Tel/Fax81138408810.Emailyosimizu@ˆsh.hokudai.ac.jp
平成 23 年度水産学会賞
魚類ウイルス病とその防疫防除に関する研究
吉
水
守
北海道大学大学院水産科学研究院
Studies on ˆsh viral diseases and those prevention and control methods MAMORU YOSHIMIZU
Faculty of Fisheries Sciences, Hokkaido University, Hakodate, Hokkaido 0418611, Japan
1. はじめに 1960 年 代 後 半 か ら , マ ス ノ ス ケ Oncorhynchus tschawytschaおよびベニザケ O. nerka をわが国に資源と して定着させようという試みが行なわれていた。その最 中の 1971 年に,道南のふ化場で飼育中のヒメマス O. nerka稚魚が大量に死亡した。翌年,道東のふ化場でも ヒメマス稚魚とアラスカから卵で移植したベニザケ稚魚 の大量死が起こった。当初は細菌病が疑われたが,原因 は伝染性造血器壊死症(IHN: infectious hematopoietic necrosis)ウイルスであることが判明した。1)当時,卒 業研究のサケ科魚類の腸内細菌叢に関する研究に没頭し ていたが,恩師木村喬久先生のニジマス O. mykiss 卵巣 由来細胞の培養を手伝い,IHN ウイルスによる細胞変 性効果を見る機会に恵まれた。それまで,わが国には IHN の発症例がなく,種卵および種苗の移動等の経緯2) から,アラスカから移植されたベニサケ卵と共に IHN ウイルスが道内に侵入したと考えられた。3)同じころ, アラスカでも本病の最初の発生が報告された。当時分離 した IHN ウイルスは,その後の遺伝子解析で,アラス カを含む北米のウイルスと同じタイプのウイルスである ことが明らかになった。4)1974 年から IHN は本州各地 のニジマス O. mykiss に広がり大きな問題となり,河川 湖沼養殖研究会で防疫対策の研究が開始され,5)後段で 紹介する防疫対策を対象魚種と病気に応じて組み合わせ る方法が功を奏し,現在,大量死は見られなくなってい る。1976 年から北海道内および東北地方のサケマス 類の採卵親魚を対象に病原ウイルスと細菌の分布調査を 始め,北海道内は現在も継続している。6)この間にサケ 科魚類からは新種のウイルスが 3 種見つかった。1978 年にサケ科魚類に肝炎を起こし,生き残った魚の口部に 上皮腫を引き起こす,魚で最初の腫瘍原性ヘルペスウイ
ルスが分離された。Oncogenic な Oncorhynchus masou 由 来 の ウ イ ル ス と し て Oncorhynchus masou virus (OMV) と名づけられた。7,8)2 番目は同年,サケからレ
オウイルスが分離され chum salmon virus (CSV) と名
づけられた。9)こちらは病原性がないウイルスであった。
3 番目は 1991 年にギンザケから分離され,神経に感染 して軸索を壊すために異常遊泳を引き起こすウイルス性 旋回病(VWD: viral wirhing disease)の原因ウイルス が分離された。10)翌 1992 年には北海道の多くの河川に 遡上したサケカラフトマスサクラマスから IHN ウ イルスが分離された。水産庁は事態を重視し,緊急に防 疫対策会議を開催し,全道の孵化場を対象に全魚種の発 眼卵を消毒することを決め,直ちに実施された。IHN ウイルスは翌年以降分離されることはなく,発眼卵消毒 は現在まで継続されている。海産魚の養殖および栽培事 業が盛んになるにつれ,1980 年代後半になって,それ ぞれの対象魚種でウイルス病が問題となり,その対策の 確立が急がれた。今回,これらのウイルス病とその防疫 対策について,その概略を紹介し,飼育魚種と対象疾病 ごとのリスク管理手法を導入した防疫対策導入の必要性 を紹介する。 2. 魚類のウイルス病研究の歴史 コイのポックス(pox)やカレイ類のリンホシスチス 病(LCD: lymphocystis disease)は,外観症状が特徴 的で慢性的に経過することから広く人々の目に留まり, 古くは 18 世紀に記載をみることができる。しかし,病 原体としてのウイルスの研究が始まったのは 1950 年代 になってからであり,11)まず,米国東部およびカナダの カワマスおよびニジマスの伝染性膵臓壊死症(IPN: in-fectious pancreatic necrosis)が濾過性の病原体による
ことが明らかになった。しかし,当時は魚類の培養細胞 がなく,ニジマスの尾鰭を用いた初代培養細胞により, 1960 年に原因ウイルスが分離された。同時期,米国西 部のベニザケならびにマスノスケに認められた風土病様 の病気も濾過性の病原体によることが明らかになった。 1960 年代に入り,ニジマスの生殖腺組織由来 RTG2 細胞12)およびマスノスケの胚由来 CHSE214 細胞が樹 立され,13)前記 IPN ウイルスをはじめベニザケおよび マスノスケからウイルスが分離された。このベニサケお よびマスノスケからのウイルスは IHN ウイルスと名付 けられ,14)IPNV および IHNV の分離が魚類ウイルスお よびウイルス病研究の始まりとなった。以来,次々と魚 類 由 来 株 化 細 胞 が 樹 立 さ れ , 1994 年 の Fryer and Lannan15)の総説には 34 科 74 種から樹立された 137 株 の魚類由来培養細胞が記載されている。これと並行して 魚類のウイルス病の研究も進み,原因が不明であったサ ケ科魚類のエグドベト病やコイの伝染性腹水症(正確に はその一部)がウイルス病であることが明らかとなり, ウイルス性出血性敗血症(VHS: viral haemorrhagic septicemia)および春ウイルス血症(SVC: spring vire-mia of carp)なる名称が提案され,原因ウイルスも分離 された。11)このような経緯から,魚類のウイルス病およ び原因ウイルスの研究は,まず,北米,欧州および日本 で産業的に被害の大きかったサケ科魚類およびコイ科魚 類が研究の対象となった。近年になって種々の魚類,甲 殻類,軟体動物が増養殖の対象になり,種苗生産施設お よび養殖場で魚類,エビ類および一部の貝類の病気が大 きな問題になっている。