九州大学学術情報リポジトリ
Kyushu University Institutional Repository
ラット大脳皮質錐体ニューロンの興奮性シナプス後 電流に対するアドレナリン受容体の活性化を介する 修飾
小嶋, 允郎
九州大学大学院歯学府口腔保健推進学講座口腔予防科学分野 / 小嶋歯科医院
https://doi.org/10.15017/14241
出版情報:Kyushu University, 2008, 博士(歯学), 課程博士 バージョン:
権利関係:
ラット大脳皮質錐体ニューロンの興奮性シナプス後電流に対する アドレナリン受容体の活性化を介する修飾
The mechanisms of adrenergic modulation of excitatory postsynaptic currents in pyramidal neurons of rat cerebral cortex
2008年 小嶋允郎
九州大学大学院歯学研究院口腔保健推進学講座 口腔予防科学分野
指導教員 :山下 喜久 教授 研究指導者:越川 憲明 教授
本論文は,細胞内記録による実験結果を以下に挙げる対象論文に加 えてまとめたものである。
対象論文
Masayuki Kobayashi, Masao Kojima, Yuko Koyanagi, Kazunori Adachi, Kazuyuki Imamura, Noriaki Koshikawa (2008) Pre- and postsynaptic modulation of glutamatergic synaptic transmission by activation of α1- and β-adrenoceptors in layer V pyramidal neurons of rat cerebral cortex, Synapse, in press
略号
ACSF 人工脳脊髄液
AMPA α-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid ATP adenosine triphosphate
BAPTA 1,2-bis(2-aminophenoxy)ethane-N,N,N’,N’-tetraacetic acid cAMP cyclic-3’,5’-adenosine monophosphate
CV coefficient of variation
D-APV D-(-)-2-amino-5-phosphonopentanoic acid DNQX 6,7-dinitroquinoxaline-2,3-dione
eEPSCs 興奮性シナプス後電流
eEPSPs 興奮性シナプス後電位
GABAA γ-aminobutyric acid type A GTP guanosine triphosphate
HEPES N-(2-hydroxyethyl)piperazine-N’-2-ethanesulfonic acid IP3 イノシトール-3-リン酸
K-S test Kolmogorov-Smirnoff test
mEPSCs miniature EPSCs,微小興奮性シナプス後電流
NMDA N-methyl-D-aspartate PKA プロテインキナーゼA
PKC プロテインキナーゼC
PPR paired-pulse ratio
QX-314 N-(2,6-dimethylphenylcarbamoylmethyl)triethylammonium bromide
sEPSCs 自発性興奮性シナプス後電流
SN比 信号―ノイズ比
要旨
アドレナリン受容体アゴニストは,受容体サブタイプ固有の働きを介して大脳 皮質における興奮伝播に対して相反する作用を及ぼすことが,光学計測による研 究から明らかとなっている(Kobayashi et al., 2000)。しかし,これらの作用がどの ような神経基盤によって実現されているかは,不明な点が多い。そこで本研究で は,脳スライス標本を用いた電気生理学的手法によって,このメカニズムの一端 を明らかにすることを目的とした。
はじめに,興奮性ニューロンである錐体ニューロンから細胞内記録を行い,電 気刺激によって誘発される興奮性シナプス後電位(eEPSPs)に対するノルアドレ ナリンと α1アドレナリン受容体アゴニストである phenylephrine,β アドレナリン 受容体アゴニストであるisoproterenolの効果を調べた。その結果,ノルアドレナリ ンは,記録ニューロンがⅡ/ⅢあるいはⅤ層のいずれにあっても,eEPSPsの振幅を 減少させ,同様の効果が phenylephrine の灌流投与によって観察された。一方,
isoproterenol は,Ⅴ層に存在する錐体ニューロンで観察される eEPSPs の振幅を有
意に増大させたが,Ⅱ/Ⅲ層の錐体ニューロンから記録される eEPSPs の振幅には 影響を与えなかった。
次に,大脳皮質Ⅴ層に存在する錐体ニューロンを対象にホールセル・パッチク ランプ記録を行い,グルタミン酸作動性シナプスを介した興奮伝達に対して,
phenylephrineとisoproterenolがどのように作用し,その作用点がどこに存在するか を調べた。
頻回刺激(33 Hz,10連矩形波)によって誘発されるα-amino-3-hydroxy-5-methyl-4- isoxazole-propionic acid(AMPA)受容体を介した興奮性シナプス後電流(eEPSCs)
に対して,phenylephrineはその振幅を抑制した。また,1発目のeEPSCsにおける 振幅の変動係数(coefficient of variation; 以下,CV)および1発目の eEPSCsに対 する2発目のeEPSCsの振幅比(paired-pulse ratio; 以下,PPR)は,phenylephrine によって変化しなかった。一方,isoproterenolは,2発目から10発目のeEPSCsと
比較して 1発目の eEPSCs の振幅を顕著に増大させ,その結果としてPPR を減少
させた。また,isoproterenol は,1 発目の eEPSCs の CV を減少させた。以上の結 果は,isoproterenol がシナプス前終末における glutamate の放出確率を上昇させる 可能性が高いのに対して,phenylephrine はシナプス前終末に影響を及ぼす可能性 が低いことを示唆するものである。
その仮説を検証するために,微小興奮性シナプス後電流(miniature EPSCs; 以下,
mEPSCs)に対するphenylephrineとisoproterenolの作用を調べた。Phenylephrineは,
mEPSCs の頻度を変化させることなく振幅を抑制し,isoproterenol は,mEPSCs の
振幅に影響を及ぼすことなく mEPSCs の頻度を増加させた。これらの結果は,
phenylephrine がシナプス後膜のグルタミン酸受容体に作用して mEPSCs の振幅を
減少させるのに対して,isoproterenol はシナプス前終末における glutamate の放出 確率を上昇させる仮説を支持するものである。
最後に,phenylephrine と isoproterenol のシナプス後膜に存在するグルタミン酸 受容体に対する効果を調べた。その結果,isoproterenolの投与は,glutamate,AMPA
および N-methyl-D-aspartate(NMDA)のパフ投与によって惹起される内向き電流
を変化させないのに対して,phenylephrine は,これらいずれの内向き電流も抑制 した。
以上の結果は,phenylephrineがシナプス後膜上に存在するAMPA 受容体および NMDA受容体を介する興奮性シナプス伝達を減弱させるのに対して,isoproterenol はシナプス後膜上のグルタミン酸受容体には影響を及ぼさず,シナプス前終末か
らのglutamate放出を促進することを示すものである。これら二つの異なった興奮
性シナプス伝達の調節メカニズムは,大脳皮質のノルアドレナリンの要となる機 能と考えられている信号―ノイズ(SN)比の向上に寄与している可能性がある。
緒言
青斑核に細胞体が存在するノルアドレナリン作動性ニューロンは,大脳皮質に投 射し(Morrison et al., 1978),シナプス形成(Blue and Parnavelas, 1982)やニューロ ンの興奮性(McCormick and Prince, 1988; Nicoll et al., 1990; Wang and McCormick, 1993),可塑性(Kasamatsu and Pettigrew, 1976; Kirkwood et al., 1999)を含む生理的 機能の調節に重要な役割を果たしている。大脳皮質のニューロンは,それぞれ違 うGタンパクに共役するα1,α2,βアドレナリン受容体を発現している(Nicholas et al., 1996)。α1アドレナリン受容体は,Gq/11タンパクと共役しホスホリパーゼC を活性化することによって,イノシトール-3-リン酸(IP3)とジアシルグリセロー ルを産生するのに対して,α2とβアドレナリン受容体は,それぞれ Gi,Gsと共役 している。Giタンパクは,cyclic-3’,5’-adenosine monophosphate(cAMP)を産生す るアデニル酸シクラーゼを阻害する一方,Gsタンパクは,アデニル酸シクラーゼ を活性化し,細胞内のcAMPを増加させる。過去の光学計測法を用いた報告では,
