87
Introduction
The venerid bivalve genus Saxidomus Conrad, 1837 commonly occurs on both the eastern and western sides of the Pacific. This genus includes Saxidomus nuttalli Conrad, 1837, S. gigantea (Deshayes, 1839) and S. purpurata (Sowerby, 1852). Of these, the first two live on the eastern Pacific side (Coan et al., 2000; Huber, 2010). Saxidomus purpurata, the only one species in the western Pacific, lives in sandy mud and gravel bottoms from the intertidal to 40 m in depth and is widely distributed, ranging from northern and eastern Hokkaido southward to Kyushu, Korea, the Bohai, Yellow and East China Seas, and the Pojet, Amur and Peter the Great Bays in Russia (Higo et al., 1999; Scarlato, 1981; Qi et al., 1989; Matsukuma, 2000, 2017; Yamazaki & Kashio, 2011; Kuwahara & Taru, 2012). As pointed out by Noda (1972), Saxidomus first appeared in a warm-water environment and gradually adapted to cooler temperate warm-water. However, it is uncertain when and where the genus adapted to the cold-water areas that correspond to the “cool-temperate” and “subarctic” areas of Nishimura (1981).
From the Pliocene Gobanshoyama Formation in Miyagi Prefecture, northern Japan, which is correlated with the Tatsunokuchi Formation, S. purpurata was collected in association with some characteristic species of the Pliocene Tatsunokuchi fauna (Nomura, 1938; Otuka, 1941) such as Anadara tatunokutiensis (Nomura & Hatai, 1936) and Fortipecten takahashii (Yokoyama, 1930) (Noda & Masuda, 1968). However, as the shell material of these fossils had dissolved, only a plaster cast of the outer surface of S. purpurata was illustrated by Noda & Masuda (1968). Moreover, the sandy facies make it difficult to determine the age of the formation from microfossils. However,
Evolution and Cold-water Adaptation of the Genus Saxidomus
(Bivalvia: Veneridae) in the Northwestern Pacific
Kazutaka Amano1* and Jun Nemoto2
1Department of Geoscience, Joetsu University of Education, 1 Yamayashiki,
Joetsu, Niigata 943-8512, Japan
2Department of Earth Science, School of Science, Tohoku University,
6-3 Aoba, Aramaki, Aoba-ku, Sendai, Miyagi 980-8578, Japan
Abstract: A subumbonal pit was found in the hinge of the venerid genus Ezocallista Kuroda in
Kamada, 1962, for the first time. This finding supports the hypothesis that Ezocallista is more closely related to Saxidomus Conrad, 1837 than to Callista Poli, 1791. Only one Recent species, Saxidomus purpurata (Sowerby, 1852) is known from the western side of the Pacific. This species lives in temperate to cold water around Japan, Korea and in cold water in Bohai Sea in China and the Pojet, Amur and Peter the Great Bays in Russia. The oldest reliable records of this species in cool-temperate water have been recovered from the uppermost Miocene to the lowermost Pliocene Tatsunokuchi Formation in Miyagi Prefecture, northeastern Japan. That occurrence suggests that Saxidomus first adapted to a cold-water environment in its northernmost habitat during the latest Miocene to early Pliocene.
Keywords: Ezocallista brevisiphonata, Saxidomus purpurata, subumbonal pit, Tatsunokuchi
Formation
* Corresponding author: [email protected]
the junior author (JN) found some well-preserved shells of this species from the Tatsunokuchi Formation in Sendai, Miyagi Prefecture.
Taxonomically, there are some opinions on the similarity between Saxidomus and Ezocallista Kuroda in Kamada, 1962 (see Matsubara, 2013 for the authorship of Ezocallista). The type species of Ezocallista is Saxidomus brevisiphonatus Carpenter, 1864, which is a senior synonym of Macrocallista chishimana Pilsbry, 1905 (Coan et al., 2000). Keen (1969) synonymized Ezocallista with Saxidomus. When Kamada (1972) redescribed Ezocallista kurodae Kamada, 1962 from the lower Oligocene Iwaki Formation, he changed Ezocallista to a subgenus of Saxidomus. On the other hand, some malacologists have treated Ezocallista as a synonym of the genus Callista Poli, 1791 (e.g., Scarlato, 1981; Matsukuma, 2000) or as a subgenus of Callista (e.g., Habe and Ito, 1965; Habe and Kosuge, 1967; Habe, 1977). However, Huber (2010) pointed out that Ezocallista is not closely related to Callista based on the hinge, pallial sinus and sculpture. Moreover, he also claimed that Macrocallista Meek, 1876 has an elongate shell and is monospecific, comprising only the Caribbean species M. nimbosa (Lightfoot, 1786). Therefore, it is necessary to clarify the status of Ezocallista in relation to Saxidomus, Callista and Macrocallista.
In this paper, we describe S. purpurata from the Tatsunokuchi Formation after establishing the taxonomical position of Ezocallista and discuss the cold-water adaptation of S. purpurata.
