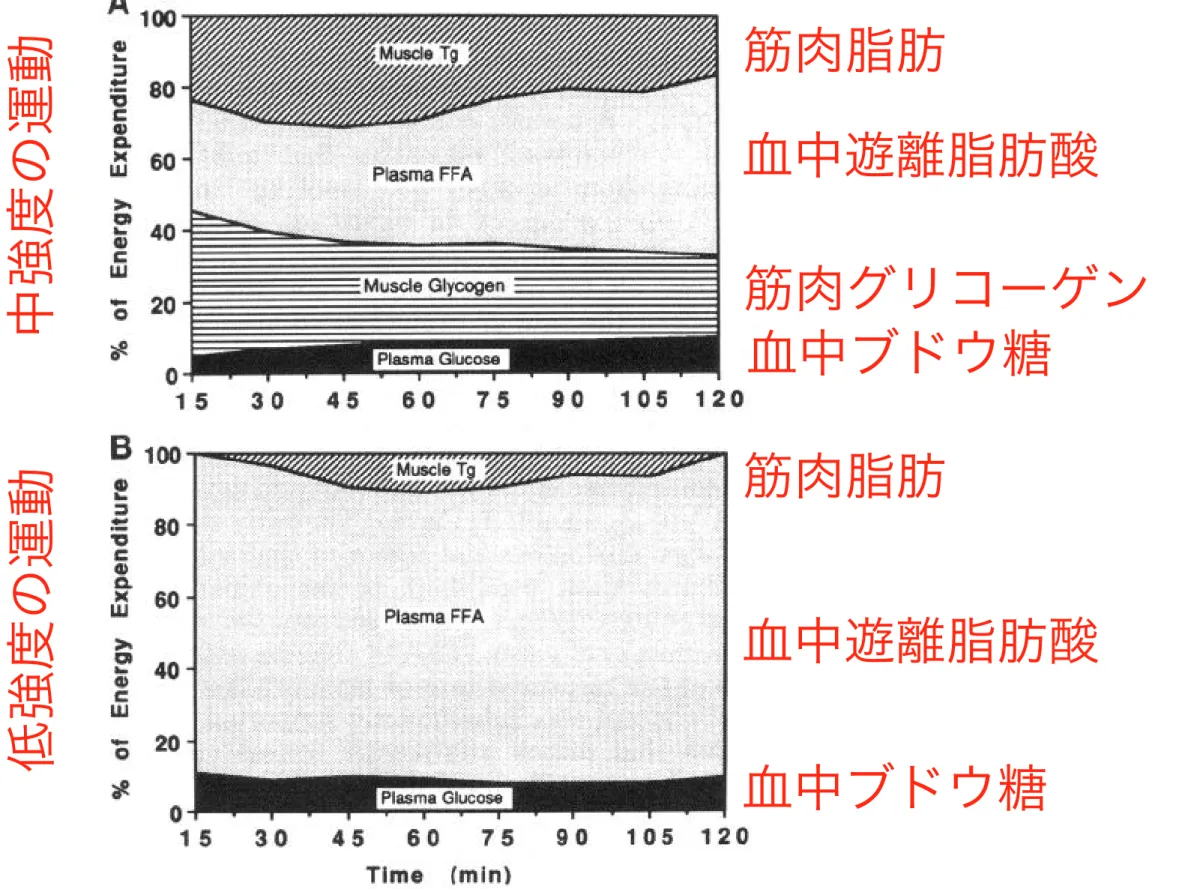
70% VO2maxでの運動時のエネルギー源。運動の後半で筋肉グリコーゲン が減少したときは、摂取した炭水化物が血糖値を維持し、主要な炭水化物 エネルギー源となる。 Coyle 1992 %エネルギー消費量 運動時間(時間) 血糖値 維持される 必要あり 脂肪 血中ブドウ糖 筋肉グリコーゲン
Romijn JA et al (1993)
SUBSTRATE UTILIZATION DURING EXERCISE E387
3oo f/$j Muscle Glycogen • l Muscle Triglycerides q Plasma FFA q Plasma Glucose v- 25 65 % of V02 max 85
Fig. 8. Maximal contribution to energy expenditure derived from glu- cose and FFA taken up from blood and minimal contribution of muscle triglyceride and glycogen stores after 30 min of exercise, expressed as function of exercise intensity. Total amount of calories (Cal) available from plasma does not change in relation to exercise intensity.