α1 アドレナリン受容体が白質の電気刺激により惹起される興奮伝達を抑制するの に対して,α2および β アドレナリン受容体アゴニストは興奮伝達を増大させるこ とが示されている(Kobayashi et al., 2000)。さらに,微小ガラス電極を使用した興 奮性シナプス後電位(EPSPs)の細胞内記録によって,α1アドレナリン受容体アゴ ニストは,プロテインキナーゼ C(PKC)の活性化を介して EPSPs を抑制し,β アドレナリン受容体アゴニストは,cAMP/プロテインキナーゼ A(PKA)のカス ケードを活性化することによって EPSPs を増大させることが明らかとなっている
(Kobayashi, 2007)。このように,ノルアドレナリンは,その受容体サブタイプによ
って効果が異なっていると考えられる。
大脳皮質におけるアドレナリン作動性ニューロンによる多様な調節機構を理解 するには,各アドレナリン受容体サブタイプのシナプス伝達に関する詳細な情報 が不可欠である。過去,シナプス前膜に存在する α アドレナリン受容体のほとん どは,α2 受容体と考えられてきた(Starke et al., 1989; Kamisaki et al., 1991)。
Lehmenkuhler ら(1991)は,α1受容体のグルタミン酸受容体に対する作用を報告
し,α1受容体アゴニストは,NMDAのイオノフォレーシスによる投与に対する応
答を減弱させること,すなわちシナプス後膜に存在するNMDA受容体を抑制する ことを明らかにした。また,Kobayashi ら(2008)は,シナプス後膜に存在してい ると想定される α1受容体の活性化により,大脳皮質錐体ニューロンの連続発火の 頻度が増加して,興奮性が高まることを報告している。その一方で最近,ラット 大脳皮質視覚野における α1アドレナリン受容体の分布が免疫組織化学的,免疫電 子顕微鏡手法により調べられ,α1 受容体がシナプス後膜のみならず,興奮性ニュ ーロンの軸索終末にも一部分布していることが明らかにされた(Nakadate et al.,
2006)。したがって,興奮性シナプス伝達に対するα1受容体の作用がシナプス前膜
を介したものか否かは,依然として不明な点が残されている。
シナプス前膜と後膜に存在する β アドレナリン受容体は,ともにニューロンの 興奮性を調節する。βアドレナリン受容体のアゴニストは,シナプス前終末におけ るglutamate放出を促進する(Herrero and Sanchez-Prieto, 1996; Wang et al., 2002)。
HuangとHsu(2006)は,シナプス前膜のβアドレナリン受容体の活性化により前
頭野でのglutamate放出を促進することを報告した。また,いくつかの細胞内記録
の研究により,シナプス後膜の β アドレナリン受容体の活性化は,細胞内通電に よる連続発火の頻度を増加させることが明らかとなっている(Foehring et al., 1989;
Dodt et al., 1991; Nowicky et al., 1992; Kobayashi et al., 2008)。しかし,シナプス伝 達に対するシナプス後膜上の β アドレナリン受容体の役割については,ほとんど 分かっていない。
本研究では,錐体ニューロンで観察される興奮性シナプス伝達に対する α1と β アドレナリン受容体の効果が,錐体ニューロンが存在する層の間で異なるか否か を明らかにすること,そして,その効果がシナプス前膜と後膜いずれに作用して 発生しているのかを明らかにすることを目的とした。実験の概要は,以下のとお りである。ラット大脳皮質の視覚野から作製した脳スライス標本を用いて,まず,
(1)大脳皮質Ⅱ/Ⅲ層およびⅤ層に存在する錐体ニューロンから微小ガラス管電極 を用いて細胞内記録を行い,ノルアドレナリンとα1およびβ アドレナリン受容体 アゴニストの投与によるeEPSPsに対する影響を調べた。さらに,ホールセル・パ ッチクランプ記録を行い,(2)電気刺激によって誘発されるグルタミン酸受容体
を介したeEPSCsに対する作用を検索した。また,(3)電位依存性ナトリウムチャ
ネル阻害薬であるtetrodotoxin存在下で観察される mEPSCsを記録し,その振幅と 頻度に対する効果を調べた。そして,(4)glutamateならびにAMPA,NMDAを記 録ニューロンにパフ投与して観察される内向き電流に対する効果を検索した。
方法 脳スライス標本の作製
Sprague-Dawley系ラット(3-5週齢)をpentobarbital(75 mg/kg, 腹腔内投与)に て深麻酔し,氷冷した人工脳脊髄液(ACSF)中に脳を取り出した。脳を冷却後,
大脳皮質後頭葉部分を切り出し,スライサー(Zero1, 堂阪 EM)に固定した。ス ライサーにて厚さ350-400 mの大脳皮質視覚野のスライス標本を切り出し,室温 のもとACSF中で培養した。スライス標本作製用ACSFの組成は次の通りである:
230 mM Sucrose, 2.5 mM KCl, 10 mM MgSO4, 1.25 mM NaH2PO4, 26 mM NaHCO3, 2.5 mM CaCl2, 10 mM D-glucose。また,スライス標本培養用および記録用ACSF(pH 7.35-7.40)の組成は以下の通りである:126 mM NaCl, 3 mM KCl, 2.0 mM MgSO4, 1.25 mM NaH2PO4, 26 mM NaHCO3, 2.0 mM CaCl2, 10 mM D-glucose。いずれのACSF も,常時,混合ガス(酸素95%, 二酸化炭素5%)で通気した。
細胞内記録
脳スライス標本は,サブマージ型の保持チャンバー内で培養後,混合ガス(酸
素95%, 二酸化炭素5%)を常時通気している記録用ACSFを連続的に灌流するイ
ンターフェース型記録用(Haas型)チャンバー(1.0-1.5 ml/min)へ移した。直径 1.2-1.5 mmのボロシリケート製ガラス管(Harvard Apparatus)をプラー(P-87およ
びP-97, Sutter Instruments)にて,加熱,牽引し,微小ガラス電極を作製した。電
極内液には2 M potassium acetateを用いた。電極内液を充填した微小ガラス電極は,
三次元水圧式マニピュレータ(MHW-3, ナリシゲ)に固定した電極ホルダーに挿 入した。電極の先端の位置を実体顕微鏡下で確認した後,マニピュレータにて電 極をスライス標本内に挿入した。細胞内への電極の刺入は,電極を発振させるこ と(Buzz)で行い,挿入後-0.5 nA程度の弱い直流電流を通電し,静止膜電位より やや過分極した状態で5分から10分維持した。その後,徐々に通電量を減少させ,
およそ10-15分後に直流通電量を0 nAにした。eEPSPsを誘発するために,白質と
Ⅵ層の境界に刺入した双極のタングステン電極を介して 100 S のパルス通電を 0.1 Hzで行った。刺激強さは,閾値の1.2-1.5倍(30-200 A)に設定した。膜電位
および電流は,細胞内記録用増幅器(Axoclamp 200B, Axon Instruments)にて増幅 し,A-Dコンバータ(Digidata 1322A, Axon Instruments)でデジタル化した後,オ シ ロ ス コ ー プ で 観 察 し つ つ ハ ー ド デ ィ ス ク に 取 り 込 ん だ (Clampex 9, Axon Instruments)。
ホールセル・パッチクランプ記録
脳スライス標本は,サブマージ型の保持チャンバー内で培養した後,記録用 ACSFを灌流しているサブマージ型記録用チャンバー(1.0-1.5 ml/min)へ移した。
ノマルスキー微分干渉装置を備えた正立顕微鏡(倍率40倍, Olympus BX51)に装 着した,赤外線高感度カメラ(C3077-78, Hamamatsu Photonics)を通して脳スライ ス標本を観察し,Ⅴ層錐体ニューロンからホールセル・パッチクランプ記録を行 った。アンプ(Axoclamp 200B, Axon Instruments)によって膜電位と電流を増幅し,
A-Dコンバータ(Digidata 1322A, Axon Instruments)でデジタル化しハードディス クに保存した(Clampex 9, Axon Instruments)。電圧固定記録のためのピペット内液 の組成は以下の通りである: 120 mM cesium gluconate, 20 mM biocytin, 10 mM N-(2-hydroxyethyl)piperazine-N’-2-ethanesulfonic acid(HEPES), 8 mM NaCl, 5 mM N-(2,6-dimethylphenylcarbamoylmethyl)triethylammonium bromide(QX-314), 2 mM
magnesium adenosine triphosphate(ATP), 0.3 mM sodium guanosine triphosphate(GTP), 0.1 mM 1,2-bis(2-aminophenoxy)ethane-N,N,N’,N’-tetraacetic acid(BAPTA)。ピペット
内液はpH 7.3で300 mOsmに調整した。液間電位差は10 mVで,全ての膜電位は
これに応じて修正した。パッチ電極(2-4 M)はFlaming-Brown micropipette puller