Materials and Methods
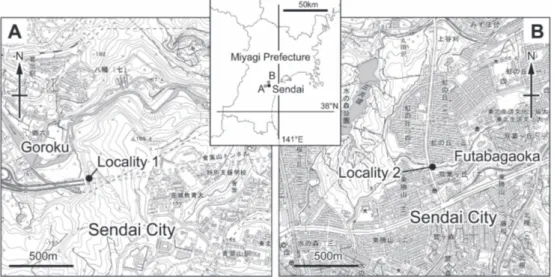
Fossil specimens of S. purpurata were recovered from two localities in the Tatsunokuchi Formation (Fig. 1). Locality 1 is a cliff at Goroku (38°15 36 N, 140°49 24 E) in Aoba-ku, Sendai City, Miyagi Prefecture. From this locality, eleven molluscan species were collected from ill-sorted tuffaceous fine- to coarse-grained sandstone yielding cobbles and boulders of andesite (Table 1). Anomia chinensis Phillippi, 1849 and Crassostrea gigas (Thunberg, 1793) are predominant, in association with some rocky-shore dwellers such as Barbatia sp., Chama sp., Haliotis sp., Serpulorbis sp. and Polyplacophora gen. et sp. indet. Based on the lithofacies and species composition, most shells were transported from the intertidal rocky shore and might have been deposited on a sandy bottom nearby. Locality 2 is an outcrop at Futabagaoka 2-chome (38°18 01 N, 140°52 01 E) in Aoba-ku, Sendai City, Miyagi Prefecture. From this locality, eleven
Fig. 1. Locality map of fossils (using the digital topographical map, scale 1:25,000, published by Geospacial
molluscan species were obtained from a shell bed (about 80 cm in thickness) whose matrix consists of pebble-bearing fine- to medium-grained sandstone. This shell bed is intercalated with fine- to coarse-grained sandstones. Among the fossils from the shell bed, Felaniella usta (Gould, 1861), Mya (Arenomya) oonogai Makiyama, 1935 and S. purpurata are predominant. The age of the Tatsunokuchi Formation was assessed at 5.5–5.0 Ma by Nakashima (2002), based on the diatom fossils.
We also have examined the characteristics of the following Recent shells collected from the beach; S. nuttalli, the type species of Saxidomus, from Monterey Bay in USA (IGPS 112411), S. purpurata from Harutachi in Hokkaido (IGPS 112412) and Amur Bay in Russia (IGPS 112413), and Callista chinensis (Holten, 1802) from Zushi in Kanagawa Prefecture (IGPS 112415). Ezocallista brevisiphonata (Carpenter, 1864) was collected from the beaches of Mutsu City in Aomori Prefecture (IGPS 112416), Nemuro City in Hokkaido (IGPS 112417), Aniva in south Sakhalin (IGPS 112418), Pachinka in southwestern Sakhalin (IGPS 112419) and from off Arutori Cape (25–30 m in depth) in Date City, Hokkaido (IGPS 112414). In the collection in the National Museum of Nature and Science, Tsukuba, we also examined S. purpurata from Shibasaki in Kanagawa Prefecture (NSMT-Mo 73980), Shinojima in Aichi Prefecture (NSMT-Mo 44172), and Koitoi Cape in northern Hokkaido as well as E. brevisiphonata from Shirikishinai in eastern Hokkaido (NSMT-Mo 76162).
All specimens treated herein are stored at the Tohoku University Museum (IGPS), Sendai, Miyagi Prefecture; the Departments of Geology and Paleontology (NMNS PM) and of Zoology (NSMT-Mo) of the National Museum of Nature and Science (NMNS), Tsukuba, Ibaraki
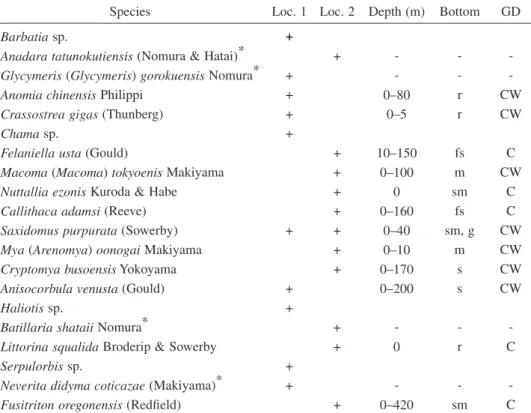
Table 1. Molluscan fossils from the Tatsunokuchi Formation. Abbreviations: m, mud; ms, muddy
sand; fs, fine-grained sand; s, sand; g, gravel; r, rock; GD, Geographic distribution; C, cold-water species; CW, temperate-water species. The data on depth, bottom and geographic distribution are after Higo et al. (1999). * extinct species.
Species Loc. 1 Loc. 2 Depth (m) Bottom GD
Barbatia sp. +
Anadara tatunokutiensis (Nomura & Hatai)* + - -
-Glycymeris (-Glycymeris) gorokuensis Nomura* + - -
-Anomia chinensis Philippi + 0–80 r CW
Crassostrea gigas (Thunberg) + 0–5 r CW
Chama sp. +
Felaniella usta (Gould) + 10–150 fs C
Macoma (Macoma) tokyoenis Makiyama + 0–100 m CW
Nuttallia ezonis Kuroda & Habe + 0 sm C
Callithaca adamsi (Reeve) + 0–160 fs C
Saxidomus purpurata (Sowerby) + + 0–40 sm, g CW
Mya (Arenomya) oonogai Makiyama + 0–10 m CW
Cryptomya busoensis Yokoyama + 0–170 s CW
Anisocorbula venusta (Gould) + 0–200 s CW
Haliotis sp. +
Batillaria shataii Nomura* + - -
-Littorina squalida Broderip & Sowerby + 0 r C
Serpulorbis sp. +
Neverita didyma coticazae (Makiyama)* + - -
Prefecture; Gunma Museum of Natural History (GMNH), Fujioka, Gunma Prefecture; the Mizunami Fossil Museum (MFM), Mizunami, Gifu Prefecture; the Department of Earth Science, Shimane University (T), Matsue, Shimane Prefecture and the Saito Ho-on Kai Museum (SM), Sendai, Miyagi Prefecture (now stored at the NMNS).