:: w 60 Plasma FFA % $ 40 5 ‘ii 20 s 0 -15 30 45 60 75 90 105 120 B loo t 15 30 45 60 75 90 105 120 Time (mln)
Fig. 9. Relative contribution of blood-borne and intramuscular sub- strates to energy production during 120 min of exercise at 25% Voz maX (B) and 65% (A) VO, mBx. Tg, triglycerides.
lipolysis was high during low-intensity exercise and did not increase further with more intense exercise. However, little lipolysis of muscle triglycerides occurred during low- intensity exercise (i.e., 25% Vo, ,,,) compared with ex- ercise at the higher intensities. Despite high lipolytic rates, the R, of plasma FFA, and presumably oxidation, was highest during exercise at 25% Vo, max, and FFA R, declined progressively as the exercise intensity increased to 65 and 85% 00, ,,,Bx. The progressive decline in plasma
Table 2. Plasma concentrations of catecholamines
at rest, during exercise, and after 60 min of recovery
Exercise Intensity,
% iTo max Rest
Exercise, min Recovery
30 120 (60 min) 25 65 85 25 65 Epinephrine 38rt6 59+3* 52+8* 36+4 58217 199+18*t 239+28*t 62ztl7 36+7 625+86*$ 4428 Norepinephrine 231rt43 388+54* 372+41* 223+27 291+31 1,694+237*t 1,851&286*t 424+130 85 266219 5;085+914*$ 293+52
Values are means rt SE in mg/l. * P < 0.05 vs. rest. t P < 0.05 vs. exercise at 25% of ~osmax.
vo2 max.
$ P-C 0.01 vs. exercise at 25 and 65% of
FFA turnover with increasing exercise intensity was off- set by progressive increases in blood glucose turnover. Therefore the contribution of plasma substrates to ca- loric expenditure remained essentially constant over this wide range of exercise intensities. The changes in lipid
metabolism occurred concurrently with changes in car- bohydrate metabolism, consistent with what would have been predicted from previous work, namely that glucose tissue uptake and especially muscle glycogen utilization
increased roughly exponentially in relation to exercise intensity.
Evaluation of Methods
The use of stable isotope tracers for quantification of glucose and palmitate turnover is well established (35). Whole body oxidation rates of fat and carbohydrate were calculated by indirect. calorimetry. This method relies on the assumption that VO, and VCO~ accurately reflect tis- sue 0s consumption and COB production (14). There is little controversy with respect to VO,, of which there are no large stores in the body. However, at exercise intensi- ties that cause hyperventilation, VCO~ may overestimate tissue CO2 production (13). This can potentially result in the overestimation of the rate of carbohydrate oxidation and, concomitantly, an underestimation of fat oxidation
(13). Therefore, before the present study, we conducted a preliminary study that developed a new breath 13C-toJ2C ratio method for the calculation of carbohydrate and fat oxidation during exercise at low to high intensities that is completely independent of the measurement of VCO~ (26). This new method proved that carbohydrate and fat oxidation calculated by indirect calorimetry during exer- cise is valid, including during exercise at 85% VO, max in trained subjects, as performed in the present study.
The rates of fat and carbohydrate oxidation in excess of the measured blood glucose and plasma FFA uptake are assumed to equal the oxidation of intramuscular triglyc- eride and glycogen, respectively. There is no reason to believe that there are major fates other than oxidation for glucose and FFA taken up by the tissues during exercise
(6, 17). The maximal rate of FFA oxidation from periph- eral lipolysis can thus be considered to be equal to plasma FFA uptake. The difference between that rate and total fat oxidation can be attributed largely to intramuscular
筋肉グリコーゲン
筋肉脂肪
血中遊離脂肪酸
血中ブドウ糖
筋肉脂肪
血中遊離脂肪酸
血中ブドウ糖
低強度の運動
中強度の運動
運動強度が高くなると炭水化物エネルギーの消費が増える
Van Loon LJ et al (2001) J Physiol 536:295–304
(Table 1), implying that plasma FFA oxidation has been overestimated in the past, and conversely that TG oxidation has been underestimated. Furthermore, the data show that the contribution of plasma FFA and TG fat sources was about 50 % each at each exercise intensity. We also correctly assumed that total fat oxidation minus plasma FFA oxidation equals the sum of intramuscular plus lipoprotein-derived TG oxidation, while others (e.g.
Romijn et al. 1993, 1995; Sidossis et al. 1997), for reasons
that are not immediately obvious, excluded lipoprotein-derived TG as an important energy source during exercise. Finally, we have recently validated the assumption that the Rd of glucose equals its oxidation rate during
moderate- to high-intensity exercise (Jeukendrup et al.