16) 魚類に致死性の病気を引き起こすウイルスのうち分離 培養が可能なものによる病気として,北米欧州日本 のサケ科魚類の IPN および IHN,日本のサケ科魚ヘル ペス ウ イル ス病 (OMVD) およ び ウイ ルス 性旋 回 病 (VWD),欧州のニジマスの VHS,伝染性サケ貧血症 (ISA: infectious salmon anemia),コイの SVC,コイヘ ルペスウイルス病(koi herpes virus disease),ウイルス 性乳頭腫症(viral papilloma),オーストラリアのレッ ド フ ィ ン パ ー チ の 流 行 性 造 血 器 壊 死 症 ( enzootic hematopoietic necrosis),アメリカナマズのウイルス病 (chanel catˆsh viral disease),シロチョウザメのイリド ウイルス病(white stergine irridoviral disease),ウナギ では日本およびヨーロッパのヘルペスウイルス性鰓弁壊 死症(herpesviral gill ˆlament necrosisus),日本のウナ ギの血管内皮壊死症(viral endothelial cell necrosis), ラブドウイルス性皮膚炎(rhabdoviral dermatitis)など があり,さらに海産魚のウイルス病としてはブリやヒラ メのウイルス性腹水症(viral ascites),ヒラメ等海産魚 のラブドウイルス病(hirame rhabdovirus disease),マ ダイ等多くの海産魚のイリドウイルス病(red seabream
iridoviral disease),ウイルス性神経壊死症(viral ner-vous necrosis)などが知られている。11,17) これらウイルスが分離されている病気に加え,原因ウ イルスの分離培養には成功していないが,ウイルス粒 子が病患部組織などに電子顕微鏡によって観察され,感 染試験によりウイルスが原因であることが確認されてい る病気も多くある。古くから知られているものとしては 種々の海産魚淡水魚の LCD(近年ヒラメやヨーロッ パヘダイの LCD ウイルスは宿主由来の培養細胞を用い ることで分離が可能になった),サケ科魚類のウイルス 性赤血球壊死症(viral erythrocytic necrosis),赤血球 封入体症候群(erythrocytic inclusion body syndrome), ウイルス性コイ浮腫症(viral edema of carp),キンギョ の ヘ ル ペ ス ウ イ ル ス 性 造 血 器 壊 死 症 病 ( herpesviral hematopoietic necrosis),アユの異形細胞性鰓病(atyp-ical cellular gill disease),ヒラメの表皮増生症(viral epidermal hyperplasia),トラフグの口白症(kuchijiro-sho)などがある。 また魚類の腫瘍の中には,上記のサケ科魚ヘルペスウ イルスによる基底細胞上皮腫,コイヘルペスウイルス (CyHV 1)による乳頭腫,パイクやウオールアイの肉 腫の他,ウイルスが原因と考えられるものがいくつか知 られている。18) わが国で報告された魚類のウイルス病を表 1 に示し た。上記のウイルス病および原因ウイルスに関しては多 くの教科書および総説解説が出版されている。代表的 なものとしては魚介類の感染症寄生虫病,19)改訂魚
病学概論,20)魚病学,21)新魚病図鑑,22)Fish Virus and
Fish Viral Disease11)などがあげられる。誌面の関係か
ら詳細については,これらを参照頂ければ幸いである。 わが国では栽培漁業の進展に伴い各地の栽培漁業セン ターあるいは種苗生産施設で多くの魚種の種苗が生産さ れるにつれ,それぞれの魚種で新しいウイルス病の被害 が報告されるようになった。特にシマアジ,キジハタ, ヒラメ,トラフグ,マツカワ等の海産仔稚に見られたウ イルス性神経壊死症およびヒラメ,キツネメバル等のウ イルス性表皮増生症は,一時各地で壊滅的な打撃を与 え,種苗生産時におけるウイルス病対策の重要性を提起 した。23,24)外国では北欧およびチリのタイセイヨウサケ
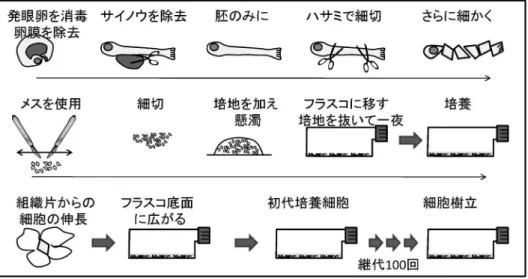
の IPN や伝染性サケ貧血症(infectious salmon anemia) が問題となっている。 3. 魚類培養細胞 魚類のウイルス病研究の基礎として,原因ウイルスの 分離同定,性状検査等に宿主由来の培養細胞が不可欠 である。前述のように 1960 年以降,サケ科魚類を中心 に魚類由来培養細胞が樹立された。日本でも,著者らの 研究室を主体に 2000 年の時点で 57 種の魚類由来細胞
表1 わが国で見られた魚類の主要なウイルス病 サケ科魚類病名 ウイルス原因 症 状 伝染性膵臓壊死症 IPNV キリモミ状旋回遊泳と突然の大量死 伝染性造血器壊死症 IHNV 貧 血 と 体 表 の V 字 状 出血を伴う大量死 サケ科魚ヘルペスウイ ルス病 OMV 肝炎と体表の潰瘍および腫瘍の形成 赤血球封入体症候群 EIBSV 赤 血 球 の 細 胞 質 に 封 入体,極度の貧血 ウイルス性旋回病 VWDV 回転を伴う旋回遊泳 ウイルス性赤血球壊死症 VENV 赤血球の細胞質に封入体 レオウイルス感染症 CSV 環境変化による死亡 温水魚病名 ウイルス原因 症 状 ウイルス性乳頭腫腫症 CyHV1 コイの鰭の上皮腫 ウイルス性コイ浮腫症 VECV 浮腫 ウイルス性眠り病 VECV 遊泳緩慢 コイヘルペスウイルス病 KHV 鰓の壊死,眼房陥没,粘液分泌 ヘルペスウイルス性造 血器壊死症 CyHV2 キンギョの造血器壊死 ウイルス性血管内皮壊 死症 VECNV ウナギの血管内皮細胞の壊死 ヘルペスウイルス性鰓 弁壊死症 AngHV1 ウナギの鰓弁壊死 ラブドウイルス性皮膚炎 EeRhabd ウナギの皮膚炎 海産魚病名 ウイルス原因 主な感染魚 リンホシスチス病 LCDV マ ダ イ , ヒ ラ メ , ス ズキ,ブリ ウイルス性腹水症 YTAV ブリ,ヒラメ ウイルス性変形症 YTAV ブリ ヒラメラブドウイルス病 HIRRV ヒラメ,クロダイ ウイルス性出血性敗血症 VHSV ヒラメ ウイルス性表皮増生症 FHV ヒラメ,キツネメバル,マツカワ マダイイリドウイルス病 RSIV マダイ,ブリ ウイルス性神経壊死症 VNNV シマアジ,キジハタ,マツカワ,クエ 口白症 sDNAV トラフグ 特定疾病 ウイルス原因 症 状 流行性造血器壊死症 ENV 体色黒化,運動失調,摂餌低下 ウイルス性出血性敗血症VHSV サケ科貧血と筋肉内出 血,成魚も発症 コイ春ウイルス血症 SVCV コイ科鰓の貧血と各臓器の点状出血 が継代維持されている。25)魚類細胞を作出する場合, 1960 年代から長くトリプシン消化法が用いられてきた が , 成 功 率 が 低 く 難 し い 手 法 で あ っ た 。 著 者 ら が Leibovitz の L15 培地を用いた直播き法(図 1)を紹介 して以来,日本国内をはじめ世界各地で本法により細胞 が樹立され,海産魚を含め多くの魚種で細胞が整備され た。 