© 2020 Japanese Proteomics Society
総合論文
感染性 HIV-1 粒子のプロテオーム解析から解き明かす
宿主性タンパク質の多彩な機能
岸本直樹 *,三隅将吾
*E-mail: [email protected] 熊本大学大学院生命科学研究部:862-0973 熊本県熊本市中央区大江本町 5-1 (受付 2020 年 9 月 12 日,改訂 2020 年 9 月 25 日,受理 2020 年 9 月 28 日)Human immunodeficiency virus type 1(HIV-1)は CD4 陽性 T 細胞を始めとする免疫細胞に感染する.これまでに,免 疫細胞内の宿主性タンパク質による HIV-1 複製制御機構は数多く報告されている.しかしながら,HIV-1 粒子内に取込ま れる宿主性タンパク質については十分に検討されていない.したがって,ウイルス粒子内宿主性タンパク質の同定および HIV-複製における機能解析は,HIV-1 複製機構の解明に向けた一つの手段となる.また,ウイルス性タンパク質は細胞内 の翻訳後修飾酵素を利用し,その機能の多様性を獲得している.したがって,ウイルス性タンパク質の受ける翻訳後修飾 の同定および機能解析も,HIV-1 複製機構の解明において重要である.本総合論文では,感染性 HIV-1 粒子そのものを試 料としたプロテオーム解析を通して明らかにした,Glyceraldehyde-3-phosphate dehydrogenase による HIV-1 複製阻害機構 とウイルス性タンパク質であるカプシドが受ける翻訳後修飾の重要性について解説する.
1 序 論
Human immunodeficiency virus type 1(HIV-1)感染症は, 世界三大感染症のひとつである.かつては「死の病気」と 恐れられていた HIV-1 感染症は,近年,治療における薬 剤貢献度が向上し,今や「コントロール可能な慢性疾患」 と再定義されつつある.しかし,根治療法が未だ確立して いないために HIV-1 感染者は一生涯服薬を続けねばなら ず,アドヒアランスの維持が難しいこと,HIV-1 の有する 最大の感染特性である易変異性を克服できていないために 耐性ウイルスが出現した場合,既存薬での治療が困難とな る症例が増加していることが問題となっている.したがっ て,HIV 感染症治療における薬剤貢献度が向上してはいる ものの,未だに HIV-1 感染症はアンメットメディカルニー ズが高い疾患として位置づけられており,既存の抗 HIV-1 薬とは作用点が異なる新規治療標的を見出すことに寄与す る基礎研究および新規知見が重要となっている. HIV-1の複製機構は,ウイルス性タンパク質だけではな く宿主性タンパク質によっても複雑に制御されており,感 染必須因子および感染阻害因子が存在することは良く知ら れている.Fig. 1 に HIV-1 複製機構の概略を示す.約 9.2 kb の HIV-1 のゲノムにはわずか 20 種類たらずのウイルス性 タンパク質がコードされているに過ぎないため,CD4 や ケモカイン受容体である CCR5 および CXCR4 に代表され るような,Fig. 1 において赤字で示した様々な宿主性タン パク質や tRNA が利用される1)∼ 5).一方で,HIV-1 複製 を阻害する宿主性タンパク質も知られており,代表的な 宿主性タンパク質として,Fig. 1 において青字で示したシ チジンデアミナーゼである Apolipoprotein B mRNA-editing enzyme-catalytic polypeptide-like-3G(APOBEC3G) は, G-to-A hypermutationを引起こすことで抗 HIV-1 活性を示 す6),7).これらの知見は,HIV-1 複製に必要とされる感染 必須因子の機能的な阻害,または感染阻害因子の作用の増 強や模倣が HIV-1 複製を破綻させることを期待させ,実際 に CCR5 阻害剤としてマラビロクが認可されている.ウイ ルス性因子を標的とした治療戦略では,ウイルスが複製す る度に薬剤の標的分子に変異が挿入され,タンパク質の構 造が変化する可能性があるが,宿主性因子を標的とした治 療戦略では,ウイルスの複製に伴って薬剤の標的分子に変 異が挿入されることはなく,タンパク質の構造が変化する ことは無いため,宿主性因子を標的とした治療戦略はウイ ルスの易変異性の問題を克服した新たな抗 HIV-1 治療法と して期待できる. 興味深いことに,HIV-1 粒子内には宿主性タンパク質が 取込まれることが知られている8)∼ 10).HIV-1 粒子の直径 はわずか 100 nm 程度であるため,ウイルス粒子内に取込 まれるタンパク質は受動的に取込まれているとは考え難 く,ウイルスが効率的なウイルス複製のために積極的に宿 主性タンパク質を取り込んでいるか,または宿主細胞側 が HIV-1 に対抗するために積極的に宿主性タンパク質を
アイソフォームとして存在し,多様な機能を獲得している. 例えば,HIV-1 外被糖タンパク質である Env はウイルスの 侵入を担うタンパク質であるが,我々の解析により,主に 糖鎖修飾構造の違いにより等電点の異なる数十のアイソ フォームからなり,ウイルス粒子表面を構成していること が明らかになっている11).Env は,このグリカンシール ドと呼ばれる糖鎖修飾を受けることで感染した宿主の液性 免疫からウイルスを逃避させるという機能を獲得する.し たがって,ウイルス性タンパク質の受ける翻訳後修飾の同 定およびその意義を明らかとすることは,新規 HIV-1 複製 制御機構の解明につながる.さらに,その翻訳後修飾機構 を解き明かすことができれば,新たに翻訳後修飾付与酵素 を標的とした抗 HIV-1 治療を構築できる可能性もある. 本論文では,新規 HIV-1 複製制御機構を明らかとするた めに,我々が実施した感染性 HIV-1 粒子そのものを試料と 取込ませているか,ということが示唆される.