1999), indicating that our estimates of blood glucose and muscle glycogen oxidation are about equal to those
published previously (Romijn et al. 1993).
In order to eliminate day-to-day variations, we quantified substrate utilisation at different exercise intensities within a single trial. Furthermore, we investigated possible accumulative effects of the incremental protocol on substrate utilisation by asking four of the subjects to perform three additional single trials at 40, 55 and 75 %
Wmax. There were no differences in total fat and
carbohydrate oxidation rates when comparing these single trials with the incremental protocol. The relative
decrease in fat oxidation at 75 % Wmax tended to be even
greater when the subjects started exercise at the high-intensity workload immediately after warming up. This indicates that emptying of the intramuscular TG stores did not seem to play a role in the reduction of fat
oxidation observed at 75 % Wmax.
In the present study, total fat and carbohydrate oxidation rates increased from rest to exercise (Table 2).
However, the relative contribution of fat oxidation to total energy expenditure did not change from resting conditions to exercise, and remained unchanged when
exercise was increased up to 55 % Wmax (Fig. 4). No
significant changes were found in the relative contribution of the different substrate sources to total energy expenditure. However, as exercise intensity was
further increased up to 75 % Wmax, substrate utilisation
changed markedly. Total body fat oxidation rate
decreased by 34 ± 7 % compared to that at the 55 % Wmax
workload (P < 0.05), and this was accounted for by a
reduction in the oxidation rate of plasma FFAs and of other fat sources (i.e. the sum of intramuscular and lipoprotein-derived TG; Table 2 and Fig. 4). A reduction in plasma FFA availability did not seem to play a role in reducing fat oxidation rates, as Ra palmitate (Table 1) and the plasma concentrations of palmitate (Fig. 1) and total FFAs (Fig. 3) did not decrease significantly at the highest workload. As blood flow will have been greatest at the highest exercise intensity, this implies that the arterial FFA supply will also have been greatest at this
time. In contrast to our data, Romijn et al. (1995)
observed a significant reduction in Ra palmitate during high-intensity exercise, which may be explained by the fact that they applied a slightly higher (relative) workload. However, elevation of plasma FFA availability by lipid and heparin infusion only partially restored fat
oxidation at the highest exercise intensity (Romijn et al.
1995). Therefore, both studies indicate clearly that the observed decrease in total fat oxidation cannot be accounted for by a decreased plasma FFA availability
and that the ‘glucose–FFA cycle’ (Randle et al. 1963) does
not operate during high-intensity exercise.
Suggestions have also been made that the transport of FFAs across the plasma membrane limits FFA uptake
Exercise intensity and fuel source utilisation
J. Physiol. 536.1 301
Figure 4. Energy expenditure and fuel selection
Values are means. FFA, free fatty acid.
) at Osaka Taiiku University on July 6, 2011
jp.physoc.org
Downloaded from J Physiol (
血中ブドウ糖 血中遊離脂肪酸 筋肉グリコーゲン その他の脂肪 運動強度 エネルギー消費量
運動時のエネルギー基質(白人)
Janyacharoen T et al. Eur J Appl Physiol (2009) 107:645-65
エネルギー源の酸化速度
運動強度 (% VO2max)
タイ人のエネルギー代謝
炭水化物の利用が多い
女性は男性よりも脂肪の利用が多い(白人)
(最大酸素摂取量の60%での90分間の運動中に利用されたエネルギー源)
Carter SL et al. (2001) Am J Physiol Endocrinol Metab 280:E898–E907
Exercise trial (90-min). By design, the percentage of
V˙
O2 peakelicited in the REL exercise trial was not
different from that elicited in the PRE-training
exer-cise trial, yet the absolute power output (in W) was
higher. Oxygen consumption (V˙
O2) was greater in the
REL exercise trial compared with both the PRE and
ABS trials (Fig. 1A). At 75 and 90 min, V˙
O2was lower
for the ABS compared with PRE trial (P
0.001)
despite the workloads being the same (Fig. 1A).
Fe-males had a lower absolute V˙
O2during the exercise
trial compared with males (Fig. 1B). Endurance
train-ing resulted in a lower RER in the ABS trial, but not in
the REL trial, compared with PRE (P
0.001). RER
increased from rest at the onset of exercise in all
exercise trials (Fig. 1C), and females had a lower
exer-cise RER compared with males (P
0.01; Fig. 1D).