Taxonomy Relationship between Saxidomus and Ezocallista
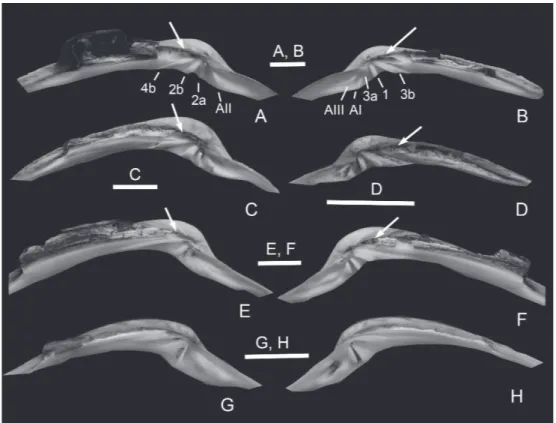
According to Habe (1977), Saxidomus differs from Ezocallista by having a subtruncated posterior margin, posterior gape, no lunule, many commarginal ribs and a “weak” anterior lateral tooth. The presence of a lunule is the most obvious distinguishing feature for discriminating between adult specimens of these genera (Fig. 2). However, some younger specimens of Saxidomus (less than 37 mm in length) have a weak lunule incision (Fig. 2C, E). Our observation of the hinge structure is slightly different from Habe’s (1977) statement regarding the anterior tooth. Namely, the anterior lateral tooth (AI, AII) of Saxidomus is not weak, but rather strong and steeply sloping like the cardinal teeth (Fig. 3A–D). In the right valve, the anterior lateral tooth (AI) is larger than the anterior cardinal tooth (3a) and almost touches it. It is easy to separate both genera based on these characters. Moreover, we have newly found a subumbonal pit in Ezocallista (Fig. 3E, F). The subumbonal pit was defined as “a deeply excavated, hollow structure just behind the posteriormost dental element” by Horikoshi (1987). Previously, he referred to the presence of the pit in Saxidomus, citing personal communication from Dr. A. Matsukuma. Consequently, both Saxidomus and Ezocallista have the subumbonal pit. The pit was neither illustrated nor described by Habe (1951, 1977) or Coan et al. (2000). The subumbonal pit of Ezocallista is usually smaller than that of Saxidomus as seen from well-preserved modern shells. However, in poorly preserved Recent shells and fossils, the subumbonal excavation becomes larger than in well-preserved shells. Such a subumbonal pit is also known in the following taxa: the arcticid Arctica Schumacher, 1817 (Horikoshi, 1987), the vesicomyids Hubertschenckia Takeda, 1953, Archivesica Dall, 1908, Ectenagena Woodring, 1938 and some Pleurophopsis van Winkle, 1919 (Horikoshi, 1987; Amano & Kiel, 2007; Amano et al., 2019), and the venerids Cyclina Deshayes, 1850, Kaneharaia Makiyama, 1936 and Dosinia Scopoli, 1777 (Horikoshi, 1987; Amano & Hikida, 1999) as well as the psammobiids Hiatula Modeer, 1793 and Nuttallia Dall, 1898 (Amano & Ogihara, 2012).
Although the hinge structure of Callista and Macrocallista is similar to that of Ezocallista, they lack a subumbonal pit. Moreover, Callista has nearly parallel anterior and middle cardinal teeth in the right valve. Based on the molecular phylogeny and shell microstructure, E. brevisiphonata is more closely related to S. purpurata rather than to Callista chione (Linnaeus, 1758), the type species of Callista, or to M. nimbosa, the type species of Macrocallista (Shimamoto, 1986; Chen et al., 2011a, b). However, as Ezocallista can be easily separated from Saxidomus in having a different hinge, the presence of a lunule in adult shells and no posterior gape, it can be considered as a separate genus.
Description of the Tatsunokuchi specimens
Family Veneridae Rafinesque, 1815 Subfamily Callocardiinae Dall, 1895
Remarks: Previously Saxidomus was included in Pitarinae Stewart, 1930 (e.g., Keen, 1969;
Coan et al., 2000) or Callistinae Habe & Kosuge, 1967 (e.g., Habe, 1977). We follow Huber (2010) and Coan & Valentich-Scott (2012) who pointed out that Pitarinae and Callistinae are junior synonyms of Callocardiinae. Their treatments are supported by the results of molecular phylogeny by Chen et al. (2011a, b).