しかしなが ら,ウイルス感染細胞を対象とした解析よりもウイルスを 対象とした解析は操作が煩雑であるため,ウイルス粒子内 宿主性タンパク質の機能解析はあまり進んでおらず,その ウイルス学的意義は未解明の部分を多く有する.したがっ て,HIV-1 粒子内に取込まれている宿主性タンパク質の同 定およびウイルス学的機能解析を行うことは,新規 HIV-1 複製制御機構の解明につながり,さらに新規 HIV-1 治療戦 略を開発するための有用な知見となると考えられる. 二次元電気泳動と質量分析を組合わせたプロテオーム解 析では,タンパク質の同定とともに,等電点や分子量の違 うタンパク質アソフォームの同定において有用である.前 述の通り HIV-1 ゲノムにコードされているタンパク質は わずか 20 種類程度しかないため,ウイルス性タンパク質 は宿主細胞の翻訳後修飾機構を有効利用することで複数の
Fig. 1 HIV-1 replication and host factors involved
The host factors required for suitable viral replication are shown in red and the restriction factors are shown in blue. HIV-1 mRNA is transcribed from the provirus and then viral proteins are translated. The endosomal sorting complexes required for transport (ESCRT) complex mediates viral assembly. In a mature virus, the incorporated host tRNALys3 is used as a primer
for viral reverse transcription. At the viral entry step, CD4 acts as a receptor and CCR5 and/or CXCR4 acts as a coreceptor. APOBEC3G induces G-to-A mutation during reverse transcription through enzymatic activity to prevent the correct reverse transcription.
2 感染性 HIV-1 粒子のプロテオーム解析 これまでに HIV-1 感染細胞を用いた研究によって,タン パク質レベルでの HIV-1 複製機構の解明は盛んに行われて いるものの,感染性 HIV-1 粒子そのものを対象としたタン パク質レベルでの解析は少ない.そこで我々は,HIV-1 複 製制御機構を詳細に理解する糸口を得るために,精製した 感染性 HIV-1 粒子を試料としたプロテオーム解析を実施 した. したプロテオーム解析を解説する.また,ウイルス粒子内 宿主性タンパク質が有する HIV-1 複製における機能を明ら かにした例として,「解糖系酵素である Glyceraldehyde-3-phosphate dehydrogenase(GAPDH)が HIV-1 複製を阻害 する」という発見について解説する12),13)とともに,「リン
酸化によるカプシドタンパク質(CA)の翻訳後修飾が脱 殻過程におけるウイルス CA コアの安定性の制御に重要で ある」という発見について解説する14)∼ 16).
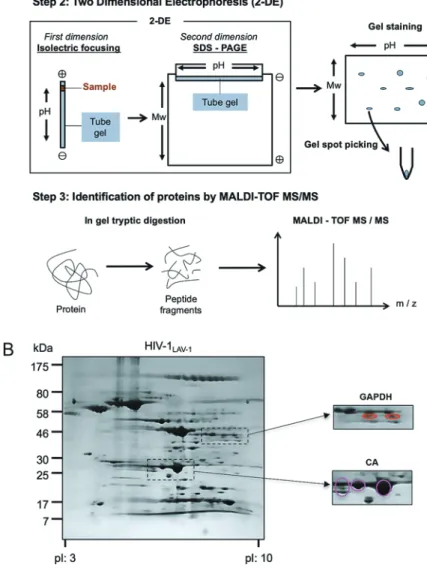
Fig. 2 Proteome analysis of infectious HIV-1 particles
(A) Schematic illustration of proteome analysis conducted in our study. Infectious HIV-1 particles were collected from the culture supernatant of CEM/LAV-1 cells. To purify them, centrifugation methods were used (Step 1). The lysate of purified infectious HIV-1 particles (1 mg protein) was separated by 2-DE. The gel obtained after 2-DE was stained and stained spots were excised to obtain gel pieces (Step 2). The gel pieces were subjected to in-gel tryptic digestion for analysis by MALDI-TOF MS/MS. (B) Representative image of CBB-stained 2DE gel. The gel obtained after 2-DE was stained with CBB G-250. Two expanded images of GAPDH and CA in the gel are shown on the right. Spots derived from GAPDH and CA are emphasized in red and magenta, respectively.
が報告されている18)∼ 21).また,肝炎ウイルスやヒトパラ インフルエンザウイルスの複製において特異的な役割を有 することも報告されている22)∼ 24).しかし HIV-1 複製にお ける GAPDH の役割報告されていなかった.そこで本項で は,GAPDH が HIV-1 粒子内に取込まれる意義について検 討を行った結果12),13)について概説する.
