日本光合成研究会 会報
第 43 号 2005 年 8 月
O 2
CO 2
日 本 光 合 成 研 究 会
NEWS LETTER No. 43 August 2005
THE JAPANESE ASSOCIATION FOR PHOTOSYNTHESIS RESEARCH
******************************************************************************************
“日本光合成研究会ワークショップ5”のお知らせ
草が作る叢(くさむら)の光環境: 生産構造図の作成 ···
1
トピックス ラン藻のチトクロム
b
6f
複合体と光合成細菌のチトクロムbc
1複合体 -最近の構造解析結果から- 栗栖源嗣··· 3
解説
FACE(開放系大気CO
2増加)を用いた冷温帯落葉樹への高CO2付加実験 小池孝良, 江口則和, 笹賀一郎··· 8
報告記事 第5回日本光合成研究会シンポジウム報告 高橋裕一郎 ··· 13
第5回日本光合成研究会シンポジウム 「光合成研究入門:地球の未来を語ろう!」に参加して 小澤真一郎 ··· 13
シアノ学会体験記
IVth ESF Research Conference on Molecular Bioenergetics of Cyanobacteria
-EuroConference on Genomics, Proteomics and Structure for Functional Understanding
-参加報告 丸山真一朗 ··· 15Gordon Research Conferences on Photosynthesis
に参加して 矢部俊樹 ··· 18集会案内
··· 21
新刊図書
··· 23
事務局からのお知らせ
··· 24
日本光合成研究会会員入会申込書
··· 25
日本光合成研究会会則
··· 27
幹事会名簿
··· 29
日本光合成研究会 会員名簿
··· 30
賛助法人会員広告「日本光合成研究会ワークショップ5」を9月2日(金)・3日(土)に行います。
単子葉植物と双子葉植物の2種類の叢(くさむら)を育て用意しました。「植物の葉のつけ方と 叢の中の光環境」の関係を学びます。野外用の装置で光合成速度やクロロフィル蛍光なども測定 し、ものすごい勢いで光合成をしているこの時期の維管束植物の光環境への応答を検討します。
皆さんが知っている「光合成」が、野外ではどの様に行われているのか、ぜひ一度実際に体験 してください。実習費用は無料で、平塚市の施設(びわ青年の家)を利用されれば、宿泊費も無 料です(寝具洗濯代三百円程度は実費)。予備知識も要求しません。ぜひお気軽にお申し込み下さ い。多くの方のご参加をお待ちしています!
"草が作る叢(くさむら)の光環境: 生産構造図の作成"
オーガナイザー:神奈川大学理学部生物科学科 鈴木祥弘・井上和仁 主催:日本光合成研究会
共催:神奈川大学総合理学研究所
神奈川大学ハイテク・リサーチ・センター
場所:神奈川大学湘南平塚キャンパス67号館生物実習室/圃場
東海道線平塚駅から、神奈川大学行きバス神奈川大学校舎前 小田急線秦野駅から、神奈川大学行きバス神奈川大学校舎前
日程
9月2日(金)
13:00 神奈川大学平塚キャンパス集合
説明・分担
14:00 圃場へ移動
光測定(PARセンサー、光ファイバー分光光度計)
光合成測定(LI-6400光合成蒸散速度計、ミニPAM)
17:00 解散
(びわ青年自然の家の宿泊者は食後、検討会)
9月3日(土)
9:00 層別刈り取り開始
10:00 生重量・葉面積 測定開始 12:00 昼食
13:00 層別刈り取り再開 16:00 集計
17:30 解散
”雨天中止”(研究会のホームページに前夜8時までに掲載します)
お申し込みは、8月20日までに、以下のフォームで下記まで
(宿泊施設のご利用は難しくなると思いますが、締め切り日が過ぎた場合でも連絡をいただけれ ば対処します。)
[email protected]
鈴木 祥弘神奈川大学理学部生物科学科(67-111B)
〒259-1205 神奈川県平塚市土屋2946
--- 申し込み票 ---
氏名:
所属・学年(役職)
住所:
電話:
宿泊施設利用を: 希望する・希望しない (どちらかを残してください)
その他、希望連絡事項
ラン藻のチトクロムb
6f複合体と光合成細菌のチトクロムbc
1複合体
-最近の構造解析結果から-
東京大学大学院総合文化研究科 栗栖源嗣
TOPICS
光合成関連蛋白質は、筆者の専門とする構造生物 学分野では古くから研究対象とされてきた蛋白質で、
関連する構造解析の話を聞かれた方も多いのではな いだろうか。今回取り上げるチトクロム複合体は、
基本的に光と直接反応せず、主に電子伝達に係わっ ている蛋白質である。日本に研究者が少ないことも あって読者の皆さんには、比較的馴染みが薄い蛋白 質かもしれない。筆者は阪大、米国Purdue大、東大 と光合成電子伝達を中心に構造研究を行ってきたの で、少し古い話も交えながら最近のチトクロム複合 体の構造研究について紹介させていただく。
チトクロムb6f複合体には、b型とc型の二種類のチ トクロムが含まれることから、呼吸鎖や光合成細菌 に存在する類縁のチトクロムbc1複合体と共に、チト クロムbc複合体と総称される。チトクロムbc複合体 には、特に膜貫通領域にアミノ酸配列の相同性が確 認され、酵素としても両複合体は、基質がキノン類
(プラストキノン:b6f 、ユビキノン:bc1)と電子 伝達蛋白質(プラストシアニン:b6f、チトクロムc:
bc1)であるという明瞭な共通点ももつ。少し年代を 溯ってみると、チトクロムbc複合体の構造研究は、
プロテアーゼなどで複合体の可溶性領域を切り出し た、部分的な構造解析から始まっている。1994年に b6f複合体中のチトクロムf1)、1996、97 年にbc1、b6f各 複合体の鉄硫黄蛋白質の結晶構造が報告されている
2,3)。膜貫通領域を含む全体構造としては、光合成反 応 中 心 の 構 造 解 析 で ノ ー ベ ル 化 学 賞 を 受 賞 し た Johann Deisenhofer教授のグループが、1997年にウシ 心筋由来チトクロムbc1複合体の結晶構造を報告した のが最初である4)。その後、ニワトリ、酵母の各ミト コンドリア由来bc1複合体の結晶構造が相次いで報告
