ジャガイモの教材開発研究
65
0
0
全文
(2) 第. ■ 章 教材としてのジャガイモのさし木の基礎的研究. 1. 材料と 方法 1)材料. 2)栽培 3)さし木の方法 4)さし穂の採取時期と位置 5)さし穂の重さと葉面積の測定. 6)光量の調節 7)日長の調節. 2 糸吉果. 6. 1)さし木による塊茎の形成および腋芽の伸長 2)塊茎の形成とさし穂重の増減 3>葉柄の肥大とさし穂重の増減 4)さし木期間中のさし穂重の増減 5)葉面積と塊茎重との関係 6)光量の違いによる葉面積と塊茎重との関係 7)日長の違いによる葉面積と塊茎重との関係. 3 考察 1)さし木の採取適期 2)さし木の最適条件 3)葉面積と光合成との関係. ジ. じめむご. 一123. 1ま. ぺ 乙. 目次. 11.
(3) ページ. 第2章ジャガイモの器内培養 一. 材料と方法. ..”....“..................一一一一一一一一一一一一一一一一一一一一一一一. @1 6. 17. 1)材料 2)塊茎からのカルス 3)種子からの無菌植物の育成 4)環境条件(日長、温度)の調節. 19. 2 糸吉果 1)カルスの形成と器官の誘導 2)器官培養による塊茎の形成 3)塊茎形成の最適条件. 。わ辞用・. 3お謝引回. 考察. 22. り tこ. 24 25. 弓1用文献. 26. 図版お. よび説明一一一一一一29.
(4) はじめに. ジャガイモは北海道から沖縄まで全国的に栽培されている作物で、. 栽培が容易なため、古くかち小学校の教材としてとりあげられてき ている。今まで、ジャガイモは、 “いも”からの栽培が中心であっ. た。しかし、これだと栽培に3∼4ヶ月かかり、しかも途申でいも の肥大の状態を観察することができないeそこで、いもの肥大の様 子を観察でき、しかも短期間で行なえる方法がないかと考え、本研 究では、さし木法と無菌培養法の導入を試みた、. さし木法は、茎葉をさし隠し、葉腋の芽を肥大させ、塊茎とする. 方法である。これだと1∼2週間で塊茎の形成の様子を観察するこ とができる。. 無菌培養法は、はじめ“いも” (塊茎)から柔組織を切り出し行. なった。しかしこの方法では、カルスは形成されたが芽の形成は見 ちれなかった。そこで、ジャガイモの種子から無菌ジャガイモを作 り、この谷底培養を行ない塊茎の誘導を試みた。これだと糖を栄養 分として与えることができるため、試験管内で塊茎の形成の過程が より早く、しかも手にとって観察できるのではないかと考えて行なっ た。. しかし、この無菌培養法は予想に反して、試みた条件では塊茎を 形成するまでに露地栽培以上の期間を要した。このため器内での塊 茎形成は、興味・関心の点ではすぐれているが、実験系として教材 にすることは問題があると結論した。このため、本研究においては 教材を指向したさし木の研究を中心に行なった。. 1.
(5) 第1章 教材としてのジャガイモのさし木の基礎的研究. さし木は植物の無性的な栄養繁殖法として、接ぎ木と共に古くか. ち行なわれ、ジャガイモのさし木の歴史も古い(鴨chting 188 7)。最近では新しい晶種や無病種いもの増殖の方法として、さし. 木がなされている(Cole and Wright 1976,Lauer 1977,. 松本 1981、大石・田中 1986>。 ジャガイモは栽培が容易なので教材として広く使われているが、 いもを用いた栽培が主体である。これを教材として用いるには、種. いもの植付けかち収穫まで最低3ヶ月を要し、結果の出るまでの興 味関心の維持が困難である、ところがさし木を用いると、時期が適. 当だと1週間で塊茎の形成が見ちれる。そこで、筆者はこのさし木 法により、形成される塊茎とさし穂の葉面積や当たる光の量とが相 関していれば、光合成を定量的にとらえた指導ができるのではない かと考え、この研究を行なった。. 一2一.
(6) 材料と方法. 1)材料 実験に用いた種いもの品種と入手先は次の通りである。 ジャガイモ(Solanum tuberosum). 品種名 入手先 入手年月日. だんしゃく(春植え、極早生) 社町農協 63年3月 メークイン( 〃 、中生) 〃 . l」. せとゆたか( 〃 、 〃.) 〃 62年9月 でじま (秋植え、晩生) 佐賀県中川種物店 63年9月. 2)栽培 さし穂用のジャガイモの栽培は、兵庫教育大学の圃場および温室. 内に置いたプランターで行なった。種いもは1週間程浴光させ、大. きいものは縦切りにした。切断後冷暗所に1∼2晩おき、圃場にう. ね幅70c皿、株間30c孤、深さ7∼8cmで植付けた。肥料は、植付 け時に油かすを2握りずつ株問に与えた。除草は適時行なった。培 土は、出芽後2週間に行なった。秋の栽培では、殺菌剤で種いもを. 消毒してから植付けし、殺菌剤、殺虫剤を出芽後10日から1週間. 毎に3回散布した。春作は、3月28日に植付けを行ない4月24 日前後に出芽した。秋作は、9月28日に植付けを行ない10月1 0日前後に出芽した。. 一3一.
(7) 3)さし木の方法 さし穂は、葉芽ざしを基本とし、葉や腋芽の数を変えて行なった。. さし穂の基部(腋芽)を2∼3cmの深さで、葉が重なちないように. バーミキ・ユライトを入れたプランター(縦21.5cmX横64c皿X 高さ18c皿)にさした(図 1)。さしたばかりのプランターは、. 1∼2日直射日光の当たちない室内に置いた後、温室に移し、1日 に二回の割で潅水した。. 4)さし穂の採取時期と位置 さし穂は、春作では開花期く出芽後約35日)以降、秋作では出 芽後22日以降の茎葉からとった。葉の位置は、頂芽である花芽を. 0として下へ葉を数え、4枚目から8枚目の葉で腋芽の伸長してい ないものを用いた。側枝の伸長のよいものについては、側枝の先端 の葉かち4枚目以降の葉を用いた。. 5)さし穂の重量と葉面積の測定 さし穂はあちかじめ生重量を求めてかち、さし木を行なった。さ. し穂の生重量は上皿電子天秤(エー・アンド・デイ社EW−60B). で10mgの単位まで量った。さし木後一一定期間(春8∼10日、 秋14∼30日)培養し、さし穂の生重量、葉軸以外の葉身、葉軸 と葉柄と茎、塊茎の生重量と乾重量、葉面積を測った。乾重量は、. 乾熱滅菌器(ミタムラ理研工業16−35)を用い、110℃で8. 時間乾燥させ、冷却後直示天秤(島津NL−200TP)で1mg. 一一一. @4 一.
(8) の単位まで量った。葉面積の測定は、合成紙感光紙(コクヨGP3 8)を用いて行なった。さし穂の葉軸を除いた葉身を合成紙上に展 開し、感光し定着を行ない(図 2)、合成紙を切り抜いて重量を 量り、その重量かち葉面積を算定した。合成紙の重量は、上皿電子. 天秤(島津EB−50)で1mgの単位まで量った。 6)光量の調節 光量の調節は、プランターを遮光ネットで覆い、ネットの枚数を. 変えて行なった(図 3)。光量は、晴天と曇天の日の正午に各プ ランターの直上の照度をルクスメーター(ミノルタデジタル照度計. T−1M)でそれぞれ10ヶ所測り、その平均値を求め、ネットで 覆わないものを100として求めた。 7)日長の調節 日照時間は、プランターをダンボール類で覆い調節した(図 4>。. 6月は0、8、14時間の3区を、11月は0、3、6、9時間の 4区を設けて行なった。. 一5一.