まず,HIV-1 粒子内に取込まれた GAPDH が,HIV-1 の 感染性に影響を与えているのかを検討した.CEM/LAV-1 細胞ヘの GAPDH 特異的 siRNA の処理によって細胞毒性 が無くウイルス産生量等にも影響を与えないレベルでウイ ルス産生細胞内 GAPDH を減少させることで,ウイルス粒 子内に取込まれる GAPDH の量のみを減少させたウイルス を作製した.作製したウイルスを用いて HIV-1 複製の各過 程を評価したところ,ウイルスの産生量やウイルスの侵入 効率に差はないものの,逆転写過程以降の複製過程が野生 型ウイルスよりも亢進しており,GAPDH の取込みが減少 したウイルスは高い感染性を有していた.また各種逆転写 反応産物を検出したところ,GAPDH の取込みが減少した ウイルスでは逆転写初期産物量が増加しており,GAPDH はウイルス粒子内に取込まれることで逆転写反応の開始そ のものを阻害することが示唆された. HIV-1はレトロウイルスであり,その複製においてウイ ルスゲノム RNA からゲノム DNA への変換,つまり逆転 写反応は非常に重要な過程である.HIV-1 の逆転写反応 は,HIV-1 ゲノム RNA をテンプレート,宿主細胞由来の tRNALys3がプライマーとなり,ウイルス性タンパク質であ る逆転写酵素によって進行する.興味深いことに,HIV-1 は出芽の際にウイルス産生細胞内の tRNALys3をウイルス ゲノム RNA および逆転写酵素とともに粒子内に取込ん でおり25)∼ 27),HIV-1 逆転写反応は HIV-1 粒子内で開始さ れ,細胞に感染後に完了する28).我々は,GAPDH は逆転 写酵素活性に直接影響を与えないという結果を得た.そこ で,GAPDH の取込みが減少したウイルス粒子内のウイル スゲノム RNA および tRNALys3を定量したところ,ゲノム
RNAの量はコントロールウイルスと同定度取込まれてい
るが,tRNALys3の取込量が増加していた(Fig. 3A).一方
で GAPDH の取込み量を増加させたウイルスを作製し検討 を行ったところ,ウイルス粒子内 tRNALys3量の減少,逆転
写効率の低下,感染価の低下が確認できた(Fig. 3A).し たがって,GAPDH は感染性 HIV-1 粒子内への tRNALys3の
取込みを阻害することが明らかになった.
次 に, ウ イ ル ス 粒 子 内 へ の GAPDH 取 込 み 機 構 と GAPDHによる tRNALys3の取込み阻害機構について検討
し た.tRNALys3は,Lysyl-tRNA synthetase(LysRS) と 複
合体を形成し,LysRS とウイルス構造タンパク質である Pr55gagとの相互作用を介しウイルス粒子内に取込まれる ことが知られている29)∼ 32).そこで,まず抗 GAPDH 抗体
プ ロ テ オ ー ム 解 析 で は, 感 染 性 ウ イ ル ス 粒 子 と し て HIV-1LAV-1を試料として用い,二次元電気泳動および
MALDI-TOF MS/MSによる解析を行った(Fig. 2A).二次 元電気泳動においてより明瞭なスポットとして,ウイルス 構成タンパク質を分離するために,ウイルスライセート (1 mg)を 1 回の電気泳動に用いた.感染性 HIV-1 粒子は 感染細胞の培養上清から得られるが,1 mg のライセート を得るためには約 1 L 程度の培養上清が必要である.一般 に感染性 HIV-1 はウイルス分子クローンをトランスフェク ションすることで作製されるが,プロテオーム解析用のウ イルスをトランスフェクションにより得ることは非効率的 である.そのため,CD4 陽性 T 細胞株 CEM 細胞に HIV-1LAV-1株を感染させた HIV-1 持続感染細胞株である CEM/
LAV-1細胞の培養上清から HIV-1LAV-1を回収した.この際,
変異の違いによってウイルス性タンパク質が複数のスポッ トとして検出されることを避けるために,CEM/LAV-1 細 胞は一度モノクローナル化したものを利用した.また,デー タベース内に報告されているウイルスタンパク質の一次配 列との相違を予め確認しておくために,CEM 細胞に組込 まれたウイルスゲノムのシークエンス解析を行った後にウ イルスを回収した.ウイルスの精製,濃縮では,0.22 μm のフィルターで培養上清を濾過した後に,100,000 xg で遠 心後,得られたウイルスペレットを PBS で洗浄を繰返し 行い,プロテオーム解析の試料となる感染性 HIV-1 粒子 を得た.O’Farrell の二次元電気泳動17)を応用して,キャ リアアンフォライトを用いた等電点電気泳動および SDS-PAGEを行った.得られたゲルを染色することで分離した 感染性 HIV-1 粒子のプロテオームを検出し,感染性 HIV-1 粒子を構成するプロテオームの二次元マップを作製した. Fig. 2Bに二次元マップの一例を示す.二次元マップ上の 各スポットのタンパク質を帰属する際は,ゲルの切り出 し,トリプシンを用いた in-gel 消化,消化産物の脱塩処理 を行い,MALDI-TOF MS/MS による解析を行った.以下 に,同定できた宿主性タンパク質の中でも解糖系酵素であ る GAPDH に着目し,GAPDH がウイルス粒子内に取込ま れる意義を検討した研究12),13)を紹介する.また,ウイル ス性タンパク質 CA の翻訳後修飾として同定したリン酸化 のウイルス学的重要性について検討した研究14)∼ 16)も紹介 する. 3 HIV-1 複製における宿主性タンパク質 GAPDH のウイ ルス学的重要性 G A P D H は , g l y c e r a l d e h y d e 3 - p h o s p h a t e か ら 1,3-bisphosphoglycerateへの変換を担う解糖系酵素として 良く知られているが,近年「moonlighting タンパク質」と して注目されており,その機能は,DNA 修復,mRNA の 安定性の調節,アポトーシスの惹起など多岐にわたること
安定構造である 4 量体 GAPDH33),34)が Pr55gagの matrix (MA) regionと capsid (CA) region の 2 箇所で相互作用し,HIV-1 粒子内に取込まれることを明らかにした(Fig. 3B).次 に,ウイルス粒子内への LysRS の取込みを検討したとこ を用いた共免疫沈降法を行ったところ,GAPDH と Pr55gag
との相互作用は見られたが,GAPDH と LysRS との相互作 用は見られなかった.また in silico docking simulation およ び変異体を用いた相互作用解析等を実施し,細胞内おいて
Fig. 3 Inhibition mechanism of viral reverse transcription by virion-packaged GAPDH
(A) The packaging level of GAPDH is inversely correlated with that of tRNALys3, which affects the subsequent reverse