魚類培養細胞は Eagle の最小必須培地(MEM)を基 礎 培 地に , 炭 酸 緩衝 液 を 使 用 し て CO2イ ン キ ュ ベ ー ターを用いて培養した場合と Tris および HEPES 緩衝 液を用いた場合,さらに前述の L15 培地あるいは 199 培地を用いた場合の細胞の増殖を比較すると,いずれの 培 地 でも 増 殖 し , 海産 魚 由 来 細 胞 を 含 め , 食 塩 濃 度 0.116~0.171 M で増殖可能である。至適発育温度は棲 息水温に依存し,サケマス類由来細胞が 15~20°C, 温水魚由来細胞は 20~30°C である。染色体は 2n のも のが大部分であるが,継代数の増加につれ染色体数が増 加して 3 倍量になっているものが多い。一方で EPC 細 胞のように,半数(n)になっている細胞も存在する。 ヒトおよび家畜の細胞培養には CO2インキュベーター が広く用いられているが,魚類細胞の多くは培養温度が 低く,冷却装置を備えたインキュベーターが必要とな る。低温 CO2インキュベーターは高価となることから, L15 培地あるいは MEMTris 培地が広く用いられ, プレートにはシールを貼ることが一般的である。26,27) 細胞を凍結する場合,FBS あるいは MEM10Tris に DMSO,グリセリンあるいはレバンを 10 の割合に加 え,凍結時の温度勾配を 0.3~1.0°C/min とし,0°C か ら-60°C あるいは-80°C までこの条件で下げ,その後 液体窒素中に保存する方法を推奨している。この温度条 件は発泡スチロール製の断熱ボックスを使用しても得ら れ,この条件で細胞は少なくとも 10 年間は生存率 85 以上で保存されている。28,29)現在,25 年が経過している が保存状態に問題はない。-80°C では数年が限界とさ れてきたが,昨年 23 年を経過した EK1 細胞の培養が 可能であった。 なお,甲殻類のエビカニ類や軟体動物の貝類由来の 株化細胞はほとんどなく,これら水棲無脊椎動物のウイ ルス病研究に大きな障害となっている。株化は困難であ るが,血リンパ細胞の初代培養は可能であり,30,31)ホタ テ貝では筋肉エキス 10 添加海水培地にインシュリン を添加することにより 6 ヵ月程度は維持が可能となっ ている。 4. 防疫対策 魚類は稚仔魚期に免疫応答が成立するまでにかなりの 時間を要する。サケ科魚類の場合,液性免疫応答が見ら れるのは 0.3~0.5 g 前後,カレイ類のマツカワでは約 15 gである。32)現在,ワクチン開発が精力的に進められ ているが,ワクチンが利用できるようになっても,この 期間およびワクチン投与後,免疫応答が成立するまでの 期間は防疫対策を実施する必要がある。陸上施設での孵 化,種苗生産ではこの期間,有効な防疫対策を講じるこ とにより,病気の発生を抑えることができるようになっ
図 1 直播き法による魚類培養細胞の作出法 図2 サケ科魚類の増養殖場で実施されている防疫対策 ている。33,34)現在採用されている孵化場あるいは種苗生 産施設における疾病防除対策は,飼育器具機材および 施設の衛生管理,病原体フリー飼育用水の確保,健康親 魚の確保あるいは選別,卵洗浄および消毒,稚仔魚の健 苗性の確認のための病原体検査,飼育水温の管理,飼育 排水の殺菌等である(図 2)。34)飼育池あるいは生簀へ の移動および放流に際しての防疫対策は健苗性の確認と ワクチン投与である。さらに養殖に際しては耐病系統の 確立などが求められている。35)魚類防疫に関しても,魚 種および病原体ごとのリスク評価を行い,重要な管理点 の抽出とその実施記録が重要である。 1) 飼育器具機材および施設の衛生管理 作業者の手指や長靴をはじめ,陸上施設では飼育水 槽,飼育器具機材の微生物管理が病原体の伝播防止上 きわめて重要である。飼育水槽,飼育池,生け簀等の飼 育施設は種苗の搬入前に病原体を殺菌あるいは排除して おく必要があり,飼育器具および機材は日頃から常時消 毒し防疫に努める必要がある。33) 消毒薬は種類が多く,その作用機序も異なり,また病 原体によって感受性も異なる。さらに水棲生物は種ごと に飼育温度や塩分濃度も異なり,病原体ごとに適切な消 毒薬を選択する必要がある。36)各種消毒薬の中で,魚介 類に対し比較的毒性が少なく,除去が容易でかつ多量に 使用しても安価なものが用いられている。幸い魚類の病 原細菌およびウイルスは,いずれも市販の公称有効濃度 で十分殺菌不活化されるが,冬期間の低温下での使用 や飼育魚の糞残餌,体表粘液等有機物が付着した対象 物の殺菌には一部不適な消毒薬がある。塩素やヨードと
表2 水産分野における電解水の利用ガイドライン9) 目 的 有効塩素濃度と作用時間 処理法 魚類飼育用水の 殺菌 0.5 mg/L5 分間(1.0 mg/L1 分間) 脱塩素後に使用 飼育排水の殺菌 0.3~0.5 mg/L 0.5 mg/L では脱塩素後に放流 カキの浄化 0.3 mg/L 掛け流し 漁船漁具の殺 菌洗浄 0.5 mg/L 洗浄 市場の床岸壁 の殺菌洗浄 0.5~1.0 mg/L 洗浄 食品加工工程の 殺菌洗浄 0.5 mg/L 食塩電解水を使用 加工品の殺菌 洗浄 0.5 mg/L 食塩電解水を使用 現在食品添加物として認められているものは,次亜塩素酸ナト リウム,強酸性電解水,弱酸性電解水,食塩水を電解した食塩 電解水である。 いったハロゲン系の消毒薬は低温下でも効果の減少は見 られないものの,反復使用は避けるべきであり,アルデ ヒド系の消毒薬は反復使用に耐えるものの温度の影響を 受けやすい。魚介類の病原ウイルス細菌に限ってみれ ば,逆性石鹸液が臭いもなく両者の条件を満たしてい る。37)後述のオゾン処理海水あるいは電解海水を消毒剤 として使用することは,消毒剤購入経費の削減になり一 石二鳥の効果がある。38)さらに,消毒済み区域への立ち 入りに際しては専用の長靴を使用し,着衣も専用のもの に着替えるといった対策が必要である。33) 2) 病原体フリー飼育用水確保のための殺菌 飼育用水の殺菌に関しては吉水笠井38)に開発の経 緯と目的に応じた殺菌法について詳述した。水そのもの の理化学的性状を変えることなく大量の水の殺菌処理が 安定して行える装置が求められる。現在のところ,紫外 線,オゾンあるいは電解による殺菌が一般的である。紫 外線を用いる場合,病原体の紫外線感受性を求め,その 値の 5~10 倍程度の線量を照射している。魚介類の病 原体は紫外線感受性から,高感受性グループと低感受性 グループに分けられる。エンベロープを有する各種ウイ ルス,DNA ウイルスおよびグラム陰性細菌が高感受性 グループに含まれ,104mWsec/cm2の紫外線照射で 99.9 以上の殺菌あるいは 99 以上の感染性ウイルス の不活化が可能である。一方,2 本鎖 RNA ウイルスや グラム陽性細菌,カビ,原虫,寄生虫は感受性が低く 106mWsec/cm2程度の照射が必要である。