doi : 10.3136/nskkk.63.464 http://www.jsfst.or.jp
研究ノート
エダマメ栽培温度がマルトース生成量に及ぼす影響
†野村知未
1,松井元子
2,大谷貴美子
2,村元由佳利
3,古谷規行
4* 1元京都府立大学大学院:(現)広島女学院大学人間生活学部管理栄養学科 2京都府立大学大学院生命環境科学研究科応用生命科学専攻 3京都府立大学生命環境学部食保健学科 4元京都府農林水産技術センター生物資源研究センター:(現)京都府山城北農業改良普及センターEffect of Growing Temperature on Maltose Production of Green Soybean
Satomi Nomura1, Motoko Matsui2, Kimiko Ohtani2, Yukari Muramoto3and Noriyuki Furutani4*
1Former: Graduate School of Life and Environmental Sciences, Kyoto Prefectural University, Present: Faculty of Human Life Studies, Hiroshima Jogakuin University,
4-13-1 Ushita Higashi, Higashi-ku, Hiroshima, Hiroshima 732-0063 2Graduate School of and Environmental Sciences, Kyoto Prefectural University,
1-5 Hangi-cho, Shimogamo, Sakyo-ku, Kyoto, Kyoto 606-8522 3Faculty of Life and Environmental Sciences, Kyoto Prefectural University,
1-5 Hangi-cho, Shimogamo, Sakyo-ku, Kyoto, Kyoto 606-8522
4Former: Biotechnology Research Department, Kyoto Prefectural Agriculture, Forestry and Fisheries Technology Center, Present: Kyoto Prefecture Institute, Agricultural Yamashiro Extension Center,
1 Tanabemyota, Kyotanabe, Kyoto 610-0331
The present study examined the effect of three growth temperatures on the starch gelatinization temperature and β-amylase activity of two cultivars of green soybean in experimental plots. The β-amylase activity of Fuki at climate chamber of 25℃ was significantly higher (p<0.01) than that grown in a field. On the other hand, we found no significant difference between three plots of Shin-tamba-guro upon comparison of their β -amylase activities. However, when comparing the starch gelatinization temperature, the values for the three plots were significantly different (p<0.01): gelatinization temperature increased with a rise in growth temperature. Thus, we suggest that the production of maltose in green soybean is affected by temperature during the ripening process.