transcription reaction. CEM/LAV-1 cells were treated with GAPDH-specific siRNA and a GAPDH expression vector to obtain a low-level-GAPDH-packaging and high-level-GAPDH-packaging virus, respectively. The low-level-GAPDH-packaging virus showed increased intravirion-tRNALys3 levels without altering the packaging levels of viral genomic RNA. In addition, the
low-level-GAPDH-packaging virus showed increased reverse transcription level and infectivity. In contrast, the high-level-GAPDH-packaging virus showed decreased intravirion-tRNALys3 level, reverse transcription level and infectivity. (B) Schematic
illustration of interaction between GAPDH and Pr55gag. Helix 10 of GAPDH composed of residues 256-267 is required for the interaction of GAPDH with the MA and CA regions of Pr55gag. The residues responsible for each interaction are shown. (C) Schematic illustration showing the mechanism by which GAPDH prevents viral reverse transcription. Upper panel: when GAPDH in HIV-infected cells is mainly used for glycolysis, the tRNALys3-LysRS complex is easily incorporated into virus
particles. Lower panel: when GAPDH in the infected cells can function as a moonlight protein, GAPDH prevents the packaging of the tRNALys3-LysRS complex into viral particles.
イマーそのものをウイルス粒子内に取り込ませない役割を 担っていた宿主性タンパク質の機能を模倣して HIV-1 逆転 写反応を阻害する方法は,HIV-1 の易変異性に対抗するた めの基盤構築となりうると考えている. 4 HIV-1 CA が受ける翻訳後修飾と HIV-1 複製 HIV-1 CAは HIV-1 の感染性の維持に非常な重要なウイ ルス性タンパク質であり,6 量体または 5 量体を最小骨格 とする円錐形の CA コアを形成し,ウイルスゲノム RNA を保護する役割を有する39)∼ 41).CA はウイルスゲノム上 の構造遺伝子 gag にコードされており,前駆体タンパク 質である Pr55gagとして発現する42).Pr55gagは HIV-1 の出 芽後に,自身の持つ HIV-1 プロテアーゼによってプロセ シングを受け,CA を含むウイルス性構造タンパク質を 生じる43).生じた CA はウイルスゲノム RNA を内包する 形で CA コアを形成し,ウイルスが標的細胞に侵入した 後に CA コアは崩壊する(この崩壊の過程を脱殻という). 感染性 HIV-1 粒子の二次元マップおよび質量分析の結果 は,CA には等電点の異なるアイソフォームがあることを 示している(Fig. 2B).そこで我々は,CA の等電点の違 いの原因およびその意義について検討を行った.本項では, CAが受ける翻訳後修飾と翻訳後修飾酵素,および CA が 受ける翻訳後修飾の脱殻過程における重要性を明らかにし た我々の結果14)∼ 16)を概説する. CAが コ ー ド さ れ て い る gag 領 域 は ス プ ラ イ シ ン グ を受けないため,等電点の異なるアイソフォームはス プライシングバリアント由来ではなく,翻訳後修飾の 違いに起因すると考えられた.実際に等電点の低いス ポット由来のペプチド消化産の物質量分析の結果より, 1PIVQNIQGQMVHQAISPR18の分子量(理論値:2016.36) からリン酸化を意味する 80 シフトしたピークが検出され た(実測値:2096.55).さらにこのピークはアルカリフォ スファターゼ処理によって消失した.したがって,Ser16 はリン酸化を受けることが明らかとなった.興味深いこと に Ser16はそれに続く Pro17と共に HIV-1 株間で高度に保
存されている.そこで,S16A 変異体ウイルス,S16E 変異 体ウイルス,S16A/P17A 変異体ウイルスの感染価を評価し たところ,S16E 変異体は予想通り感染性の低下を示さな いが,S16A 変異体および S16A/P17A 変異体は感染性の低 下を示し,S16A/P17A 変異体が最も感染性の低下を示した. したがって,Ser16のリン酸化はそれに続く Pro17と共に HIV-1の感染性の維持に重要であることが明らかとなった. Ser16のリン酸化はウイルス粒子内で検出されたもので ある.そこで,リン酸化 Ser16-Pro17がコアの形成に重要で あるのか,それともコアの崩壊に重要であるのかを検討 した.まず,電子顕微鏡を用いてウイルス粒子内 CA コア の観察を行ったところ,S16A/P17A 変異体ウイルスは WT ろ,GAPDH の取込み量が減少すると LysRS の取込み量は 増加し,GAPDH の取込み量が増加すると LysRS の取込み 量は減少していた.一方で,ウイルス産生細胞内 LysRS を過剰発現させLysRSの取込みを増加させたウイルスでは, GAPDHの取込み量が減少していた.また,Pr55gagとの 相互作用に必要な GAPDH の helix10 内のアミノ酸に変異 を導入し Pr55gagと相互作用を欠落させた GAPDH 変異体 (D256R/K260E/K263E/E267R)をウイルス産生細胞に発現 させた結果では,ウイルス粒子内への tRNALys3の取込み 量の低下は見られなかった.これらの結果より,GAPDH は Pr55gagと直接相互作用することでウイルス粒子内に取 込まれること,GAPDH は Pr55gagと tRNALys3-LysRS複合