水深は紫外 線の透過率(5 cm で 10 減衰)を考慮してなるべく浅 くとり,影になる部分がないよう水中の大型粒子を除去 する必要がある。また低温下では効力が若干低下する。 現在の紫外線ランプの出力は 8,000 時間経過で約 20 低下する。年一回の交換が望ましい。 海水をオゾン処理すると海水中のブロム等のハロゲン と反応してオキシダントが生成され,これが殺菌効果を 示す。もちろん,魚にも毒性を示す。殺菌効果はオゾン ガスの酸化力とオキシダントの効果がほぼ 11 であ る。淡水では曝気によりオゾンガスを除去後飼育用水と して使用可能であるが,海水ではまずオゾン処理水槽で 殺菌後,活性炭槽を過し残留オキシダントを除去して飼 育水とする。魚類病原細菌およびウイルスを 99.9 以 上殺菌あるいは不活化させる残留オキシダント濃度と処 理時間は 0.5 mg/L で 1 分程度であり,安全率を考慮し て 通 常 0.5 mg / L で 5 分 間 処 理 さ れ て い る 。38)こ の 場 合,飼育水中の一般細菌の生菌数は 99.9 以上減少す る。上記中圧紫外線殺菌装置を用いた 106mWsec/cm2 照射時の生菌数の減少率も 99.9 以上となり共に 3 桁 以上の減少を見る。この数値を魚類病原微生物を殺菌す る際の指標値としてもよい。オゾン殺菌装置はヨーロッ パを中心に水道水の殺菌に広く利用され普及している が,高価であり,メンテナンス経費等から水産増養殖分 野では,後述の海水電解装置への置き換えが進んでいる。 海水を電気分解すると食塩が分解され,塩化物イオン が酸化力の強い次亜塩素酸に転化し,これが殺菌効果を 示す。海水を白金コートチタン電極間を流すのみという 簡単な構造であり装置が単純で,かつ海水は無尽蔵であ り,経費的に最も安価である。飼育にはオゾン殺菌同 様,活性炭を用いた脱塩素処理が必要である。38)表 2 に 機能水学会が示した水産分野における電解水利用のガイ ドラインを示した。飼育海水の殺菌に用いる場合,有効 塩素濃度 0.5 mg/L で 5 分あるいは 1 mg/L で 1 分間の 処理後,活性炭を通して脱塩素を行って使用する。有効 塩素濃度 0.5 mg/L で 5 分の処置で海水中の一般細菌の 生菌数は 99.9 以上減少する。カキなど貝類やウニの 浄化には有効塩素 0.3 mg/L 以下の電解海水で 24 時間 飼育すれば大腸菌は陰性となり,39)ウニ腸管内の腸炎ビ ブリオは排除される。40)水産物の食品加工には食品添加 物として認められている食塩電解水を用いる。 その他,中空糸濾過膜を用いた濾過除菌やホットプ レートを用いた加熱殺菌あるいはヨードを滴下する方法 も有効であるが,電解殺菌以外は経済的な面や魚毒性等 で問題があり,実用化には至っていない。41) 3) 健康親魚の確保あるいは選別 採卵用親魚の健康状態の把握とその管理は,種苗生産 の成否を左右する。魚類の場合,一般に感染耐過してキ ャリアーになった個体は,成熟期に生殖産物,特に卵巣 腔液あるいは精液に病原体が出現する。催熟畜養中に病 原体を出す個体が存在すると,群全体に水平感染が起こ り,生み出された卵あるいは精子は病原体に汚染され, 孵化仔魚に感染する。このリスクを避けるために,採卵 用親魚候補個体の検査を実施し,催熟中の水平感染を防
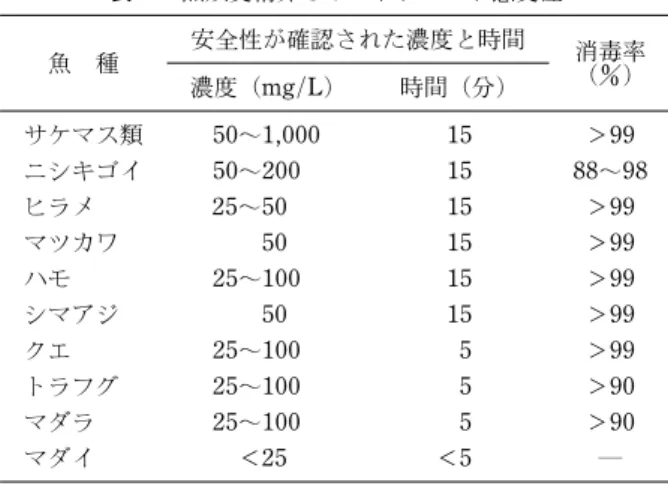
表3 魚類受精卵のポピドンヨード感受性 魚 種 安全性が確認された濃度と時間 消毒率() 濃度(mg/L) 時間(分) サケマス類 50~1,000 15 >99 ニシキゴイ 50~200 15 88~98 ヒラメ 25~50 15 >99 マツカワ 50 15 >99 ハモ 25~100 15 >99 シマアジ 50 15 >99 クエ 25~100 5 >99 トラフグ 25~100 5 >90 マダラ 25~100 5 >90 マダイ <25 <5 ― 表4 代表的な魚類病原ウイルスのポピドンヨード感受性 ウ イ ル ス 不活化濃度(mg/L) 時間 伝染性膵臓壊死症ウイルス 25 15 秒 伝染性造血器壊死症ウイルス 30 5 分 Oncorhynchus masouvirus 40 30 秒 ウイルス性出血性敗血症ウイルス 8 5 分 コイヘルペスウイルス 12 30 秒 ウイルス性神経壊死症ウイルス 25 5 分 止する。さけます類では,受精後発眼に至るまでに約 1 ヶ月を要するために,採卵時に卵巣腔液を採取し,細 菌およびウイルスの検査を行い,病原体保有状況を把握 している。42)北日本のヒラメおよびマツカワなどの異体 類では,天然海域での捕獲後,親魚候補個体は全て個体 標識され,施設への搬入時に抗体検査を実施し,高リス ク個体を排除している。43)さらに,成熟 3 ヵ月前に再度 検査を行い,親魚候補個体を選別している。採卵時に卵 および精子を対象に,RTPCR を用いてウイルス遺伝 子の有無を検査し,陽性個体があれば受精卵を廃棄して いる。上記の飼育用水の殺菌と親魚の検査により,ウイ ルス性神経壊死症の発生は見られなくなっている。 4) 卵洗浄および消毒 魚 の ウ イ ル ス が 卵 子 や 精 子 に 吸 着 す る こ と が 分 か り,44)ウイルスが受精時に精子とともに卵内に侵入する と,生まれてくる仔魚への感染が避けられないとの論議 が起こった。しかし,受精時に卵内に侵入したウイルス は胚に感染したのち,ある程度は増えるものの胚は死亡 し,ウイルスは臍嚢内容物によって不活化されることが 分かり,45)死亡した卵を除去し,正常発生卵を胚の安定 期である発眼期にポピドンヨード剤(50 ppm15 分) で消毒すると,卵表面に付着している病原体を殺すこと ができ,その感染を防止することができるようになった。 消毒した発眼卵を病原体がいない飼育用水でふ化させ る方法が世界的に広く用いられるようになり,その結 果,仔稚魚期の病気は激減し,サケマス類の増養殖事 業は順調に進展し,放流事業はもちろんニジマスやギン ザケ O. kisutch 養殖も産業として成り立つようになった。 等張液洗卵は,受精率を上げる目的で古くから実施さ れてきたが,卵表面の生菌数を減少させることにも効果 があることが報告され,垂直感染防止に有効であると注 目されている。細菌性腎臓病原因菌と冷水病原因菌は卵 内感染するとされてきたが,107CFU/ml 以上の細菌を 含む卵巣腔液(体腔液)中に卵が存在すると,細菌は卵 門から囲卵腔に侵入することが明らかになった。卵表面 の細菌およびウイルスの数は,1 回の洗卵で 1 桁減少 し,通常の洗卵 2 回,シャワー洗卵 1 回で約 4 桁減少 することが報告されている。46)受精後,ポピドンヨード で卵表面を消毒すれば,孵化施設への病原体搬入が阻止 できる。 他の魚種でも,この卵消毒が導入されているが,魚種 により卵径卵膜の厚み,消毒剤感受性が異なり,表 3 に見られるように,それぞれの魚種に適した消毒剤の濃 度と処理時間が検討されている。