(Received Mar. 22, 2016 ; Accepted Jun. 22, 2016)
Keywords : green soybean, maltose, β-amylase, growth temperature, starch gelatinization temperature キーワード : エダマメ,マルトース,栽培温度,β-アミラーゼ活性,糊化温度 エダマメは,日本や中国・台湾で主に食されており,国 内では夏から秋にかけての代表的な野菜として地位を築き 上げている.2000 年代に入るとエダマメは「世界の食べ 物」1)と位置付けられ,さらなる良食味品種の育成が国内 各地で行われている.良食味のエダマメの品種育成では甘 味・旨味を呈する遊離糖量や遊離アミノ酸含量が重要視さ れ,それらの成分はエダマメの食味に大きく影響する.エ ダマメには,アラニンなどの甘味を呈す遊離アミノ酸を含 むことが報告されているが,アミノ酸含量に比べて,糖量 はゆで加熱後で約 2 g∼6 g/100g-f.w.2)∼4)と非常に多くエダ マメの食味に大きく関与する.その中でも含有量が最も多 いスクロースは,エダマメの食味向上にとって欠かせない 要因である1).ゆで加熱後の糖の約 60 % を占めるスクロー ス5)は,収穫後,エダマメの呼吸に利用されるため常温で は急激に低下する6).そのためスクロースの減少を防ぐた めに流通時の徹底した温度管理や,青果物自身の呼吸を低 く抑え品質低下のスピードを遅延させる P-プラス(住友 ベークライト株式会社)などの鮮度保持フィルム7)8)の導入 が不可欠である.一方,マルトースはエダマメが持つ β-アミラーゼによって加熱中に糊化デンプンに作用して生成 され2)9),流通時の保存温度にあまり影響されない10).そのた め,調理後の糖の増強,補強が期待されるものの,品種2)や 作期4)によって生成量が異なることが報告されている. 京都のブランド野菜の丹波黒大豆エダマメ「紫ずきん®」 1〒732-0063 広島県広島市東区牛田東 4-13-1,2〒606-0823 京都府京都市左京区下鴨半木町 1-5 3〒606-0823 京都府京都市左京区下鴨半木町 1-5,4〒610-0331 京都府京田辺市京田辺明田 1 *連絡先(Corresponding author),[email protected]
は,加熱中にマルトースを生成することが報告2)されてお り,早生品種から順に 紫ずきん 2 号,紫ずきん,新丹波 黒 で構成される.紫ずきん 2 号 は,他の 2 品種と異なり マルトース生成量の年次変動が著しく,古谷等3)は,紫ず きん 2 号 の子実肥大期が 8 月の高温期となるためデンプ ンの糊化温度や β-アミラーゼ活性の変動によるものであ ると考察している.我々はこれまでに,エダマメを加熱中 に生成されるマルトースの量は,デンプン量と有意な相関 はなく,品種による β-アミラーゼ活性の強さとデンプン の糊化温度が関与していること,また,新丹波黒 のマル トース生成能は高く,一方でマルトースの生成を検出でき なかった品種があることを報告2)した. そこで本研究では,先行研究2)にてマルトースは生成さ れずスクロース含量が高かった高スクロース型9)の 富貴 と,マルトース生成量が高かったデンプン型9)の 新丹波黒 を試料として栽培温度がエダマメのマルトース生成量に及 ぼす影響について検討した. 1. 実験方法 (1)材 料 2012 年に京都府農林水産技術センター・生物資源研究セ ンターのほ場で栽培された丹波黒大豆エダマメの 新丹波 黒 および黄大豆エダマメの 富貴 の 2 品種を用いた.こ のほ場区に加えて,栽培温度の違いによるマルトース生成 量の変動を把握するため,さらに,ほ場より高温のビニル ハウス区11)12)(換気扇設定温度 35℃)と,25℃に制御したガ ラス人工気象室区(以下,25℃区)の 3 区で栽培し,適期に 収穫したエダマメを試料とした. 