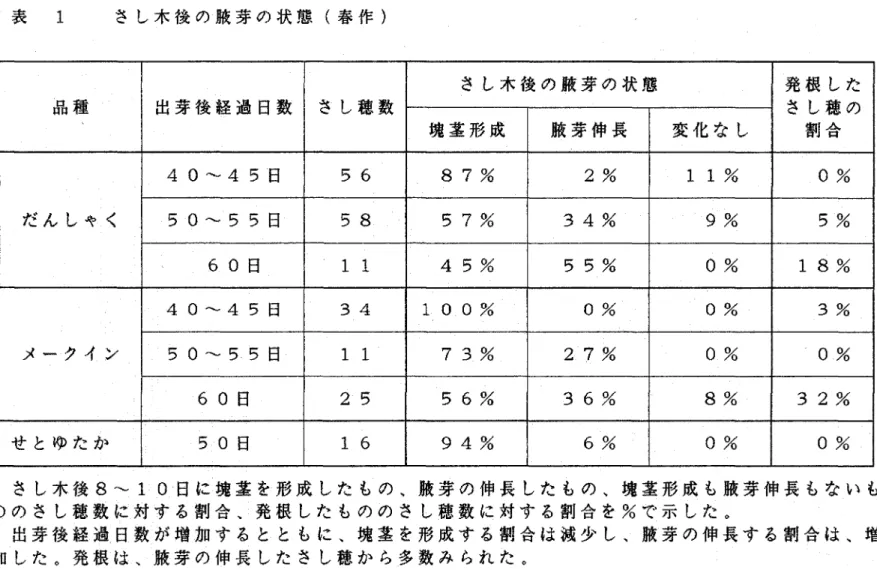
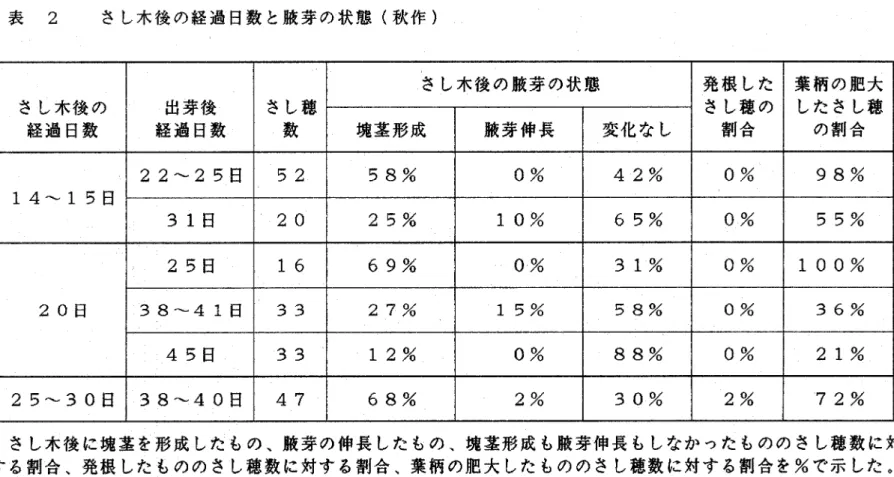
(9) 結果. 1)さし木による塊茎の形成および腋芽の伸長 春作のジャガイモを用いたさし木実験において、さし木鼻の塊茎 の形成および腋芽の伸長の割合は、共に出芽後日数とともに変化す. る傾向がみちれた。出芽後40∼45日は塊茎を形成する割合が高 いが、順に低下し、出芽後60日になると塊茎形成率はだんしゃく で50%以下になる。これに対し腋芽の伸長する割合は逆に加齢と. ともに高くなり、出芽後40∼45日ではほとんど伸長しなかった. ものが、出芽後60日ではだんしゃくで55%、メークインで36 %が伸長するようになった。発根は、腋芽の伸長したさし穂から多. 数みちれた(表 1、図 5>。出芽後70日のメークインの伸長 した側枝からとったさし穂をさし訳すると、出芽後日数の少ないも. のと同様に塊茎の形成率が100%となった。またこれは、腋芽の 伸長、発根共に全くみちれなかった。. 秋作のさし木実験においても、出芽後の日数が増え、加齢が進む と塊茎形成率が低下する傾向がみちれた。しかし、春作の実験でみ ちれたように、塊茎形成率の低下と共に腋芽の伸長する割合は高く ならず、塊茎形成も腋芽伸長もしないものの割合が高かった。さし 木日数が増加すると塊茎を形成する割合が高くなった。腋芽の伸長 したさし穂の割合はきわめて低く、最も高率のものでも15%であっ た。発根もほとんどみちれなかった(表 2)。. 春作の実験においては、頂芽かち数えて1∼3枚目の葉をさし表 した時に葉柄の基部の肥大がみちれたが、秋作の実験においては、. 一6一.
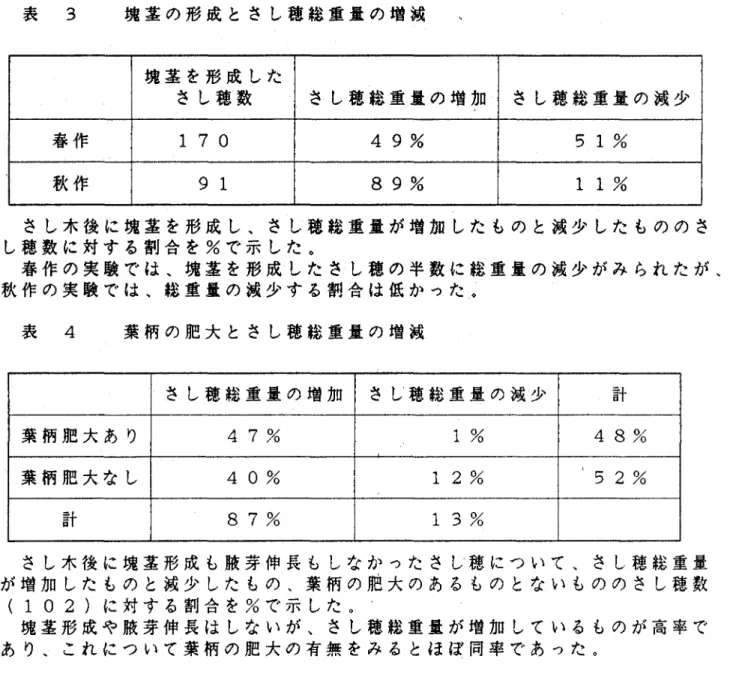
(10) 葉柄の基部の肥大しているものが高率でみられ、塊茎を形成したさ. し穂の87%で葉柄の肥大がみられた(図 6)。 2)塊茎の形成とさし穂総重量の増減 塊茎を形成したさし穂をさし木前後の総重量でみると、春作の実. 験では、塊茎を形成したさし穂の半数(51%)にさし穂総重量の 減少がみちれた。秋作の実験では、塊茎を形成したさし穂は、さし. 穂総重量の減少の割合が11%と低かった(表 3)。 3>葉柄の基部の肥大とさし穂総重量の増減 秋作の実験において、塊茎形成や腋芽伸長をしなかったさし穂に ついて葉柄の基部の肥大とさし穂総重量の増減との関係をみると、. 塊茎形成や腋芽伸長はしないが、さし穂総重量が増加しているもの が87%と高率であった。これについて葉柄の肥大の有無をみると、. ほぼ同率であった(表 4)。葉柄が肥大しているさし穂は、さし 木後のさし穂総重量の増加の平均が0.52±0.036gであるのに対し、. 肥大していないさし穂は、0。25±0.024gであり、葉柄が肥大して いるものは肥大していないものより増加重量が多かった。. 4)さし木鼻間中のさし穂総重量の増減. 秋作の実験で、さし穂総重量をさし木後2∼4日ごとに測定した ところ、さし穂総重量が重いものでも軽いものでも、同様な増減を. 一7一.
