実験 群には1 匹あたり 10 µl の感染用人工餌を 24 時間経口摂取させた。一方,対照 群には滅菌生理食塩水にて2 倍希釈した人工餌を 10 µl 与えた。実験 2 日目以 降は毎日,全ての幼虫を新しい人工餌上に移して飼育した。実験は最大5 日間 行った。 3 感染幼虫および人工餌からのM. plutonius 分離 上記方法によって M. plutonius の感染が成立することを確認するため, DAT606 株(典型株)摂取群および DAT561 株(非典型株)摂取群を用い,各 感染幼虫および人工餌残渣からKSBHI 培地によりM. plutonius の分離培養を 行った。幼虫を生存させたまま感染菌量を測定することは不可能なため,生存 曲線作製のための群とは別に幼虫に菌を摂取させ,感染が十分に成立したと考 えられる実験4 日目に無作為に幼虫および人工餌残渣を抽出し,実験に供試し た。両群の感染菌量の差の有無については,P 値が 0.05 未満になる場合を有意 とし,t 検定により判定した。 4 生存曲線の作製 生存曲線作製のために,全ての実験群および対照群について,実験期間中毎 日,実体顕微鏡下で幼虫を観察し,その生死を確認した。幼虫の死亡は呼吸の 消失,虫体の不透明性および黄色化により判定した。実験は 5 日目まで行い, 各群の生存曲線間の差の有無については,P 値が 0.05 未満になる場合を有意と し,ログランク検定により判定した。
34 結果 1 ミツバチ幼虫へのM. plutonius 感染の確認と臨床症状 感染用人工餌摂取開始4 日後において,供試した全ての DAT606 株(典型株) 摂取幼虫およびDAT561 株(非典型株)摂取幼虫からM. plutonius が分離さ れ,1 匹あたりそれぞれ 7.32×107±6.26×107 cfu(n = 8)および 6.94×106 ±8.24×106 cfu(n = 13)の菌が回収された。一方,人工餌残渣の培養では, 2 検体からM. plutonius のコロニーが1つだけ発育したものの,他の検体から は全くM. plutonius は分離されなかった。このことから,幼虫から分離された M. plutonius は,虫体表層に付着した人工餌に残存していた M. plutonius の クロスコンタミネーションによるものではなく,幼虫体内に感染していた M. plutonius の数を反映したものであり,いずれのタイプの株を用いた場合でも 本試験法で幼虫へのM. plutonius 感染が成立することを確認した。 しかし,実験 4 日目の時点での感染菌量は,DAT561 株摂取群に比べ, DAT606 株摂取群で有意に多かった(t 検定,P < 0.05)ものの,DAT606 摂取 群では,取り扱いの過程で物理的に傷ついた個体を除き,全ての幼虫が良好に 発育し,臨床的な異常は全く認めなかった。一方,DAT561 株摂取群では全て の幼虫の成長が完全に停止し,既に死亡していると考えられる個体も存在した (図2-1(A))。 2 生存率および生存曲線 過去の報告(3, 30)と同様に,継代培養した典型株はミツバチ幼虫に対する 病原性を失っていた。典型株を摂取させた幼虫では,実験終了まで良好に発育 し,ほとんどの幼虫が生存した。実験終了時の生存率は 83.3~94.3%であり,
35 対照群(91.4%)と同等であった(ログランク検定, P = 1)(表 2-1 および図 2-1(B))。 一方,非典型株を摂取させた幼虫は摂取後 2~3 日で成長が停止し,呼吸緩 慢になり,実験終了時までにほとんどの幼虫が死亡した。実験終了時の生存率 は 5.7~29.2%であり,対照群,典型株摂取群と比較して有意に低かった(ロ グランク検定, P < 0.05)(表 2-1 および図 2-1(B))。