体の相互作用を阻害することでウイルス粒子内の逆転写反 応のプライマー,tRNALys3量を低下させ HIV-1 逆転写反応
を阻害することが明らかとなった. GAPDHが細胞内に豊富に存在するにも関わらず HIV-1 が複製できる点について現在までに詳細には明らかにで きていない.しかしながら,HIV-1 感染に伴って細胞内代 謝は好気的解糖にシフトすること35),好気的解糖下では GAPDHは moonlight タンパク質としての機能を発揮でき ないことが知られている36).そこで我々は,HIV-1 感染に 伴う好気的解糖へのシフトは GAPDH の抗 HIV-1 活性を阻 害するために必要であり,好気的解糖へのシフトを解除し, GAPDHが moonlight タンパク質として抗 HIV-1 活性を発 揮できれば,効果的にウイルス粒子内への tRNALys3-LysRS
複合体の取込み阻害することによって逆転写反応を阻害で きると考えている(Fig. 3C). 以上は感染性 HIV-1 粒子のプロテオーム解析の情報を基 に明らかにした宿主性タンパク質による HIV-1 複製阻害機 構の一例である.HIV-1 の変異獲得は逆転写反応のエラー に起因する.GAPDH そのものを治療に応用することは難 しいと考えられるが,tRNALys3-LysRS複合体の取込みを阻
害する知見はこれまでに無いため,tRNALys3そのもののウ
イルス粒子内への取込みが阻害されれば,逆転写反応の開 始そのものが阻害されるという我々の発見は,HIV-1 の易 変異性に対抗する手段となりうる.GAPDH が tRNALys3の
取込みを阻害するという知見を得る中で,GAPDH と相互 作用するウイルス前駆体タンパク質の領域(MA 領域内の R58, Q59, Q63,および CA 領域内の R214, E208)(Fig. 3B) に変異を持つウイルスは,複製ができないという知見も 得ている.さらに,著者らは GAPDH 以外の解糖系酵素 の中でも Alpha-enolase や Pyruvate kinase isozymes M2 は HIV-1粒子内に取込まれていること,そしてそれぞれの解 糖系酵素は独立して逆転写過程を阻害するという知見も 得ている37),38).HIV-1 逆転写酵素活性を阻害する既存薬は,
逆転写酵素内にアミノ酸変異が蓄積するとその効果が減弱 する可能性があるが,逆転写過程の開始に必須であるプラ
ウイルスと同様の円錐状のコアを形成できていることがわ かり,リン酸化 Ser16-Pro17はコアの形成ではなく脱殻過程 に関与することが示唆された.そこで,Stremlau らが構 築した fate-of-capsid assay44)により脱殻過程の評価を行っ たところ,S16A/P17A 変異体ウイルスは脱殻しにくいと いう結果が得られ,S16A/P17A 変異体ウイルスは脱殻過 程が進行しないことで感染性が低下することが明らかと なった. CAコアは,ウイルス粒子内では安定な構造体として存 在し,標的細胞内において崩壊することから,脱殻過程に は標的細胞内に存在する因子,つまり宿主性因子が必要で あると考えられる.そこで,リン酸化 Ser16-Pro17を認識す る宿主性タンパク質に着目した.その結果,ペプチジルプ ロリルイソメラーゼ Pin1 が,リン酸化 Ser16-Pro17を認識
していることを突き止めた.Pin 1 は,cis-trans 異性化反
応触媒する酵素である45).我々の結果は,CA のリン酸化
Ser16-Pro17が Pin1 の基質であること,そして脱殻は Pin1
の cis-trans 異性化反応によって制御されることを示してい る(Fig. 4A). 次に我々は,Ser16のリン酸化機構について検討を進め た.まず CA の一次配列から予測されたキナーゼ候補とリ ン酸化 Ser16-Pro17を含む前後 12 残基の直鎖ペプチドを反 応させ,リン酸化反応を質量分析によって追跡した.そ の結果,Extracellular signal-regulated kinase 2(ERK2)が HIV-1 CAの Ser16をリン酸化する候補として得られた.実 際に ERK2 をノックダウンした細胞由来のウイルス粒子 内 CA ではリン酸化 Ser16-Pro17のシグナルが消失してお り,ERK2 が Ser16にリン酸化を付与する酵素であること が明らかとなった.次に,ERK2 による Ser16のリン酸化は, ウイルス産生細胞内で起こるのか,それともウイルス粒
Fig. 4 Viral uncoating mechanism mediated by phosphorylation at Ser16 of CA
(A) Schematic illustration of Pin1-dependent uncoating. The serine residue in the Ser16-Pro17 motif of a CA protein in the CA
core is phosphorylated and peptidyl-prolyl isomerase Pin1 specifically recognizes the phosphorylated Ser16-Pro17 motif of the
HIV-1 core. Finally, Pin1 prolyl isomerase activity is required for the uncoating of the HIV-1 core. (B) Schematic illustration of phosphorylation mechanism of Ser16. Activated ERK2 is packaged into viral particles and catalyzes the phosphorylation of Ser16
約 100 nm という限られた空間であるため,ウイルス複製 を制御するプロテオームが選択的に取込まれていると考え られる.このような背景から,感染性 HIV-1 粒子を用いた プロテオーム解析のデータは,ウイルスの複製に深く関わ る宿主細胞由来のタンパク質や宿主翻訳後修飾システムを 明らかにし,ゲノム情報からでは解明できないプロテオー ムレベルでの「真のウイルスの姿」を捉えるために重要で ある.今後技術開発がさらに発展し,HIV-1 複製の本態の 解明,そして根治療法の開発にプロテオーム解析がもっと 貢献できる日が来ることを期待する. 謝 辞 本研究の遂行にあたり,科学研究費助成事業および熊薬 研究助成会から助成を受けた. 著者らに開示すべき利益相反状態は無い. 文 献
1) Alkhatib G, Combadiere C, Broder CC, et al. CC CKR5: a RANTES, MIP-1alpha, MIP-1beta receptor as a fusion cofactor for macrophage-tropic HIV-1. Science. 1996;272(5270):1955– 1958.