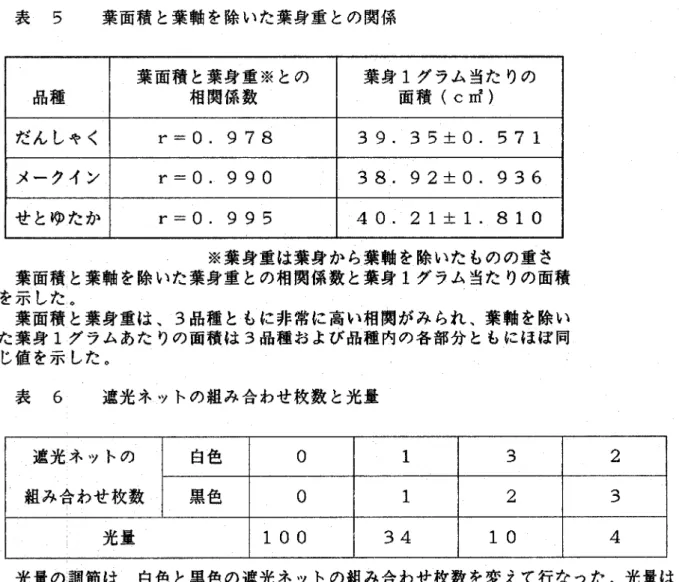
(11) 示した(図 7>。しかし、さし穂総重量が重い程、増減の幅が広 い傾向がみちれた。さし穂総重量はさし木後6日までは減少し、そ れ以後増加しはじめた。塊茎の形成は早いものでさし木後6日にみ. ちれた。さし木盃3∼4週間すると、さし穂の葉先の枯れや黄変が みちれるようになった。. 5)葉面積と塊茎重との関係 だんしゃく、メークイン、せとゆたかの3品種を用い、さし木後 に総重量が増加したさし穂で、葉面積と形成された塊茎重との関係. をみると、3品種とも葉面積が大きくなると塊茎重も増加し、比例 傾向がみちれた。だんしゃくとメークインでは、葉面積が2倍にな. ると塊茎重もおよそ2倍になったが、せとゆたかでは、1。5倍程 度であった(図 8)。合成紙感光紙で求めた葉軸を除いた葉身の 面積と重量との関係をみると、葉面積と葉身重は3品種共に非常に 高い相関がみられた(r=・ O. 978∼0.995)。葉軸を除いた葉身1グ. ラムあたりの面積は、各品種および品種内の各部分ともに38,92±0. .936∼40.21±L810c㎡でほぼ同じ値であった(表 5)。葉軸を. 除いた葉身1グラムあたりの面積を40c㎡として、葉身重から求 めた葉面積と塊茎重との関係をみると、合成紙かち求めた葉面積と. 塊茎重との関係と同様な比例傾向がみちれた(図 9)。葉身重か ら求めた葉面積と塊茎重との比例傾向は、出芽後経過日数の異なる. もの(図 10)や、さし木後経過日数の異なるものにおいても同 様にみちれた。葉軸を除いた葉身重と塊茎重は生重量で関係を求め たが、乾燥重量で行なっても同様な比例傾向を示した(図 11)。. 一8一.
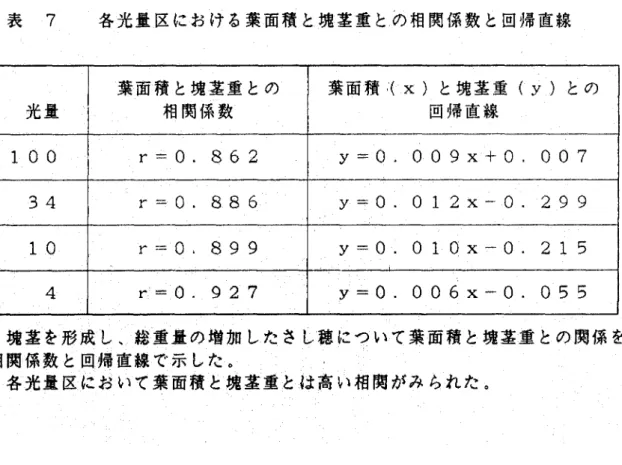
(12) 頂芽を0として葉を数え、4枚目かち8枚目の位置の異なる葉で葉 面積と形成された塊茎重との関係を調べたが、いずれも比例傾向を. 示し、葉の位置による違いはみられなかった(図 12)。 秋作のさし木実験で、合成紙から求めた葉面積と葉軸を除いた葉 身重との関係を調べたところ、春作の結果と同様に非常に高い相関 (r=・ e.989) がみられた。しかし、葉身1グラムあたりの面積は. 31.45±0.368c㎡となり、春作の3品種よりこの値は低かった。葉. 軸を除いた葉身1グラムあたりの面積を31c㎡として、さし予後 に総重量が増加したさし穂について葉身重かち求めた葉面積と塊茎. 重との関係を調べると、比例傾向はみられなかった(図 13)。 秋作の実験では、出芽後経過日数やさし木曜経過日数の異なるもの においても、葉面積と塊茎重とは比例傾向を示さなかった。. 6)光量の違いによる葉面積と塊茎重との関係 光量の調節は、白色と黒色の遮光ネットを組み合わせ、4区で行 なった(表 6)。各光量区において、塊茎を形成し総重量の増加 したさし穂について、葉面積と塊茎重との関係を調べると(図14). 重宝量区とも葉面積と塊茎重との間に高い相関がみちれた(r=o.. 862∼0.927、表7)。そこで各光量区における葉面積と塊茎重と. の関係を回帰直線で比較した結果、光量100かち10までは、葉 面積に対する塊茎重はほとんど差がなかったが、光量が4まで下が. ると差がみちれるようになった(図 15>。 秋作の実験では、遮光ネットでプランターを覆い20日間培養を. 行なったが、塊茎を形成したさし穂は48本中3本で形成率は6%. 一9一.
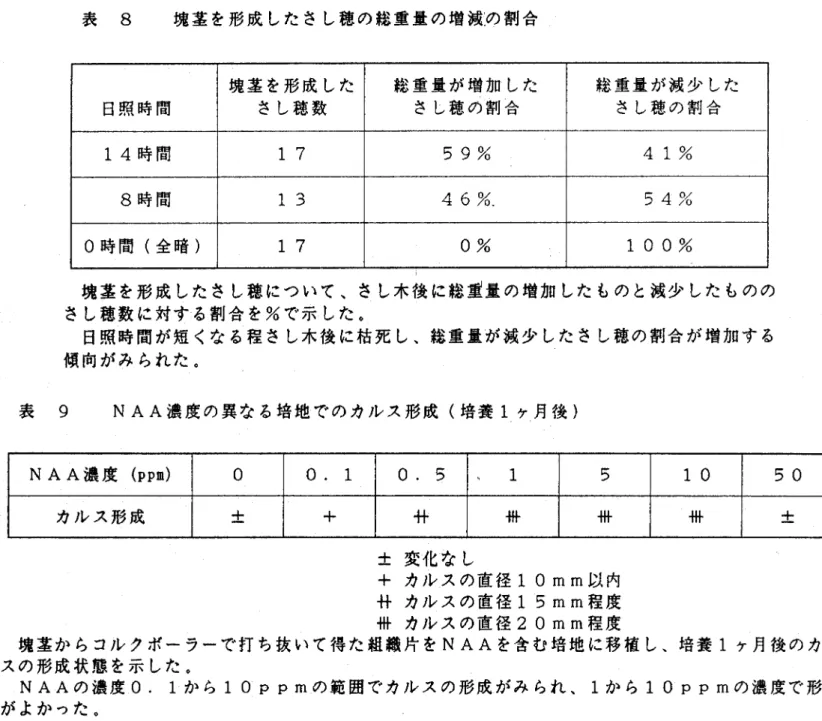
(13) と低く、光量の違いによる葉面積と塊茎重との関係をみることはで きなかった。. 7)日長の違いによる葉面積と塊茎重との関係 春作の実験で、日長を変えた3区とも塊茎の形成がみちれ、全暗. にして光を当てなかった区でも、22本中17本のさし穂に塊茎の 形成がみちれた。日照時間が短くなる程さし木後に枯死し、総重量. の減少したさし穂の割合が増える傾向がみられた。特に全暗く0時 間)区では、黄変し枯死するさし穂が多く、塊茎を形成したさし穂. は全て、さし木後に総重量が減少した(表 8)。日長の異なる2. 区(8時間と14時間)では、葉面積と塊茎重との関係に有意差は みちれなかった。. 秋作の実験では、さし木により塊茎を形成したさし穂の率は32 %と低く、腋芽の伸長したものの率も2%と低かった。全暗にした ものは、葉が黄変、枯死し、塊茎の形成もみちれなかった。残る3. 区く3時間、6時間、9時間)において、形成された塊茎は、重量 がO.01∼0.04gの小型のものが多く、日照時間の増加により単位葉 面手あたりの塊茎重が増加する傾向は認められなかった(図 16)。. 10一.