47,48)このときの消毒対 象となるウイルスの不活化濃度を表 4 に示した。卵消 毒では完全殺菌を求める必要はなく,対象とする病原体 を不活化あるいは殺菌できれば目的は達成される。魚類 ではマダイ卵が一番感受性が高く,安全に消毒できる濃 度は 10 mg/L5 分処理程度である。クルマエビのホワ イトスポット病対策には 5 mg/L5 分が採用されてい る。ウイルス性神経壊死症ウイルスのオキシダントによ る不活化は残留オキシダント濃度 0.5 mg/L で 30 秒で ある。マツカワ卵の場合,モルラ期に残留オキシダント 濃度 0.5 mg/L5 分あるいはポピドンヨード 50 mg/L 15 分で消毒するのが一番安定であり,ヒラメでも同 濃度での消毒の安全性が報告されている。 5) 稚仔魚の病原体検査健苗性の確認 孵化仔魚は親魚群毎に水槽に収容し隔離飼育を行う。 当然,飼育器具は各水槽専用とし,定期的に消毒を行 う。異常遊泳個体あるいは発症個体を見つけた場合は, 速やかに検査する。発症魚の検査には病患部を含む部位 のスタンプ標本を作製し,モノクローナル抗体を用いた 蛍光抗体法を用いて行う検査が最も早く精度良く診断で きる。発症の有無にかかわらず定期的に検査する場合 は,培養可能ウイルスは一度細胞に接種して 1~2 日培 養し,培養細胞を PCR に供する培養併用 PCR 法が最 も精度が高い。海産魚ではウイルス性神経壊死症,ウイ ルス性腹水症,ヒラメラブドウイルス病,マダイイリド ウイルス病には培養併用 RTPCR が,ウイルス性表皮 増生症には蛍光抗体法がリンホシスチス病には RT PCR が適している。採血が可能なサイズになれば,抗 体検査を行い感染履歴の把握に務めることも重要である。
6) 飼育水温の調節 サケマス類の伝染性造血器壊死症や赤血球封入体症 候群,ウイルス性出血性敗血症およびヒラメラブドウイ ルス病などのウイルス感染症は,水温が 20°C あるいは 15°C を越えると自然終息することが知られている。ヒ ラメラブドウイルス病では感染試験でも 15°C では死亡 が見られなかったことから,以後飼育水温を 18°C に設 定するよう指導がなされ,翌年から,わが国では発症報 告はなくなっている。49)細菌病ではコイキンギョの通 称“穴あき病”は 30°C への昇温療法が有効であり,鰻 養殖では加温養鰻の普及により路地池で見られた赤点病 等の病気は見られなくなっている。 7) 飼育排水の殺菌 飼育排水はその量が多く,前述の紫外線あるいはオゾ ンでの殺菌はコスト的に困難である。しかし,魚病対策 はもちろん環境対策からも効果的な排水の殺菌が必要で ある。前述のように海水を電気分解すると,次亜塩素酸 が発生する。この次亜塩素酸は,魚類病原微生物に対し 0.1~0.5 ppm,1 分の処理で良好な殺菌不活化効果を 示す。これに必要な装置は,チタン電極間に海水を通す のみの簡単な構造でよく,小型で安価であり,毎時 200 ~500 トンの飼育排水の生菌数を 99.9 以上減少させ ることができる。50)排水中に含まれる塩素の環境影響評 価を行い,適切な運転条件を設定すれば,排水の殺菌処 理が可能である。タンパク質を凝集させ加圧浮上などの 処理を行えば飼育排水の COD,SS,TOC,全リン量, 全窒素量およびアンモニア態窒素量を減らすことが可能 である。 8) 有用細菌による細菌叢の安定化 受精卵をヨード剤あるいはオキシダント海水で消毒 後,紫外線あるいはオゾンで殺菌した飼育用水を用いて 孵 化 仔 魚 を 飼 育 す る と , い わ ゆ る 病 原 体 フ リ ー (speciˆc pathogen free: SPF)魚が得られる。これによ り孵化場や種苗生産施設での大量死は見られなくなり, ワクチン開発の実験魚,バイオテクノロジー研究,病理 学研究に大いに貢献した。しかし,一部で放流時に細菌 感染症に罹りやすい,環境適応が悪いとの指摘があり, 飼育魚の細菌叢を正常細菌叢に近づける試みを実施して いる。サケマス類の場合,正常細菌叢の形成時期は免 疫応答成立時とほぼ一致し,それまでは環境中の細菌叢 の影響を受ける。51)そのため,なるべく早く正常細菌叢 に近づける必要がある。この場合,病原性がなく且つ抗 ウイルス物質や免疫賦活物質を産生する細菌を投与した 方が,より効果的と考えた。サケマス類やヒラメマ ツカワで,抗ウイルス物質産生腸内細菌を経口投与する ことにより,ウイルス病を制御することが可能となっ た。52)淡水養殖サケマス類の腸内細菌は Aeromonas 属であり,その中から抗 IHNVOMV 活性のあるもの を選び,その培養液を飼料(ペレット)に 10 の割合 で添加して与えると,腸管内でも抗ウイルス物質が産生 され,糞と共に排泄され,水槽内に蓄積した。他方,海 産稚仔魚の場合は,生物餌料のワムシやアルテミア卵を まず消毒し,孵化後,抗ウイルス物質産生細菌を添加す るとワムシやアルテミアの細菌叢を制御することがで き,海産魚の腸内細菌は Vibrio 属が優性なため,この 中 か ら 抗 IHNV OMV NNV 活 性 の あ る も の を 選 び,この抗ウイルス物質産生細菌が優勢となった生物餌 料を給餌すると,稚仔魚の腸内細菌叢も添加細菌が優勢 となり,腸管内容物に抗ウイルス活性が認められた。こ れらは糞と一緒に飼育水槽中に放出される。ヒラメとマ ツカワの場合,ともに飼育水槽の換水率が低いため,抗 ウイルス物質は水槽内に蓄積される。マツカワについて は 8 年間,抗ヘルペスウイルスラブドウイルスおよ びノダウイルス活性を有する細菌の経口投与を続けた が,投与菌による障害はなく,上記の防疫対策の効果と 相まって,ウイルス性表皮増生症およびウイルス性神経 壊死症の発生は見られなくなった。52) 9) 耐病系の選抜 病気が蔓延しても,必ず生き残る個体が存在し,一般 的には 10 世代で抵抗性を獲得した個体が優性となり生 物は耐病性を獲得すると云われている。魚類でも主要養 殖魚種を対象に選抜育種が行われているが,被害の大き いウイルス病では伝染性造血器壊死症のようにウイルス の変異が魚の耐病化を上回り,未だ耐病性獲得には至っ ていない。53)現在まで,伝染性造血器壊死症に抵抗性の あるギンザケとニジマスとの異種間交配により感受性の 低いギンザケの性質を受け継いだニジマスの選抜や,伝 染性造血器壊死症抵抗性を示すクローンニジマスの選 抜,54)ニジマス 4 倍体とブラウントラウトを掛け合わせ OMV 抵抗性の信州サーモン55)などが報告されている。 さらにリンホシスチス病耐病性遺伝子座をマーカーに選 抜した LCD 耐性ヒラメやホワイトスポット病抵抗性ウ シエビの樹立などが報告されている。しかし,伝染性造 血器壊死症抵抗性ニジマスおよびホワイトスポット病抵 抗性ウシエビはビブリオ病に弱く,ニジマスではワクチ ン投与で対応可能であるが,ウシエビは養殖系統とはな っていない。 10) ワクチン開発の現状 現在市販されているワクチンは,投与方法により浸漬 ワクチン,経口ワクチンおよび注射ワクチンに大別され る。まず,細菌性疾病であるビブリオ病に対する浸漬ワ クチンが承認され,ついで連鎖球菌症に対する経口ワク チンが販売された。マダイイリドウイルス病や連鎖球菌 症,類結節症に対する注射ワクチンが開発され,現在 3 種混合ワクチンも市販されている。56)注射は最も有効な 投与方法であるが,対象魚数万尾に接種するのは容易な