Fig. 2. Recent shells of Saxidomus, Ezocallista and Callista. A, B, D. Saxidomus nuttalli Conrad, IGPS
112411, Loc. Monterey Bay, California in USA; A, outer surface of left valve; B, inner surface of left valve; D. dorsal view. C, E, F, G. Saxidomus purpurata (Sowerby); C, E, NSMT-Mo 44172, Loc. Shinojima, Aichi
Prefecture; F, G, IGPS 112412, Loc. Harutachi in Hokkaido; F, outer surface of left valve; G, inner surface of left valve. H–J. Ezocallista brevisiphonata (Carpenter), IGPS 112414, Loc. off Arutori Cape in Hokkaido;
H, dorsal view; I, outer surface of left valve; J, inner surface of left valve. K–M. Callista chinensis (Holten),
IGPS 112415, Loc. Zushi in Kanagawa Prefecuture; K, dorsal view; L, outer surface of left valve; M, inner surface of left valve. White arrows show the deepest point of pallial sinus. Abbreviation: lu, lunule. Scale bars = 10 mm.
Genus Saxidomus Conrad, 1837
Type species: Saxidomus nuttalli Conrad, 1837 (original designation). Saxidomus purpurata (Sowerby, 1852)
(Figs 2C, E–G; 3C, D; 4A, B, D, E, G–L; 5A–D, F–H) Tapes purpurata Sowerby, 1852: 692, pl. 150, figs 124, 125.
Saxidomus purpuratus – Lischke, 1871: 127, pl. 9, figs 4, 5; Brauns, 1881: 40, pl. 5, fig. 20; Yokoyama, 1920: 127, pl. 9, figs 8, 9; Yokoyama, 1922: 153, pl. 12, fig. 9; Kinoshita & Isahaya, 1934: 16, pl. 12, fig. 88; Habe, 1951: 165, fig. 381; Taki & Oyama, 1954: 43, pl. 10, figs 8, 9, pl. 32, fig. 9; Fujie, 1958: 673, pl. 27, figs 20, 21; Ozaki, 1958: 128, pl. 23, fig. 1; Kira, 1959: 142, pl. 56, fig. 9; Yamamoto & Habe, 1959: 95, pl. 7, figs 7, 8; Sawada, 1962: 84, pl. 7, fig. 6; Kira, 1962: 160, pl. 57, fig. 9; Kaseno & Matsuura, 1965: pl. 19, fig. 14; Habe & Ito, 1965: 138, pl. 46, fig. 5; Sakagami et al., 1966: pl. 8, fig. 2; Golikov & Scarlato, 1967: 108, pl. 11, fig. 4; Habe & Kosuge, 1967: 152, pl. 57, fig. 8; Ohara, 1968: 82, pl. 14, fig. 4; Noda & Masuda, 1968: 5, pl. 1, fig. 9; Iwai & Shiobara, 1968: pl. 2, fig. 7; Kuroda et al., 1971: 645, pl. 93, figs 5–7; Noda, 1972: 685, pl. 4, figs 1, 2; Matsuura, 1977: pl. 4, fig. 7; Habe, 1977: 270, p. 56, fig. 3; Scarlato, 1981: 380, fig. 383; Zhao et al., 1982: 108, pl. 15, figs 7, 8; Fujiyama, 1982: 350, pl. 175, fig. 1965a, b; Masuda et al., 1983: 21, pl. 4, fig. 15; Akamatsu, 1984: 18, pl. 5, fig. 8; Fig. 3. Hinge structure of Saxidomus, Ezocallista and Callista. A, B. Saxidomus nuttalli Conrad, IGPS
112411, Loc. Monterey Bay, California in USA; A, hinge of left valve; B, hinge of right valve. C, D. Saxidomus purpurata (Sowerby); C, hinge of left valve, IGPS 112412, Loc. Harutachi in Hokkaido; D, hinge of right valve, IGPS 112413, Loc. Amur Bay in Russia. E, F. Ezocallista brevisiphonata (Carpenter), IGPS 112414, Loc. off Arutori Cape in Hokkaido; E, left valve; F, right valve. G, H. Callista chinensis (Holten), IGPS 112415, Loc. Zushi in Kanagawa Prefecuture; G, hinge of left valve; H, hinge of right valve. White arrows show a subumbonal pit. Scale bars = 10 mm.
Ogasawara et al., 1986: pl. 76, fig. 1; Okamoto & Ibaraki, 1988: pl. 1, figs 10, 11, pl. 2, figs 1, 2; Okutani et al., 1989: 164, fig. 144; Qi et al., 1989: 215, pl. 11, fig. 14; Noda et al., 1993: 164, figs 21–2, 3; Sakanoue, 1998: 44, fig. 4; Nagamori, 2000: 11, pl. 11, fig. 5; Noda et al., 2004: 86, fig. 12–15; Nakagawa, 2018: pl. 17, fig. 2.
Saxidomus nuttalli Conrad – Nomura & Hatai, 1936: 127, pl. 14, fig. 9.
Macrocallista brevisiphonata (Carpenter) – Nomura, 1938: 259, pl. 36, figs 1–3. Saxidomus nomurai Hatai & Nisiyama, 1952: 133 [nomen nudum].