(P-97, Sutter Instruments)を用いて,ボロシリケート製ガラス管(直径1.5 mm, Harvard Apparatus)から作製した。
記録時の温度は30-31℃に設定した。本研究では,シール抵抗は 5 G以上で,
データはアクセス抵抗が 8-20 Mのもののみ採用した。アクセス抵抗が,記録中
に 20%以上変化したものは,データとして採用しなかった。容量補正は 70%とし
た。γ-aminobutyric acid type A(GABAA)受容体アンタゴニストであるbicuculline methiodide(10-20 M)とD-(-)-2-amino-5-phosphonopentanoic acid(D-APV, 50 M)
の灌流投与下で,eEPSCsを記録した。保持電位は-70 mVとした。eEPSCsを誘発
するために,Ⅳ層とⅤ層の境に置いた単極のタングステン電極を介して,100 S の10連パルスを33.3 Hzで与えた。刺激強さは,閾値の1.2-1.5倍の強さに設定(4-20
A)した。mEPSCsは,tetrodotoxin(1 M, Wako)とbicuculline methiodide(20 M) を投与した状態で,保持電位-70mVにて記録した。Glutamate,AMPAおよびNMDA をパフ投与するため,パッチ電極と同様の形状をもつガラスピペットをⅤ層に設 置し,ピペットの先端と記録ニューロンの細胞体との間の距離を約50 mに保持 した。Gluamate,AMPAおよびNMDAは,ガラスピペット内に充填し,PicoPump
(PV830, World Precision Instruments)を使用してピペットの内圧を高める(15 psi,
20 ms)ことにより,その先端から放出させた。Gluamate,AMPA と NMDA が誘
発する電流は, tetrodotoxin(1 M)と bicuculline methiodide(20 M)が入っ た記録用ACSF投与下で記録した。保持電位は,それぞれ-70 mV,-70 mV,-30 mV とした。パフ投与の間隔は0.03 Hzとした。
本研究では,bicuculline methiodide,D-APVおよびtetrodotoxinに加えて,以下 の薬剤を灌流液に直接投与した: 6,7-dinitroquinoxaline-2,3-dione(DNQX, 30 M), isoproterenol, phenylephrine, prazosin(100 M), propranolol(100 M)。
データ解析
直流通電量を0 nAにした時の膜電位を静止膜電位とした。細胞の電気生理学的 な基本的性質を調べるために,脱分極性の短矩形波を通電して活動電位を発生さ せた(Fig. 1A)。活動電位の振幅は,静止膜電位からピークまでの値とした。その 持続時間は,振幅の1/2になる電位において活動電位の脱分極相および再分極相が 交叉する時点を求め,その時間差とした(1/2 持続時間)。EPSCs と,glutamate,
AMPA お よ び NMDA に よ る 内 向 き 電 流 の 解 析 に は ,pClamp ver. 9(Axon Instruments)とWindetecta(Ulrich and Huguenard, 1996)を用い,回帰直線の解析 には,Origin(ver. 6, OriginLab)を用いた。
統計
ニューロンの電気生理学的特性に対する灌流投与した薬物が及ぼす影響につい ては,対応のあるt検定(paired t-test)により有意差の有無を判定した。mEPSCs
の解析における累積度数分布曲線については,Kolmogorov-Smirnoff test(K-S test)
により検定した。有意水準は,それぞれ 5%,0.1%とした。統計解析には,SPSS
(ver. 12, SPSS)を用いた。
結果
第Ⅱ/Ⅲ層およびⅤ層錐体ニューロンから記録される EPSPs に対するアドレナリ ン受容体アゴニストの効果
大脳皮質Ⅱ/ⅢおよびⅤ層のニューロンから微小ガラス電極による細胞内記録 を行った。なお記録したニューロンのうち,静止膜電位が-60 mV以下,活動電位
の振幅が70 mV以上のニューロンを解析の対象とした(n = 92)。記録したニュー
ロンの電気生理学的特性をTable 1に示す。記録したニューロンは,短矩形パルス
(5 ms)によって単一(regular spiking)もしくは2発以上(bursting)の活動電位 を発生し,活動電位および連続発火の特徴から,グルタミン酸作動性錐体ニュー ロンであると推定された(Fig. 1A)。白質とⅥ層の境界に刺入した双極のタングス テン電極から電気刺激を行うことによって誘発される興奮性シナプス後電位
(eEPSPs)の電気生理学的特性をTable 1に示す。細胞内記録実験において,regular
spiking ニューロンと bursting ニューロン間で,アドレナリン受容体アゴニストの
効果に有意な差はなかったため,共に錐体ニューロンとして扱った。
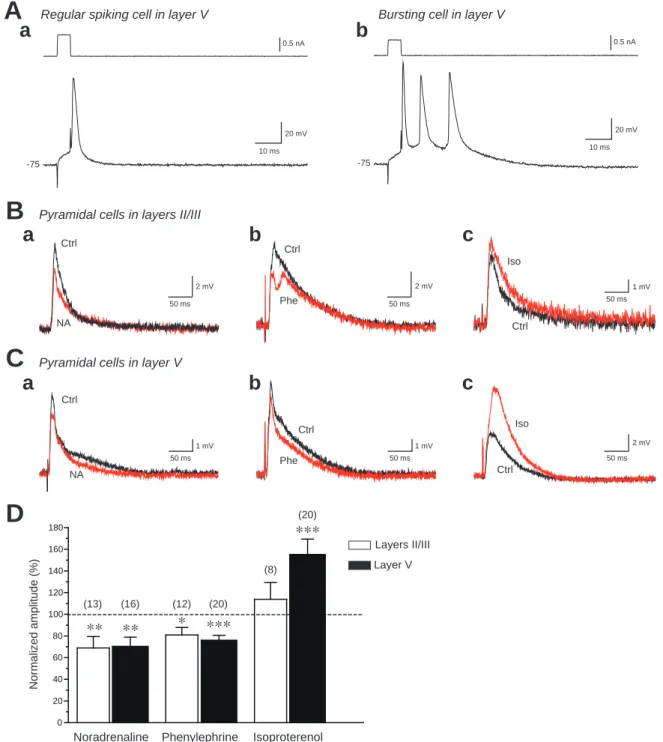
ノルアドレナリン(100 M)では,錐体ニューロンがⅡ/ⅢおよびⅤ層のいずれ にあっても,eEPSPsの振幅を減少させた(P < 0.01, paired t-test; Fig. 1)。同様の効 果は,α1アドレナリン受容体のアゴニストであるphenylephrine(100 M)の灌流 投与でも観察された(II/III: P < 0.05, V: P < 0.001, paired t-test; Fig. 1B, C and D)。第
Ⅱ/Ⅲ層とⅤ層に存在する錐体ニューロン間で振幅減少の程度に有意な差は認め られなかった。一方,βアドレナリン受容体アゴニストであるisoproterenol(100M) 投与では,Ⅴ層に存在する錐体ニューロンで観察されるeEPSPsの振幅を有意に増 大させたが(P < 0.001, paired t-test),Ⅱ/Ⅲ層の錐体ニューロンから記録される
eEPSPsの振幅には影響を与えなかった。
以下の実験では,Ⅴ層の錐体ニューロンに焦点を絞り,phenylephrine および
isoproterenol の glutamate を介する興奮性シナプス伝達に対する作用と,その神経 基盤を明らかにするために,ホールセル・パッチクランプ記録を行った。
eEPSPsのPPRおよびCVは細胞外Ca2+濃度に依存する
グルタミン酸作動性のシナプス伝達における α1およびβアドレナリン受容体ア ゴニストの効果を調べるために,記録ニューロン近傍に刺激電極を設置し,10連 続刺激を33 Hzで与えた。eEPSPsは,GABAA受容体アンタゴニストであるbicuculline methiodide(10-20 M)とNMDA受容体アンタゴニストであるD-APV(50 M)存 在下で記録した。eEPSCsは,AMPA受容体アンタゴニストであるDNQX(30 M)
の投与によってほぼ完全に消失したことから,AMPA受容体を介した電流と考えら れた(n = 6; Fig. 2A)。
PPRは,シナプス前膜における伝達物質の放出確率を反映し,シナプス後膜の反 応には影響されないと考えられている(Zucker, 1989; Manabe et al.,1993)。そこで,
本標本でも同様の仮説が成立するか否かを検証するために,細胞外Ca2+濃度を変化 させることにより,様々な放出確率の条件下でeEPSCsを記録した。すなわち,細 胞外Ca2+を標準濃度である2 mMから,3 mM Ca2+と0.5 M Ca2+に変化させたACSF を灌流投与することにより,それぞれ放出確率の高い条件と低い条件を作り出し た。なお,細胞外の2価の陽イオン濃度は,細胞膜の安定性に関わるため,細胞外 Mg2+濃度を調整することによって,細胞外の2価の陽イオン濃度を4 mMに維持し た。Mg2+,Ca2+とも2 mMの条件下をコントロールとし,-70 mVでeEPSCsの記録を 行ったところ,振幅は127.4 ± 21.8 pAであった(n = 41)。2発目の振幅(Fig. 2B, A2)
を1発目の振幅(Fig. 2B, A1)で割って算出されるPPRは,1.36 ± 0.11であった。32.7%
のニューロンは,paired-pulse depressionを示し,55.1%のニューロンにおいて paired-pulse facilitationが観察された。0.5 mMのCa2+を灌流投与すると,1発目と2発 目のeEPSCsの振幅は減少し,PPR値は有意に増加した(P < 0.05, paired t-test; Fig. 2C, E)。一方,3 mMのCa2+投与下では,eEPSCsの振幅は増強され,PPRは有意に減少 した(P < 0.001, paired t-test)。
CVは,PPR同様にシナプス後膜における変化には影響されず,シナプス前膜に おける放出確率に影響されると考えられている(Bekkers and Stevens, 1990)。そこ
で次に,3 mM,および0.5 mM Ca2+を含むACSFの投与下で,1発目の振幅のCVを 調べた。PPRと同様,CVは,0.5 mM Ca2を含むACSFを投与した場合増加し(P < 0.01, paired t-test),3mM Ca2+を含むACSFを投与した場合には減少した(P < 0.001, paired t-test; Fig. 2F)。これらの結果により,1発目と2発目のEPSCsの振幅によって導き出 されるPPRとCVは,シナプス前終末における放出確率に感受性があることが示唆 された。