各々のエダマメは,収穫適期に達したところで速やかに 収穫・脱莢し 新丹波黒 は沸騰後 10 分,富貴 は 6 分ゆで た後,直ちに実験に供するまで−30℃で冷凍保存した. (2)遊離糖の分析 遊離糖の分析は,前報2)と同様の方法で行った.すなわ ち,上記の凍結保存サンプルを 8 倍量の 80 % エタノール 中で粉砕抽出し,遠心分離後の沈殿物を再度 8 倍量の 80 % エタノール中で撹拌抽出し,遠心分離した上清をメンブラ ンフィルター(0.45 µm)で濾過し,抽出は 3 反復行った. 得られた抽出液は,示差屈折計検出器(RID)を装着した 高速液体クロマトグラフ(1260Infinity/Agilent technol-ogies,カラム:Asahipak NH2 P-50 4E/Shodex,カラム温 度 40℃)を用いて分析した.移動相は,70 % アセトニトリ ル(流速 1.0 ml/min)を用いて分離した.標品としてフル クトース,グルコース,スクロースおよびマルトース(和 光特級/Wako)を使用し,各遊離糖の含量に関する検量線 を作成して 4 種の糖の定量に用いた. (3)耐熱 β-アミラーゼ活性の測定 β-アミラーゼの活性は,前報2)と同様に,Betamyl-3Ⓡ キット(Megazyme/Bray)を用いて測定を行った.エダ マメ子実に 1 M Tris/HCl 緩衝液(pH 8.0, 20 mM EDTA, 0.02 % アジ化ナトリウム含)を加えホモゲナイズし室温で 1 時間撹拌抽出した.その後懸濁液を遠心分離(15 000 rpm,常温,10min)し,得られた上清を粗酵素液とした. 基質の p-ニトロフェニルマルトトリオースに抽出した粗 酵素液を加え 60,65,70,75℃の各温度で 10 分間反応させ た.その後 1 %(w/v)Trizma-base を加えて反応を停止し, 解離した p-ニトロフェノールを 400 nm で測定した.結果 は,1 分間に基質から 1 µmol の p-ニトロフェノールが解 離した量を 1unit とし,エダマメ重量当たりの β-アミラー ゼ活性を比較した.なお,抽出した粗酵素液は,分析に用 いるまで−30℃の冷凍庫で保存し,測定は 3 反復行った. (4)デンプンの分画 デンプンの分画は,Matsunaga 等13)の方法で行った.つ まり,エダマメを 10 倍量の 0.05 M の水酸化ナトリウム中 で粉砕し懸濁させることにより除タンパクした.その後, 沈殿物を蒸留水に懸濁させさらに 1 時間撹拌させた.沈殿 物に再び蒸留水を加え 0.1 M の塩酸で中和した後そのまま 撹拌を 1 時間行い遠心分離(5 500×g,10min)した.その 後,沈殿物を蒸留水に懸濁させ撹拌し洗浄して 355 メッ シュ(東京スクリーン株式会社)に通し,30℃のオーブン (DRYING OVEN Mov-112/SANYO)で 24 時間乾燥させ た.なお,Matsunaga 等の方法は,中和により生成された 塩を除去するために十分な洗浄が行われるため,デンプン 糊化温度へのナトリウムの影響はなかった14)といえる. (5)示差走査熱量測定(DSC) 無水単離デンプン 4 mg および蒸留水 20 µl をサンプル パンに封入し,示差走査熱量計(DSC-60/SHIMADZU)を 用いて分析した.昇温速度は 2℃/min とし,対照として水 20 µl をサンプルパンに注入したものを用いた.DSC の特 性値として糊化の開始温度(To),ピーク温度(Tp),終了 温度(Tc)を求めた. (6)統計処理 新丹波黒 および 富貴 各々の品種について,栽培温度 が異なる 3 区のエダマメの遊離糖量,デンプン量,糊化温 度,β-アミラーゼ活性(unit)の値についてはエクセル統 計 Statcel3 アドインソフト(オーエムエス版)を用い,一 元配置分散分析後,Scheffe の F 検定を行った.また,各 栽培条件下における 2 品種の差異は,student の t 検定を 行った. 2. 実験結果 (1)エダマメの子実肥大期と温度 ほ場で栽培された各エダマメの開花日および収穫日,子 実肥大期の最高気温および最低気温の平均値を Table 1 に 示した.エダマメ子実は,受精後,莢の伸長が始まり一定 の長さとなった後に充実し,子実が肥大し始めるのは 富 貴 のような早生エダマメは開花日から数えて約 20 日 後15)より,新丹波黒 のような晩生エダマメは開花より約 40 日後16)である.また,デンプン蓄積が顕著となる時期が