されている5,6)。直接光合成に係わるものでは、2003 年にラン藻および緑藻由来チトクロムb6f複合体の結 晶構造が7、8)、そして2004年に光合成細菌R. capsulatus 由来のチトクロムbc1複合体の結晶構造が報告され9)、 光合成分野のチトクロムbc複合体の構造研究も、(光 化学系も同様だが)最近になって一気に加速度を増 しているように思われる。他誌に幾つか総説を書い ているので、多分に重複しているところもあるが、
折角頂いた機会なので、直接構造解析に携わったラ ン藻由来チトクロムb6f複合体の全体構造を中心に、
光合成細菌のチトクロムbc1複合体など、最近の構造 研究も交えながらチトクロム複合体の構造研究につ いて紹介したいと思う。
チトクロムb6f複合体は、葉緑体のチラコイド膜に 存在し、2量体あたり16個のサブユニットを含む分 子量22万の超分子複合体である。光化学系ⅡとⅠを 電気的に繋ぐ役割を担うとともに、膜のストローマ 側からルーメン側へとH+を輸送するポンプの役割も 担っている10)。もう 2 年前の話になるが、筆者の所 属していた米国Purdue大学のCramer研究室で好熱性 ラン藻由来の複合体を 3.0Å分解能で構造解析した7)。 ほぼ同時期にフランスのD. Picot等のグループが緑藻 クラミドモナス由来の複合体を 3.1Å分解能で構造 解析している8)。両グループにより報告されたラン藻 およびクラミドモナス由来b6f複合体の結晶構造は驚 くほど良く似た構造であり、3つの新しい補欠分子族
(ヘム、クロロフィルaおよびβ-カロチン)が両方の 構造中の同じ場所に確認された。中でも 5 配位型の 新奇なヘム(heme x or heme ci)はスペクトル法など 他の物理化学的手法で予想すらされていなかった非 常に目新しいヘム鉄であった。今回紹介するもうひ
とつの複合体である光合成細菌由来チトクロムbc1複 合体は、反応中心との間でユビキノンとチトクロム を介して電子のやり取りを行い、2量体あたり6個の サブユニットを持っている膜蛋白質複合体である。
やはり電子伝達と連動する形でH+を輸送するポンプ の役割を担っている。ニワトリのチトクロムbc1複合 体を構造解析したUCバークレーのE. Berryらにより
2004年に3.8Å分解能の構造が報告されている9)(表
1、図1)。
チトクロムb6f複合体はチラコイド膜のストローマ 側に可溶性のドメインを持たないため、界面活性剤 による可溶化が難しく活性型のダイマーよりも不活 性なモノマーになり易いと言う特性をもっている。
我々が構造解析に用いた好熱性ラン藻由来のb6f複合 体は、界面活性剤で可溶化した後も高い電子伝達活 性を有し、構造解析に用いた良質の結晶は合成脂質 を添加剤として加えた時に析出した11)。X線回折強度 データは米国と日本の大型放射光施設のビームライ ンにおいて収集し、構造解析はPtとPb化合物誘導体 結晶による同型置換法と多波長異常分散法により行 った。3.0Å分解能で立体構造を決定した蛋白質の分 子サイズは膜のルーメン側で約120 x 75Å、膜貫通部
分で約90 x 55 Åであった(図1)。光合成細菌由来
のチトクロムbc1複合体も非常に良く似た分子サイズ をしているが、サブユニット構成は、ラン藻のb6f複 合体よりもウシやニワトリの呼吸鎖にあるbc1複合体 のコアサブユニットのそれに近い。
次に構造の細部を見ていくことにしよう。ラン藻、
光合成細菌の両bc複合体とも、膜のルーメン側にはc 型ヘムをもつチトクロム(チトクロムfとチトクロム c1)の可溶性ドメインと[2Fe-2S]クラスターを持つ鉄 硫黄蛋白質の可溶性ドメインのみが配置していた。
これらの構造では、二つの酸化還元中心の距離が30 Å近く離れている為、そのままでは直接電子を伝達 することは出来ない。ウシやニワトリのチトクロム bc1複合体の場合、鉄硫黄蛋白質の鉄・硫黄クラスタ ーとチトクロムc1の酸化還元中心間の電子移動は鉄 硫黄蛋白質の可溶性ドメインによるシャトル運動に より成り立っていると提唱されている。ラン藻、光 合成細菌の両bc複合体共に、ウシやニワトリのbc1複 合体と同様に、鉄硫黄蛋白質の可溶性ドメインによ るシャトル運動によって酸化還元中心間を電子が移 動すると推測される5,6)。
チラコイド膜を貫通するチトクロムb6f複合体の 膜貫通領域は2量体あたり26本の膜貫通ヘリックス から構成されている。膜貫通領域中に電子密度図か
表1 チトクロムbc複合体のサブユニット構成の比較 チトクロム b6f(チラコイド) チトクロム bc1(光合成細菌)
基質 プラストキノン ユビキノン プラストシアニン チトクロム c
- - -
サブユニット 補欠分子族 サブユニット 補欠分子族- - -
チトクロム f c-型 ヘム チトクロム c1 c-型 ヘム サブユニット 鉄硫黄タンパク 鉄硫黄クラスタ 鉄硫黄タンパク 鉄硫黄クラスタ 構成 チトクロム b6 b-型 ヘム チトクロム b(n-末端) b-型 ヘム*新型ヘム
サブユニット IV
*クロロフィル
a チトクロム b(c-末端) -PetG - -
PetL *β-カロチン - -
PetM - -
PetN - -
*新型ヘム,
クロロフィル a 、 β-カロチンは結晶構造解析により存在が確認された。ら3種の補欠分子族を同定した。サブユニットIVの2 つのヘリックスに挟まれる形でクロロフィルa分子 を確認し、b6f分子の外側に突き出る形でβ-カロチン を同定した。驚いた事にすでに分光学的な研究があ る2つのb型ヘム(heme bpおよびheme bn)以外に分 子のストローマ側の端にもう一つ余計なヘムを発見 した。この初めて同定された新しいヘムは 5配位型 で、配位子がb型ヘム(heme bn)のプロピオネイト基 に結合した水分子であるという非常に新奇なもので あった。さらに興味深いことにc型ヘムと同じように チトクロムb6のCys35 とヘムのビニル基がチオエー テル結合していたのである。ではこのヘムはc型ヘム なのであろうか。c型チトクロムの中には5配位型の もの(光合成細菌の持つチトクロムc’など)も有るが 5 番目の配位子は共通してHisの側鎖である。通常c
型チトクロムは 2つのチオエーテル結合によりヘム と結合し、-CXXCH-という保存配列を持つはずであ る。しかし、チトクロムb6のCys35近傍にはこのよう な保存配列は無く、アミノ酸からヘムへの配位子が 無い。したがって我々はこのヘムを既存のカテゴリ ーに分類できない新しいヘムであると結論付けた。