Saxidomus purpurata (Sowerby) – Oyama, 1973: 102, pl. 45, fig. 8, pl. 46, figs 4, 5; Ishii, 1987: 18, pl. 17, fig. 4; Baba, 1990: 302, pl. 38, fig. 8; Nemoto & Akimoto, 1990: pl. 13, fig. 3; Tsuchida & Kurozumi, 1996: 9, pl. 4, fig. 1; Matsukuma, 2000: 1017, pl. 506, fig. 70; Min, 2004: 463, fig. 1526; Qi, 2004: 300, pl. 164A; Xu & Zhang, 2008: 249, fig. 786; Huber, 2010: 409; Kurihara, 2010: 63, fig. 25 E–G; Lutaenko & Noseworthy, 2012: 83, pl. 37, figs A–D; Zhang et al., 2016: 347, fig. 432; Matsukuma, 2017: 1249, pl. 544, fig. 7.
Saxidomus purpuratus (Sowerby) – Takayasu, 1980: 144, pl. 4, figs 2–5, pl. 5, fig. 4. [= Saxidomus cf. purpurata (Sowerby)]
? Saxidomus cf. purpuratus (Sowerby) – Arkhipova et al., 1992 [1994]: 226, pl. 42, fig. 1; Noda et al., 1995: 68, figs 9–9, 10.
non Saxidomus purpuratus (Sowerby) – Sakanoue & Itoigawa, 1997: 9, figs 5.1–5.4. [= Saxidomus sp.]; Sakanoue, 1998: 44, figs 18.5–18.7, 19.1–19.3. [= Saxidomus sp.]; Sakanoue, 2000: figs 5.1, 5.3 [= Saxidomus? sp.].
non Saxidomus purpurata (Sowerby) – Nemoto & O’Hara, 2003, pl. 6, fig. 1. [= Pseudamiantis? sp.]
Material examined: One specimen from Loc. 1 (IGPS 112410) and six specimens from loc. 2
(IGPS 101966).
Description: Shell medium in size, attaining 77.7 mm in length, ovate, equivalve and
inequilateral. Antero-dorsal margin broadly arcuate, gradually transitioning into semi-circular anterior margin; postero-dorsal margin straight and gently sloping, forming blunt angle with subtruncated posterior margin; ventral margin broadly arcuate. Posterior ventral end flexed outward, forming narrow gape. Umbo well inflated and located at anterior one-fourth to one-third of shell length (AL/L = 0.27, 0.32; Table 2); beak distinctly prosogyrate. Lunule and escutcheon absent. Pallial sinus probably deep but its top unobservable. Shell surface weathered but sculptured by fine commarginal ribs. Inner ventral margin smooth. In right valve, anterior cardinal tooth (3a) small and vertical, almost touching steeply sloping anterior lateral tooth (AI); middle cardinal tooth (1) inclined posteriorly; posterior cardinal tooth (3b) strong and weakly bifid; subumbonal pit deep just above posterior cardinal tooth. In left valve, anterior lateral tooth (AII) rather strong and inclined anteriorly; anterior cardinal tooth (2a) thin and vertical; middle cardinal tooth (2b) stout and slightly inclined posteriorly; posterior cardinal tooth (4b) long and very thin; subumbonal pit deep just above posterior cardinal tooth.
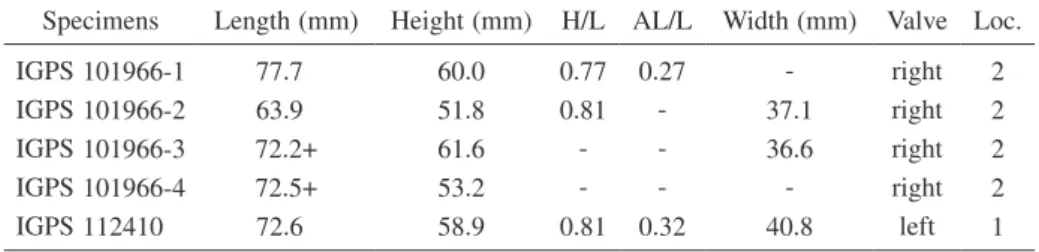
Table 2. Measurements of Saxidomus purpurata (Sowerby) from the Tatsunokuchi Formation.
H = height, L = length, AL = anterior length before beak, + = slightly longer than the value because of imperfect shells.
Specimens Length (mm) Height (mm) H/L AL/L Width (mm) Valve Loc.
IGPS 101966-1 77.7 60.0 0.77 0.27 - right 2
IGPS 101966-2 63.9 51.8 0.81 - 37.1 right 2
IGPS 101966-3 72.2+ 61.6 - - 36.6 right 2
IGPS 101966-4 72.5+ 53.2 - - - right 2
Remarks: The Tatsunokuchi species has a hinge referable to Saxidomus, and a high shell shape
(H/L = 0.77, 0.81; Table 2) as well as a well inflated umbo, both of which are characteristics of S. purpurata. When Sato & Matsushima (2000) studied the relation between the shell morphology and the habitat of Recent specimens of S. purpurata, they showed the range of mean values for four populations (H/L = 0.74 to 0.81) which includes the ratio of the Tatsunokuchi specimens. Although Nomura (1938) described some specimens as Macrocallista brevisiphonata (Carpenter, 1864) [= E. brevisiphonata] from the Tatsunokuchi Formation at Goroku (Loc. 1 in this study), his specimens have the same characteristics as S. purpurata, such as shell outline, surface sculpture, absence of lunule, pallial sinus shape and hinge structure (Fig. 5A–D, F–H). Thus, M. brevisiphonata of Nomura (1938) is misidentification of S. purpurata.