Heart rate was lower during exercise in the ABS trial
compared with both the PRE and REL trials (P
0.001; data not shown). Resting heart rate was
signif-icantly lower after endurance training in both the ABS
and REL compared with the PRE trial (P
0.001; data
not shown). Males had a lower resting heart rate
com-pared with females (P
0.05; data not shown).
Endurance training resulted in an increase in the
proportion of fat oxidized during exercise at the same
ABS workload (P
0.01); however, the proportion of
fat oxidized at the same REL intensity was not affected
by training. Consequently, training resulted in a
de-crease in the proportion of CHO oxidized during
exer-cise at the same ABS (P
0.01), but not REL, intensity
(Fig. 1E). Females oxidized a greater percentage of
energy from fat during exercise compared with males
(P
0.001; Fig. 1F); conversely, males oxidized a
higher percentage of CHO compared with females
dur-ing exercise in all trials (P
0.01; Fig. 1F).
Plasma lactate concentration was lower during
exer-cise (P
0.001) in the ABS trial compared with both
the PRE and REL trials (Fig. 2F). Plasma lactate
Fig. 1. Oxygen consumption (V˙
O2),
respira-tory exchange ratio (RER), and substrate
ox-idation. A: V˙
O2during 90 min of exercise
before (PRE) and after [absolute (ABS) and
relative (REL)] training.
aSignificantly
differ-ent from PRE and ABS (P
0.001);
bsignifi-cantly different from PRE (P
0.001). B:
gender difference in V˙
O2during 90 min of
exercise.
csignificantly different from males
(P
0.001). C: RER during 90 min of exercise
before (ABS) and after (ABS and REL)
train-ing. Main effect for condition:
dP
0.01
(ABS
PRE and REL). D: gender difference
in RER during 90 min of exercise. Main effect
for gender:
eP
0.01 (females
males). E:
effect of training on the proportion of
sub-strate utilized during 90 min of exercise.
Main effect of condition:
dP
0.01 (ABS
PRE and REL);
fP
0.001 (ABS ⇥ PRE and
REL). F: gender differences in the proportion
of substrate utilized during 90 min of
exer-cise. Main effect for gender:
eP
0.01
(fe-males
males);
gP
0.001 (males
fe-males).
E901
GENDER AND METABOLISM DURING EXERCISE
on July 6, 2011
ajpendo.physiology.org
Downloaded from
男性
女性
たんぱく質
脂質
炭水化物
%エネルギー消費量
運動強度 (% VO2max) 脂質 炭水化物 男性 女性 %エネルギー 消費量 %エネルギー 消費量
タイ人のエネルギー代謝
炭水化物の利用が多く性差はない
Janyacharoen T et al. (2009)366 運動生理学的研究I
68.糖 質食 あ るいは脂肪 食 を長期間摂 取 したラッ トにお ける持久運 動 中の エネルギ ー代謝 とグ リコーゲン ・ロ ーデ ィングの効 果
Effects of long-term feeding of carbohydrate diet or fat diet on the glycogen loading and energy metabolism during execise in rats
glycogen loading dietary composition
rat
○岡村浩嗣、菊地範昭、田口優子(大 塚製薬(株)佐賀研究所)
•› KOJI OKAMURA, NORIAKI KIKUCHI, YUKO TAGUCHI (Saga Reserch Institute, Otsuka Pharmaceutical Co.Ltd.)