(14) 考察. 1)さし穂の採取適期 さし木により高率に塊茎を形成させるためのさし穂の採取適期は、. 春作の場合は、開花始期(出芽後約35日)かち(極)早生品種で. 出芽後45日まで、中生品種で55日までである、中生品種はこれ 以降でも、伸長した側枝から採取すると高率に塊茎を形成すること が明らかになった。. ジャガイモの塊茎の肥大開始期や開花始期は、多くの品種で詳し. い研究がなされている(農山漁村文化協会 1981)。これによ ると塊茎の肥大開始期は、最も早い品種で出芽後約3週間であるが、. これは植付け時期によっても変動がみちれる。しかし、開花始期に. 関しては、いずれの品種も出芽後約5週間で、出芽時期が同じなら. 品種間差異はみちれないという(田口ら 1969)。各品種を用 いて行なった本実験でも、春作での開花始期は出芽後5週間であり、 ほぼ同じ結果が得られている。しかし、開花は全ての株でみちれず、. 出芽後5週間以後の株から採取したさし穂は、開花の有無と関係な く高率で塊茎を形成することから、母株の開花とさし穂の塊茎の形 成とは直接的には関係がないように思われる。すなわち、母株の開 花の有無は光周性によって決まり、さし穂の塊茎の形成は母株の加 齢(エイジ〉によって決まるものと思われる。. さし穂を母株かち採取する位置としては、頂芽(花芽)かち数え. て4枚圏から8枚目までのものが最適であるという結果が得ちれて いる。1枚目かち3枚目までの先端上位にある若い葉は、葉柄基部. 11.
(15) が肥大し、9枚目以下の下位にある老化した葉は、腋芽が伸長して しまい不適当である。このことは、植物ホルモンによって調節され ていることを示している。すなわち、腋芽の伸長はオーキシンによっ て抑制され、ジベレリンによって促進される。したがってこの場合、. オーキシンは先端部分に多く下部に少なく、逆にジベレリンは上部 に少なく下部に多いことが考えられる。中位の葉がさし穂として適 しているということは、オーキシンとジベレリンの濃度によって塊 茎形成が調節されていることを示唆している。. 以上のことかち、教材としてさし穂を採取する場合は、母株のエ. イジが出芽後5週間以後のもの、そして、葉の位置としては、中位 の葉を用いることが適当であることが結論された。春作の場合は、 エイジの代わりに開花始期も目安となることが明らかになった。. 2)さし木の最適条件 ジャガイモのいもは茎の変形したものであり、腋芽を除くと塊茎 の形成はみちれない。したがって、さし木による塊茎形成の観察実 験の材料には、必ず腋芽をつけたさし穂が必要である.これは腋芽 を切除したり、葉だけをさし:本すると、発根はするが塊茎は形成さ れないことかちも明ちかである。. 光は、塊茎の形成自体には直接には関わっていないものと思われ る。このことは、斜光下でも暗黒下でも塊茎が形成されることから 明らかである。しかしこの場合は、光合成が行なわれないためにさ し穂中の養分が使われ、さし木後の塊茎を含むさし穂全体の重量は、. さし木前の重量より減少することになる。さし木により光合成量を. 12一.
(16) 塊茎の重量でみようとする場合には、光を当てる必要がある。 温度は、生育にも塊茎形成にも影響している。ジャガイモの葉の. 光合成能力の最大値は、葉齢の老若や照度の大小にかかわらず20. ℃付近にあるとされている(Winkler,1971)。これより高温 でも低温でも光合成能力は低下し、極端な低温(6℃〉では茎葉の. 成長が停止するという(Smith,1968>。さし木においても実 験の適温は、栽培の適温と同じ20∼24℃である。高温でさし穂 の枯死が多かったのは、呼吸量や葉からの蒸散量が増加して光合成 量が減少したためと思われる。低温条件では、枯死は少ないが、塊 茎形成の速度が遅く不揃いで、低温になる程塊茎形成率は減少した。. このため、さし木期間を長くする必要があり、短期間で結果を出す には不適当である。. これちのことから、さし木実験に適する時期は、春作では5∼6. 月にかけての2か月間、秋作では10月の1ヶ月間である。特に日. 中の気温が20∼24℃、夜問の気温が10℃内外の条件が最適で ある。 1. 秋作で気温が低下する時期には、プランターによるさし木実験は 不適当なので、教室の日当たりのよい窓際などで水さしを行なうの もよい。これだと塊茎の形成、腋芽の伸長、ストロンの伸長とその 先の肥大、塊茎かちの芽の伸長などを継続的に観察できる。. 3)葉面積と光合成との関係 ジャガイモの栽培で、最適葉面積指数を上限として、株の葉面積 が増えればいもの収量が増えることは確かめられている(田口ら. 13.
(17) 1969)。さし木を用いるとこれが短期間により正確に求めちれ、 光合成量をある程度まで定量的にとらえるための教材になるのでは ないかと考える。. 最適条件のもとで葉面積とその葉で光合成によって形成された塊 茎重との関係をみると、はっきりとした比例関係がみられる。葉面 積を求める場合、ジャガイモの葉は奇数羽状複葉であり、葉面積を 直接求めることは非常に困難である。しかし、本実験に用いた感光 紙に転写しその紙を切り抜いて重量を量り、それかち葉身の面積を 算出する方法は、小学生にでもできる方法であり優れている。この 葉軸を除いた葉身部分の面積と重量とは、同一条件で栽培した母株 からの葉では、正確に一致し、強い相関がみられる。しかし、葉の 厚さは温度や照度によって多少変わるので、その都度葉軸を除いた 葉身の面積と重量を測定し、これかち算出した値を用いるようにし なくてはならない。. この葉面積と光合成量とが.lk例関係にある条件のもとで、光量を. 変えて光合成量を定量的にみる実験を行なった結果、光量を遮光し て変えた場合も日照時間を調節して変えた場合も、ともにはっきり. とした差はみられなかった。遮光の場合、天空光の照度を1とする. と、25分の1以下の光量にしないと光合成量にはっきりとした違 いがみられず、光量の増加に伴う光合成量の増加を理解する実験の. 系には不適当であった。ジャガイモの葉は、照度が3千かち1万ル クスの間では光量増加にしたがって光合成が急激に盛んになるが、. 2万かち6万ルクスでは増加がゆるやかになり、6万ルクス以上で. はほとんど増加しないという(Burton 1966)。したがってこ れは、3千から1万ルクスの間の、光合成が光量増加にしたがって. 14一.
(18) 盛んになる範囲の光量調節が、遮光ネットではうまくできなかった ことに起因しているものと思われる。. 一15一.
(19) 第2章. ジャガイモの器内培養. ジャガイモのさし木による塊茎形成と平行して器内の組織培養を 行なった。これはさし木法では調節できない培地条件など、全ての 環境条件を調節することによって、教材として塊茎の形成条件や形 成過程を観察できるのではないかと考え行なったものである。 ジャガイモの組織培養は、ウィルスフリーのジャガイモの作出の ための茎頂培養や、カルス組織からの植物体の誘導、形態形成や分. 化に関する研究など既に多くの研究がなされている(Bajaj 19. 87)o 本研究は、いもかちのカルス化とともにジャガイモの種子からの 無菌植物を用いて、上記の塊茎形成条件を調べると同時に、最近高 等学校の教科書にも取り上げちれるようになったバイオサイエンス、. バイオテクノロジーの基礎学習の教材の開発をも意図した。. 16一.
(20) 材料と方法. 1) 材料 ジャガイモ(Solanum tuberosum)の品種としてはだんしゃくと メークインを用いた。種いもは加東郡社町農協より入手した。種子 は、タキイ種苗株式会社より入手したシードポテト ホワイトアン. ドホワイトを用いて行なった(図 17)。 2)塊茎かちのカルス 塊茎は中性洗剤でていねいに洗った後、塩素系漂白剤(ハイター). の10%液に30分浸漬して表面を殺菌した。これをクリーンベン チ内に移し、滅菌蒸留水で2回よく撹拝しながら水洗を行ない、滅 菌シャーレの上に置き、殺菌したメスで厚く皮をむいた。これを再. び5%ハイター液で10分滅菌し洗浄後、滅菌した直径5mmのコ ルクボーラーで軸方向に打ち抜いた。打ち抜いた組織の両端を切除. し、これを長さ2mmの切片にし培地に移植した(図 18)。培 地は、ムラシゲスクーグ培地(MS培地、Murashige and Skoog. 1962)を基本培地とし、これにインドール酢酸(IAA)、α. 一ナフタレン酪酸(NAA)、2,4一ジクロロフェノキシ酢酸 (2,4−D)、およびカイネチンを培地1リットルに0.1∼5 0mgの範囲で加えたものを用いた。オーキシンは主にNAAを用 いた。培養容器としては、100mlの三角フラスコを用い、これ. に培地を40ml分注した。培養は、26℃±1℃の全暗と全明の. 一17一.