投稿論文中で”heme x”と名づけることにした7)。なぜ このヘムxは長い間存在が確認されなかったのであ ろうか。答えは 2つあると考えられる。まず第一に
「チトクロムbc1複合体と同じである」という強い固 定概念。第二に「膜からの可溶化、精製の難しさ」
である。Heme xを発見した後に、b6fに関する分光学 の論文を確認したが、その幾つかに 5 配位型ヘムに 見られるHighSpinシグナル(総シグナルの10%程度)
の存在を記述している物があった12)。しかし精製の
図 1. 好 熱 性 ラ ン 藻M.laminusus由 来 チ ト ク ロ ムb6f複 合 体 と 光 合 成 細 菌 R.capsulatus由来チトクロムbc1複合体の結晶構造
難しさを踏まえて、それは不純物由来もしくは部分 分解したb型ヘム由来のシグナルであろうと結論し ている。プロテオミクス研究においても同様である。
実際にはマススペクトルの分子量でヘム 1つ分の分 子量差が観測され13)、電気泳動ゲルをヘムで染色す るとチトクロムb6に対応するバンドが染色されてい た。これらの場合も部分的に分解されたb型ヘムが残 っている物として十分な考察がなされていなかった のである。
一方、光合成細菌のチトクロムbc1複合体には、新 しいヘム鉄もクロロフィルもβカロチンも含まれて いない。新しいヘム鉄に対応する部分はキノン結合 サイトとして空いているし、クロロフィルaが結合し ている部分にはチトクロムbのc末端ヘリックスが存 在している。また、b6f複合体がβカロチンを挟み込 んでいる分子量 4千程度の小さなサブユニットは、
光合成細菌には元々存在していない。つまり完全に、
ウシやニワトリの呼吸鎖にあるbc1複合体の補欠分子 族のコピーなのである。ラン藻のb6f複合体と光合成 細菌のbc1複合体の構造は、外見上非常によく似てい るがその中身は、必要なところだけ似ていて、違う ところは結構違うのである。
それでは、どうしてチトクロムb6f複合体にだけ、
ヘムxおよびクロロフィル、βカロチンが結合してい るのであろうか。ヘムxは膜貫通領域のストローマ側 の端に位置し、チトクロムbc1複合体には存在しない ことから、アンチマイシン感受性のフェレドキシン 依存循環電子伝達に関与している可能性が高いと考 えられる。読者の皆さんも良くご存知の通り、光合 成の研究分野では古くからチラコイド膜上にフェレ ドキシン依存性のプラストキノン還元反応が存在す ることが知られている。植物やラン藻は生育環境に よってATPを過剰に必要とする場合、フェレドキシン からプラストキノンへと電子を循環させH+ポンプの みを駆動する仕組みをもっているのである。しかし、
この反応を触媒する直接の酵素(フェレドキシン-プ ラストキノン還元酵素)はいまだに発見されていな い。チトクロムb6f複合体のストローマ側の分子表面 は非常に塩基性でLysやArgなどの塩基性のアミノ酸 が多く配置している。フェレドキシンはAspやGluの
多い酸性蛋白質なので直接b6f複合体とフェレドキシ ンが相互作用する可能性も考えられる。また他の蛋 白質を介した電子伝達カスケードが存在する可能性 も 否 定 で き な い 。 高 等 植 物 で は フ ェ レ ド キ シ ン -NADP+還元酵素(FNR)がb6f複合体のサブユニットと して存在しているという報告もあり14)、ヘムxがフェ レドキシン用のヘムであると言う仮説は大変興味深 いところである。
一般にクロロフィルは光エネルギーを吸収するア ンテナの役割を担うが、光に依存しないチトクロム
b6f複合体の反応系では非常に危険なラジカルを発生
させる“劇物“になってしまう。クロロフィルによ るラジカルの発生を抑えると考えられるβカロチン は、クロロフィルからは遠すぎる位置にあり、その 役割は不明である。チトクロムb6f複合体はLHCのリ ン酸化に働いていると言う話もあるので、もしかし たらこのクロロフィルは電子伝達とは全く関係の無 い働きを担っているのかもしれない。
光合成電子伝達を行う高等植物由来の膜蛋白質複 合体の結晶構造は、長い間報告されてこなかったが、
2003 年にイスラエルのNelson等によって高等植物由 来の光化学系Iの中分解能構造が報告された15)。高等 植物の光化学系Iは3量体構造を取らず、LHC 1と結 合した状態で存在しプラストシアニンとの結合に有 利なサブユニット構造を取っていると報告されてい る。高等植物では葉緑体中のチラコイド膜の形態に もバラツキがあり、ラメラ構造を取る領域では光化 学系IIとb6f複合体が、それ以外の領域では光化学系I
とb6f複合体が多いと言われている。先に述べた通り
高等植物のチトクロムb6f複合体ではラン藻と異なり
FNRをサブユニットとするとも報告されている14)。高
等植物とラン藻の光合成電子伝達において、分子レ ベルで異なった機能制御が行われている可能性があ るとするとクロロフィルaやβカロチンは高等植物 の複合体中で主に働くということも考えられる。今 後、高等植物由来のチトクロムb6f複合体の結晶構造 が明らかになると、そのあたりの役割が明らかにな ることであろう。
最後に、本稿で紹介したチトクロムb6f複合体の構 造研究は筆者が在籍していた米国Purdue大学生物科
15. A. Ben-Shem, F. Frolow and N. Nelson (2003) Nature, 426, 630-635.
学科のH.Zhang博士、J.L.Smith教授、W.A.Cramer教授 との共同研究の一部である。特に在籍中大変お世話 になったCramer教授に深く感謝したい。
1. S. E. Martinez, D. Huang, A. Szczepaniak, W. A.
Cramer and J. L. Smith (1994) Structure, 2, 95-105.
2. S. Iwata, M. Saynovits, T. A. Link and H. Michel (1996) Structure, 4, 567-579.
3. C. J. Carrell, H. Zhang, W. A. Cramer and J. L.
Smith (1997) Structure, 5, 1613-1625.
4. D. Xia, C.-A. Yu, H. Kim, J.-Z. Xia, A. M.
Kachurin, L. Zhang, L. Yu and J. Deisenhofer (1997) Science, 277, 60-66.