Saxidomus purpurata can be distinguished from the type species, S. nuttalli by its smaller and higher shell (maximum length = 156 mm; H/L = 0.64–0.72 in S. nuttalli; Coan & Valentich-Scott, 2012) and well inflated umbo. Another Recent species, S. gigantea from Alaska to California, resembles S. purpurata. Apart from a differently colored inner surface, S. gigantea has a larger shell (maximum length = 136 mm; Coan et al., 2000) than S. purpurata, a less inflated umbo and coarse and low commarginal ribs.
Nomura & Hatai (1936) described and illustrated one imperfect right valve from the lower upper Miocene Kubota Formation in Fukushima Prefecture as S. nuttalli. They treated S. purpurata as a junior synonym of S. nuttalli. The illustrated specimen (SM 2668 [= NMNS PM 21427]; Fig. 4D, E, K) is slightly different from the Recent S. purpurata in having a wider hinge plate and less inflated umbo. Hatai & Nisiyama (1952) proposed a new name, Saxidomus nomurai for the specimen illustrated by Nomura & Hatai (1936). However, this is a nomen nudum because they did not give any definition of the new species (International Commission on Zoological Nomenclature, 1999: Article 13.1). Kurihara (2010) described and illustrated specimens with a similar outline from the lower upper Miocene Itahana Formation in Gunma Prefecture. Examination of the left valve hinge of the voucher specimens (GMNH PI 2238) shows it to have a similar hinge plate to that of the extant S. purpurata. These morphological differences from the extant species can be interpreted as intraspecific variation and the Kubota and Itahana species are thus here identified as the oldest known records of S. purpurata. Takayasu (1980) described some deformed venerids from the uppermost middle Miocene Matsue Formation at Nange in Shimane Prefecture as S. purpurata. Although the specimens examined (T1017, T1026) have the characteristic hinge of Saxidomus (Figs 4C, 5E), it is difficult to certainly identify the species because of their poorly preserved surface, produced umbo and subtruncate anterior margin.
Some specimens from the middle Miocene Omori Formation in Shimane Prefecture were illustrated and described as S. purpurata by Sakanoue & Itoigawa (1997), Sakanoue (1998) and Sakanoue (2000). Among them, Sakanoue (1998) illustrated a Recent specimen of S. purpurata (fig. 4 in Sakanoue, 1998) for comparison with his fossil specimens. However, the poor preservation of the shell surface and the lack of description or illustration of the inner structure including the hinge prevent us from definitely identifying these species.
Arkhipopa et al. (1992) [1994] described Saxidomus cf. purpurata from the Pliocene middle part of the Maruyama Formation in the Makarov Region of southeast Sakhalin. Although the outline is similar to S. purpurata, they did not describe the hinge of their specimen. It is difficult to identify the species with certainty as S. purpurata.
When Nakagawa (2018) described Saxidomus sp. from the upper lower Miocene Shimo Formation at Oguroi in Fukui Prefecture with an illustration of the Recent S. purpurata (pl. 17, fig. 2 in Nakagawa, 2018), he separated his species from S. purpurata by its higher and more inflated shell and strong commarginal ribs.
Nemoto & O’Hara (2003) illustrated a well-preserved right valve of “Saxidomus purpurata” from the Kubota Formation mentioned above. However, their specimen has the wide commarginal
Fig. 4. Fossil shells of Saxidomus and Ezocallista. A, B, G–J, L. Saxidomus purpurata (Sowerby) from the
Tatsunokuchi Formation; A, hinge of left valve, IGPS 6, Loc. 2; B, hinge of right valve, IGPS 101966-4, Loc. 2; G, right valve, IGPS 101966-3, Loc. 2; H, right valve, IGPS 101966-1, Loc. 2; I, dorsal view of both valves, IGPS 112410, Loc. 1; J, left valve, IGPS 101966-1, Loc. 2; L, left valve, IGPS 112410, Loc. 1.
C. Saxidomus cf. purpurata (Sowerby); hinge of left valve of Saxidomus purpuratus (Sowerby) by Takayasu
(1980) from the Matsue Formation, T1017, Loc. Nange in Matsue City, Shimane Prefecture. D, E, K. Saxidomus purpurata (Sowerby) from the Kubota Formation described as Saxidomus nuttalli Conrad by Nomura & Hatai (1936); SM 2668 [= NMNS PM 21427], Loc. Okada in Tanagura Town, Fukushima Prefecture; D, hinge; E. dorsal view; K, right valve. F. Ezocallista brevisiphonata (Carpenter) recorded as
Saxidomus nuttalli Conrad by Nomura & Hatai (1939), NMNS PM 19642, Loc. Kurosawa in Akita Prefecture, dorsal view. White arrows show a subumbonal pit. Abbreviation: lu, lunule. Scale bars = 10 mm.
ribs and many fine radial threads near the umbo that are never observed in Saxidomus but that do occur in Pseudamiantis Kuroda, 1933.