子実肥大開始時期と一致する1)17).これらのことから,本研 究では,夏エダマメの 富貴 は開花日より数えて 20 日後 から収穫日までの 15 日間を,新丹波黒 は開花日より 40 日後から収穫日までの 25 日間を子実肥大期と定めた. 子実肥大期のほ場の最高および最低気温を比較したとこ ろ,両者とも最高および最低気温の差は約 10℃であった が,新丹波黒 の子実肥大期の最高および最低平均気温は 富貴 に比べて約 5℃低い結果となった.高温のビニルハ ウス区は,ほ場区よりも最高気温の平均値で 新丹波黒 は 4℃,富貴 は 5℃高くなったが,最低気温の平均値では両 品種とも 2℃しか高くならなかった.25℃区は,ほ場区よ り最高気温は 新丹波黒 で 4℃,富貴 で 9℃低くなった. 一方,25℃区の最低気温は,ほ場区よりも 新丹波黒 で 9℃,富貴 で 2℃高くなった. (2)遊離糖量 生およびゆでエダマメの遊離糖量の結果を Fig. 1 に示し た. 両品種ともに,栽培条件が異なっても生エダマメにおい てスクロースが遊離糖量の約 90 % を占めていた.全遊離 糖量は,新丹波黒 では,ビニルハウス区(2.47 g/100g-f.w.) >ほ場区(1.69 g)>25℃区(1.59 g)であり,富貴 におい てもビニルハウス区(2.40 g/100g-f.w.)>25℃区(2.39 g)> ほ場区(2.27 g)の順となった.ゆでエダマメでは,富貴 のビニルハウス区以外のエダマメでマルトースが生成さ れ,新丹波黒 は,ほ場区>ビニルハウス区>25℃区の順 となりほ場区とビニルハウス区では有意な差は認められな かった.一方,富貴 は 25℃区>ほ場区>ビニルハウス区 の順となり 3 つの栽培条件で有意に(p<0.01)マルトース の生成量が異なった. (3)β-アミラーゼ活性 栽培温度の異なる 2 品種のエダマメの β-アミラーゼの 温度依存および活性の強さを比較し,Fig. 2 に示した. 前報と同様に,各々のエダマメから抽出した粗酵素液と 基質の p-ニトロフェニルマルトトリオースを 60,65,70, 75℃で反応させた場合 65℃で最大値を示し後,70℃から 減少し始め 75℃では急激に減少した.この傾向は,栽培条 件を変えても変化は認められず,ほ場区,ビニルハウス区 および 25℃区において同様だった. 新丹波黒 の場合,いずれの栽培条件においても酵素反 応の各温度で活性の強さに有意な差は認められず,ほぼ同 等の値を示した.しかし,富貴 の場合では,3 つの栽培条 件において β-アミラーゼ活性の強さは有意に異なってお り,最大活性を示す 65℃に着目すると,25℃区(416U)> ほ場区(309U)>ビニルハウス区(251U)の順で活性は異 なった.また,富貴 の β-アミラーゼ活性を 65℃の最大 値(=a)を 70℃の値(=b)で割ったものを低下率(=b/a ×100)とした場合,25℃区(80 %)>ほ場区(78 %)>ビニ ルハウス区(71 %)となり,富貴 は栽培条件によって β-アミラーゼ活性の低下率が違っていた. (4)デンプンの糊化温度 エダマメの単離デンプンの糊化特性を DSC で調査した (Table 2).品種および栽培条件に関わらず単一ピークを 持つ曲線が得られた.糊化開始温度,ピーク温度,終了温 度供に 2 品種で差が認められ,通常栽培条件のほ場区にお いて,新丹波黒 の糊化ピーク温度は 富貴 に比べて有意 に(p<0.01)低く,約 12℃異なった. さらに,2 品種ともに Table 1 に示したように,子実肥 大期の気温が異なることで同品種であっても,糊化温度に 差が認められた.糊化ピーク温度(Tp)は 新丹波黒 では 25℃区(70.2℃)>ビニルハウス区(60.2℃)>ほ場区(58.9 ℃)となり 25℃一定で栽培した際に最も高かったが,一方, 富貴 ではビニルハウス区(72.8℃)>ほ場区(71.1℃)>25 ℃区(68.6℃)となり 25℃一定条件で栽培したエダマメデ ンプンの糊化ピーク温度が最も低い結果であった. 3. 考 察 本研究で用いたエダマメ品種の夏エダマメの 富貴 は, スクロース含量が高い2)ものの,通常の栽培条件ではマル トースはほとんど生成されないことが明らかとなってい る.本実験において通常の栽培条件(ほ場区)では,0.30 g/100g-f.w.とわずかに生成が認められたが,ビニルハウス 区では検出されず,逆に 25℃区では 1.19 g/100g-f.w.生成さ れた.加熱によるマルトース生成には,β-アミラーゼが失