5. Z. Zhang, L. Huang, V. M. Shulmeister, Y-I. Chi, K.
K. Kim, L-W. Hung, A. R. Crofts, E. A. Berry and S-H. Kim (1998) Nature, 392, 677-684.
6. C. Hunte, J. Koepke, C. Lange, T. Rossmanith and H. Michel (2000) Structure, 8, 669-684.
7. G. Kurisu, H. Zhang, J. L. Smith and W. A. Cramer (2003) Science, 302, 1009-1014.
8. D. Stroebel, Y. Choquet, J-. L. Popot and D. Picot (2003) Nature, 426, 413-418.
9. E. A. Berry, L-S. Huang, L. K. Saechao, N. G. Pon, M. Valkova-Valchanova and F. Daldal (2004) Photosynthesis Res., 81, 251-275.
10. W. A. Cramer and D. B. Knaff (1991) Energy Transduction in Biological Membranes, chap. 6, Springer-Verlag, New York.
11. H. Zhang, G. Kurisu, J. L. Smith and W. A. Cramer (2003) Proc. Natl. Acad. Sci. USA., 100, 5160-5163.
12. V. Schunemann, A.X. Trautwein, J. Illerhaus and W.
Haehnel (1999) Biochemistry, 38, 8981-8991.
13. J. P. Whitellegge, H. Zhang, R. Taylor and W. A.
Cramer (2002) Mol. Cell. Proteomics, 1, 816-827.
14. H. Zhang, J. P. Whitelegge and W. A. Cramer (2001) J. Biol. Chem., 276, 38159-38165.
FACE(開放系大気CO
2増加)を用いた冷温帯落葉樹への高CO
2付加実験 小池
こ い け
孝良
た か よ し
1)
・江口
え ぐ ち
則和
の り か ず
2)
・笹
さ さ
賀
か
一郎
い ち ろ う
1)
1)北海道大学・北方生物圏フィールド科学センター
2)北海道大学大学院・環境科学院・森林圏環境学コース(日本学術振興会研究員)
解 説
はじめに
化石燃料の大量消費と森林の伐採などによって、
大気中CO2濃度は急激に増加し、温室効果ガスとされ るCO2によって地球温暖化が懸念されている。現在、
大気中CO2濃度は約 350~400ppmVであるが、2100
年には約540~970ppmVに達すると予測されている1)。
さらに、温暖化の影響は特に中高緯度地帯で顕著で あると言われている2)。陸域生態系では広い面積を占 める北方冷温帯林への影響が大きく、それに伴う地 球環境への影響も深刻であると予測されている。こ の予測の精度を上げるために、北方森林を構成する 樹木の高CO2環境下での生理的応答と成長反応につ いての多面的な研究が必要とされる。
北大北方生物圏フィールド科学センター・森林圏 ステーションでは、札幌研究林実験苗畑全体を実験 室として造り替え、高CO2環境での北方林構成樹木 の成長反応を研究している。このような大規模実験 を行う理由は、1991年に指摘されたポットサイズ効 果に端を発した1)。従来、CO2濃度を厳密に制御した 自然光型人工気象室でポット植えの材料を生育させ て実験してきた1)。しかし、人工気象室で育てられた 材料は、対照区との比較においてのみ変化を解析で きるが、その結果を野外での現象説明には一部しか 適用出来ない。
現在CO2の付加実験は、このような人工気象室を用 いた閉鎖的環境で制御する方法と、開放的な環境で 制御する方法の2種類に大別される。本稿では、CO2
付加実験における手法の特徴と問題点を紹介し、札 幌研究林のFACEでの研究結果とこれからの研究の 展望を紹介する。
CO2付加手法の特徴と問題点
生育環境を厳密に制御する方法としては、人工気 象室が挙げられる(写真1A)。人工気象室ではCO2濃 度だけではなく、温度、湿度、光量なども制御でき る。しかし、人工気象室のサイズの制限から、ポッ トに植栽した苗木を用いることが多く、ポットのサ イズによって根系が制限を受け3)、成長や光合成作用 が左右されることが指摘されている1)。また、ガラ スで外界と遮断された人工気象室内での実験では、
光の量や質が屋外と異なり4)(特に通常のガラスで は紫外線領域が遮断される)、室内の実験結果を屋外 の現象説明に直には適用できない。
その後、根系の制限をなくし比較的自然に近い状 態でCO2を付加するため、天井の開いたチャンバーを 用いる方法(オープン・トップ・チャンバー(OTC)) が使用された(写真1B)。1980年代後半からはOTC を用いた研究が盛んに行われた。しかし、周囲をガ ラス等で覆っているために、チャンバー内の環境は 外部環境とは大きく異なる。事実、Moya et al.5)はOTC では気温上昇や日射量の減少が見られた点を指摘し た。
現在、最も自然に近い状態でCO2を付加することの できる施設がFACEである(写真1C~E)。FACEは「Free Air CO2 Enrichment」の略であり「開放系大気CO2増 加」と訳される6)。FACEは閉鎖的な囲いをせず、CO2 が大気より重いことを利用して囲いの中へ風上から CO2を供給する方法であり、高CO2への植物の反応を 調査するには理想的な方法である。しかしながら、
維持経費(主にCO2経費)が人工気象やOTC法に比べ て20倍以上になる、気温上昇の影響を調べるのは困 難である、など課題も多い1)。
1987年に米国で始まったFACEは、現在、世界中で 農作物、牧草、樹木(写真C)、自然植生を対象に研 究が実施されている7,8)。スイス・バーゼル大学には、
樹高 35mに達するヨーロッパブナ、サクラ、シナノ キ類等とヨーロッパトウヒの針広混交林に樹冠クレ ーンを設置し、16 本の個体の枝先全てにCO2付加用 チューブを巻き付け、葉のごく近くにCO2付加する方 式のweb-FACEが設けられた8,9)(写真1D)。
日本ではイネのFACE実験(Rice-FACE)が1998年 から岩手県雫石町で行われたが6)、これまで樹木など の野生植物のFACE実験は行われていなかった。その ため、北海道大学札幌研究林に設置されたFACEは、
樹木を扱うFACE研究としては日本初の試みとなっ た(写真E)。
研究の概要
我々のFACEは札幌研究林実験苗畑に設置された
直径6.5メートル、高さ5メートルの六角柱を概観と
し(写真1E)、合計3基が約80m離れて設置されて
いる。CO2は枠組みの側面に蛇行状に配置したチュー ブから放出される。FACE枠の一角に設置したセンサ ーが風向を計測し、常に風上側のチューブを開けて
FACE内のCO2濃度が約500ppmVになるように制御し