(21) 条件で行なった。. 3)種子かちの無菌植物の育成. ジャガイモの種子を2枚重ねたガーゼで包み、5%ハイター液に 5分間浸漬し滅菌後、これをクリーンベンチに移し、クリーンベン. チ内でガーゼごと滅菌蒸留水で2回水洗した、この滅菌種子をMS. 培地およびハイポネックス培地(水11にハイポネックス1g、寒. 天10gを加えたもの)に置床し、26℃±1℃の全明条件下で培 養し無菌植物を得た。増殖は、無菌植物から節部(葉の着生部)を. 含む茎の切片を切り出し、オーキシンを含まないMS培地に移植し. て行なった(図 19、20)。 4)環境条件(日長、温度)の調節 日長の調節は、無菌植物をインキュベーーター内(温度 26℃±. 1℃、照度 3000∼4000ルクス)で培養しながら、照明時 間を変えた3区(24時間、8時間、0時間〉で行なった。. 温度の調節は、連続照明(照度 3000∼4000ルクス)の 条件下で温度の異なるインキュベーターで行なった。同時に三角フ ラスコの下部をアルミホイルで包み、下方かちの光量を少なくした ものや、室内の変温自然無下などでも培養を行なった。. 18一.
(22) 結果. 1)カルスの形成と器官の誘導 塊茎からコルクボーラーで打ち抜いて得た組織片は、移植してか. ち1週間後にオーキシンを含む培地でカルス化した。オーキシンの. 種類は、IAA,NAA,2,4−Dの全てにおいてカルス化がみ られた。NAAは0かち50ppmの濃度で行なったが、培養1ヶ 月後には、0,1かち10ppmの範囲でカルスの形成がみられ、 1から10ppmの濃度で形成がよかった(表 9)。カルスの継 代培養は、NAAlppmを含むMS培地で、明所と暗所で行なっ た。暗所で培養したカルスは自色であるが、明所で培養したカルス. は、クロロフィルの形成がみられ緑色となった(図 21)。NA. Alppmを含む培地上では、長期間培養を行なっても芽や根の分 化はみちれなかった。. カルスから芽や根を誘導する目的で、増殖したカルスをNAAと カイネチンを含む培地に移植すると、カルスの増殖や根の分化がみ ちれたが、長期間培養しても芽の分化はみられなかった。また、塊. 茎から切り出した組織片の、NAAとカイネチンとの組み合わせ培 地での培養でも、カルスを生じ、カルスかち根は分化したが、シュ ートの形成はみちれなかった。. 2)器官培養による塊茎の形成. ジャガイモの種子は、培地に置床後5∼7日で発根がみちれ、7. 一19一.
(23) ∼10日で子葉の展開がみちれた。本葉の展開は更に7∼10日か かった。子葉は倒皮針形で初出の第1葉は丸みを帯びた単葉である. (図 22)。MS培地とハイポネックス培地との差は、第1葉展 開時までの初期生育ではみちれないが、その後の生育はMS培地の 方がよかった。全藤下での培養では、子葉基部からストロンの伸長 がみちれるが、先端に塊茎を形成せずそのまま腋芽として伸長し、. 葉を着けるようになった(図 23>。節部を含む茎の切片を切り だしMS培地に移植すると生育状態のそろった無菌植物を多数つく ることができ、この方法で繁殖させた植物体には、露路栽培でみち れる複葉の形成がみられた。しかし、露路のものに比べると小葉の. 数は少なかった(図 24)。 3)塊茎形成の最適条件 温度を一定にし、照明時間を変えて培養しても、3ヶ月内では塊 茎の形成はみちれなかった。全暗区で培養したものは、茎が白く徒. 長した(図 25)。 光条件を一定にし、温度を10℃から35℃に変えて培養したが、 培養3ヶ月内では塊茎の形成はみちれなかった。茎葉の生育は、培. 養10日後にみると28℃が最もよかった(図 26)。35℃で は、茎葉が枯死した。. 塊茎の形成は、培養4ヶ月以後に、室内の変温自然光下に置いた. ものや、10℃から15℃の低温で、三角プラス1の下部をアルミ ホイルで包んで低照にしたものに多くみちれた。. 塊茎は、腋芽が肥大したものや、芽の基部が肥大したもの(図. 一20一.
(24) 27)、また節部からストnンを生じその先端やストロンの腋芽が. 肥大したものく図 28)などみちれた。長期間(6ヶ月かち10 ヶ月)器内培養を続けると、茎葉の枯しゅうが始まる。この時、培. 養条件のいかんに関わちず塊茎を形成するものがみられた(図 2 9)。更に、形成された塊茎から茎葉を伸ばすものもみちれた(図. 30)e. 一21.
(25) 考察. 器内培養により、塊茎形成の条件や過程の観察を意図して本研究 を行なったが、現段階では器内培養を行なっても、塊茎の形成をさ し木以上に短期間にみることは不可能であることが明らかになった。. ジャガイモのカルスからの不定芽の誘導は、ゼアチンを用いて成 功している例がある(Kikuta,Y. and Y. Okazawa, 1984)。. KikutaらはオーキシンとしてIAAをサイトカイニンとしてゼアチ ンを用い、さちにマニトールを添加することによって不定芽の形成 率をあげている。本研究は形態形成の教材として考えているため、 なるべく安価で入手の容易な試薬のみを用いて行なうことを意図し た。このためサイトカイニンとしてカイネチンのみを用いて行なっ た。. 無菌植物の塊茎の形成は、当初、定芽(腋芽)を含む茎の切片を 糖を含む培地で培養することにより、培地中の糖を短期間に吸収転 流させ、塊茎を形成させることができるのではないかと考えた。こ のため、温度、光条件などを変えて芽を含む節部の培養を行なった が、いずれも短期間で塊茎の形成はみられず、塊茎形成の条件を求 めることは不可能であった。一般のジャガイモ品種は、短日条件下. で塊茎形成が早まるという報告がある(北農試 1971∼73)。 本研究では、短日条件で培養しても全心条件で培養したものに比べ て、塊茎の形成が早まることはなかった。成長点を切り出して培地. に移植し、低温(4℃)弱光下で4ヶ月間培養したものに塊茎の形 成がみられたという報告がある(Taoら 1987)。本研究では、. 節部を移植したものを用いたが、低温(10∼15℃)弱塗下で塊. 一22一.
(26) 茎の形成がみられるまで4ヶ月以上かかった。露路栽培において、 出芽後5週間で塊茎形成がみちれるようになるが、器内培養におい ては塊茎形成を観察するためにこれの3倍以上の期間を必要とする こととなる。. ジャガイモの器内培養は絶えず身近に置いて塊茎の形成部位や形 成過程を観察することができる点で優れている。しかし、塊茎の形 成に時間がかかり過ぎる点で、教材としては問題であり不適当であ ることが明らかになった。. 一23一.
(27) おわりに. ジャガイモのさし木法は、小学生にもでき、しかも2週間という 短期間で結果が得られるので教材として極めて優れている。しかし、. 問題はさし木の時期である。夏の高温期と冬の低温期を避け、春と 秋の気温の適期に、さし穂をとる母株のエイジの適期を・一致させて. おく必要がある。気温の適期に成熟した適期の旧株が得ちれれば、 葉面積と光合成量との相関関係をみることや、器内培養で意図した 塊茎形成の位置や過程の観察が可能である。. 本研究は基礎的なものであり、またこれを実践するまでには至ち なかった。今後この実践を通して更に改良を加え、光合成の指導の 開発研究に取り組んでいきたい。. 一24一.
(28) 謝辞. 本研究は、1987年4月より1988年12月まで兵庫教育大 学自然系生物学教室で行なったものである。終始懇切丁寧にご指導 頂いた植物学教室、山田卓三博士、なちびに渥美茂明博士に謹んで 感謝の意を表します。. また、専門的な立場からご指導下さった北海道大学農学部、吉田 稔助教授に深く感謝の意を表します。. 一25.
(29) 引用文献. 1 . Bajaj, Y. P. S. (198 7>. Biotechnology in Agriculture and Forestry 3 Springer−Verlag. 2 . Burton, W. G. (1966). The Potato. Veenman & Zonen.. 3 . Cole,EF. a.nd N. S.Wright (1976). Propagation of p,otato by ste’m cuttings.. American potato jourfial 44’:3el−304. 41北海道農業試験場.(1971、1972、1973) ばれいしょ試験成績書. 5 . Kikllta, Y. afid’ Y. Okazawa (1984). Control of root and shootbud formatioB from potato tuber tissue cultured in vitro. Physiol,Plant.,61:8’12. 6 . Lauer, F. L (1977). Turbers from leaf−bud cuttings:A.tool for potato seed certification and breeding. programs.. 一26一.