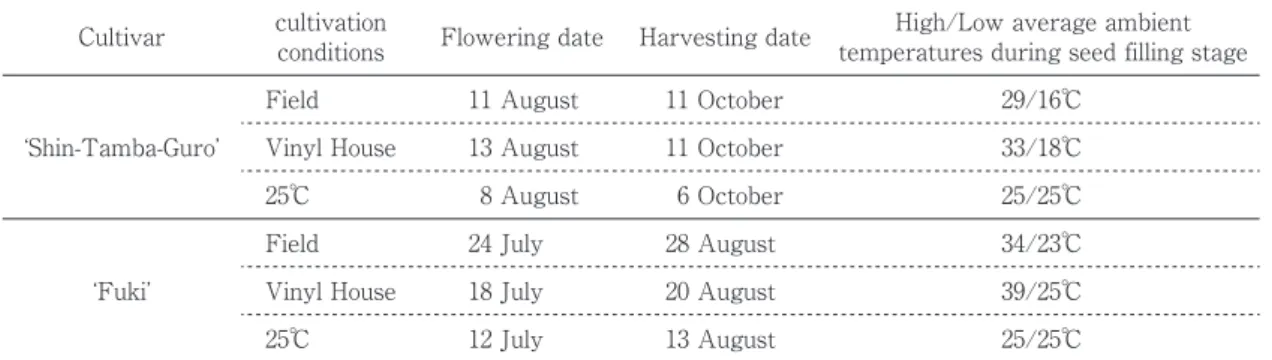
Table 1 Green soybean samples used in this study
Cultivar cultivationconditions Flowering date Harvesting date temperatures during seed filling stageHigh/Low average ambient Field 11 August 11 October 29/16℃
Shin-Tamba-Guro Vinyl House 13 August 11 October 33/18℃ 25℃ 8 August 6 October 25/25℃ Field 24 July 28 August 34/23℃ Fuki Vinyl House 18 July 20 August 39/25℃ 25℃ 12 July 13 August 25/25℃
活する前にデンプンの糊化が開始されていることが条件と なり,糊化温度の低い方がマルトース生成に当然有利とな る.これまでに,米では登熟期ごろ18),サツマイモにおいて は,塊根の肥大期温度19)がアミロペクチン短鎖/長鎖比お よびデンプンの糊化温度に強い相関を示し,側鎖が長くな ると糊化温度が高くなることが報告されている.このよう に,多数の植物においてデンプンの糊化温度が栽培時の気 温に影響しており,エダマメにおいても,デンプンが急激 に蓄積される子実肥大期の温度がデンプンの糊化温度に作 用する可能性が考えられる.本研究において 富貴 をほ 場区より気温が低い 25℃区で栽培した結果,デンプンの糊 化温度が他の試験区よりも有意に(p<0.01)低くなり,マ ルトースが生成されたと示唆された.また,富貴 をビニ ルハウス区で栽培した場合,Table 1 に示したように,3 区 の中で子実肥大期の温度が最も高くなり,糊化温度が有意 に(p<0.01)高くなったと考えられる.一方,富貴 とは 異なり,適期の栽培でマルトースを生成する 新丹波黒 の マルトース生成量は,25℃区のマルトース生成量が最も低 かった(0.78 g/100g f.w.).新丹波黒 は,子実肥大期の最 低気温が,25℃区よりもほ場区で 9℃,ビニルハウス区で 7℃低いために,25℃区のデンプン糊化温度(To, Tp, Te) は,他の試験区の糊化温度に比べて高くなったと示唆され る. 一方,マルトースを生成させる β-アミラーゼについて も,栽培温度を変えたことにより違いが認められた.犬飼 等20) は,サツマイモ塊根の場合,栽培温度を下げても,β-アミラーゼ活性の強さは変化しないことを報告している. 本研究において,エダマメ重量当たりのマルトース生成量 に及ぼす アミラーゼ活性を比較したところ,富貴 の β-アミラーゼ活性の強さ(最大活性 65℃時)は,ほ場区より も最高気温が低い 25℃区が,有意に(p<0.01)1.4 倍高い 値を示した.新丹波黒 においては,β-アミラーゼ活性の 耐熱性および強さは,3 区において変化が認められず,新 丹波黒 では 富貴 と異なり栽培温度が β-アミラーゼ活性 に与える影響は小さいことが考えられる.これまでに,稲 において β-アミラーゼはデンプンの代謝に関わる21)こと
Fig.1 Free sugar contents of three growth temperatures in experimental plots in two cultivars ( , ),Field;( , ),Vinyl House;( , ),25℃.