ている。この濃度は現状のCO2増加速度(1.5ppmV/
年)が維持されると約2040年頃に実現する値である。
CO2が十分にある場合には、土壌の栄養条件は大き な成長制限要因になる6,7)。そこでFACE内の地面を半 分に区切り、半分が褐色森林土、残りが北海道に特 徴的な未成熟火山灰土壌(以下、火山灰土壌)とし、
土壌の栄養条件がどのような影響を樹木の成長にも たらすかを調査している。なお、火山灰土壌では褐 色森林土に比べて窒素量は約1/2、可給態リンは1/10 程度、カリ量は約1/15と少ない。植栽樹種はすべて 2年生稚樹であり、シラカンバやケヤマハンノキなど
写真1.CO2付加システムの変遷
A: 8連式人工気象室(森林総研北海道支所)
B: 16基のオープントップ・チェンバー(森林総研東北支所)
C: FACE(米・ディーク大学7基のサークルが設置されている)
D: web-FACE(スイス・バーゼル大学のシステム)
E: mini-FACE(北大札幌研究林)
遷移前期樹種、ヤチダモやハリギリなどの中間樹種 とイタヤカエデやブナなど遷移後期樹種の計11樹種 である。FACEの半面当たり各樹種8~9本、合計95 本を植栽した。予算の制約からCO2の過度な消費を抑 えるため、風速が3m/秒、光強度が光補償点以下(夜 間)になるとCO2付加を停止している。
CO2を付加しない対照区も主風の方向を考慮して
FACEから距離を置いて 3つ作り、FACE内と同じ条
件 下 で 植 栽 し た 。 対 照 区 の 日 中 のCO2濃 度 は 約
370ppmVであった。本研究で高CO2環境下での針広混
交林成立の初期過程を予測できることを期待してい る。
FACE での研究項目
高CO2環境下で生育させた植物の多くで光・CO2飽 和での光合成速度の低下が報告されている6,7)。これ は光合成産物のシンクとソースのバランスが崩れる ためで、光合成産物の転流が阻害され、葉に過剰に 蓄積したためと考えられている9)。特に、小さなポッ トで生育させた場合、シンク器官となる根系の発達 が抑制され根端にもストレスがかかり、光合成能力 の低下が顕著となる(負の制御)3,6,7)。ところがFACE によるCO2付加では根系に制限はかからない1)。この ため、FACEで生育させた植物はグロースチャンバー で生育させた時の光合成反応と異なる可能性がある。
最近の測定によると、土壌条件にかかわらずケヤ マハンノキは高CO2処理によって光・CO2飽和の光合 成速度が増加したが、シラカンバでは低下した。光 合成速度の「負の制御」の原因を知るために葉のル ビ ス コ ( Rubisco: ribulose-1,5-bisphosphate carboxylase/oxygenase)とデンプン量を調べた。
この結果、ケヤマハンノキとシラカンバの高CO2処 理では葉のルビスコ量が対照区に比べると低下して いた。これに対して、デンプン量はケヤマハンノキ での集積はシラカンバに比べて少なかった。この原 因 は ハ ン ノ キ 類(Alnus sp.)に 特 徴 す る 共 生 菌 類
(Frankia sp.)が、光合成産物のシンクとして働 いていることが原因と考えられる。
さらに、光合成特性が変わるならば、個体の成長 特性にも変化が生じると考えられる10)。そこで、
FACEで生育した樹木の成長特性と形態変化を追跡 している。特に、将来の森林の光合成生産力をリモ ートセンシングによって広域解析・推定を行うため、
そのパラメータとなる葉面積指数(LAI:m2·m-2)を経 時的に計測中である。また、窒素・クロロフィル、
光阻害回避機構とされるキサントフィル系の分析11) も予定している。
一方、FACEによる高CO2環境下での地上部の成長 特性を調べた報告は数多くあるが12,13)、地下部の成長 特性に関しては、依然として不明な点が多い。そこ で、モジュールFACEでは、ライゾトロン(根圏観察 装置)を用いて、根の成長率や回転率を測定する予 定である14)。
さらに、高CO2環境で植物を生育させた場合、葉の C/N比が上昇することが知られており、これによって、
植食者-植物の関係に重大な影響をもたらすことが 予想される15,16)。例えば、葉の窒素濃度が低下する ことで、幼虫の成長が抑制されて世代交代が進まな い可能性がある。しかし反対に、一定量の窒素を幼 虫が摂取しようとするために被食量が増加すること も考えられる。植物側の変化では、高CO2環境で被食 防衛物質である総フェノールやタンニンが増加した という報告もある7,15,16)。しかし、これらは人工気象 室を用いた実験の結果であり、実際の屋外環境での 予測は依然として不明である。そのためFACEによる 研究が必要であると考えられる。
また、葉のC/N比が上昇すると、落葉落枝(リター)
は微生物にとって魅力の低下したものへと変化する。
特にC/N比の増加は難分解性のリグニンの増加にも つながり、リターの分解速度が低下することが考え られる。リターは森林内の物質循環の根幹を成すも のであり、リターの変化は生態系に大きな影響をも たらす17)。従って、高CO2環境でのリターの成分、
分解速度も測定し、「土壌機能」も測定する予定であ る。
これまでの研究結果と今後の課題
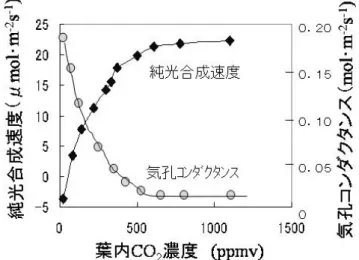
ここで、初年度の調査から興味深い結果を紹介し たい。高CO2環境では気孔が閉じ気味になることが知 られている7,8,10)(図1)。この事実がどのような生理
生態的現象をもたらすのかを検討するために、土壌 水分と葉温を測定した。すると、高CO2環境下では通 常環境に比べて、土壌水分(図2)、葉温ともに高い 値を示した。これらは気孔が閉じ気味になったこと による影響と考えられ、将来、植物の成長や、葉温 変化に伴う葉での酵素活性が変化する可能性も示唆 される。ただ、個葉の蒸散機能は低下するが個体と しては葉数が増加するので、個体としての蒸散機能 がどのように変化するかを検討中である。
ここで、気孔が閉じ気味になるということは、植 物体内の水分が葉から外に出づらくなるということ である。そのため、水の運搬を行う道管の形態にも 変化が起こることが予想される7,8)。つまり、水の利 用効率が上がるので、水を運ぶ道管の直径が小さく なると考えられる18)。ところが、Yazakiら13)は、人工 気象室でシベリアカラマツを育て、その細胞内腔が 富栄養の高CO2環境下で増加したことを報告した。し かしながら、これ以外の報告は少なく、FACEを用い て木部構造を調べた結果はほとんどない18)。今後、
数多くの樹種で調べていく必要がある19)。
図
1.葉内CO
2濃度と気孔コンダクタンス、純光合成速度の関係
モジュール FACE の今後の目標
これまで見てきたように、FACEを用いて研究すべ き課題は数多く挙げられる。モジュールFACEの目標 は、これらの研究課題を明らかにし、最終的には将 来の環境下で樹木がどれだけ炭素を固定、貯留でき るかを解明することにある。そこで、実験が終了す る2007年には育てた樹木を伐採して、炭素の貯留場
図
2
.土壌水分の変化の例。測定には「Dielectric Aquameter, Decagon Devices, Inc. NE, USA
」 を用いた。土壌水分の高い値は降雨時の値である。(Eguchi, N. et al.