36 考察 M. plutonius(典型株)は人工培地上で急速に病原性を失うと考えられており, 人工培養菌を用いた場合,EFB を再現することは非常に困難であることが報告 されている(3, 30)。実際,McKee らの感染実験では,人工培養菌を用いた場 合,1.1 × 109 organisms/ml のM. plutonius を与え続けたにもかかわらず, EFB を再現することは出来なかった(30)。過去の報告と同様に,本研究にお いても人工培養したM. plutonius 典型株では EFB を再現することは出来なか った。一方,驚くべき事に,M. plutonius 非典型株は,人工培地での継代を典 型株とほぼ同じ回数繰り返しているにもかかわらず,また,実験 4 日目の感染 菌量は発症しなかった典型株摂取群より有意に少ないにもかかわらず,摂取 5 日以内にほとんどのミツバチ幼虫を死亡させた。死亡幼虫はEFB の典型的な臨 床症状を示し,また,菌分離を試みた全ての発症幼虫から非典型株が分離され たことから,非典型株は人工培地で継代を繰り返しても病原性を維持すること が確認されたと同時に,確かにEFB の原因になり得ることが証明された。 EFB の診断は病性鑑定指針(33)に基づき,各都道府県の家畜保健衛生所に て実施されている。現行の病性鑑定指針(33)には典型株に関しての情報しか 記載されていないことから,非典型株が分離される事例については EFB とし て確定診断されず,農林水産省への発生報告もほとんど行われてきていない可 能性が高い。また,発生報告もアメリカ腐蛆病(American foulbrood; AFB) との区別なく「腐蛆病」とだけ報告・集計されるため,法定伝染病であるにも かかわらず国内における EFB の発生状況は良く把握できていないのが実状で ある。EFB の正確な診断や発生状況把握のためには,病性鑑定指針の記載内容 見直しと発生報告におけるAFB との区別が必要とされる。
37 人工培養したM. plutonius 典型株では EFB を再現することは出来なかった が,本研究で用いた全ての典型株はEFB の臨床症状を示すミツバチ幼虫から分 離されている。また,菌株提供者から,典型株感染幼虫の症状と非典型株感染 幼虫の症状は見分けがつかなかったと報告を受けている。従って,今回供試し た典型株も野外ではEFB を引き起こす能力があったと考えられる。実際,一旦, 病原性が低下したM. plutonius 株でも,ミツバチ幼虫の腸内を数回通過するこ とで病原性が次第に復帰していくことが確認されており(5),典型株の病原性 関連遺伝子の発現は人工培養時には抑制されている可能性が考えられる。また, 二次感染菌である‘B. eurydice’や P. alvei を M. plutonius とミツバチ幼虫に 共感染させることにより,EFB を再現,もしくは死亡率が上昇したという報告 もある(3, 21)。このことから,M. plutonius 典型株が病原性関連遺伝子を発現 し,十分な病原性を発揮するためには,P. alvei のような二次感染菌の存在も必 要とされるのかもしれない。一方,非典型株では人工培養時でも,また二次感 染菌の非存在下でも,ミツバチ幼虫に対して強力な病原性を発揮したことから, 典型株と非典型株の両者では病原性制御メカニズムは異なることが示唆された。 し かし, 非典 型株の 詳細な発 病メ カニズ ムはまだ 解明 されて いない。M. plutonius は通常,ミツバチ幼虫の腸内でのみ増殖することから,増殖した M. plutonius が栄養成分を消費してしまい,幼虫を飢餓状態にすることで死に至ら せるというEFB 発病機序が Bailey により提唱された(7)。しかし,McKee ら の実験では,腐蛆から直接抽出したM. plutonius を実験感染させた幼虫は,十 分量の食餌を与えているにもかかわらず発症し,死亡した(30)。さらに,本研 究においても,非典型株摂取群は十分な餌を与えられていたにもかかわらず, 感染後数日以内に成長が停止した。対照的に,典型株摂取群は濃厚感染してい るにもかかわらず,良好に発育した。これらの結果は,Bailey が提唱した餓死