が報告されているため,エダマメにおいても β-アミラー ゼ活性に栽培温度が影響する可能性が考えられる. 以上のように,栽培温度がエダマメのデンプンの糊化温 度,エダマメ重量当たりの β-アミラーゼ活性に作用し,マ ルトース生成量に差異が認められた.栽培温度がデンプン の構造に及ぼす影響や β-アミラーゼの内在酵素の種類や 優位性についてはさらなる検討が必要である. 4. 要 約 栽培温度の異なる 3 つの試験区でエダマメ 2 品種を栽培 し,マルトース生成量に関与する β-アミラーゼ活性およ びデンプンの糊化温度の影響を検討した.富貴 は,子実 肥大期の温度が低い 25℃区がほ場区に比べて,β-アミラー ゼ活性の強さが有意に(p<0.01)高くなったが,新丹波黒 の場合,栽培温度の違いにより活性の強さは変化しなかっ た.一方,デンプンの糊化温度は両品種供に 3 つの試験区 で有意に(p<0.01)異なり,子実肥大期の温度が高いほど 大きく上昇した.これらのことから,エダマメ加熱後のマ ルトース生成量は,子実肥大期の温度に大きく影響を受け ることが認められた. 最後に,本研究を進めるにあたり京都府立大学大学院生 命環境科学研究科故本杉日野教授にご協力頂いた.また, DSC 分析には畿央大学健康科学部岩城啓子教授にご協力 頂いた.ここに記して心より感謝申し上げる. 文 献 1) 笹原健夫,作物としての特性,「農業技術大系野菜編 10 マ メ類・イモ類・レンコン」追録第 25 号,(農山漁村文化協 会,東京),pp. 1-14 (2000).
2) Nomura, S., Furutani, N., Ohtani, K., Muramoto, Y., Shibata, M. and Matsui, M., Production of maltose in various cultivars of soybean vegetable. Glycine max(L.)Merr., after Boiling. Food Sci. Technol. Res., 20, 401-407 (2014). 3) 古谷規行,野村知未,大谷貴美子,松井元子,丹波黒大豆エ ダマメにおける食味評価法の開発,園芸学研究,11,309-314 (2011). 4) 本庄 求,篠田光江,武田 悟,田口多喜子,エダマメ新品 種 あきた香り五葉 の作期による内部品質と食味官能の違 い,東北農業研究,60,189-190 (2007). 5) 阿部利徳,氏家隆光,笹原健夫,生およびゆでエダマメの遊 離アミノ酸および糖含量の品種間差異,日本食品科学工業 学会誌,51,172-176 (2004). 6) 廣田智子,田畑広之進,福嶋 昭,井上喜正,中川勝也,丹 波黒大豆冷凍エダマメの品質に及ぼす収穫後の保存状態, 日本食品保蔵科学会誌,29,11-16 (2003). 7) 曽根一純,八分着色イチゴ果実の MA 包装と低温貯蔵を組 み合わせた鮮度保持技術,農業および園芸,86,713-719 Fig.2 Total activities of β-amylase per 1 g fresh weight
(f.w.) of three growth temperatures in experimen-tal plots in two cultivars
,Field; ,Vinyl House; ,25℃.
Superscript letter indicate the level of significance (*p <
0.05,**p<0.01).