Nomura & Hatai (1939) also listed S. nuttalli from Kurosawa in Akita Prefecture. However, as the poorly preserved specimen (NMNS PM 19642; Fig. 4F) has a lunule, a shallow pallial sinus,
Fig. 5. Fossil shells of Saxidomus. A–D, F–H. Saxidomus purpurata (Sowerby) from the Tatsunokuchi
Formation at Goroku misidentified as Macrocallista brevisiphonata (Carpenter) by Nomura (1938); SM 2277 [= NMNS PM 18148]; A, hinge of left valve; B, hinge of right valve; C, dorsal view of left valve of A; D, dorsal view of right valve of B; F, inner surface of left valve; G, outer surface of right valve; H, inner surface of left valve. E. Saxidomus cf. purpurata (Sowerby) from the Matsue Formation described as Saxidomus
purpuratus (Sowerby) by Takayasu (1980); T1026, Loc. Nange in Matsue City, Shimane Prefecture. White arrows show a subumbonal pit in A, B and deepest point of pallial sinus in F, H. Scale bars = 10 mm.
a small subumbonal pit and a long anterior lateral tooth, it can be identified as E. brevisiphonata. As the generic name Saxidomus was previously misunderstood as masculine, most researchers treated S. purpurata as S. purpuratus. Although Oyama (1973) noticed it to be feminine and corrected the species name as purpurata, many researchers continued to misuse the masculinized species name as purpuratus after 1973.
Measurements: See Table 2.
Distribution: ?late middle Miocene, Matsue Formation in Shimane Prefecture; early late
Miocene, Kubota Formation in Fukushima Prefecture (Nomura & Hatai, 1936) and Itahana Formation in Gunma Prefecture (Kurihara, 2010); latest Miocene to early Pliocene, Tastsunokuchi Formation (this study); early Pliocene, Gobanshoyama Formation in Miyagi Prefecture (Noda & Masuda, 1968), ?Hitachi and Kume Formations in Ibaraki Prefecture (Noda et al., 1993, 1995, 2004); ?Pliocene Middle Maruyama Formation in Makarov Region on Sakhalin (Arkhipova et al., 1992 [1994]); late Pliocene, Kitakanegasawa Formation in Aomori Prefecture (Amano, in press) and Ogikubo Formation in Nagano Prefecture (Nagamori, 2000); early Pleistocene, Nakanokawa [= Setana] and Tomikawa Formations in Hokkaido (Sawada, 1962; Sakagami et al., 1966), Daishaka and Noheji Formations in Aomori Prefecture (Iwai, 1965; Iwai & Shiobara, 1968); Omma Formation in Ishikawa Prefecture (Kaseno & Matsuura, 1965), the deposit from off Yamaguchi Prefecture (Okamoto & Ibaraki, 1988) and Haneji Formation in Okinawa Prefecture (Noda, 1972); middle Pleistocene, Zaimokuzawa Formation in Hokkaido (Fujie, 1958; Akamatsu, 1984) and Toriyamahama Formation in Miyazaki Prefecture (Nakao, 1993); late Pleistocene, Katanishi Formation in Akita Prefecture (Ogasawara et al., 1986), Hiratoko Shell Bed in Ishikawa Prefecture (Matsuura, 1977), Katori and Narita Formations in Chiba Prefecture (Yokoyama, 1922; Ozaki, 1958; Ohara, 1968), Tokyo Formation in Tokyo Metropolis (Brauns, 1881), Otsu Formation (Yokoyama, 1920) in Kanagawa Prefecture; Holocene, Nanba Formation in Osaka Prefecture (Ishii, 1987); Recent, Rebun Island in northern Hokkaido (Kuwahara & Taru, 2012) and Akkeshi Bay in eastern Hokkaido (Yamazaki & Kashio, 2011) to Kyushu, Japan Sea, Korea, Yellow Sea and Bohai Sea, Pojet and Peter the Great Bays (Higo et al., 1999; Scarlato, 1981; Qi et al., 1989; Matsukuma, 2000, 2017; Qi, 2004).
Discussion
Possibly the oldest record of Saxidomus is Saxidomus? noblei Dickerson, 1914 from the Paleocene Martinez Formation in California (Dickerson, 1914). Sinelnikova (1997) recorded Saxidomus noblei from the Paleocene to lower Eocene Tkaprabayamskaya Formation in western Kamchatka. Macrocallista? andersoni Dickerson, 1915 from the middle Eocene Cowlitz Formation in California looks like a younger shell of S. nuttalli in its outline. It has a distinct lunule incision. However, as the length of this species is small (26 mm) it resembles young shells of Saxidomus, which sometimes have a lunular incision. Saxidomus? sp. indet. was illustrated from the middle Eocene Tainohata Formation in Hyogo Prefecture, southwest Honshu by Matsubara et al. (2010). Saxidomus vaquerosensis (Arnold, 1909) which is similar to S. nuttalli, first appeared from the upper Oligocene in California, USA (Weaver, 1969). Although the shell outline of this Paleogene species is similar to species of Saxidomus, it is difficult to confidently identify it as Saxidomus because there is no description of the hinge or inner structure.
Kamada (1962) described E. kurodae as a new species from the lower Oligocene Iwaki Formation in Fukushima Prefecture. Judging from the figures of the hinge illustrated by Yabe et al. (1995), the species can be safely allocated to Saxidomus for its large subumbonal pit, small anterior cardinal tooth (3a) touching the anterior lateral tooth (AI) in the right valve and large anterior lateral tooth (AII) in the left valve. Moreover, examination of the specimens illustrated by Nemoto & O’Hara (2001) show this species to lack a lunular incision. Thus, Saxidomus kurodae
(Kamada, 1962) is the oldest certain record of the genus Saxidomus in Japan. This age corresponds to the global cooling at the Eocene and Oligocene transition (Prothero, 1994).