38 説とは別の機序で幼虫が死亡していることを示唆している。 興味深いことに,Giersch らは,幼虫 1 匹あたりの摂取菌数が本研究で行った 感染実験の約200 倍量に相当するものの,腐蛆から分離培養したM. plutonius 株を用いてミツバチ幼虫への実験感染を成功させており,感染 169 時間後の致 死率は80%以上であった(21)。同グループが過去に行った感染実験(30)では 成功しなかった人工培養菌での EFB の再現を,何故,2010 年の報告では再現 できているのかは,Giersch ら(21)の文献内では全く考察されていない。2010 年の報告で用いたM. plutonius 株は人工培地上で 1 回しか継代されていないた め,ミツバチ幼虫に摂取された時,まだ病原性関連遺伝子を発現していたのか もしれない。または,詳細な性状は記載されていないが,我々の用いた非典型 株同様,人工培地上でも病原性を維持可能な特殊な株であった可能性も考えら れる。 これまで,人工培地上での継代を繰り返すことで急速に病原性を失うという M. plutonius 典型株の性状が EFB の発病メカニズム解明の大きな妨げとなって いた。しかし,人工培養条件下でも病原性を維持し続け,実験室内において容 易に感染実験を成立させることが出来る非典型株の発見は,今後のEFB 発病メ カニズム解明の大きな足掛かりになると考えられた。特に,人工培養した典型 株と非典型株の遺伝子発現プロファイルを比較解析することで,EFB 発病メカ ニズム解明のヒントとなる有用な情報が得られる可能性が高いと考えられる。
39 表2-1 実験終了時における各群の生存率 生存率(%) 対照群 91.4 典型株摂取群 DAT583 87.5 DAT585 83.3 DAT606 94.3 非典型株摂取群 DAT351 8.3 DAT561 5.7 DAT573 29.2
40 図2-1 感染幼虫および生存曲線 (A) 実験開始 4 日目の幼虫(5 日齢)。矢頭は幼虫を示す。撮影のために,幼虫を新しい 24 穴細胞培養用プレートに移し,十分量の人工餌上に浮遊させた。 (B) 対照群(灰色線),典型株(DAT606 株)摂取群(黒色線)および非典型株(DAT561 株)摂取群(黒色点線)の生存曲線。他の典型株摂取群,非典型株摂取群もそれぞれ DAT606 株摂取群,DAT561 株摂取群と同様な生存曲線を示した。
41
第三章
M. plutonius
典型株/非典型株検出・識別用
42 序論 M. plutonius には典型株と非典型株の2つのタイプの株が存在し,さらに, 感染実験の結果から,両者は異なるメカニズムで病原性を制御している可能性 が明らかとなった。従って,より正確に EFB を診断し,より有効な対策を考 える上で,野外の各EFB 症例の原因となっているM. plutonius 株のタイプを 識別し,EFB の疫学情報を蓄積することが重要となってくる。 EFB の診断はまず特徴的な臨床症状を確認することから始まる。しかし, EFB 以外の原因でも幼虫が EFB と類似の症状を示すことがあり,EFB であっ ても,P. larvae によるアメリカ腐蛆病(American foulbrood; AFB)と類似の 症状である有蓋幼虫の死亡が引き起こされることもある。そのため,EFB の確 定診断にはM. plutonius の検出が必要とされる。 M. plutonius を検出するための方法としては第一に分離培養法が挙げられる。 特に,実際に病気を引き起こしている株そのものを手に入れる必要がある場合 には,分離培養法は必須の方法である。しかし,M. plutonius のように複雑な 条件で培養する必要がある菌では,その取扱いが難しく,培養操作が煩雑であ り,分離頻度に検査担当者の熟練度が影響する。さらに,M. plutonius は発育 速度が比較的遅い菌種であることから,分離・同定には2週間以上の時間を必 要とする。従って,診断の簡便性・正確性・迅速性の点において,分離培養法 には様々な欠点が存在する。 近年,病原体を簡便・迅速に検出・同定するための方法として PCR 法が広 く活用されている。EFB においても,16S rRNA 遺伝子を標的とした M. plutonius 特異的 PCR 法が報告されている(14, 23, 29)。しかし,これらの方 法は典型株と非典型株の両タイプを検出出来るものの,これらを識別すること
43