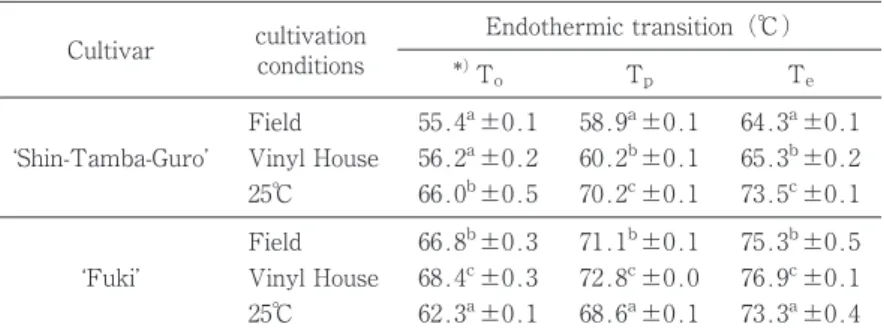
Table 2 Gelatinization properties by DSC of starches from Edamame different cultivars or environment
Cultivar cultivationconditions *)Endothermic transition(℃) To Tp Te
Field 55.4a±0.1 58.9a±0.1 64.3a±0.1
Shin-Tamba-Guro Vinyl House 56.2a±0.2 60.2b±0.1 65.3b±0.2
25℃ 66.0b±0.5 70.2c±0.1 73.5c±0.1
Field 66.8b±0.3 71.1b±0.1 75.3b±0.5
Fuki Vinyl House 68.4c±0.3 72.8c±0.0 76.9c±0.1
25℃ 62.3a±0.1 68.6a±0.1 73.3a±0.4 *)Each value showed mean±S.D.
(2011).
8) 石川 豊,MA 包装の国内動向,日本包装学会誌,17,145-150 (2008).
9) 増田亮一,エダマメの食味向上に関わるマルトース生成反 応の解明,農業および園芸,79,1085-1093 (2004). 10) Sugimoto, M., Goto, H., Otomo, K., Ito, M., Onuma, H., Suzuki,
A., Sugawara, M., Abe, S., Tomita, M. and Soga, T., Meta-bolomic profiles and sensory attributes of edamame under various storage duration and temperature conditions. J. Agric. Food Chem., 58, 8418-8425 (2010).
11) 永畠秀樹,山元皓耕二,温度勾配ビニルハウスを用いた水 稲の高温登熟性の評価,育種学研究,7,95-101 (2005). 12) 藤本幸平,内藤 潔,卜部昇治,ビニールハウスにおける換 気扇による強制換気に関する研究(第 1 報)換気扇の設置 基準の設定および効果について,奈良県農業試験場研究報 告,2,32-38 (1968).
13) Matsunaga, N., Takahashi, S. and Kainuma, K., Rice starch isolation from newly developed rice cultivars by the Improved alkali method. J. Appl. Glycosci., 50, 9-13 (2003). 14) 高橋幸資,白井邦郎,和田敬三,澱粉の糊化と保持塩の関 係,澱粉科学,28,1-8 (1981). 15) 古谷義人,久木井基二,大豆の結実に就いてⅠ,落花の様相 及び莢の初期発育経過,九州農業研究,6,45-46 (1950). 16) 岩本孝幸,桜井敏雄,黒大豆エダマメの安定多収・高品質栽 培技術の確立,京都府農業総合研究所 平成 10 年度 単年 度試験研究成績,23-24 (1999). 17) 増田亮一,エダマメの品質─おいしさに関与する成分,エ ダマメ研究,1,4-9 (2003). 18) 奥田将生,橋爪克己,上用みどり,沼田美子代,後藤奈美, 三上重明,イネ登熟期気温と酒造用原料米のデンプン特性 の年次・産地間変動,日本醸造協会誌,105,97-105 (2010). 19) Noda, T., Kobayashi, T. and Suda, I., Effect of soil tempera-ture on starch properties of sweet potatoes. Carbohyd. Polym., 44, 239-246 (2001). 20) 犬飼義明,松林隆宗,芝山秀次郎,カンショの甘味に関わる 栽培環境要因,根の研究,16,147-154 (2007). 21) 平野達也,太田千尋,平野美奈子,道山弘康,イネ茎葉部の デンプン代謝における β アミラーゼ遺伝子. OsBAM2 の役 割,第 235 回日本作物学会講演会要旨集,pp. 428-429,神 奈川 (2013). (平成 28 年 3 月 22 日受付,平成 28 年 6 月 22 日受理)