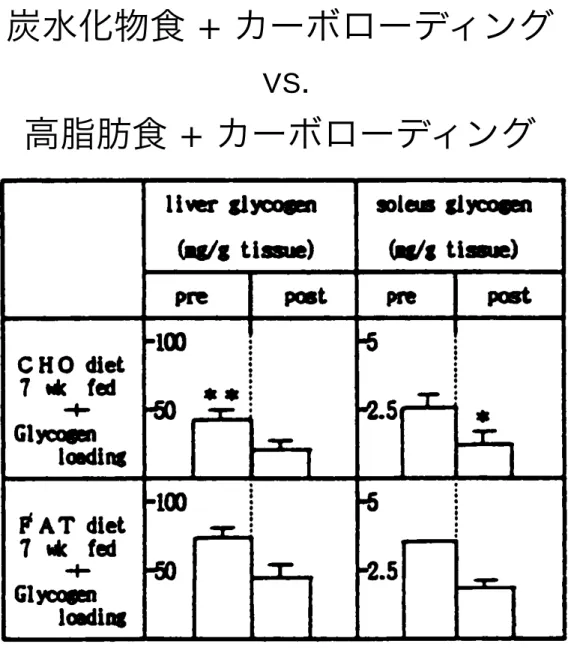
持 久 的 運 動 能 力 の 向 上 に は 、 運 動 開 始 時 に組 織 グ リ コ ー ゲ ン貯 蔵 量 が 多 い こ と と、運 動 中 に 脂 肪 を利 用 し て グ リコ ー ゲ ンの 枯 渇 を防 止 す る こ と が重 要 と 考 え ら れ て い る。 と こ ろ で 、運 動 中 に利 用 され る糖 と脂 肪 の 割 合 は 種 々 の 要 因 に よ っ て 影 響 さ れ る こ と が知 ら れ て い る が 、 日常 摂 取 す る 食 事 の 組 成 も その ひ とつ と 考 え ら れ る 。 ま た 、持 久 力 向 上 の た め の 栄 養 処 方 で あ る グ リ コ ー ゲ ン ・ロ ー デ ィ ン グ の 効 果 に対 し て も 、 日常 の 食 事 組 成 が影 響 す る可 能 性 が あ る。 そ こ で本 研 究 で は 、 脂 肪 食 あ るい は 糖 質 食 を長期 的 に 摂 取 した ラ ッ トを 用 い て 、 持 久 的 運 動 中 の エ ネ ル ギ ー 代 謝 と グ リ コー ゲ ン ・ロー デ ィ ン グ の 効 果 に つ い て 検 討 した。 <方 法> Sprague・Dawley系〓 ラ ッ トを離 乳 直 後 か ら 糖 質 食(タ ンパ ク 質20%、 糖 質69%、 脂 肪11%、 カロ リ ー%)あ る い は脂 肪 食(タ ンパ ク質20%、 糖 質1 %、 脂 肪79%)で7週 間 飼 育 した 。7週 間 飼 育 後 、
FIG. 1 TIE EFFECT OF F7mtCISE ON TIE TISSIE GLYQOGEN DONTFRTS OF RATS FED 010 OR FAT DIET FOR 71915.
Mean±SE
*, ** ; sigai f icxntly different between two groups by t-test (*;p<0.05 **<O.01) 各 群 の 約 半 数 の ラ ッ ト を1時 間 の遊 泳 運 動 の 前 後 で 屠 殺 し た 。 残 りの ラ ッ トに は2時 間 の遊 泳 運 動 を負 荷 し て グ リ コー ゲ ン を 枯 渇 させ た後 高 糖 質 食 を給 与 し、 翌 日1時 間 の遊 泳 運 動 の 前後 で屠 殺 した 。 <結 果> 脂 肪 食 を7週 間摂 取 す る こ とに よ っ て 、 運 動 開 始 時 の 肝 及 び ヒ ラメ 筋 の グ リ コ ー ゲ ンは減 少 した 。 し か し 、1時 間 の遊 泳 に よ る グ リコ ー ゲ ンの 減 少 は 糖 質 食 群 で 多 か った(図1)。 ま た 、 一 度枯 渇 した グ リ コ ー ゲ ンの 肝 及 び ヒ ラ メ 筋 へ の 再 補 充 は 、脂 肪 食 群 に 高糖 質 食 を与 え た と き の 方 が 速 や か だ っ た 。 さ ら に 、 この よ う に し て 再 補 充 さ れ た グ リコ ー ゲ ンの 運 動 中 の利 用 は 、 脂 肪 食 群 で 節 約 さ れ る傾 向 に あ っ た 。(図2)。 以 上 の こ とは 、 日常 摂 取 す る食 事 の 組 成 が 持 久運 動 中 の エ ネ ル ギ ー代 謝 や グ リコ ーゲ ン ・ロ ー デ ィ ン グ の 効 果 に 対 して影 響 す る こ とを示 唆 して い る と思 わ れ る 。
FIG. z TIE FExr OF (LYOGEN IAIDIIG ON GLYQ)G 4 UTILIZATION OF RATS FED QI)OR FAT DIET FOR 7 *E13 INKING E RCISE.
Metn•}SE
*, ** ; sigiificantiy different bet*een two sroi s by t-test (* ; p <O.( , ** <O.O1)
岡村ら
366 運動生理学的研究I
68.糖 質食 あ るいは脂肪 食 を長期間摂 取 したラッ トにお ける持久運 動 中の エネルギ ー代謝 とグ リコーゲン ・ロ ーデ ィングの効 果
Effects of long-term feeding of carbohydrate diet or fat diet on the glycogen loading and energy metabolism during execise in rats
glycogen loading dietary composition
rat
○岡村浩嗣、菊地範昭、田口優子(大 塚製薬(株)佐賀研究所)
•› KOJI OKAMURA, NORIAKI KIKUCHI, YUKO TAGUCHI (Saga Reserch Institute, Otsuka Pharmaceutical Co.Ltd.)