は出来ない(図1-1,第三章 結果 2 各 PCR 法の特異性)。そこで,本章では, 2 組のプライマーセットを同時に用いることにより,1つの反応で典型株と非 典型株の両者を検出し,それぞれを識別できる新たな M. plutonius 特異的 Duplex PCR 法を開発した。さらに,本 Duplex PCR 法の EFB 診断における 有用性について既報のPCR 法および分離培養法との比較検証を行った。
44 材料および方法 1 供試株 M. plutonius 特異的 Duplex PCR 法の開発とその特異性の検証のために, M. plutonius 典型株 24 株・非典型株 26 株,M. plutonius 以外の 19 菌種 23 株および健康なミツバチ幼虫の腸由来菌13 株を用いた(表 3-1)。 M. plutonius 株の内,33 株は第1章で供試した株(表 1-1)と同一であり, 残り 17 株は以下に示す方法で腐蛆から新たに分離された。健康なミツバチ幼 虫腸由来菌株は下記の方法で分離された。これら以外の菌株は,埼玉県中央家 畜保健衛生所および独立行政法人農業・食品産業技術総合研究機構動物衛生研 究所に保存されていた株を用いた。 (1)腐蛆からのM. plutonius 分離 EFB の臨床症状を示した腐蛆を乳剤化後,KSBHI 培地に塗抹し,37℃・ 3~4 日間嫌気培養を行った。M. plutonius を疑うコロニーを KSBHI 培地 に継代し,単離した後,形態学的特徴,培養・生化学性状,M. plutonius 特 異的PCR 法(23)により,M. plutonius 典型株または非典型株と同定した。 (2)健康幼虫からの細菌分離
健康なセイヨウミツバチおよびニホンミツバチ(Apis cerana japonica) 幼虫は独立行政法人農業・食品産業技術総合研究機構畜産草地研究所で飼育 されている蜂群から集められた。それぞれ 10 匹以上の幼虫を分離培養に用 いた。幼虫の体表面を 70%エタノールで消毒後,滅菌蒸留水で洗浄した。 腸を外科的に摘出し,その内容物を100-500 µl の PBS または BHI(Difco) ブロスと混合して,乳剤を作製した。乳剤をBHI 寒天培地,5%馬血液加 BHI 寒天培地,Lactobacilli MRS 寒天培地(Difco)および卵黄加 GAM 寒天培
45 地(日水製薬(株))に塗抹し,いずれも35~37℃で,BHI 培地は好気培養, 5%馬血液加 BHI 培地は 5%炭酸ガス培養,これら以外の培地は嫌気培養し, 発育してきたコロニーを初代分離と同じ培養条件で数回継代培養した。グラ ム染色,カタラーゼ試験およびオキシダーゼ試験により分離菌の一次鑑別を 行った後,良く分離した菌株について後述の方法で DNA を抽出し,16S rRNA 遺伝子の部分塩基配列を前述の方法(第一章 材料と方法 7 16S rRNA 遺 伝 子 解 析 ) に よ り 決 定 し た 。 EzTaxon-e (http://eztaxon-e.ezbiocloud.net)(26)を用いて,決定した配列とデータベ ースに登録されている各菌種の基準株の16S rRNA 遺伝子配列を比較し,分 離した各菌株の種,属,科,目または綱を決定した。 2 DNA 抽出 各菌株をそれぞれの至適条件で培養した後,前述の方法(第一章 材料および 方 法 6 ゲ ノ ム DNA の 抽 出 ) ま た は InstaGene Matrix ( Bio-Rad Laboratories)により DNA を抽出した。
3 特異遺伝子領域の抽出およびプライマー設計
Duplex PCR の標 的遺伝 子の 選択 のた め ,in silico MolecularCloning Genomics Edition(インシリコバイオロジー(株), 横浜, 日本)を用いてM. plutonius 基準株(典型株: DDBJ/EMBL/GenBank アクセッション番号 AP012200)および DAT561 株(非典型株:アクセッション番号 AP012282) の全ゲノム配列を比較解析し,典型株および非典型株それぞれに特異的な遺伝 子領域を抽出した。その結果に基づき標的遺伝子候補を選択し,電気泳動をし た際に容易に識別可能なサイズの特異的 PCR 産物が得られるプライマーセッ