(30) American potato journal 54:457−464. 7.松本工(1981) ばれいしょのさし木繁殖による増殖と病害防除〔1〕 農業および園芸 56:1399−1402. 8 . Murashige,T.,F. Skoog. A revised medium for rapid growth and bloassays with. tabacco tissue cultures. Physiol,Plant.,15:473−497. 9.農山漁村文化協会(1981). 畑作全書 イモ類編 基礎生理と応用技術. 農山漁村文化協会 10.大石 顕一・田中 智(1986) さし木繁殖による無病種芋の高速増殖. ポテトサイエンス6:65−74 1 1 . Smith, O. (1968). Potatoes, Produetion, Storing, Processing.. Avi.. 12.田口 啓作・吉田 稔・中世古 公男・由田 宏一(1969) ばれいしょの生理生態学的研究 第2報 乾物生産について. 一27一.
(31) 北大農付属農場報告.7=33−41. 13.. Tao Guoqing,Yin Weiyi,Gong Guopu, and Cui Cheng (1987). In Vitro Production and Release of Potato Varieties in. China Biotechnology in Agrieulture and Forestry 3:62L79. Springer−Verlag. 14.. Vochting (1887). Pflanzenphysiologie. Verlag von Julius Springen. 15. WiRkler, E. (1971>. Kartoffelbau in Tirol II.Photosynthesevermogen uRd Res−. piration von verschiedenen Kartoffelsorten. Potato Res.14:1−18. 一28一.
(32) 表. 図版および説明.
(33) 表 1. さし木後の腋芽の状態(春作〉. さし木後の腋芽の状態 品種. 出芽後経過日数. ウし穂の. さし歯数. 塊茎形成. だんしゃく. せとゆたか. 腋芽伸長. 変化なし. @割合. 40∼45日. 56. 87%. 2%. 11%. 0%. 50∼55日. 58. 57%. 34%. 9%. 5%. 11. 45%. 55%. 0%. 18%. 40∼45日. 34. 100%. 0%. 0%. 3%. 50∼55日. 11. 73%. 27%. 0%. 0%. 60日. 25. 56%. 36%. 8%. 32%. 50日. 16. 94%. 6%. 0%. 0%. 60日. メークイン. 発根した. さし木後8∼10日に塊茎を形成したもの、腋芽の伸長したもの、塊茎形成も腋芽伸長もないも ののさし同数に対する割合、発根したもののさし同数に対する割合を%で示した。 出芽後経過日数が増加するとともに、塊茎を形成する割合は減少し、腋芽の伸長する割合は、増 加した。発根は、腋芽の伸長したさし穂から多数みられた。.
(34) 表 2 さし木後の経過日数と腋芽の状態(秋作). さし木牌の腋芽の状態 さし木後の. o過日数. 14∼15日. 20日. 25∼30日. 出芽後. さし穂. 塊茎形成. o過日数. 腋芽伸長. 変化なし. 発根した. 葉柄の肥大. ウし穂の. オたさし穂. @割合. @の割合. 22∼25日. 52. 58%. 0%. 42%. 0%. 98%. 31日. 20. 25%. 10%. 65%. 0%. 55%. 25日. 16. 69%. O%. 31%. 0%. 100%. 38∼41日. 33. 27%. 15%. 58%. 0%. 36%. 45日. 33. 12%. 0%. 88%. 0%. 21%. 38∼40日. 47. 68%. 2%. 30%. 2%. 72%. さし木後に塊茎を形成したもの、腋芽の伸長したもの、塊茎形成も腋芽伸長もしなかったもののさし穂数に対 する割合、発根したもののさし丁数に対する割合、葉柄の肥大したもののさし穂数に対する割合を%で示した。 出芽後経過El数が増加するとともに、塊茎を形成する割合は減少したが腋芽の伸長する割合は低いままであり、 塊茎形成も腋芽伸長もしない割合は、出芽後経過日数に関わりなく高かった。さし木後の経過日数が増加すると、 塊茎を形成する割合が高くなった。発根したさし穂はほとんどみられず、葉柄の肥大したさし穂の割合が高かっ た。.
(35) 表 3. 塊茎の形成とさし穂総重量の増減 塊茎を形成した. @さし穂数. さし穂総重量の鋤ロ. さし穂総重量の減少. 春作. 170. 49%. 51%. 秋作. 91. 89%. 11%. さし木後に塊茎を形成し、さし穂総重量が増加したものと減少したもののさ し亭亭に対する割合を%で示した。 春作の実験では、塊茎を形成したさし穂の半数に総重量の減少がみられたが、 秋作の実験では、総重量の減少する割合は低かった、。. 表 4. 葉柄の肥大とさし穂総重量の増減 計. さし穂総重量の増加. さし’穂総重量の減少. 47%. 1%. 48%. 葉柄肥大なし. 40%. 12%. 鋼52%. 87%. 13%. 計. 葉柄肥大あり. さし木後に塊茎形成も腋芽伸長もしなかったさし’穂について、さし穂総重量 が増加したものと減少したもの、葉柄の肥大のあるものとないもののさし穂数. (102)に対する割合を%で示した。』 塊茎形成や腋芽伸長はしないが、さし穂総重量が増加しているものが高率で あり、これについて葉柄の肥大の有無をみるとほぼ同率であった。.
(36) 表 5. 葉面積と葉軸を除いた葉身重との関係 葉面積と葉身重※との. @ 相関係数. 品種. 葉身1グラム当たりの @ 面積(c㎡). だんしゃく. r=0.978. 39. 35±0. 571. メークイン. r=0.990. 38. 92±0. 936. せとゆたか. r=0.995. 40. 21±:1. 810. ※葉身重は葉身かち葉軸を除いたものの重さ 葉面積と葉軸を除いた葉身重との相関係数と葉身1グラム当たりの面積 を示した。. 葉面積と葉身重は、3品種ともに非常に高い相関がみられ、葉軸を除い た葉身1グラムあたりの面積は3品種および品種内の各部分ともにほぼ同 じ値を示した。. 表 6. 遮光ネットの組み合わせ枚数と光量. 遮光ネットの. 白色. 0. 1. 3. 2. gみ合わせ枚数. 黒色. 0. 1. 2. 3. 100. 34. 10. 4. 光量. 光量の調節は、自色と黒色の遮光ネットの組み合わせ枚数を変えて行なった。光量は、 晴天と曇天の日の正午に各プランターの直上の照度をルクスメーターでそれぞれ10ヶ 所測り、その平均値を求め、ネットで覆わないものを100として求めた。.
(37) 表 7. 光量. 三光三三における葉面積と塊茎重とめ相関係数と回帰直線. 葉面積と塊茎重との. 葉面旧くx)と塊茎重(y)との. @ 相関係数. @ 回帰直線. 100. r=O.862. y・=・0. 009x十〇. 007. 34. r=0。886. y=・0. 0 12x−0. 299. 10. r=0、899. y・=・0. 01 0x−0. 21 5. 4. r’=0.927. y=0. 006x−0. 055. 塊茎を形成し、総重量の増加したさし穂について葉面積と塊茎重との関係を 相関係数と回帰直線で示した。 各光量区において葉面積と塊茎重とは高い相関がみちれた。.
(38) 表 8. 塊茎を形成したさし穂の総重量の増減の割合. 日照時間. 塊茎を形成した. 総重量が増加した. 総重量が減少した. @さし穂数. @さし穂の割合. @さし穂の割合. 14時間. 17. 59%. 41%. 8時間. 13. 46%、. 54%. 0時間(全暗). 17. 0%. 100%. 塊茎を形成したさし穂について、さし木後に総重量の増加したものと減少したものの さし月数に対する割合を%で示した。 日照時間が短くなる程さし木後に枯死し、総重量が減少したさし穂の割合が増加する 傾向がみられた。. 表 9 NAA濃度の異なる培地でのカルス形成(培養1ヶ月後) NAA濃度(PP皿). 0. 0.5. 十. 昔. 、 1. 5. 10. 50. 帯. 帯. ±. ±. 0.1. カルス形成. 帯. ±変化なし. +カルスの直径10mm以内 昔カルスの直径15mm程度 帯カルスの直径20mm程度 塊茎かちコルクボーラーで打ち抜いて得た組織片をNAAを含む培地に移植し、培養1ヶ月後のカル スの形成状態を示した。. NAAの濃度0.1かち10ppmの範囲でカルスの形成がみられ、1かち10ppmの濃度で形成 がよかった。.