持 久 的 運 動 能 力 の 向 上 に は 、 運 動 開 始 時 に組 織 グ リ コ ー ゲ ン貯 蔵 量 が 多 い こ と と、運 動 中 に 脂 肪 を利 用 し て グ リコ ー ゲ ンの 枯 渇 を防 止 す る こ と が重 要 と 考 え ら れ て い る。 と こ ろ で 、運 動 中 に利 用 され る糖 と脂 肪 の 割 合 は 種 々 の 要 因 に よ っ て 影 響 さ れ る こ と が知 ら れ て い る が 、 日常 摂 取 す る 食 事 の 組 成 も その ひ とつ と 考 え ら れ る 。 ま た 、持 久 力 向 上 の た め の 栄 養 処 方 で あ る グ リ コ ー ゲ ン ・ロ ー デ ィ ン グ の 効 果 に対 し て も 、 日常 の 食 事 組 成 が影 響 す る可 能 性 が あ る。 そ こ で本 研 究 で は 、 脂 肪 食 あ るい は 糖 質 食 を長期 的 に 摂 取 した ラ ッ トを 用 い て 、 持 久 的 運 動 中 の エ ネ ル ギ ー 代 謝 と グ リ コー ゲ ン ・ロー デ ィ ン グ の 効 果 に つ い て 検 討 した。 <方 法> Sprague・Dawley系〓 ラ ッ トを離 乳 直 後 か ら 糖 質 食(タ ンパ ク 質20%、 糖 質69%、 脂 肪11%、 カロ リ ー%)あ る い は脂 肪 食(タ ンパ ク質20%、 糖 質1 %、 脂 肪79%)で7週 間 飼 育 した 。7週 間 飼 育 後 、
FIG. 1 TIE EFFECT OF F7mtCISE ON TIE TISSIE GLYQOGEN DONTFRTS OF RATS FED 010 OR FAT DIET FOR 71915.
Mean±SE
*, ** ; sigai f icxntly different between two groups by t-test (*;p<0.05 **<O.01) 各 群 の 約 半 数 の ラ ッ ト を1時 間 の遊 泳 運 動 の 前 後 で 屠 殺 し た 。 残 りの ラ ッ トに は2時 間 の遊 泳 運 動 を負 荷 し て グ リ コー ゲ ン を 枯 渇 させ た後 高 糖 質 食 を給 与 し、 翌 日1時 間 の遊 泳 運 動 の 前後 で屠 殺 した 。 <結 果> 脂 肪 食 を7週 間摂 取 す る こ とに よ っ て 、 運 動 開 始 時 の 肝 及 び ヒ ラメ 筋 の グ リ コ ー ゲ ンは減 少 した 。 し か し 、1時 間 の遊 泳 に よ る グ リコ ー ゲ ンの 減 少 は 糖 質 食 群 で 多 か った(図1)。 ま た 、 一 度枯 渇 した グ リ コ ー ゲ ンの 肝 及 び ヒ ラ メ 筋 へ の 再 補 充 は 、脂 肪 食 群 に 高糖 質 食 を与 え た と き の 方 が 速 や か だ っ た 。 さ ら に 、 この よ う に し て 再 補 充 さ れ た グ リコ ー ゲ ンの 運 動 中 の利 用 は 、 脂 肪 食 群 で 節 約 さ れ る傾 向 に あ っ た 。(図2)。 以 上 の こ とは 、 日常 摂 取 す る食 事 の 組 成 が 持 久運 動 中 の エ ネ ル ギ ー代 謝 や グ リコ ーゲ ン ・ロ ー デ ィ ン グ の 効 果 に 対 して影 響 す る こ とを示 唆 して い る と思 わ れ る 。
FIG. z TIE FExr OF (LYOGEN IAIDIIG ON GLYQ)G 4 UTILIZATION OF RATS FED QI)OR FAT DIET FOR 7 *E13 INKING E RCISE.
Metn•}SE
*, ** ; sigiificantiy different bet*een two sroi s
by t-test (* ; p <O.( , ** <O.O1)
炭水化物食 vs. 高脂肪食 炭水化物食 + カーボローディング vs. 高脂肪食 + カーボローディング 岡村ら 体力医学会1986