作業ではない。ブリ属およびヒラメを対象としたワクチ ン注射装置を開発し57)製造承認も得たが量販には至っ ていない。本装置には電気麻酔装置を組み込むことも可 能であり,注射時の麻酔の効き過ぎによる死亡や逆に麻 酔がかからない等の事故の防止に役立つ。課題としては 稚仔魚へのワクチン投与法の検討が挙げられる。現状と しては浸漬,噴霧あるいは経口投与法が有望である。弱 毒ウイルスの使用が効果的であり,ワクチン処理施設を 設け,確実に排水処理を実施すれば実現可能と考える。 11) インターフェロン誘導剤の活用 天然生薬の利用 中国やベトナム,タイでは,古くから薬草の直接経口 投与が行われている。漢方生薬あるいはハーブ抽出液に 抗ウイルス活性が認められ,これら抽出物を用いた,ウ シエビのイエローヘッド病やキャットフィッシュの運動 性エロモナス症の改善例が報告されている。5860)抗ウイ ルス活性とインターフェロン誘導活性を有する漢方生薬 抽出物を添加した餌料を,マダイに投与してマダイイリ ドウイルス病の死亡率軽減効果を観察し,餌料添加物と しての利用が可能となっている。
二本鎖 RNA ウイルスあるいは Poly (I:C)の利 用 インターフェロンを誘導する例として 2 本鎖 RNA ウ イルスの感染が知られている。IPN ウイルスに感染し た後に IHN ウイルスあるいは Aquabirnavirus に感染し た後に VHS ウイルスで攻撃すると大幅な死亡率の低減 が見られる例61,62)や CSV と IHN ウイルスの組み合わせ 等が報告されている。弱毒とは云え生きたウイルスを使 用することは環境への配慮を考えると好ましくなく,合 成 2 本鎖 RNA である Poly (I:C)を使用する感染防御 法が検討され,同様の効果が認められ,容量用法も提 案されている。63,64)魚体内残留もなく注射ワクチンとな らぶ将来の一つの防御法として有望視されている。さら に系統保存個体の発症時の治療法にも有効と考える。 5. おわりに 魚介類の種苗の生産放流を中心とする栽培漁業や沿 岸域での養殖が盛んになるにつれ,魚を人為環境下で管 理することが多くなり,病原体が侵入すれば抵抗力の弱 くなっている個体が感染発病し,飼育群全体に病気が 伝播する環境が形成されている。なかでもウイルスによ る病気は被害が大きく,その対策確立が急務であり,水 産関係のウイルス研究者の労力の大半はこちらに注がれ てきた。魚類ウイルスの種類は,今のところヒトや家畜 に比べればかなり少なく,これは魚類ウイルスの研究が 産業的に被害の大きい病気の原因ウイルスを対象に行わ れてきたことと,魚類甲殻類軟体動物のウイルスで 人や家畜に病原性を有するウイルスが分離されていない ことにより,医学獣医学領域の研究者の関心を引かな かったことも一因と考える。増養殖対象魚介類がより広 範囲になれば,今後も未知のウイルスによる病気が発生 する可能性があり,水産業の発展のためにも,増養殖の 対象となり得る魚種のウイルス保有状況調査を行うと共 に,現在実施されている防疫対策に加え,より効果的な ウイルス病対策を検討する必要がある。その場合,現在 飼育している魚を対象にした危害分析を行い,◯どうい うウイルスに感染する可能性があるか,◯感染したとき の発症率は,◯来源として考えられるものは,◯親魚は 健康か,◯洗卵および卵消毒は可能か,◯飼育用水は汚 染されていないか,◯飼育担当者は対応マニュアルを遵 守しているか等をあらかじめ検討し,リスク分析を行 う。次いで,◯親魚候補魚の健康状態評価を行ったか, ◯洗卵および卵消毒を行ったか,◯飼育用水は汚染され ていないか,可能性がある場合,殺菌処理を実施したか, ◯殺菌装置は正常に作動しているか,◯飼育者の作業衣 は適切に管理されているか,◯手洗器および長靴の消毒 槽の消毒液は交換されているか,◯魚の健康診断を実施 したか,◯対応ワクチンは準備されているか,◯そのワ クチン投与法は,等をチェックし,実施の有無およびそ の評価を記録として残しておく必要がある。防疫対策も 日進月歩であり,最新の知見手法を随時導入する必要 がある。 水圏に生息する生き物は多種多様であり,魚類のみな らず,甲殻類をはじめ軟体動物,藻類さらにはプランク トンまでも研究の対象に入れれば,多くの未知の病原体 が存在すると考えられる。現在まで,主に産業的に被害 の大きい魚類ウイルスや細菌,真菌,原虫を対象に研究 が展開されてきたが,非病原ウイルスや病原体の生態学 的な研究が今後の課題である。同じ増養殖現場でも,施 設の形態や規模の違いで,採用される対策が異なる。ま た海面をそのまま利用する場合は,病原微生物に感受性 の高い時期を避けるか,ワクチンを利用する以外に現実 的な対処法がない。今後新たな病原微生物に水産業が脅 かされないためにも,国内未侵入の疾病に対する防疫体 制を整えると共に,国内でも未侵入の地域に対しては同 様の対策を講じる必要がある。 魚類は陸上に暮らすヒトや家畜とは生活環境が大きく 異なる。ウイルスの侵入門戸も異なる。変温動物であり 実験動物としては扱いにくいように思われるが,適切な 管理をすればマウスやラットよりも容易であり,多数の 同腹飼育群が得られる利点がある。ニジマスを中心に実 験動物としての系群の確立やクローンの樹立が進み,ま た魚類の培養細胞もすでに数多く樹立されている。魚類 由来細胞は宿主の棲息温度に近い温度で培養しなければ ならないが,発育温度域が広く管理もしやすいなど多く の利点を有している。ウイルスの科が同じであれば抗ウ
イルス物質の作用は同一であり,ヒトや家畜に重篤な危 害を及ぼすウイルスに対する抗ウイルス物質の検索等利 用が広がっている。さらに魚類ウイルスや他の脊椎動物 のウイルスを比較することでウイルスの進化の過程も明 らかになってくると考えられる。今後の研究の益々の発 展を期待したい。 文 献
1) Kimura T, Awakura T. Current status of disease of cul-tured salmonids in Hokkaido, Japan. In: Proceedings from the International Symposium on Diseases of Cultured Salmonids, Sponsored by Tavolec Inc. Seattle, Washing-ton, April 46. 1977, 124160.