を改作)20)12) Oren, R. et al. 2001. Nature, 411: 469-472.
所となる木部の構造が高CO2環境下でどのように変
化するのか調べることを当面の課題としている。 13) Yazaki, K. et al. 2001. Tree Physiol., 21 : 1223 -1229.
これまで経験しなかった速度で変化し続ける環境 の中では、生物間相互作用も変化する可能性が高い。
そのため、今後これら様々な疑問をより自然条件に 近い環境で解明しなければならない。このためには、
モジュールFACEは有力な手法となり得るであろう。
14) 里村多香美ら.2000. 根の研究, 11:15-23.
15) 小池孝良ら. 2003. 北方林業,55 : 14 -17.
16) Drake, B. G. et al. 1997. Ann. Rev. Plant Physiol. Plant Mol. Biol., 48 : 609 – 639.
17) 小池孝良ら. 1998. 北方林業, 50: 241-244.
なお、本研究の予算は文部科学省RR2002研究による
(代表:安岡善文:東大生産研)。実験施設の管理は、
奥谷 昭、福井富三、石田亘生、高畠 守諸氏(北 方生物圏フィールド科学センター:生物圏セ)の協 力を得た。丸山 温、北尾光俊、飛田博順(森林総研 北支)、岡田哲夫(東北農研)、小林和彦(農環研・現 東大)、Ch. Kőrner(Basel大), R. Oren (Duke大)、高木 健太郎、日浦 勉、柴田 英昭(生物圏セ)、船田 良、
平野高司(北大院農)各博士には数々の助言を頂い た。以上、記して感謝する。
18) Saxe, H. D. et al. 1998. New Phytol., 139:
395-436.
19) 江口則和. 2004. 日林北支論,52:66-68.
20) Eguchi, N. et al. 2005. Phyton (in press).
引用文献
1) 小池孝良ら. 1995. 森林立地学会誌, 37: 28-34.
2) IPCC 2001. Climate Change. The third assessment report of IPCC working group I.
Cambridge University Press, Cambridge, UK.
3) Arp, W. P. 1991. Plant Cell Environ., 14 : 869-875.
4) Lawlor, D. R. and R. A. C. Mitchell. 1991. Plant Cell Environ., 14 : 807-818.
5) Moya T. B. et al. 1997. Trans. ASAE., 40 : 739-747.
6) 小林和彦. 2001. 日作紀, 70 : 1-16.
7) 小池孝良. 2002. 河川文化. 10: 183-243.
8) 小池孝良. 2003. 光と水と植物のかたち (種生物 学会編). 119-138. 文―総合出版.
9) Pepin, S. and Ch. Kőrner. 2002. Oecologia 133:
1-9.
10) 牧野 周. 1999. 植物の環境応答 (渡邊 昭ら編 著). 134-141. 秀潤社.
11) 中路達郎ら 2003. 日林誌, 85: 205-213.
報 告 記 事
第5回日本光合成研究会シンポジウム報告
日本光合成研究会常任幹事 高橋裕一郎(岡山大学)
日本光合成研究会は村田紀夫前会長の時から年1回のシンポジウムを開催しています。昨年の名古 屋での常任幹事会で、本年の第5回シンポジウムは、光合成の若手研究者や専門外の研究者に光合成 研究の全体をカバーする入門的な内容にすること、そして常任幹事の高橋裕一郎(岡山大)、藤田祐一
(名古屋大)、寺島一郎(大阪大)、大政謙次(東京大)がお世話することが決定されました。シンポ ジウムのタイトルは伊藤繁会長と相談した結果、「光合成研究入門:地球の未来を語ろう!」としまし た。シンポジウムでの話題は広い分野をカバーするため、各世話人が「アンテナ色素系のつくり方」
(藤田)、「光エネルギー変換系研究の新展開」(高橋)、「光合成の代謝系」(寺島)、「光合成と環境・
生態・応用」(大政)のセッションを分担し、12名の演者の選定と講演依頼をしました。
シンポジウムは2005年5月28日(土)から29日(日)に名古屋大学野依記念学術交流館で 開催されました。シンポジウムの講演の他にポスター発表が16題、機器展示が5件ありました。こ の期間は愛知万博が開催中の週末にあたり、参加者は宿の確保に苦労したことと思いますが、大学院 生を含めた若手と光合成の専門家以外の方を含めおよそ100名の参加がありました。本シンポジウ ムでは日本光合成研究会の会員に講演を依頼する時は旅費などを自前でお願いすることが原則となっ ています。しかし、今回は会員以外の方にも講演をお願いしましたが、依頼したときに会員になって いただいたりして、手弁当で駆けつけて講演していただいたケースがありました。そのお陰で予算が 乏しかったにもかかわらず、シンポジウムの内容は予想以上に充実することができたと感謝しており ます。また、シンポジウム会場と懇親会の準備・運営は名古屋大学理学部の伊藤研究室のスタッフと 学生の方々のお世話なりましたが、この場を借りてお礼を申し上げます。
本シンポジウムにより、若手の研究者や大学院生および光合成以外を専門とする研究者が、光合成 研究の重要さや面白さを理解するきっかけとなったなら、目的は達成できたと思います。
**********************************************************************************
第5回日本光合成研究会シンポジウム
「光合成研究入門:地球の未来を語ろう!」に参加して
岡山大学大学院自然科学研究科 小澤真一郎
本シンポジウムは、5月28日から29日にかけて名古屋大学野依記念国際交流館で開催された。
シンポジウムでは、総説的な話が中心であったが、話題はタンパク質複合体の分子レベルの議論から、
代謝系、生態系や、さらに応用面に至るまでの幅広い分野を網羅していた。色素系を人工的に構築し ようとする試みや、光エネルギー変換システムも静的ではなく動的に捉えることを前面に打ち出し、
また、フィールド実験の結果から、光合成能力を強化するための試みも紹介された。二酸化炭素固定 の様式と、葉の構造との関わりを、二酸化炭素を葉がどのように取り込むかという疑問から始めて、
細胞に到達したあとの二酸化炭素の分子レベルでの拡散の議論にまで及んだ。ポスター発表も行われ、