2) Cocchi F, Devico AL, Garzinodemo A, et al. Identification of rantes, Mip-1-alpha, and Mip-1-beta as the major Hiv-suppressive factors produced by Cd8(+) T-cells. Science. 1995;270(5243):1811–1815.
3) Deng HK, Liu R, Ellmeier W, et al. Identification of a major co-receptor for primary isolates of HIV-1. Nature. 1996; 381(6584):661–666.
4) Dragic T, Litwin V, Allaway GP, et al. HIV-1 entry into CD4(+) cells is mediated by the chemokine receptor CC-CKR-5. Nature. 1996;381(6584):667–673.
5) Feng Y, Broder CC, Kennedy PE, et al. HIV-1 entry cofactor: Functional cDNA cloning of a seven-transmembrane, G protein- coupled receptor. Science. 1996;272(5263):872–877.
6) Sheehy AM, Gaddis NC, Choi JD, et al. Isolation of a human gene that inhibits HIV-1 infection and is suppressed by the viral Vif protein. Nature. 2002;418(6898):646–650.
7) Mangeat B, Turelli P, Caron G, et al. Broad antiretroviral de-fence by human APOBEC3G through lethal editing of nascent reverse transcripts. Nature. 2003;424(6944):99–103. 8) Arthur LO, Bess JW, Jr. Sowder RC 2nd, et al. Cellular proteins
bound to immunodeficiency viruses: implications for patho-genesis and vaccines. Science. 1992;258(5090):1935–1938. 9) Ott DE. Cellular proteins in HIV virions. Rev Med Virol.
1997;7(3):167–180.
10) Chertova E, Chertov O, Coren LV, et al. Proteomic and bio-chemical analysis of purified human immunodeficiency virus type 1 produced from infected monocyte-derived macro-phages. J Virol. 2006;80(18):9039–9052.
11) Misumi S, Fuchigami T, Takamune N, et al. Three isoforms of cyclophilin A associated with human immunodeficien-cy virus type 1 were found by proteomics by using two- dimensional gel electrophoresis and matrix-assisted laser 子内で起こるのかを検討した.その結果 ERK2 は,Pr55gag
ではなくプロセシングの過程で Ser16をリン酸化すること,
ERK2の活性化フォームおよび ATP が感染性 HIV-1 粒子 内に取り込まれていることが明らかとなり,ERK2 による Ser16のリン酸化はウイルス粒子内で起こることが示され た(Fig. 4B). 以上は感染性 HIV-1 粒子を用いてプロテオーム解析を実 施したからこそ明らかにできたウイルス性タンパク質の翻 訳後修飾の一例である.脱殻過程は,観察することが難しく, 円錐形の CA コアを試験管内で構築できないなどの理由で HIV-1の複製過程において最も解明が遅れていたが,我々 の研究により脱殻メカニズムの一端を明らかにすることが できた.これまで脱殻過程の理解が不十分であった原因は, ゲノムレベルの解析だけでは分からない翻訳後修飾によっ て脱殻が制御されていたためであり,タンパク質そのもの を捉える重要性を我々の結果は示している.我々は,ウイ ルス粒子内 ERK2 の活性化や Pin1 によるリン酸化 Ser16
-Pro17の認識を阻害することができれば,易変異原性を有 するウイルス性タンパク質を標的とした従来の治療戦略と は異なる方法で,薬剤耐性ウイルスの出現の問題を克服し た治療が可能になると考えている. 5 結 論 本論文では,感染性 HIV-1 粒子を試料に用いたプロテ オーム解析によって明らかにできた HIV-1 複製制御機構 についてまとめた.HIV-1 感染症が世界で取り沙汰され約 40年が経過しようとする中で,これまでに行われてきた 世界中の膨大な研究をもってしても,HIV-1 感染に対する 根治療法を見出せていない.周知のように HIV-1 は単独で は増殖できず,ウイルスが複製するためには,宿主との共 存,つまり宿主細胞の代謝機構やエネルギー産生機構に依 存しなければならない.したがって,HIV-1 複製制御機構 を理解し根治療法の開発につなげるためには,宿主細胞が どのような仕組みで動作しているかを理解することが欠か せない.2000 年代前半にはヒトのゲノム配列の解読が完 了し,ポストゲノム時代が到来した.ゲノムはプロテオー ムに変換され,その中で構築された生命システムが動作し, 細胞が固体内でその役割を果たしている.しかしながら, 個々の細胞において各々の遺伝子は常に一定であるのに対 して,タンパク質は極めて多様な機能や性質を有している だけで無く,広い動的な存在様式を持っており,その状態 が刻一刻と変化することで,細胞の機能が維持されている. そのため,ウイルスの複製そのものの理解においても,プ ロテオームが織りなすタンパク質間分子ネットワークの構 造や機能を明らかにすることが極めて重要であり,プロテ オーム解析をしなければ明らかにできないウイルスの複製 機構が数多く有るといえる.加えて,HIV-1 粒子内は直径
tRNA(Lys)(3) concentration in human immunodeficiency vi-rus type 1 upon its annealing to viral RNA, GagPol incorpora-tion, and viral infectivity. J Virol. 2002;76(18):9096–9102. 27) Guo F, Cen S, Niu M, et al. Specific inhibition of the