(39) F. 一. ぐ 駈一. 、,㌧. 、 ・ 、. \. 噌謡. ド .爵−. 3プ. ∼た. 2れ. を入 ︶を 芽ト 腋イ ︵ラ 部ユ 基キ のミ. 穂一. しバ さに. 、・つ. しょ とい 本な 基ち をな し重 法ざが。 方芽葉た の葉でし 木はささ し穂深に. さしの一. さmタ cン. 図. 1. 、. 険’. ’. ?@ 一.
(40) e,,,.一一一一一.. 図. た。. 感光定着を行な一. 2 葉軸を除いた葉身の合成紙感光紙への転写 さし穂の葉軸を除いた葉身を合成紙上に展開し. し 節黒 調、. を3. ・え 、. 量 光白 て︵. 変一 を− 数 枚黒 せ、. わ1. 合 。. 節み白た 調組︵し ののらに 量トか一. 光ツ左3. るネ 、. よ光は黒 に遮数、. トの枚2. ッ色の. ネ黒ト白 光とッー 遮色ネ、 白。︶. た2. 図. 3. e. ●▼「膠●●. b。’. Ω● θ.
(41) 名 爆 ノ. し≒. 匪 、 節し 調を. のい. 間覆。 時でた 照箱し 日ル節 る一調 よボを に段間 箱に時 ル一照. 一定時間ごとに覆いを取V. r隔、、. 二. 一タ日. ボンて 段うし プし 外. 図. 4. 千. 7. 図. 5. 腋芽の伸長したさし穂かちの発根 さし木後に伸長した腋芽の基部から多数の発根がみられた.
(42) 笥. A. B. 葉大 肥. の 。も たた れれ一 らら作 みみ秋 がで一 積木の 蓄しも 穂のさた しんのれ さぷ葉ら たんいみ しで若で 大にる木 肥分あし が部にさ 部い位の 基白上期 のた端温 柄し先低. (春作). AB. 図. 6.
(43) 12. 5. 12. 0. IL5. 切. A. v 11.0 9. 5. 淵職.蜘鰹潭 ︶. 9. 0. 8. 5. 駒. 8. 0. 5e .5. 5. 0. 4. 5. IL一一Lt一一一一一“一一一一.一.一一“ny.一“一一一一一. O 246810 12. 16. 20 一24 28. 29. さし木後経過日数 ( 日 ). 図 7. さし木期間中のさし穂総重量の増減. さし穂総重量をさし木後2∼4日ごとに測定した。 さし穂総重量が重いものでも軽いものでも同様な増減を示したが、 さし穂総重量が重い程、増減の幅が広い傾向がみられた。さし穂織 重量はさし木後6日までは減少し、それ以後増加しはじめた。.
(44) ユ.5. だんしゃく. 塊藝里. 1. a u 日. a. g e.s. ε晶. z ユ.5. 華. ・ 雲. メークイン. 1. 口. o a. a. g Z.5. 。 o. oo. 0. 0. 一=・ g. 4ヨ5.. せとゆた一. ヘ」. 1. a 、,. o. o. g. e.s o. 0◎. 0 0. a. o. o. o. z e. le an 3e ca sa om 7Z en su 12”b 11Z l n 13Z. 葉面積。㎡. 図 8. 葉面積と塊茎重との関係. さし木後に塊茎を形成し総重量が増加したさし穂で、だんしゃく、 メークイン、せとゆたかの3品種の葉面積と塊茎重との闘係を示し た。. 葉面積と塊茎重とは3品種ともに比例する傾向がみちれ、だんしゃ くとメークインでは、葉面積が2倍になると塊茎重もおよそ2倍に なったが、せとゆたかでは、1.5倍程度であった。.
(45) だん しゃ く. 1.5. 垂. 1. o o. 口. g. Z.5. oo a$. 1.. 05. メーーク イ ン. 垂. o. 1. 口 i一 O.. a. g e.s. o o. 0. oo. 0 〇一5. 1. せとゆたカ、. 垂. 1 o. a. g Z.5. a a. 口. .o. a ao o . a. a. 1 2e. 図 9. 4g 6e so 葉面積。㎡. leo. 120. 葉軸を除いた葉身重から求めた葉面積と塊茎重との関係(春作). さし木後に塊茎を形成し総重量が増加したさし穂で、葉軸を除い. 、だんしゃく、メー・一 た葉身1グラムあたりの面積を40c㎡として. クイン、せとゆたかの3品種の葉身重から求めた葉面積と塊茎重と の関係を示した。 、 葉身重かち求めた葉面積と塊茎重とは、3品種ともに比例する傾 向がみちれた。.
(46) 出芽後40一一45日メークイン 1.5. D. 塊茎重. 1 o. g. 口 口 0. Z.5. ロロ. ロ。. D. 口. o ロロ. 口. n. 口. z 4e. 20. 60 80 100. 16e. 120 140. 葉面積。㎡ 出芽後60日メークイン 1.5. 塊 蚤里. 1. 口. 口. g. ・Z.5. o 口. ロロ、. 0. 0. 40. 60. 80. 1. 1. 20. 14. を. Cm. oo. む. 2e. 積 面 葉. z. 璽 10. 口. D. 出芽後経過日数の異なるものでの葉面積と塊茎重との関係. さし木後に塊茎を形成し総重量が増加したさし穂で、出芽後経過. 日数が40∼45日のものと60日のものの葉身重から求めた葉面. 積と塊茎重との関係を示した。 葉面積と塊茎重の比例傾向は、出芽後経過日数の異なるものでも みられた。. 1. α.
(47) Z.4. 塊華里 (乾). Z.3 o. 口 か’. Z.2 口 口. g. a. o o. 口. 口. a. 口. Z.1. a b. z. z. Z.1 Z.2 Z.3 Z.4 Z.5 Z.6 Z.7 Z.8 ,葉身重(乾)g. 図 11 乾燥重量を求めたものでの葉身重と塊茎重との関係. 1 さし木後に塊茎を形成し総重量が増加したさし穂で、葉軸を除い た葉身と塊茎を乾熱滅菌器で乾燥させ、冷却後直視天秤で計量し、 乾燥重量での葉身重と塊茎重との関係を示した。 乾燥重量を求めたものでも、葉身重と塊茎重とは比例する傾向が みちれた。.
(48) ・1.ら. 塊茎重. 口. 04枚目. 1 口. ●5枚目. g. ロ6枚目. eim. Z.5. ■7枚目. 哉 。 o. z 20. 凶 4e. △8枚目. N e. 60 so loe. 120 140. 160. 葉面積cm2. 図 12. 葉の位置の違いによる葉面積と塊茎重との関係. 頂芽を0として葉を数え、4枚目から8枚目の位置の異なる葉で、 さし木後に塊茎を形成し総重量が増加したさし穂の葉面積と塊茎重 との関係を示した。. 位置の異なる葉でも葉身重かち求めた葉面積と塊茎重とは比例傾 向を示し、葉の位置による違いはみられなかった。.
(49) e.s. 書. 臼.6 口. a. Z.4 口. g 口. 口 . ロ. ロ. 臼.2. ロロ. ・口. 30. O. 口. 8. z. 0. ロO. ロ ロ. O口. 口. 601, 90 120. 口口. 口 150. 180. 葉面積cm2. 図 13. 葉軸を除いた葉身重から求めた葉面積と塊茎重との関係(秋作). さし木後に塊茎を形成し総重量が増加したさし穂で、葉軸を除い. た葉身1グラムあたりの面積を31c㎡として、葉身重から求めた. “葉面積と塊茎重との関係を示した。. 葉面積と塊茎重とは、比例する傾向はみちれなかった。.
(50) 図 14. 光量の違いによる葉面積と塊茎重との関係. さし木後に塊茎を形成し総重量が増加したさし穂で、100 から0の光量の異なる区での葉面積と塊茎重との関係を示した。 光量の異なるものにおいても 、葉面積と塊茎重とは比例する 傾向がみちれた。.