eEPSPsに対するアドレナリン作動性アゴニストの効果
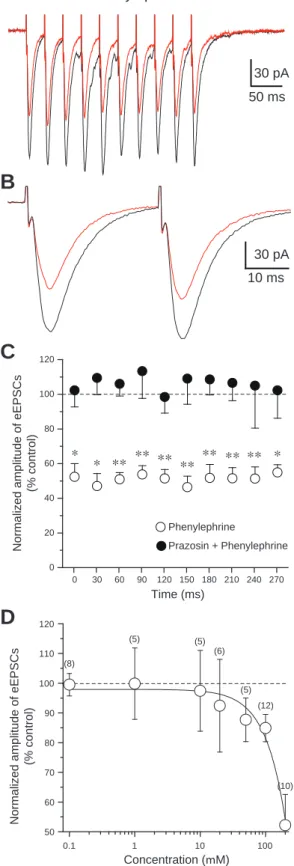
Phenylephrine(200 M)の灌流投与は,1発目のeEPSCsの振幅を47.8 ± 7.6%に減 少させた(n = 10, P < 0.05, paired t-test)。eEPSCsの振幅に対するphenylephrineの効 果の典型例をFig. 3A, Bに示した。Phenylephrineは,1発目のeEPSCsだけでなく2-10 発目のeEPSCsも同程度に抑制した。Phenylephrineによって抑制されたeEPSCsの平 均値,すなわち1発目から10発目までの抑制率を比較したものをFig. 3Cに示した。
PhenylephrineによるeEPSCsの抑制は,α1アドレナリン受容体アンタゴニストである prazosin(100 M)を前投与することによって阻害された(n = 6)。Phenylephrine 投与下での1発目のeEPSCsのCV(0.39 ± 0.06, n = 10)は,コントロールにおける CV(0.39 ± 0.06, n = 10)と有意な差はなかった。Phenylephrine投与時のeEPSCsの1 発目と2発目のPPR(125.2 ± 17.3%)も,コントロール(133.8 ± 14.6%)とほとん ど変わらなかった。1発目に対する3発目から10発目までのPPRについても,
phenylephrine投与時とコントロールにおいてほとんど変わらなかった。
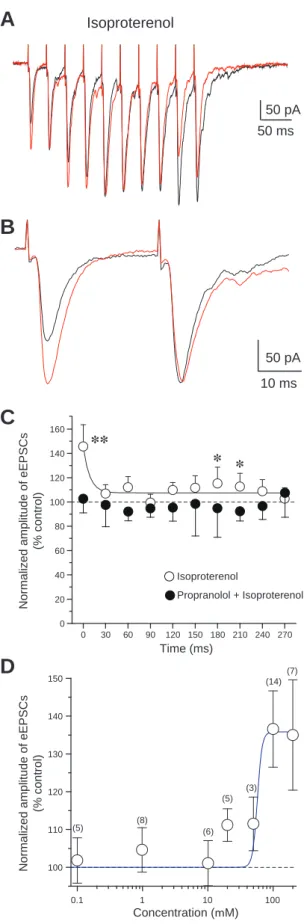
Isoproterenol(100 M)の灌流投与は,1発目のeEPSCsを有意に増大させた(45.7
± 17.9%, n = 13, P < 0.01, paired t-test)。興味深いことにisoproterenolは,2発目から 10発目までのeEPSCsも増大させる傾向にあるが,その程度は1発目のeEPSCsより も小さかった(Fig. 4A-C)。βアドレナリン受容体のアンタゴニストである propranolol(100 M)により,isoproterenolの促進効果は消失した(n = 7, Fig. 4C)。 Isoproterenolは,eEPSCsを増大させるとともに,CVを0.38 ± 0.06から0.29 ± 0.05へ 有意に減少させた(n = 13, P < 0.05, paired t-test)。さらにisoproterenolは,1発目に 対する2発目のeEPSCsのPPRを174.2 ± 27.0%から122.9 ± 10.7%へ減少させ(n = 13, P < 0.05, paired t-test),1発目に対する3発目から10発目までのPPRも減少させた。
PhenylephrineによるeEPSCsの抑制と,isoproterenolによるeEPSCsの増大は,濃度依 存的であった(Figs. 3D, 4D)。以下の実験におけるphenylephrineとisoproterenolの濃 度は,用量依存曲線と過去の報告(Kobayashi, 2007)を参考に,電気生理学的特性 や一定の効果を誘発するのに十分な濃度であると考えられる100 Mに設定した。
アドレナリン受容体アゴニストによるmEPSCsの頻度および振幅の調節
本研究では,phenylephrineおよびisoprotelenolによるEPSCsの調節が,シナプス前 膜あるいはシナプス後膜のいずれで行われているかを決定するために,電位依存 性 ナ ト リ ウ ム チ ャ ネ ル 阻 害 薬 で あ るtetrodotoxin(1 M), お よ びbicuculline
methiodide(20 M)を灌流投与した条件下で,Ⅴ層錐体ニューロンからmEPSCs
の記録を行った。保持電位は-70 mVに設定した。なおmEPSCsは,DNQX(30 M)
とD-APV(50 M)の投与により消失した(data not shown)。
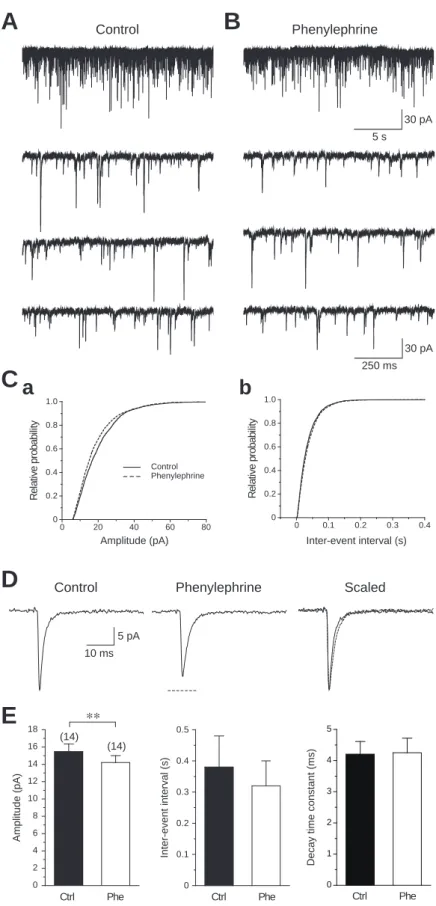
mEPSCsの発生頻度におけるphenylephrineの効果を調べるため,14ニューロンを
解析した(Fig. 5)。コントロールとphenylephrine(100M)投与時の典型的なmEPSCs の記録をFig. 5A, Bに示す。累積度数分布曲線が示すように,phenylephrineは,
mEPSCsが発生する時間間隔に影響を及ぼさなかった(K-S test; Fig. 5C)。また phenyleprineは,その平均値にも有意な変化を及ぼさなかった(コントロール: 0.38
± 0.10 s; phenylephrine: 0.32 ± 0.08 s, n = 14, paired t-test; Fig. 5E)。一方,
phenylephrineは,mEPSCsの振幅の累積度数分布曲線を左方へ移動させ(P < 0.0001, K-S test; Fig. 5C),平均値は減少した(コントロール: 15.5 ± 0.9 pA; phenylephrine:
14.2 ± 0.8 pA, n = 14, P < 0.05, paired t-test; Fig. 5D, E)。Phenylephrineは,立ち上がり 時間(コントロール: 1.0 ± 0.06 ms; phenylephrine: 1.1 ± 0.08 ms, n = 14),1/2持続時 間(コントロール: 5.5 ± 0.6 ms; phenylephrine: 5.4 ± 0.7 ms, n = 14),減衰時間(コン トロール: 4.2 ± 0.4 ms; phenylephrine: 4.3 ± 0.5 ms, n = 11; Fig. 5E)において何の変化 も生じさせなかった。
IsoproterenolのmEPSCsに及ぼす効果をFig. 6に示す。コントロールとisoproterenol
(100 M)投与時の典型的なmEPSCsの記録をFig. 6A, Bに示す。Isoproterenol(100
M)の投与によって,mEPSCsの発生頻度は有意に増加した(P < 0.0001, K-S test;
Fig. 5A-C)。コントロールにおいてmEPSCsの時間間隔(0.2 ± 0.06 s)は,isoproterenol
によって有意に減少した(0.2 ± 0.05 s, n = 10, P < 0.05, paired t-test; Fig. 6E)。一方,
isoproterenolは,mEPSCsの振幅にほとんど影響を与えなかった(K-S test; Fig. 6C)。 Isoproterenol投与時におけるmEPSCsの平均振幅(15.2 ± 1.5 pA, n = 10; Fig. 6D, E) は,コントロール(15.5 ± 1.7 pA)と差がなかった。Isoproterenolもまた,立ち上が り時間(コントロール: 1.0 ± 0.1 ms; isoprpterenol: 1.0 ± 0.1 ms, n = 10),1/2持続時間
(コントロール: 5.9 ± 0.7 ms; isoproterenol: 6.0 ± 0.7 ms, n = 10),減衰時間(コント ロール: 4.1 ± 0.4 ms; isoproterenol: 4.4 ± 0.5 ms, n = 10; Fig. 6E)に影響を与えなか った。
Phenylephrineはglutamate,AMPAおよびNMDA誘発電流を抑制する
AMPA受容体を介したeEPSCsとmEPSCsの解析結果は,phenylephrineがシナプス 後膜のAMPA受容体に作用することにより,EPSCsの振幅を抑制することを示唆す る。そこで,phenylephrineのシナプス後膜に存在するグルタミン酸受容体への効果 を確認するために,tetrodotoxin(1 M)とbicuculline methiodide(20 M)の灌流 投与下で, glutamate(200 M)を投与して誘発される電流を記録した。Glutamate の投与は,パフ投与により行った。保持電位は-70 mVとし,glutamateによる電流 を内向き電流として観察した。Glutamateによって発生した電流の振幅は,219.2 ± 40.7 pAであった(n = 31)。Glutamateを介した内向き電流は,DNQXだけでなく
D-APVにも感受性があった(Fig. 7A)ことから,この電流はnon-NMDA受容体と
NMDA受容体の両方を介して構成されていると考えられた。Glutamateによって発 生した電流の振幅は,phenylephrine(100 M)の灌流投与により85.7 ± 2.6 %(n = 16, P < 0.001, paired t-test; Fig. 8A, C)に減少した。
次に,phenylephrineがAMPA型とNMDA型のどちらの受容体に効果を及ぼしてい
るのかを調べるため,tetrodotoxin(1 M)とbicuculline methiodide(20 M)の灌 流投与下で,錐体ニューロンにAMPA(200 M)またはNMDA(200 M)のパフ 投与を行った。AMPAの投与による反応曲線は,直線状のⅠ-Ⅴ 関係を示した(Fig.