2) Urawa S. A review of sockeye salmon production in the Nijibetsu River in eastern Hokkaido, Japan. Tech. Rep. Hokkaido Salmon Hatchery1991;160: 310.
3) Yoshimizu M. Disease problems of salmonid ˆsh in Japan caused by international trade. Rev. Sci. Tech. OŠ. Int. Epiz. 1996;15: 533549.
4) Nishizawa T. Higashi S. Yoshimizu M. Nucleotide diversi-ty of Japanese isolates of infectious hematopoietic necro-sis virus (IHNV) based on the glycoprotein gene. Dis. Aquat. Org. 2006;71: 267272.
5) 山崎隆義,原 武.伝染性造血器壊死症,「養鱒の研究」 全国湖沼河川養殖研究会,養鱒部会編,緑書房,東京. 1976; 8388.
6) Kasai H. Nomura T. Yoshimizu M. Surveillance and con-trol of salmonid viruses of wild salmonid ˆsh returning to the northern part of Japan, from 1976 to 2002. In: Proceedings of the 3rdFiSCUP JapanKorea Joint Semi-nar on Fisheries Sciences, December 1516, 2003, Jinju-Tongyeong, Korea. 2003; 142147.
7) Kimura T. Yoshimizu M. Tanaka M. Sannohe H. Studies on a new virus (OMV) from Oncorhynchus masouI. Characteristics and pathogenicity. Fish Pathology 1981; 15: 143147.
8) Kimura T. Yoshimizu M. Tanaka M. Studies on a new vi-rus (OMV) from Oncorhynchus masouII. Oncogenic na-ture. Fish Pathology 1981; 15: 149153.
9) Winton JR. Lannan CN. Fryer JL. Kimura T. Isolation of a new reovirus from chum salmon in Japan. Fish Pathology 1981;15: 155162.
10) Oh MJ. Yoshimizu M. Kimura T. Ezura Y. A new virus isolated from salmonid ˆsh. Fish Pathology 1995;30: 23 32.
11) Wolf K. 「Fish virus and ˆsh viral disease」 Cornell Univer-sity Press, New York 1988; 476.
12) Wolf K. Quimby MC. Established eurythermic line of ˆsh cell in vitro. Science 1962; 135, 10651066.
13) Fryer JL. Yusha A. K. S. Pilcher KS. The in vitro cultiva-tion of tissue and cells of Paciˆc salmon and steelhead trout. Ann. N.Y. Acad. Sci. 1965;126, 566586. 14) Amend DF. Yasutake WT. R. W. Mead RW. A
hematopoietic virus disease of rainbow trout and sockeye salmon. Trans. Amer. Fish. Soc. 1969;98: 769804. 15) Fryer JL. Lannan CN. Three decades of ˆsh cell culture:
A current listing of cell lines derived from ˆshes. J. Tissue Culture Methods1994;16, 8794.
16) OIE World Organisation for Animal Health. 「Aquatic Animal Health Code」 14thEdition, Paris 2011; 286. 17) Kimura T. Yoshimizu M. Viral diseases of ˆsh in Japan.
Annual Rev. Fish Dis. 1991;1: 6782.
18) Yoshimizu M. Kasai H. Chapter 7, Oncogenic viruses and Oncorhynchus masouvirus, In 「Fish Diseases and Disor-ders」 Vol. 3, 2ndEdition: Viral, Bacterial and Fungal In-fections (eds. P.T.K. Woo and D.D. Bruno), CAB Inter-national. 2011; 276301. 19) 江草周三監修,若林久嗣,室賀清邦編.「魚介類の感染症 寄生虫病」恒星社厚生閣,東京.2004; 424. 20) 小川和夫,室賀清邦編.「改訂魚病学概論」恒星社厚生 閣,東京.2008; 192. 21) 畑井喜司雄,宗宮弘明,渡邉 翼.「魚病学」学窓社,東 京.1998; 157. 22) 畑井喜司雄,小川和夫.「新魚病図鑑」緑書房,東京. 2006; 295.
23) Yoshimizu M. (2003) Control strategy for viral diseases of salmonids and ‰ounder. In: Biosecurity in Aquaculture Production System: Exclusion of Pathogens and Other Undesirables, C. S. Lee and P. J. O' Bryen (Eds.), World Aquaculture Society, Baton Rouge, Louisiana, USA. 2003; 3541.
24) Muroga K. Viral and bacterial diseases of marine ˆsh and shellˆsh in Japanese Hatcheries. Aquaculture 2001; 202: 2344.
25) 吉水 守,木村喬久,西澤豊彦.日本国内で保管されて いる魚類由来株化細胞,「動物細胞工学ハンドブック」日 本動物細胞工学会編,朝倉書店,東京.2000; 319334. 26) Yoshmizu M. Kamei M. Dirakubusarakom S. Kimura T.
Fish cell lines: Susceptibility to salmonid viruses. In 「Invertebate and Fish Tissue Culture」 Kuroda, Y., E. Kurstak and K. Maramorosch (eds.), Jap. Sci. Soc. Press, Tokyo/Springer-Verlag, Berlin. 1988; 207210. 27) 吉水 守.魚類由来培養細胞のウイルス感受性,日水誌 1997;63: 245246. 28) 吉水 守.動物培養細胞および癌細胞の凍結保存―魚類 培養細胞,「凍結保存―動物植物微生物」酒井 昭 編,朝倉書店,東京.1986; 9496. 29) 吉水 守.魚類培養細胞の凍結保存法,「海洋生物のジー ンバンク―系統保存凍結保存」海洋1990;22(3): 154 158. 30) 小坂善信,吉水 守.ホタテ貝の閉殻筋着色異常,ワー クショップ「貝類の新しい疾病」,魚病研究 1999;34: 222.