(51) id. 雌. 1 .00. 1.5}. 三 董 . 三 9. 1. P回. .口. .、日. .臼.S. 日占b 幽 ..’. . .撃. 一.. 02﹁.. ち4.. 光量1.. 塊芯董. 1・ .5. ロ。. 1. 9 oO @D V“’ ’. ・Z.5. o”. 例︸2. ロ ロ. 光it1. ’1.0. 塊華里. ・p.. ・.1.;.5.F. o 1. .9. .口. o 0 O. z. ・2. 0ロ. Z.5. 1’. :う罎..4 ,. 塊 三里. s. .1し5. 1. 0. 9. 口 口. 巳L5.. 口. 口. 。. 口 口. 日 z・. .2Z. 4e. ca. 8tZ) IZ[]. ’121.・. 葉面積。㎡. 図14.. .光量の違いによる葉面積と塊茎重との関係. 1 4’. ’16{b. 1 821.
(52) う阻 . うヒ量 . 1.5. ■00. ■0. り ■. 肇璽. 光ヒ匠 4. g O.5. o. 20 40. 60 ・ 80・ 100 .120 140 160 − 180. 葉面積 図 15. 光量の違いによる葉面積と’. ’c nf. 茎重との関係(回帰直線). 各光量区における葉面積と塊;茎重との関係を回帰直線で示した。. 光量100から10までは、葉面積に対する塊茎重はほとんど差 がなかったが、光量が4まで下がると差がみられるようになった。.
(53) 1.2. 塊華璽. 1. ●3時間 ロ6時間 轟9時間. e. Ze8. ,Z.6 :g. 璽. 一口. e.4. 臼.2. 置. 掴−’. .1. 50. 10Q. 富. 1口’『■. z. 150 葉面積. 図 16. 口. .200. 250. dtV. 日照時間の違いによる葉面積と塊茎重との関係. さし木後に塊茎を形成し総重量が増加したさし穂で、3から9時 間の日照時間の異なるものでの葉面積と塊茎重との関係を示した。. 形成された塊茎は重量が0.01から0.04gの小型のものが 多く、日照時間の増加により単位葉面積あたりの塊茎重が増加する. 傾向はみちれなかった。. 300.
(54) L:.: :. :a. F−i.i.. ・!4やセ. .rt一一 e一. ’ trtt ↓.亨 ’tr. t. φ 卓t.,.. Et一. レ。 ・. 二←r. .哲・. 一一Lr” 一’一’r. 一ウセ讐 . 一一. −一. t’ ”. 溢. 一→. pt.’1”. }1:1三ヨ. 幽. 織. 旧. . 乱 置雫 拝 ・ i .セ ごこ コ. +..e−i一’. ...噛し↓.. “’t’十’. “V4・. ヰコ , ﹁ 誹. .みΨ。. ’ 算 .季. をヰトヤヰるすをヰ. +一1 ’ t. +:一一・ ;一. ・“. . ﹂ ﹁舜. 養麟. 一.一L... 7tr .一 t.卜十.. 誓+.. .了 ↓噌.一幽↑ . ’..←つ t. 曽L. 4.嚢コ、蔦慧. コ. .一+. †+十.峯. 十’t’. ←↓. .噌レ,. 4. 葺. 環. :t. →転’噌 1やし.. ゆ一. . ↑ . 9﹂﹁. #肩一け.. 箕. ト;・. .. o ﹁ ・. 榊. 一一+. ←移軍. rト 審 ・. 一一一. 拝. ﹁ . ←︸ #畔 . 藩憶澤糊..、 コ 曹. 4十十一. .1軍. 羅 灘 事 醸 ÷. 一ti. 麺認㍑滋. ⋮. 耐. 「. 墾蹴導一∴滋. T. ,. ¥. 「 ’. 齢. 」?r‘一唱.過φ ・. き. s , . 一. 一 ↓,幽. 一曽マ←.. 昏一?.曹. 嘱 ’7 1. L†う一. 酔 . u . t’一“i’. 9t.’ ェ’噂」f「辱・. t”P.セヒ.’ ’t. ☆÷∫脳÷÷モ. ,. ヤ. ’ 一’t’:’. ド. 1』’幽↑”. ∵享「’ 一 一 一一一’. ト テ ポ ド. 一. シ. た し 手 入 り よ 子社一 種会種 の式生 モ田田 イ豊中 が種ー. ヤイト. 7. ジキイ タワ ホ. 図 1. D一..一一H一一F.. ホワイトアン1.
(55) x N. ①ハイター10%液で30分. 滅菌し、クリーーンベンチ内で. 滅苗蒸留水で2回水洗する。. ② 滅菌シャーレ上に置き.、殺 菌したメスで淳く皮をむ’く。. ③ハイター5%液で10分滅 菌し、滅菌蒸留水で2回水洗 する。. o. 9 9 9 9. ④滅菌した直径5mmのコル. クポーラーで両方向に打ち抜 く。. 図 ユ8. ⑤打ち抜いた組織の両端を. 10mm程切り落とし、長さ 2mmに切断する。. 塊茎組織の切り出し方法. ⑥ 円盤上分組織を培地に移植 する。.
(56) //一. ①クリーンベンチ内で無菌植物. ②殺菌したメスで節部を含む茎. を滅菌シャーレ上に取り出す。. の切片を切り出す。. 図 19. ③茎の切片を培地に移植する。④節部からシュートが伸長する。. 種子からの無菌三物あ増殖方法.
(57) 7轍.. 警’一”. ’. 拳,、. ・=轟薩. e)(’i.1”. Dtt/. 壁. sM−ee 図 20. A. 無菌植物の増殖. A 培地に移植した茎の切片 B 節部から伸長したシュート(移植後10日めのもの).
(58) 図 21 塊茎から得た組織片のカルス形成 塊茎から打ち抜いて得た組織片は、オーキシンを含む培地で カルス化したv A 暗所で培養したカルス B 明所で培養し、緑色になったカルス.
(59) しご:「. θ・. @ 覧嚢. @ ・幣噸. ( \. 聡・醍一蟹. IN−puV 叛『貫\. 。繍購.蟄. ¥触 講”. 載璽、. 勺 e. ヒ し. ヘシ. ..、 ♂桝 デー一 些. ノ∼馴. 向. A. 図22 ジャガイモの子葉と本葉 ジャガイモの種子は、培地に置床後7∼10日で子葉を展開した。. A 子葉 B 本葉. B.
(60) 鞭蝋 図 23 上方に伸長し、腋芽となったストロン 子葉の基部からストロンの伸長がみられたが塊茎を形 成せず、腋芽として伸長し葉を着けるようになった.. 図 24. 増殖した無菌植物に見られた複葉 節部を含む茎の切片を切り出し、培地に移植を繰り返 すと、複葉の形成がみられた。しかし、露地のものに比 べると小葉の数は少ない。.
(61) 図 25. 茎の切片を暗所で培養したもの 節部を含む茎の切片を暗所で培養すると、白く徒長した シュートがみられた。. 訟. ♪ノ. 9障︶=. 唾. iEs= iS :1. 図 26. 温度の異なる条件での無菌植物の成長. 光条件を一一定にし、温度を10から35℃に変えて培養 した。培養10日後では、茎葉の成長は28℃が最もよく、 35℃では枯死した。.
(62) 図 27 五内培養で形成した塊茎 A 腋芽の肥大したもの B 芽の基部の肥大したもの.
(63) 1. ド. A聯、 4. ?’t’︶’. .. d’ 1’B 図 28 節部からストロンを生じて形成した塊茎. A ストロンの先端が肥大したもの B ストロンの腋芽が肥大したもの.
(64) t ’Se.s一.一rv.’ i・. 酔5. 、麺 噛. 韓. 召. 静 鴨. ,,罪. 図 29 長期間(6N.・10ヶ月)培養後に形成した塊茎 長期間培養を続けると茎葉の枯しゅうが始まり、培養条 件に関わらず塊茎を形成するものがみちれた。.
(65) 驚『㍗. 爆.,… ”Mi.,,g..,一一,.. / 隻声叱. 図 30 形成された塊茎から伸長した芽 形成された塊茎から茎葉を伸ばすものがみられた.. ‘. A.
(66)
図
+6



関連したドキュメント
事務局 そのとおりである。. 委員
この見方とは異なり,飯田隆は,「絵とその絵
この数字は 2021 年末と比較すると約 40%の減少となっています。しかしひと月当たりの攻撃 件数を見てみると、 2022 年 1 月は 149 件であったのが 2022 年 3
それから 3
経済学研究科は、経済学の高等教育機関として研究者を
夫婦間のこれらの関係の破綻状態とに比例したかたちで分担額
かつての広葉樹は薪炭林としての活用が主で、20〜40年の周期
これらの事例は、照会に係る事実関係を前提とした一般的