7Da, Dc)。保持電位が-70 mVの時,AMPAによって発生した電流の振幅は,192.9 ± 21.0 pA(n = 36)であり,この電流はDNQX(30 M)によって完全に消失した(Fig.
7B)。Phenylephrineは,AMPAによって誘発された電流を9.7 ± 1.3%抑制した(n = 20,
P < 0.001, paired t-test; Fig. 8A, C)。一方,NMDAによって誘発された電流は,Obokata ら(1997)の研究と同様に,J字型のⅠ-Ⅴ 関係を示した(Fig. 7Db, Dc)。NMDA によって発生した電流(保持電位 = -30 mV)の振幅は,132.3 ± 16.6 pAであった
(n = 36)。NMDAが誘発した電流は,D-APV(50 M)によってほぼ完全に消失し
た(Fig. 7C)。Fig. 8Aに示すように,phenylephrineは,NMDAが誘発した電流を13.9
± 2.7%抑制した(n = 22, P < 0.05, paired t-test)。
Isoproterenolはglutamate,AMPAおよびNMDA誘発電流に影響しない
Isoproterenolは,mEPSCsの振幅を変化させなかったことから,βアドレナリン受 容体の活性化は,シナプス後膜に存在するグルタミン酸受容体にはほとんど影響 を及ぼさないと予想される。そこで,tetrodotoxin(1 M)とbicuculline methiodide
(20 M)の灌流投与下で,glutamate,AMPAまたはNMDA(200 M)のパフ投与 に対するisoproterenolの効果を調べた。Isoproterenol(100 M)の灌流投与では
glutamate,AMPAおよびNMDAによって誘発される電流の振幅は変化しなかった
(glutamate: 95.4 ± 2.1%, n = 15; AMPA: 97.3 ± 2.0%, n = 16; NMDA: 96.4 ± 3.3 %, n
= 14; Fig. 8B, C)。
考察
本研究では,電気刺激によって惹き起こされた eEPSPs に対する α1アドレナリ ン受容体アゴニストによる抑制効果と,βアドレナリン受容体アゴニストによる増 強効果について調べた。その結果,α1 アドレナリン受容体アゴニストである
phenylephrine の効果は,Ⅱ/Ⅲ層およびⅤ層の錐体ニューロンいずれに対しても
eEPSPs の振幅を抑制した。一方,isoproterenol は,Ⅴ層の錐体ニューロンにおい
て eEPSPs の振幅を増大するのに対して,Ⅱ/Ⅲ層の錐体ニューロンに対しては増
強効果が小さいことが明らかとなった。そこで,Ⅴ層の錐体ニューロンからホー ルセル・パッチクランプ記録を行って,glutamate 放出による興奮性シナプス伝達 に対する phenylephrine と isoproterenol の作用機序を調べた。Phenylephrine は,
mEPSCs の発生頻度を変化させずに eEPSCs と mEPSCs の振幅を減少させ,
glutamate,AMPAおよびNMDAのパフ投与によって誘発される内向き電流の振幅 を減少させた。一方,isoproterenolは,eEPSCsの振幅を増大させると同時に,CV とPPRを減少させた。またisoproterenolは,mEPSCsの振幅の変化を伴わずに発生 頻度を増加させる一方,glutamate,AMPAおよびNMDAのパフ投与によって誘発 される電流にはほとんど影響を与えなかった。以上の所見から,phenylephrine に
よる EPSCs の抑制は,AMPA と NMDA 受容体を介した興奮性シナプス後電流を
減少させるのに対して,isoproterenolによるEPSCsの振幅の増大は,シナプス前終
末からのglutamateの放出を促進していることが示唆される。
シナプス前膜における放出確率を推定しうる指標
ある薬物がEPSCs の振幅を変化させる場合,その作用機序については大きく二 つに分けて考えられる。一つは,シナプス前膜に作用して神経伝達物質を含有し たシナプス小胞の放出確率を変化させるものである。もう一つは,シナプス後膜 に作用して受容体の感受性を変化させたり,細胞内セカンドメッセンジャー系を 介してイオンチャネルの開口確率や開口時間などを変化させるものである。この ように,薬物のシナプスレベルにおける作用機序を明らかにすることは,薬物の 神経伝達に対する効果を明らかにする上で,最も基本的な情報を提供するもので あり,極めて重要なテーマである。しかし,直接的に神経伝達物質の放出確率を 定量化する手法はないことから,多くの場合,いくつかの傍証を積み重ねること によって推定することが多い。本研究では,シナプス前膜からの放出確率を予想 するために,三つの手法を用いた。第一は,eEPSCs の CV である。CV は,シナ プス前膜における放出確率を反映することが知られており,放出確率の上昇にと もなってCVは減少すると考えられている(Bekkers and Stevens, 1990)。第二は,
PPRである。PPRは,CV同様,シナプス前膜の放出確率の指標として知られてお り,放出確率が上昇すればPPRは減少するとする報告が多い(Zucker et al., 1989;
Manabe et al., 1993)。第三は,mEPSCsの頻度であり,放出確率の高いシナプスで
は,偶然伝達物質が量子的に放出される頻度は高いと考えられている。
本研究では,細胞外 Ca2+濃度を変化させることによって,CV と PPR について 上述した仮説が当てはまるか否かを検証した。伝達物質の放出は,細胞外カルシ
ウムの細胞内への流入量に依存することが知られている。したがって,細胞外Ca2+
濃度が高い場合は,流入量が多く放出確率が上昇し,細胞外 Ca2+濃度が低い場合 は,放出確率が低いと考えられる。本研究で用いた大脳皮質視覚野の脳スライス 標本でも,上述した仮説と同様の結果が得られたことから,放出確率が高いとき にはCVおよびPPRは減少し,放出確率が低いときは逆にCVとPPRは増加する ことが明らかとなり,放出確率の推定において,PPR と CV は信頼できる指標で あると考えられた。
Phenylephrineはシナプス後膜に作用し,グルタミン酸受容体を抑制する
α1 アドレナリン受容体の局在性は,特異性の高い抗体がなかったため,不明な 点が多かった(Papay et al., 2004)。過去の研究では,シナプス前終末に存在するα アドレナリン受容体はすべて α2タイプで,α1アドレナリン受容体はシナプス後膜 に存在することが示唆されてきた(Starke et al., 1989; Kamisaki et al., 1991)。しか し最近,電子顕微鏡による免疫組織化学的研究により,α1アドレナリン受容体は,
シナプス後膜に豊富に存在するのみならず,グルタミン酸作動性ニューロンのシ ナプス終末と考えられている GrayⅠ型シナプスの前膜にも存在することが確認さ れた(Nakadate et al., 2006)。本研究では,phenylephrineによってeEPSCsの振幅は 減少するものの,そのCVは有意に変化はしないことを明らかにした。また,PPR は,phenylephrine によってはほとんど変化をしないことを観察した。これらの所 見は,phenylephrineがシナプス前膜でglutamate放出の確率を変化させる可能性が 少ないことを示唆している。mEPSCsの記録では,phenylephrineがmEPSCsの発生 頻度に影響を及ぼさないことを明らかとし,CV と PPR の結果から推定される結 論を支持する結果が得られた。MarekとAghajanian(1999)は,α1アドレナリン受 容体の活性化が,ラット前頭皮質Ⅴ層に存在する錐体ニューロンの末端からの神 経伝達物質の放出を増強すると報告した。本研究成果は,この報告に矛盾する。
その理由としては,視覚野と前頭皮質の違い,動物の年齢の違いなどが考えられ るが,最も重要であるのは,記録条件の違いである。彼らの研究では,mEPSCs ではなく,自発性EPSCs(sEPSCs)の発生頻度を記録している。sEPSCsは,偶然 小胞がシナプス前膜に融合して生じる微小なEPSCs のみならず,シナプス前ニュ
ーロンの活動電位によって放出される神経伝達物質によっても惹き起こされる。
したがって,もし phenylephrine がシナプス前ニューロンの活動電位の発生頻度を 上昇させる効果があったと仮定すると,sEPSCsに対する効果は,前膜からの放出 確率の上昇ではなく,単にシナプス前ニューロンの発火頻度が上昇したに過ぎな い可能性が残されている。よって,自発的に起こるEPSCs をシナプス前膜の放出 確率の推定に用いるのは,不適当であると考えられる。