31) Nagai T. Nakatsugawa T. Nishizawa T. Muroga K. Pri-mary culture of hemocytes from Japanese Black Abalone Nordotis discus discus. Fish Pathology1998;33: 147148. 32) 渡辺研一,吉水 守.大腸菌発現 VNN ウイルス外被タ
ンパク質に対するマツカワの液性免疫応答開始時期,魚 病研究2002;37: 9294.
33) 吉水 守,笠井久会.魚類ウイルス病の防疫対策の現状 と課題,化学と生物2005;43: 4858.
34) Yoshimizu M. Control strategy for viral diseases of salmonid ˆsh, ‰ounders and shrimp at hatchery and seeds production facility in Japan. Fish Pathology 2009; 44: 9 13.
35) 望月万美子,阿久津哲也,鴻上 繁,岡本信明,吉水 守.染色体操作により得られたニジマス 2 系統の耐病性 ならびに再生産形質に見られた差異,日水誌 2007;73: 844850.
36) Ahne W. Winton JR. Kimura T. Prevention of infectious diseases in aquaculture. J. Veter. Med. B/Zentralblatt fuer Veterinaer-medizin Reihe1989;36: 561567.
37) 木村喬久,吉水 守.水産養殖システムの殺菌,「新殺菌 工学実用ハンドブック」高野光男,横山理雄監修,サイ
エンスフォーラム,東京.1991; 220226.
38) 吉水 守,笠井久会.種苗生産施設における用水および 排水の殺菌,工業用水2002;523: 1326.
39) Kasai H. Kawana K. Labaiden M. Namba K. Yoshimizu M. Elimination of Escherichia coli from oysters using elec-trolyzed seawater. Aquaculture 2011; 319: 315318. 40) 木村 稔,三上加奈子,干川 裕,森 立成,笠井久 会,吉水 守.電解海水を用いた蓄養によるウニ内蔵か らのVibrio parahaemolyticus 除菌効果について,日水誌 2006;72: 15. 41) 吉水 守(共著)「海水殺菌装置評価基準」マリノフォー ラム21,東京.1991; 220. 42) 吉水 守,野村哲一.サケマス採卵親魚の病原微生物 検査法,魚と卵1989;158: 4959.
43) Watanabe K. Nishizawa T. Yoshimizu M. Selection of brood stock candidates of barˆn ‰ounder using an ELISA system with recombinant protein of barˆn ‰ounder ner-vous necrosis virus. Dis. aquat. Org. 2000;41: 219223. 44) Mulcahy D. Pascho RJ. Adsorption to ˆsh sperm of
verti-cally transmitted ˆsh virus. Science 1984; 225: 333335. 45) Yoshimizu M. Sami M. Kimura T. Survivability of
infec-tious haematopoietic necrosis virus in fertilized eggs of masu salmon and chum salmon. J. Aquat. Anim. Health 1989;1, 1320. 46) 小原昌和,小川 滋,笠井久会,吉水 守.養殖サケ科 魚類の人工採卵における等調液洗卵法の除菌効果.水産 増殖 2010;58: 3743. 47) 渡辺研一吉水 守.海水のオゾン処理により生成され るオキシダントを用いた飼育器具類および受精卵の消 毒,魚病研究 1998; 33: 145146. 48) 渡邉研一.病原体の制御および宿主機能活用による病害 防除技術の開発,水産増殖部会,2008. 49) 大迫典久,吉水 守,木村喬久.Rhabdovirus olivaceus (HRV)人工感染に及ぼす水温の影響.魚病研究 1988; 23: 125132. 50) 笠井久会,渡辺研一,吉水 守.流水式海水電解装置に よる飼育排水の殺菌,日水誌 2001;67: 222225. 51) 吉水 守,木村喬久,坂井 稔.サケ科魚類の稚仔魚期 における腸内細菌叢の形成時期について.日水誌1980; 46: 967975. 52) 吉水 守,絵面良男.抗ウイルス物質産生細菌による魚 類ウイルス病の制御,Microbes Environ. 1999; 14: 269 275.
53) Mochizuki M, Kim HJ. Kasai H. Nishizawa T. Yoshimizu M. Virulence change of infectious hematopoietic necrosis
virus against rainbow trout Oncorhynchus mykiss with viral molecular evolution. Fish Pathology 2009; 44: 159 165. 54) 望月万美子,阿久津哲也,鴻上 繁,岡本信明,吉水 守.染色体操作により得られたニジマス2 系統の耐病性 ならびに再生産形質に見られた差異,日水誌 2007;73: 844850. 55) 小原昌和,傳田郁夫.染色体操作による異質三倍体品種 「信州サーモン」の開発,水産育種 2008;37: 6166. 56) 吉水 守,吉田夏子.水産業で使用されている医薬品, 環境技術2008; 37: 860864. 57) 吉水 守笠井久会.水産用ワクチン注射装置の開発. 養殖 2004;41(9): 8283.
58) Direkbusarakom S. Herunsalee A. Yoshimizu M. Ezura Y. Antiviral activity of several Thai traditional herb ex-tracts against ˆsh pathogenic viruses. Fish Pathology 1996;31: 209213.
59) Direkubusarakom S. Tuangpan L. Yoshimizu M. Ezura Y. Protective e‹cacy of Clinacanthus nutans on yellow-head disease in black tiger shrimp (Penaeus monodon). Fish Pathology1998;33: 401404.
60) Direkubusarakom S. Ezura Y. Yoshimizu M. Herunsalee A. E‹cacy of Thai traditional herb extract against ˆsh and shrimp pathogenic bacteria. Fish Pathology. 1998;33: 437441.
61) Kim HJ. Oseko N. Nishizawa T. Yoshimizu M. Protection of rainbow trout from infectious hematopoietic necrosis (IHN) by injection of infectious pancreatic necrosis virus (IPNV) or poly (I:C). Dis. Aquat. Org. 2009;83: 105113. 62) Pakingking R. Jr. Takano R. Nishizawa T. Mori K. Iida Y. Arimoto M. Muroga K. Experimental coinfection with Aquabirnavirus and viral hemorragic septicemia virus (VHSV), Edwardsiella tarda or Streptococcus iniae in Japanese ‰ounder Paralichthys olivaceus. Fish Pathology 2003;38: 1521.
63) Nishizawa T. Takami I. Kokawa Y. Yoshimizu M. Fish-immunization using a synthetic double-strand RNA Poly (I:C) an interferon inducer, oŠers protecrion against RGNNV, a ˆsh nodavirus. Dis. Aquat. Org. 2009; 83: 115 122.
64) Nishizawa T. Takami I. Yoshimizu M. Oh MJ. Required dose of ˆsh nervous necrosis virus (NNV) for Poly (I:C) immunization of sevenband grouper Epinephelus septem-fasciatus. Aquaculture2011;311: 100104.