synthe-sis of human lysyl-tRNA synthetase results in decreases in tRNA(Lys) incorporation, tRNA(3)(Lys) annealing to viral RNA, and viral infectivity in human immunodeficiency virus type 1. J Virol. 2003;77(18):9817–9822.
28) Isel C, Ehresmann C, Marquet R. Initiation of HIV reverse transcription. viruses. 2010;2(1):213–243.
29) Cen S, Khorchid A, Javanbakht H, et al. Incorporation of Lysyl-tRNA synthetase into human immunodeficiency virus type 1. J Virol. 2001;75(11):5043–5048.
30) Cen S, Javanbakht H, Niu MJ, et al. Ability of wild-type and mutant lysyl-tRNA synthetase to facilitate tRNA(Lys) incor-poration into human immunodeficiency virus type 1. J Virol. 2004;78(3):1595–1601.
31) Kovaleski BJ, Kennedy R, Hong MK, et al. In vitro charac-terization of the interaction between HIV-1 Gag and human lysyl-tRNA synthetase. J Biol Chem. 2006;281(28):19449– 19456.
32) Kovaleski BJ, Kennedy R, Khorchid A, et al. Critical role of Helix 4 of HIV-1 capsid C-terminal domain in interac-tions with human lysyl-tRNA synthetase. J Biol Chem. 2007;282(44):32274–32279.
33) Jenkins JL, Tanner JJ. High-resolution structure of hu-man D-glyceraldehyde-3-phosphate dehydrogenase. Acta Crystallogr D Biol Crystallogr. 2006;62(Pt3):290–301. 34) White MR, Khan MM, Deredge D, et al. A dimer
inter-face mutation in glyceraldehyde 3-phosphate dehydroge-nase regulates its binding to AU-rich RNA. J Biol Chem. 2015;290(7):4129.
35) Frauwirth KA, Thompson CB. Regulation of T lymphocyte metabolism. J Immunol. 2004;172(8):4661–4665.
36) Chang CH, Curtis JD, Maggi LB, et al. Posttranscriptional control of T cell effector function by aerobic glycolysis. Cell. 2013;153(6):1239–1251.
37) Kishimoto N, Iga N, Yamamoto K, et al. Virion-incorporated alpha-enolase suppresses the early stage of HIV-1 reverse transcription. Biochem Biophys Res Commun. 2017;484(2):278– 284.
38) Mouree KR, Kishimoto N, Iga N, et al. Virion-packaged pyruvate kinase muscle type 2 affects reverse transcrip-tion efficiency of human immunodeficiency virus type 1 by blocking virion recruitment of tRNA(Lys3). Biol Pharm Bull. 2018;41(4):612–618.
39) Ganser BK, Li S, Klishko VY, et al. Assembly and anal-ysis of conical models for the HIV-1 core. Science. 1999;283(5398):80–83.
40) Li S, Hill CP, Sundquist WI, et al. Image reconstructions of helical assemblies of the HIV-1CA protein. Nature. 2000;407(6802):409–413.
41) Zhao GP, Perilla JR, Yufenyuy EL, et al. Mature HIV-1 capsid structure by cryo-electron microscopy and all-atom molecular dynamics. Nature. 2013;497(7451):643–646.
42) Freed EO. HIV-1 Gag proteins: Diverse functions in the virus life cycle. Virology. 1998;251(1):1–15.
43) Pettit SC, Moody MD, Wehbie RS, et al. The P2 Domain of desorption ionization-time of flight mass spectrometry. J
Virol. 2002;76(19):10000–10008.
12) Kishimoto N, Onitsuka A, Kido K, et al. Glyceraldehyde 3-phosphate dehydrogenase negatively regulates human immunodeficiency virus type 1 infection. Retrovirology. 2012;9:107.