本研究では,α1 アドレナリン受容体の活性化によるシナプス後ニューロンにお ける影響を調べるために,phenylephrine の灌流投与下でglutamate,AMPA および NMDAのパフ投与を行った。Lehmenkuhlerら(1991)は,ノルアドレナリンがシ ナプス後膜に存在するα1アドレナリン受容体を介してNMDA電流を抑制し,興奮 性シナプス伝達を調節することを報告した。本研究では,NMDA だけでなく glutamateやAMPAによるパフ投与によって誘発される電流がphenylephrineによっ て抑制されることを明らかにした。また,細胞内記録を用いた過去の研究におい
て,phenylephrine は PKCの活性化を介して eEPSPs を抑制することが示されてい
る(Kobayashi, 2007)。実際,GluR2のサブユニットには,PKCによってリン酸化 される領域がある(McDonald et al., 2001)ことから,α1アドレナリン受容体の活 性化は,PKCによるGluR2サブユニットのリン酸化を誘導し,その結果,eEPSCs を抑制する可能性が考えられる。
以上のことをまとめると,興奮性シナプス伝達におけるphenylephrineの標的は,
シナプス前膜ではなくシナプス後膜の α1アドレナリン受容体である。ある研究で は,グルタミン酸作動性シナプスのシナプス前領域での PKA と PKC 間の相互作 用が報告されていて(Evans et al., 2001),α1アドレナリン受容体がシナプス前終末 の情報伝達を修飾する可能性を示唆している。しかし,大脳皮質シナプス前終末 の α1アドレナリン受容体の生理的な機能については,これを除いてほとんど情報 がないのが現状である。
Isoproterenolのシナプス前膜におけるglutamate放出の促進
Nowicky ら(1992)は,ラットの知覚運動野において β アドレナリン受容体に
よるeEPSPsの増大効果はほとんどないことを報告しているが,いくつかの研究で
は,その増強効果を報告している(Law-Tho et al., 1993; Kobayashi, 2007)。電圧固 定法を用いた研究では,ラットの前頭葉Ⅴ層錐体ニューロンにおいて,isoproterenol
が cAMP/PKAのシグナリング・カスケードを介して eEPSCs を増強することを示
している(Huang and Hsu, 2006)。本研究では,isoproterenolが誘発するeEPSCsの 振幅の増加と,CV と PPR の減少が同時に起こり,シナプス前終末における β ア ドレナリン受容体の活性がeEPSCsを増強することを示した。またIsoproterenolは,
mEPSCsの振幅にはほとんど影響を与えずに発生頻度を増加させたことから,シナ
プス前終末のβアドレナリン受容体活性化によりglutamateの放出確率が増加した と考えられる。さらに,isoproterenolがglutamate,AMPAまたはNMDAのパフ投 与によって惹起された電流に対してほとんど影響を及ぼさなかったことは,シナ プス後膜の β アドレナリン受容体は,グルタミン酸作動性シナプス伝達に対して ほとんど関与しないことを示唆するものである。以上の結果から,isoproterenolは,
グルタミン酸作動性シナプス前終末に作用して glutamate 放出を増大させる一方,
シナプス後膜のグルタミン酸受容体にはほとんど作用しないと考えられる。この 結論は,以下に示すいくつかの解剖学的,生化学的所見と合致する。電子顕微鏡 を用いた免疫組織化学的研究では,シナプス後膜だけでなく,シナプス前終末に もβアドレナリン受容体が存在することが報告されている(Aoki et al., 1987)。ま た,大脳皮質から得られたシナプトソーム標本において,βアドレナリン受容体ア ゴ ニ ス ト は ,glutamate 放 出 を 増 強 す る こ と が 示 さ れ て い る (Herrero and Sanchez-Prieto, 1996; Wang et al., 2002)。なお,扁桃体ニューロンや(Ferry et al., 1997),海馬CA1錐体ニューロン(Gereau and Conn, 1994),歯状回顆粒ニューロ ン(Dahl and Sarvey, 1990)においても同様に,βアドレナリン受容体の活性化によ
って,glutamate放出が増加することが示されている。
アドレナリン受容体アゴニストは,シナプス後膜におけるグルタミン酸受容体 ではなく,他の神経伝達物質の放出を調節している可能性も考えられる。Roerig ら(1997)は,自発的に発生するsEPSCsが,大脳皮質視覚野のニコチン性アセチ ルコリン受容体またはセロトニン 5-HT3 受容体により調節されていることを報告 している。しかし本研究では,eEPSCsがDNQXによってほぼ完全に消失すること から,アドレナリン受容体アゴニストがグルタミン酸作動性シナプス以外の伝達
を調節することで作用を発揮している可能性は低いと考えられる。
過去のいくつかの研究から,isoproterenolは,cAMP/PKAカスケードの活性化を
介して,glutamateの放出確率を増加させていると考えられる。まず,(1)PKAに
よる小胞結合タンパクであるシナプシンⅠのリン酸化が,シナプス小胞のシナプ ス前膜への移動を促進することが,ラット海馬(Parfitt et al., 1992; Bonanomi et al., 2005)およびネコ大脳皮質のニューロン(Foehring et al., 1989)で報告されている。
そして,(2)シナプス小胞タンパクであるrabphilin 3Aは,cAMP依存性のプロテ インキナーゼの基質となりうること(Fykse et al., 1995)が報告されている。さら に,(3)小脳の顆粒ニューロン→プルキンエニューロンへのシナプス前終末にお いて,カルシウム流入の下流に cAMP が作用することが示されている(Chen and Regehr, 1997)。そして最近,(4)HuangとHsu(2006)が,PKAの活性化がシナプ ス前終末の eEPSCs を増強させることを示した。本研究で用いた標本においても,
isoproterenolによって,類似したメカニズムにより視覚野の eEPSCsが調節されて
いる可能性が考えられる。
機能的意義
大脳皮質視覚野において,ノルアドレナリンは,視覚刺激により放出され
(Marrocco et al., 1987),ノルアドレナリン作動性ニューロンを変性させると,最 初期遺伝子であるc-fosの発現が抑制される(Yamada et al., 1999)。これらの研究 は,視覚野においてノルアドレナリンがグルタミン酸作動性シナプスにおける伝 達に対して重要な役割を果たすことを示唆している。βアドレナリン受容体の活性 化によって生じるglutamateの放出確率の増加と,α1アドレナリン受容体の活性化 によって起こるEPSCs の抑制は,一見相反する効果であって,グルタミン酸作動 性シナプス伝達を効率的に調節していない可能性がある。すなわち,ノルアドレ ナリンは,視覚野の興奮性シナプス伝達を抑制するため(Kobayashi et al., 2000), 結果としてβアドレナリン受容体の活性化によるglutamate放出の増加は,生体で 効果を発揮しない可能性がある。視覚野の錐体ニューロンは,光刺激によって惹 起される興奮性入力だけでなく,自発性の振幅の小さな EPSPs の入力も受けてい る。この際,光刺激による興奮性入力(信号)が,振幅の小さな自発性の EPSPs
(ノイズ)によって攪乱される可能性がある。本研究結果から,シナプス小胞が シナプス前膜へ偶然結合することで伝達物質が放出される活動電位非依存性の
EPSPsは,βアドレナリン受容体の活性化後に発生頻度が増加し,α1アドレナリン
受容体の活性化により EPSPs の振幅が減少することが予想される。この現象は,
結果的にはノイズの基線を減少させることに繋がり,刺激により惹起されたEPSPs の振幅がたとえ抑制されても,光刺激が十分であれば確実にglutamateの放出が誘 発され,結果として伝達の信頼度が上がると考えられる。つまり,α1および β ア ドレナリン受容体の活性化による相反する効果は,ノルアドレナリンの要となる 機能と考えられている SN 比の向上のメカニズムの一つであるかもしれない
(Waterhouse and Woodward, 1980; Armstrong-James and Fox, 1983; Mantz et al., 1988;
Waterhouse et al., 1988)。
文献
Aoki C, Joh TH, Pickel VM. (1987) Ultrastructural localization of β-adrenergic receptor-like immunoreactivity in the cortex and neostriatum of rat brain. Brain Res, 437, 264-282.