熱心な議論が交わされた。ポスターは横長であったので、話の流れをつかむ上ではたいへん見やすか ったのであるが、ポスターボード全体が低かったので、横長の特徴を活かしきることができていなか ったと感じた。今回のシンポジウムでは、自分の研究領域以外の話題を聞くことができ、またポスタ ーも見ることができ、ともすれば自分の研究領域に閉じこもりがちな傾向を打破することができたと 考えている。また、別の分野からのアプローチを学ぶことで、今後の研究方針や計画に幅ができ、よ り自由な発想で研究を進めることができるようになったと思う。今後も、機会があればこういったシ ンポジウムには積極的に参加していきたい。
会場:野依記念国際交流館
ポスター発表風景
報 記 告 事
シアノ学会体験記
IVth ESF Research Conference on Molecular Bioenergetics of Cyanobacteria
-
EuroConference on Genomics, Proteomics and Structure for Functional Understanding
-参加報告 東京大学分子細胞生物学研究所 丸山真一朗スペインに行ってきました。
太陽と乾いた大地が僕を呼んでいたので。
空と海は旅人を祝福し、一週間よく晴れて暖かい日が続きました。
ラテンの國の人たちは、アジアの辺境の民を手厚く迎えてくれました。
その辺境の民の中に僕がいました。
園池先生もいらっしゃいました。
そんな訳で、この原稿を書いています。
初めまして。丸山真一朗と申します。
光合成研究とはほぼ無縁の日々を送って参りましたが、ひょんなことからお邪魔することになりまし た。しばしお付き合い頂ければ幸いです。
事の発端は我が研究室の廊下に(ここで言う我が研究室とは田中寛先生率いる東大分生研分子遺伝 研究分野のことですが)、一枚の簡単な学会の案内用ポスターが貼られていたのを目にしたことでした。
まず「シアノバクテリア」という文字に目が行き、次に開催地の「スペイン」という文字に目を惹か れ、次の瞬間には決意は固まっていました。そんな不純な動機で学問の世界に土足で踏み込もうとは
学会会場の遠景(矢印が会場となったホテル)
何とけしからん、と思われる方も多 い事でしょうが、まあそれはそれで そういうこともありますが、あの、
それはそれとして、ええと、ともか く場所は重要だと思う訳です。例え ば旧大陸と新大陸では、同じタイト ルで学会を行っても全く異なる内 容になるだろう、というように。
こ の 学 会 は
European Science Foundation (ESF)
からの支援を受け、2005
年5月21日から26日の6 日間に渡りスペインのSant Feliu de
Guixols
という、バルセロナから北上してジローナという街に出てさらに東へ行ったところにある地中海沿いのリゾート地で行われました。まだオフシーズンということで海水浴客、観光客は少なかった ものの、春先の逗子辺りを思わせる風光明媚なところでした。空港からバスがアレンジされており、
道中、旅の疲れと時差ぼけとでうつらうつらと揺られているうちに、バスはいつの間にか学会会場で
ある
Eden Roc
ホテルに着いていました。夕暮れに佇むホテルの玄関上の屋根には少しだけくたびれたネオンが煌煌と灯っており、初めて来たはずなのに何故かノスタルジーを喚起するような不思議な趣 を漂わせていました。
セッション会場の様子
Hans C. P. Mattijs
先生を議長として執り行われたこの会議では、ストラクチュローム全盛の世にあってか、構造に焦点を当てた発表が印象に残りました。
NDH-1
複合体やPSI
、PSII
の高次構造に関する 発表がいくつもあり、それぞれ独自の視点から高等植物研究に追いつけ追い越せと切磋琢磨されてい る様子が伺えました。個人的には、グローニンゲン大のBoekema
先生らによる、電子顕微鏡観察によ る多数のPSI
超複合体からの単一粒子投射再構成 像が圧巻でした。特にisiA
複合体の構造と機能に 関しては他の発表でも盛んに議論されており、注 目度の高さが伺えました。かずさDNA
研究所の 田畑先生によるY2H
解析に関する発表では、「興 味のある遺伝子があったら私宛にメイルを下さ い」という言葉に、会場からは、シアノコミュニ ティへの貢献に対する感謝と敬意、そしておそら くは労いの意味も込めて、大きな拍手が送られて いました。ワシントン大のPakrasi
先生はAMT
タ グを用いたPSII
に関する最新のプロテオーム解析 オーシャン・ビューのポスター会場と筆者の結果を発表されるなど、アメリカから参加されている方々の中には、技術面も含め全てにおいてア メリカは進んでるぜ、とでも言わんばかりの圧倒的な先進性を顕示されていた方も見受けられたよう に思いました。東大新領域の園池先生は自信作のジョークで会場全体に笑顔の華を咲かせ、ご自身も ご満悦のようでした。私も含め、数十のポスター発表も行われましたが、地中海に臨むガラス張りの ポスター会場などこの先いつ体験出来るか分からないような貴重な時間を過ごすことが出来ました。
会期の途中では近くの(といってもバスで高速を一時間くらいかっ飛ばして)ダリ美術館へのエク スカーションなどもあり、最終日前日の最後の晩餐ではフラメンコショーを堪能するなど、地中海に 面したカタルーニャ地方の自然から恵みを受け、それを芸術として精一杯昇華させようとする人間の 営みを目の当たりにすることが出来ました。願わくば、この自然の恵みを単なる技術としてではなく、
科学という形でも地球に還元することが出来れば、このような素敵な場所で学会を行って頂けたこと、
そしてそれに私自身が参加出来た意味がそこで初めて結実するのではないか、と日本へと向かう飛行 機の中でハリウッド版「
Shall we dance?
」を見ながらぼんやりと思ったのでした。私事で恐縮ですが、普段はシゾンという原始紅藻における光シグナル伝達経路の解析を行っている のですが、比較生物学的な視点を取り入れようということでシアノバクテリアの解析も始めたところ に、このような学会に参加する機会を得たのは本当に幸運でした。私のような門外漢を快く受け入れ て下さったシアノコミュニティの温かさは、じんわりと骨身に沁みました。いつか生物種の枠組みを 超えて、藻類もシアノも高等植物も入り乱れて活発な交流が行われることで、葉緑体共生進化の謎が 紐解かれて行くような時代が来ることを、趣味の問題もあるので飽くまで個人的な願望ですが、願っ て止みません。
最後になりましたが、写真をご提供などご協力頂いた埼玉大・日原先生、東大・村松君、及び日本
版「
Shall we
ダンス?