13) Kishimoto N, Onitsuka-Kishimoto A, Iga N, et al. The C-terminal domain of glyceraldehyde 3-phosphate dehydro-genase plays an important role in suppression of tRNALys3 packaging into human immunodeficiency virus type-1 parti-cles. Biochem Biophys Rep. 2016;8:325–332.
14) Misumi S, Inoue M, Dochi T, et al. Uncoating of human im-munodeficiency virus type 1 requires prolyl isomerase pin1. J Biol Chem. 2010;285(33):25185–25195.
15) Dochi T, Nakano T, Inoue M, et al. Phosphorylation of human immunodeficiency virus type 1 capsid protein at serine 16, required for peptidyl-prolyl isomerase-dependent uncoating, is mediated by virion-incorporated extracellular signal- regulated kinase 2. J Gen Virol. 2014;95:1156–1166.
16) Dochi T, Akita A, Kishimoto N, et al. Trametinib suppress-es HIV-1 replication by interfering with the disassembly of human immunodeficiency virus type 1 capsid core. Biochem Biophys Res Commun. 2018;495(2):1846–1850.
17) O’Farrell PH. High resolution two-dimensional electrophore-sis of proteins. J Biol Chem. 1975;250(10):4007–4021. 18) Volker KW, Reinitz CA, Knull HR. Glycolytic enzymes and
assembly of microtubule networks. Comp Biochem Physiol B Biochem Mol Biol. 1995;112(3):503–514.
19) Ishitani R, Chuang DM. Glyceraldehyde-3-phosphate dehy-drogenase antisense oligodeoxynucleotides protect against cytosine arabinonucleoside-induced apoptosis in cultured cer-ebellar neurons. Proc Natl Acad Sci U S A. 1996;93(18):9937– 9941.
20) Fermani S, Ripamonti A, Sabatino P, et al. Crystal structure of the non-regulatory A(4 )isoform of spinach chloroplast glycer-aldehyde-3-phosphate dehydrogenase complexed with NADP. J Mol Biol. 2001;314(3):527–542.
21) Glaser PE, Han X, Gross RW. Tubulin is the endogenous in-hibitor of the glyceraldehyde 3-phosphate dehydrogenase isoform that catalyzes membrane fusion: Implications for the coordinated regulation of glycolysis and membrane fusion. Proc Natl Acad Sci U S A. 2002;99(22):14104–14109.
22) Schultz DE, Hardin CC, Lemon SM. Specific interaction of glyceraldehyde 3-phosphate dehydrogenase with the 5’-nontranslated RNA of hepatitis A virus. J Biol Chem. 1996;271(24):14134–14142.
23) Petrik J, Parker H, Alexander GJ. Human hepatic glyceralde-hyde-3-phosphate dehydrogenase binds to the poly(U) tract of the 3’ non-coding region of hepatitis C virus genomic RNA. J Gen Virol. 1999;80(Pt12):3109–3113.
24) Choudhary S, De BP, Banerjee AK. Specific phosphorylated forms of glyceraldehyde 3-phosphate dehydrogenase associ-ate with human parainfluenza virus type 3 and inhibit viral transcription in vitro. J Virol. 2000;74(8):3634–3641.
25) Huang Y, Mak J, Cao Q, et al. Incorporation of excess wild-type and mutant tRNA(3Lys) into human immunodeficiency virus type 1. J Virol. 1994;68(12):7676–7683.
human-immunodeficiency-virus type-1 gag regulates sequen-tial proteolytic processing and is required to produce fully in-fectious virions. Journal of Virology. 1994;68(12):8017–8027. 44) Stremlau M, Perron M, Lee M, et al. Specific recognition and
accelerated uncoating of retroviral capsids by the TRIM5 alpha
restriction factor. P Natl Acad Sci U S A. 2006;103(14):5514– 5519.
45) Ryo A, Liou YC, Lu KP, et al. Prolyl isomerase Pin1: a catalyst for oncogenesis and a potential therapeutic target in cancer. J Cell Sci. 2003;116(5):773–783.
Various Functions of Host Proteins Revealed by Proteome Analysis Using
Infectious HIV-1 Particles
Naoki Kishimoto*, Shogo Misumi
*E-mail: [email protected]
Department of Environmental and Molecular Health Sciences, Faculty of Medical and Pharmaceutical Sciences, Kumamoto university, 5-1 Oe-honmachi, chou-ku, Kumamoto 862-0973, Japan
(Received on September 12, 2020; Revised on September 25, 2020; Accepted on September 28, 2020) Human immunodeficiency virus type 1 (HIV-1) infects immune cells including CD4+ T cells. Previous studies
demonstrated that host proteins in immune cells regulate HIV-1 replication. However, it has not yet been clarified how virion-packaged host proteins affect viral replication. Thus, the identification of virion-packaged host proteins and the elucidation of their role in HIV-1 replication are among the important approaches to clarifying further the HIV-1 replication process. Furthermore, because viral proteins utilize intracellular post-translational modification enzymes to acquire protein diversity, the identification and functional analysis of post-translational modifications in viral proteins are also important for elucidating the mechanism of HIV-1 replication. In this review, among the novel HIV-1 replication mechanisms that we have discovered by proteome analysis using infectious HIV-1 particles, we present the inhibition of HIV-1 replication by intravirion glyceraldehyde-3-phosphate dehydrogenase and the importance of post-translational modification on viral capsid protein.