Armstrong-James M, Fox K. (1983) Effects of ionophoresed noradrenaline on the spontaneous activity of neurones in rat primary somatosensory cortex. J Physiol (Lond), 335, 427-447.
Bekkers JM, Stevens CF. (1990) Presynaptic mechanism for long-term potentiation in the hippocampus. Nature, 346, 724-729.
Blue ME, Parnavelas JG. (1982) The effect of neonatal 6-hydroxydopamine treatment on synaptogenesis in the visual cortex of the rat. J Comp Neurol, 205, 199-205.
Bonanomi D, Menegon A, Miccio A, Ferrari G, Corradi A, Kao HT, Benfenati F, Valtorta F. (2005) Phosphorylation of synapsin I by cAMP-dependent protein kinase controls synaptic vesicle dynamics in developing neurons. J Neurosci, 25, 7299-7308.
Chen C, Regehr WG. (1997) The mechanism of cAMP-mediated enhancement at a cerebellar synapse. J Neurosci, 17, 8687-8694.
Dahl D, Sarvey JM. (1990) β-adrenergic agonist-induced long-lasting synaptic modifications in hippocampal dentate gyrus require activation of NMDA receptors, but not electrical activation of afferents. Brain Res, 526, 347-350.
Dodt HU, Pawelzik H, Zieglgansberger W. (1991) Actions of noradrenaline on neocortical neurons in vitro. Brain Res, 545, 307-311.
Evans DI, Jones RS, Woodhall G. (2001) Differential actions of PKA and PKC in the regulation of glutamate release by group III mGluRs in the entorhinal cortex. J Neurophysiol, 85, 571-579.
Ferry B, Magistretti PJ, Pralong E. (1997) Noradrenaline modulates glutamate-mediated neurotransmission in the rat basolateral amygdala in vitro. Eur J Neurosci, 9, 1356-1364.
Foehring RC, Schwindt PC, Crill WE. (1989) Norepinephrine selectively reduces slow Ca2+- and Na+-mediated K+ currents in cat neocortical neurons. J Neurophysiol, 61, 245-256.
Fykse EM, Li C, Sudhof TC. (1995) Phosphorylation of rabphilin-3A by Ca2+/calmodulin- and cAMP-dependent protein kinases in vitro. J Neurosci, 15, 2385-2395.
Gereau RW 4th, Conn PJ. (1994) Presynaptic enhancement of excitatory synaptic transmission by -adrenergic receptor activation. J Neurophysiol, 72, 1438-1442.
Herrero I, Sanchez-Prieto J. (1996) cAMP-dependent facilitation of glutamate release by β-adrenergic receptors in cerebrocortical nerve terminals. J Biol Chem, 271, 30554-30560.
Huang CC, Hsu KS. (2006) Presynaptic mechanism underlying cAMP-induced synaptic potentiation in medial prefrontal cortex pyramidal neurons. Mol Pharmacol, 69, 846-856.
Kamisaki Y, Hamahashi T, Okada CM, Itoh T. (1991) Clonidine inhibition of potassium-evoked release of glutamate and aspartate from rat cortical synaptosomes.
Brain Res, 568, 193-198.
Kasamatsu T, Pettigrew JD. (1976) Depletion of brain catecholamines: failure of ocular dominance shift after monocular occlusion in kittens. Science, 194, 206-209.
Kirkwood A, Rozas C, Kirkwood J, Perez F, Bear MF. (1999) Modulation of long-term synaptic depression in visual cortex by acetylcholine and norepinephrine. J Neurosci, 19, 1599-1609.
Kobayashi M, Imamura K, Sugai T, Onoda N, Yamamoto M, Komai S, Watanabe Y.
(2000) Selective suppression of horizontal propagation in rat visual cortex by norepinephrine. Eur J Neurosci, 12, 264-272.
Kobayashi M. (2007) Differential regulation of synaptic transmission by adrenergic agonists via protein kinase A and protein kinase C in layer V pyramidal neurons of rat cerebral cortex. Neuroscience, 146, 1772-1784.
Kobayashi M, Sasabe T, Shiohama Y, Koshikawa N. (2008) Activation of α1-adrenoceptors increases firing frequency through protein kinase C in pyramidal neurons of rat visual cortex. Neurosci Lett, 430, 175-180.
Law-Tho D, Crepel F, Hirsch JC. (1993) Noradrenaline decreases transmission of NMDA- and non-NMDA-receptor mediated monosynaptic EPSPs in rat prefrontal neurons in vitro. Eur J Neurosci, 5, 1494-1500.
Lehmenkuhler C, Walden J, Speckmann EJ. (1991) Decrease of N-methyl-D-aspartate responses by noradrenaline in the rat motorcortex in vivo. Neurosci Lett, 121, 5-8.
Manabe T, Wyllie DJ, Perkel DJ, Nicoll RA. (1993) Modulation of synaptic transmission and long-term potentiation: effects on paired pulse facilitation and EPSC variance in the CA1 region of the hippocampus. J Neurophysiol, 70, 1451-1459.
Mantz J, Milla C, Glowinski J, Thierry AM. (1988) Differential effects of ascending neurons containing dopamine and noradrenaline in the control of spontaneous activity and of evoked responses in the rat prefrontal cortex. Neuroscience, 27, 517-526.
Marek GJ, Aghajanian GK. (1999) 5-HT2A receptor or α1-adrenoceptor activation induces excitatory postsynaptic currents in layer V pyramidal cells of the medial prefrontal cortex.
Eur J Pharmacol, 367, 197-206.
Marrocco RT, Lane RF, McClurkin JW, Blaha CD, Alkire MF. (1987) Release of cortical catecholamines by visual stimulation requires activity in thalamocortical afferents of monkey and cat. J Neurosci, 7, 2756-2767.
McDonald BJ, Chung HJ, Huganir RL. (2001) Identification of protein kinase C phosphorylation sites within the AMPA receptor GluR2 subunit. Neuropharmacology, 41,
672-679.
McCormick DA, Prince DA. (1988) Noradrenergic modulation of firing pattern in guinea pig and cat thalamic neurons, in vitro. J Neurophysiol, 59, 978-996.
Morrison JH, Grzanna R, Molliver ME, Coyle JT. (1978) The distribution and orientation of noradrenergic fibers in neocortex of the rat: an immunofluorescence study. J Comp Neurol, 181, 17-39.
Nakadate K, Imamura K, Watanabe Y. (2006) Cellular and subcellular localization of α1- adrenoceptors in the rat visual cortex. Neuroscience, 141, 1783-1792.
Nicholas AP, Hokfelt T, Pieribone VA. (1996) The distribution and significance of CNS adrenoceptors examined with in situ hybridization. Trends Pharmacol Sci, 17, 245-255.
Nicoll RA, Malenka RC, Kauer JA. (1990) Functional comparison of neurotransmitter receptor subtypes in mammalian central nervous system. Physiol Rev, 70, 513-565.
Nowicky AV, Christofi G, Bindman LJ. (1992) Investigation of β-adrenergic modulation of synaptic transmission and postsynaptic induction of associative LTP in layer V neurones in slices of rat sensorimotor cortex. Neurosci Lett, 137, 270-273.
Obokata K, Kamiya H, Ozawa S. (1997) Differential effects of phorbol ester on AMPA and NMDA components of excitatory postsynaptic currents in dentate neurons of rat hippocampal slices. Neurosci Res, 29, 171-179.
Papay R, Gaivin R, McCune DF, Rorabaugh BR, Macklin WB, McGrath JC, Perez DM.
(2004) Mouse α1B-adrenergic receptor is expressed in neurons and NG2 oligodendrocytes.
J Comp Neurol, 478, 1-10.
Parfitt KD, Doze VA, Madison DV, Browning MD. (1992) Isoproterenol increases the phosphorylation of the synapsins and increases synaptic transmission in dentate gyrus, but not in area CA1, of the hippocampus. Hippocampus, 2, 59-64.
Roerig B, Nelson DA, Katz LC. (1997) Fast synaptic signaling by nicotinic acetylcholine and serotonin 5-HT3 receptors in developing visual cortex. J Neurosci, 17, 8353-8362.
Starke K, Gothert M, Kilbinger H. (1989) Modulation of neurotransmitter release by presynaptic autoreceptors. Physiol Rev, 69, 864-989.
Ulrich D, Huguenard JR. (1996) GABAB receptor-mediated responses in GABAergic projection neurones of rat nucleus reticularis thalami in vitro. J Physiol, 493, 845-854.
Wang SJ, Coutinho V, Sihra TS. (2002) Presynaptic cross-talk of β-adrenoreceptor and 5-hydroxytryptamine receptor signalling in the modulation of glutamate release from cerebrocortical nerve terminals. Br J Pharmacol, 137, 1371-1379.