」における既婚男性心理の機微に関するご高見を頂いた埼玉大・仲本先生にこの場をお借りして御礼申し上げます。
タマネギ農家の皆さんと自然の恵み
(学会会場付近の畑にて)
記 告 事 報
Gordon Research Conferences on Photosynthesis に参加して
大阪大学蛋白質研究所 矢部俊樹
2005年7月3日から8日まで,米国ロードアイランド州,ブライアント大学において光合成に 関するゴードン会議が行われました。同じく参加されていた東工大の久堀先生からこの会議から感じ たことでよいので参加記事を書いて欲しい,との依頼を受けました。参加記事など書いた経験がない にも関わらず,安請け合いしてしまった自分を今頃になって後悔しつつ,単身で参加した一人の大学 院生が感じたものを今回報告したいと思います。
会議の行われたブライアント大学は,米国東海岸に位置するロードアイランド州の北部に位置し,
ボストンより車で約1時間のところにあります。私自身,ボストンは知っていたのですが,大学名ど ころかロードアイランド州自体知らず,‘島なのか?’等と愚かなことを考えてしまったのですが,全 米最長の州名である“State of Rhode Island and Providence Plantations”を正式名称として持つが,州面 積は全米最小なのだそうです。ブライアント大学は経済系の大学らしく,向こうの人も知らない人の 方が多かった様です。大学構内や建物は非常にきれいで,構内にある池には噴水もあり,木からはリ スも降りてくるような自然溢れる場所(田舎?)であります(写真)。参加者は基本的に全員同じ宿舎 に寝泊まりします。宿舎は,普段は学生寮として使われているものを使用するので,否応なしに典型
ブライアント大学構内の写真:大半の日本の大学とは 大きく異なった雰囲気を感じる
的なアメリカの大学生生活を味わうことになります。私の泊まった部屋は,大きな部屋の中に4つの 個室があり,トイレ,洗面台,シャワーを各個室に泊まっている人と共用することになります。私は 他人と寝起きまで共にするのは勘弁願いたかったので一人部屋にしましたが,望めば赤の他人とでも 部屋をシェアする事も可能で,その分参加費も少し安くなり,アメリカの大学院生気分をより満喫で きることになります。
会議は,日曜の夜から始まり,金曜の朝まで合宿形式行われました。1日のスケジュールは“朝食
〜午前のセッション〜昼食〜自由時間〜ポスターセッション〜夕食〜午後のセッション〜ポスターセ ッション”という感じで進行し,月曜の朝食後には集合写真の撮影が行われました(写真)。私はこの ことをすっかり忘れて部屋に帰っていたため,残念ながらこの写真には入れませんでした。食事はバ イキング形式で,自由席です。ですから,テーブル毎にグループができあがっている場合も多く,そ の構成メンバーを見ると,誰と誰が仲良しなのかが大方分かるのが興味深かったです。午後の自由時 間は各々がテニスに興じたり,大学構内にあるジムで体を動かしたり,研究に関するディスカッショ ンをしたり,時差ぼけなどで疲れていれば部屋で寝ていたり,と自由に過ごせることができます。私 はというと,有志により催されたサッカーに参加し,90分間走り回り汗を流していました。
会議の内容ですが,セッションのタイトルに関しては
HP
に掲載されているのでご興味のある方はそ ちらの方をご参照して頂けると幸いです(http://www.grc.org/programs/2005/photosyn.htm
)。ポスターは,100題を超えるほどあり,非常に充実している印象がありました。自身の英語力不足と分野外の内 容が多かった事によりセッションやポスターの内容の全ては理解できなかったのですが,光合成を研 究する人にとっては非常に魅力ある内容だったのではないかと思います。今回ポスターセッション時
集合写真:真ん中最前列の小さい方が今回のChairであるSabeeh Merchanta 氏である.
(写真は紺野氏(東工大)の提供による)
にワインテイスティング,参 加者が持ち寄ったワインを 飲み比べするということも 行われました。これに限らず ポスターセッションの時間 には毎回お酒などの飲み物 と軽食が提供されるため,ど ちらかというと懇親会的な 色合いがより濃くでている という印象でした。私も結局 その雰囲気に飲み込まれ(誘 惑に負け?),そこで知り合 った人たちと話をしたり,卓 球をしたり(久堀先生と),
真面目にポスターの前に立っていたのは結局初日だけとなってしまいました。最終日にはポスター賞 の発表があり,大学院生,ポスドク等若手研究者のポスター発表の中から私を含めた数名選ばれ,賞 状(写真)と賞金(後に判明)が授与されました。私の発表は,ある蛋白質がフェレドキシンや光化 学系Ⅰの鉄硫黄クラスターの生合成に関わるという昨年発表した論文の内容と,最近解いたこの蛋白 質の結晶構造についてでした。ポスターセッションの時間には残念ながら2−3名の人しか来なかった
(しかも初日だけ)のですが,受賞してしまいました。しかし,ポスターの前に10部近く別刷りを 置いていたのですが,私の気づかない間に全部持って行かれていたので,興味を持ってくれた人は多 かった様です。
ポスター賞の賞状
雑感として,ゴードン会議は普通の学会より多くの人と知り合うことが出来る,のがこの会議の最 もよい点ではないかと私は感じました。もちろん仲間内の同窓会的な側面もありますが,この会議で は今まで論文上で名前しか見ることの無かった人の顔が拝めるだけでなく,勇気さえあれば会話も交 わすことも出来るし,顔見知りになることも可能で,私のような立場の人はポスドク先を見つけるこ とも出来る。そして,その機会を様々な場(食事の時間,自由時間等)で提供してくれている事を感 じました。そして,これは欧米の研究者との交流にとどまらず,日本の研究者同士の交流も可能にし ており,今回に関して言えば,同じく参加されていた久堀先生や鹿内先生(九大)といった方々と私 の様な学生が会話する機会は,日本の学会では滅多にない事であると思われました。もしこのような 会議が日本でも定期的に行われれば,日本においても同世代や世代を超えた研究者間の交流が進むの ではないかと思いました。ということで,私のような若輩者が言うのもおこがましいことですが,日 本でも是非この様な会議が定期的に,出来れば光合成研究会が主催して,行って欲しいことを最後に 希望したいと思います。