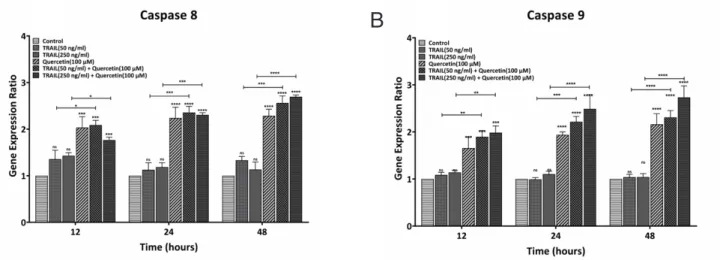
Quercetin elicits proapoptotic effect through downregulation of antiapoptotic and upregulation of proapoptotic genes in human myeloid leukemia KG-1 cells
6
0
0
全文
図

関連したドキュメント
An example of a database state in the lextensive category of finite sets, for the EA sketch of our school data specification is provided by any database which models the
Research Institute for Mathematical Sciences, Kyoto University...
A NOTE ON SUMS OF POWERS WHICH HAVE A FIXED NUMBER OF PRIME FACTORS.. RAFAEL JAKIMCZUK D EPARTMENT OF
Department of Orthopedic Surgery Okayama University Medical School Okayama Japan.. in
The inclusion of the cell shedding mechanism leads to modification of the boundary conditions employed in the model of Ward and King (199910) and it will be
A lemma of considerable generality is proved from which one can obtain inequali- ties of Popoviciu’s type involving norms in a Banach space and Gram determinants.. Key words
In [9], it was shown that under diffusive scaling, the random set of coalescing random walk paths with one walker starting from every point on the space-time lattice Z × Z converges
de la CAL, Using stochastic processes for studying Bernstein-type operators, Proceedings of the Second International Conference in Functional Analysis and Approximation The-