Wang Z, McCormick DA. (1993) Control of firing mode of corticotectal and corticopontine layer V burst-generating neurons by norepinephrine, acetylcholine, and 1S,3R-ACPD. J Neurosci, 13, 2199-2216.
Waterhouse BD, Woodward DJ. (1980) Interaction of norepinephrine with cerebrocortical activity evoked by stimulation of somatosensory afferent pathways in the rat. Exp Neurol, 67, 11-34.
Waterhouse BD, Sessler FM, Cheng JT, Woodward DJ, Azizi SA, Moises HC. (1988) New evidence for a gating action of norepinephrine in central neuronal circuits of mammalian brain. Brain Res Bull, 21, 425-432.
Yamada Y, Hada Y, Imamura K, Mataga N, Watanabe Y, Yamamoto M. (1999) Differential expression of immediate-early genes, c-fos and zif268, in the visual cortex of young rats: effects of a noradrenergic neurotoxin on their expression. Neuroscience, 92, 473-484.
Zucker RS. (1989) Short-term synaptic plasticity. Annu Rev Neurosci, 12, 13-31.
謝辞
本研究は,九州大学大学院歯学府口腔保健推進学講座 山下喜久教授ならびに 日本大学歯学部薬理学講座 越川憲明教授のご指導の下に行われたもので,先生 方の御懇篤な御指導と御校閲に心より深く感謝いたします。 本研究を遂行する にあたり,終始御指導,御援助頂きました日本大学歯学部薬理学講座 小林真之 准教授に謹んで御礼申し上げます。本研究に御協力下さいました九州大学大学院 歯学府口腔保健推進学講座ならびに日本大学歯学部薬理学教室の皆様に,心から 感謝いたします。
最後になりましたが,大学院生活に多大なる支援をいただき,終始温かい目で 見守ってくれた家族に心から感謝いたします。
総括
1: ノルアドレナリンは,記録ニューロンがⅡ/ⅢおよびⅤ層のいずれにあっても,
eEPSPs の振幅を減少させた。同様の効果は,α1アドレナリン受容体のアゴニ
ストであるphenylephrineの灌流投与でも観察された。第Ⅱ/Ⅲ層とⅤ層に存在 する錐体ニューロン間で振幅減少の程度に有意な差は認められなかった。
2: βアドレナリン受容体アゴニストであるisoproterenolは,Ⅴ層に存在する錐体 ニューロンで観察される eEPSPs の振幅を有意に増大させたが,Ⅱ/Ⅲ層の錐 体ニューロンから記録されるeEPSPsの振幅には影響を与えなかった。
3: Phenylephrineは,頻回刺激(33Hz,10連矩形波)によって惹起される AMPA
受容体を介したeEPSCsの振幅を抑制した。また,1発目のeEPSCsのCVと1
発目のeEPSCsに対する2発目のPPRに影響を与えなかった。
4: Isoproterenol は,2 発目から 10 発目の eEPSCs と比較して,1 発目の eEPSCs の振幅を顕著に増加させ,結果としてPPRを減少させた。また,Isoproterenol が誘発するeEPSCsの振幅の増加は,CVの減少を伴った。
5: Phenylephrineは,mEPSCsの頻度に影響することなく,振幅を抑制した。
6: Isoproterenol は,mEPSCs の振幅に影響を及ぼすことなく,mEPSCs の頻度を
増加させた。
7: Phenylephrineは,glutamate,AMPAまたはNMDAのパフ投与によって惹起さ れる内向き電流を抑制した。
8: Isoproterenol は,glutamate,AMPA または NMDA のパフ投与によって惹起さ れる内向き電流を変化させなかった。
9: 以上の結果より,phenylephrine は,シナプス後膜上の AMPA 受容体または NMDA受容体を介してeEPSCsを減少させるのに対し,isoproterenolは,シナ プス前終末からのglutamate放出を促進することが示唆された。
図表
Table 1. Electrophysiological properties of pyramidal cells in layers II/III and V
Layers II/III Layer V
Regular spiking cell Regular spiking cells Bursting cell Resting membrane potential (mV) -75.9 ± 0.8 (46) -69.6 ± 1.0 (29) -72.1 ± 1.0 (19) Input resistance (MΩ) 33.6 ± 1.4 (48) 44.0 ± 4.4 (26) 41.9 ± 2.0 (18) Action potential
Amplitude (mV) 105.3 ± 1.1 (42) 93.1 ± 1.7 (22) 96.6 ± 1.6 (11) Half duration (ms) 1.9 ± 0.0 (39) 1.5 ± 0.1 (20) 1.3 ± 0.1 (10) eEPSPs
Amplitude (mV) 9.0 ± 0.5 (45) 4.8 ± 0.3 (33) 5.4 ± 0.4 (23) 10-90% rise time (ms) 7.9 ± 0.7 (40) 6.0 ± 0.9 (23) 5.4 ± 0.5 (22) 90-10% decay time (ms) 146.5 ± 10.0 (41) 72.0 ± 9.9 (30) 55.6 ± 9.3 (22) Half duration (ms) 36.4 ± 2.8 (43) 26.3 ± 3.4 (33) 21.0 ± 2.4 (23)
Layer V Layers II/III
Noradrenaline Phenylephrine Isoproterenol
2 mV 50 ms
Pyramidal cells in layers II/III
2 mV 50 ms
1 mV 50 ms
1 mV 50 ms
1 mV 50 ms
2 mV 50 ms
Pyramidal cells in layer V
B
C
a b c
D
a b c
Ctrl
NA
Phe
Iso
Figure 1. Effects of noradrenaline, phenylephrine and isoproterenol on the amplitude of evoked exitatory postsynaptic potentials (eEPSPs) recorded from layers II/III and V pyramidal cells. A: Regular spiking and bursting cells in layer V.
Resting membrane potential was -75 mV. B: An example of effects of noradrenaline (NA), phenylephrine (Phe) and isoproterenol (Iso) on eEPSP amplitude recorded from pyramidal cells in layers II/III. Noradrenaline and phenylephrine decreased eEPSP amplitude, while isoproterenol had little effects on it. C: Noradrenaline and phenylephrine decreased eEPSP amplitude recorded from pyramidal cells in layers V. In contrast, isoproterenol increased eEPSP amplitude. D:
Summary of the effects of noradrenaline, phenylephrine and isoproterenol on the mean amplitude of eEPSPs recorded from pyramidal cells in layers II/III and in layer V. Noradrenaline and phenylephrine consisitently decreased eEPSP amplitude, while isoproterenol significantly increased eEPSP amplitude recorded from layer V but not layers II/III pyra- midal cells (*P < 0.05, **P < 0.01, ***P < 0.001, paired t-test). Error bars indicate SEMs. The number in the paren- theses above the bars represent the number of neurons in each group.
20 mV 10 ms -75
20 mV 10 ms
0.5 nA 0.5 nA
-75
A
Regular spiking cell in layer V Bursting cell in layer Va b
(13) (20)
(20)
(8)
(12) (16)
NA
Phe
Iso Ctrl
Ctrl
Ctrl
Ctrl
Ctrl
0 20 40 60 80 100 120 140 160 180
Normalized amplitude (%)
** ** *
***
***
0 20 40 60 80 100 120 140
160 1st EPSC amp 2nd EPSC amp
0 10 20 30 40 50 60 70 80
0 1 2 3 4 5
Time (min) 3 mM 2 mM 0.5 mM
2 mM [Ca2+]o
EPSC amplitude (pA)Paired-pulse ratio
20 pA 50 ms
C D
E a
b
a
b
2.0
0.5 3.0
0 0.5 1.0 1.5 2.0 2.5 3.0
Paired-pulse ratio
[Ca2+]o (mM)
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8
Coefficient of variation
[Ca2+]o (mM)
* *** ** ***
2.0
0.5 3.0
F
A1 A2
50 pA 50 ms Bicuculline + D-APV Bicuculline + D-APV + DNQX
A B
Figure 2. Effects of DNQX and [Ca2+]o on evoked EPSCs (eEPSCs) recorded from layer V pyramidal neurons. A.
Bath application of 30 µM DNQX effectively suppressed eEPSCs. B. Measurement of peak amplitude of eEPSCs (A1 and A2). C. Time course of the peak amplitudes of 1st (closed circles) and 2nd eEPSCs (open circles). The paired-pulse ratio (PPR) calculated by dividing the 2nd eEPSC amplitude by the 1st amplitude was shown in the bottom panel. Ten consecutive EPSCs were evoked by repetitive stimulation at 33 Hz under application of 20 mM bicuculline methiodide and 50 mM D-APV. [Ca2+]o was changed from 2.0 to 0.5 or 3.0 mM, as shown on the top of the panel. D. Averaged traces of 10 consecutive eEPSCs recorded during the periods a and b indicated in C. E.
![Figure 2. Effects of DNQX and [Ca 2+ ] o on evoked EPSCs (eEPSCs) recorded from layer V pyramidal neurons](https://thumb-ap.123doks.com/thumbv2/123deta/9879211.1905637/36.892.37.876.54.977/figure-effects-evoked-epscs-eepscs-recorded-pyramidal-neurons.webp)