In the late early to early middle Miocene, the marine climate around southwestern Hokkaido to Kyushu was tropical and subtropical (Chinzei, 1986; Ogasawara, 1994) and that of southwestern Sakhalin correspondingly temperate (Amano et al., 1996). Some species of Saxidomus have been recorded from tropical to temperate faunas. Nomura (1935) described S. gigantea [= Saxidomus? sp.] from the upper lower Miocene Tanosawa Formation in Aomori Prefecture, along with many warm-water species. Nomura (1940) described an inner mold of a venerid from the lower middle Miocene Moniwa Formation in Miyagi Prefecture as S. cf. nuttalli Conrad. Michinoku Amateur Paleontologists’ Club (1991) recorded S. cf. purpurata (Sowerby) in a nest of a rock-boring bivalves from the Moniwa Formation. Moreover, Shibata in Itoigawa et al. (1974) described Saxidomus sp. from the upper lower Miocene Yamanouchi Formation in Gifu Prefecture. Based on the observation of the hinge of the specimen whose outer surface only was illustrated by Shibata in Itoigawa et al. (1974) (MFM 11261), the specimen has three cardinal teeth, as in Saxidomus, and a large subumbonal pit. The outlines of the Moniwa and Yamanouchi species are similar to each other, and differ from S. purpurata by having a less inflated umbo and a horizontal postero-dorsal margin. Saxidomus sp. was illustrated from the upper lower Miocene Higashi-innai Formation in Ishikawa Prefecture by Matsuura (2009) and from the lower middle Miocene Ausi Formation in southwestern Sakhalin by Amano et al. (1996). Saxidomus sp. was described from the lower middle Miocene Aratani and Shimo Formations in Fukui Prefecture by Nakagawa (1998, 2018). Saxidomus kakertensis Sinelnikova in Gladenkov et al., 1984 was described from the middle Miocene Kakert Formation in western Kamchatka (Gladenkov et al., 1984). The outline of the latter species is similar to S. purpurata, but Sinelnikova did not describe the hinge and compared it with S. nuttalli and not with S. purpurata. Saxidomus kamchaticus Krishtofovich, 1969 and Saxidomus conspectus Krishtofovich, 1969 were described and illustrated from the Miocene Hot Spring (Goryachiv Klyuchei) Bed in the Kronotskiy Region of eastern Kamchatka (Krishtofovich, 1969). Most of them were internal molds and the hinge plates were not illustrated. A Saxidomus species identified as S. pupurata was described from the middle Miocene Omori Formation (Sakanoue & Itoigawa, 1997; Sakanoue, 1998, 2000). However, it is difficult to confidently identify as S. purpurata because of poor preservation and no illustration of the hinge.
As set out above, S. purpurata certainly first appeared in the lower upper Miocene Kubota Formation in Fukushima Prefecture and the Itahana Formation in Gunma Prefecture, whose fauna represents the warm- to mild-temperate Shiobara-type fauna (Chinzei, 1963; Chinzei & Iwasaki, 1967; Iwasaki, 1970). Saxidomus purpurata appeared and adapted to the warm- to mild-temperate water after the global cooling at 14.7 Ma and 13.9–13.8 Ma along with the stepwise expansion of Antarctic ice sheets (Holbourn et al., 2014).
The Tatsunokuchi fauna (Nomura, 1938; Otuka, 1941) represents an embayment with a shallow cold-water fauna (Chinzei, 1978; Chinzei, 1986) or cool-temperate fauna (Ogasawara, 1994) on the Pacific side in the latest Miocene to Pliocene. Judging from the lithofacies and associated species from the uppermost Miocene to lowermost Pliocene Tatsunokuchi Formation in Miyagi Prefecture described herein, it is plausible that the molluscs lived on a cold-water sand or gravelly sand bottom in the intertidal to 10 m deep in the embayment (Table 2). Today, S. purpurata occupies a wide marine climate range from the warm-temperate to subarctic zones classified by Nishimura (1981). Barinov (1992) illustrated only a fragment of a shell as Saxidomus sp. together with cold-water species from Bed 12 of the upper Pliocene Limimtevayamskaya Formation in Karaginsky Island near eastern Kamchatka, whose age was based mainly on diatom fossils. However, this specimen differs from S. purpurata in having a very wide hinge plate. Consequently, the occurrence of S. purpurata from the Tatsunokuchi Formation is the oldest certain record of the genus Saxidomus from the cool-temperate zone.
Acknowledgements
We thank Geerat J. Vermeij (University of California at Davis) for his critical reading of the manuscript and Takashi Matsubara (Hokkaido University of Education at Kushiro) for his plentiful information on the literature and his suggestions regarding Ezocallista kurodae. We also thank Yukito Kurihara (Mie University) and one anonymous reviewer for their critical reading and many useful suggestions. We are grateful to Takuma Haga and Atsushi Yabe, Kazunori Hasegawa, Hiroshi Saito (National Museum of Nature and Science), Toshiaki Irizuki (Shimane University), Yuji Takakuwa (Gunma Museum of Natural History), Yusuke Ando (Mizunami Fossil Museum) and Nagayuki Nemoto (Hironomachi Education Board, Fukushima) for their help in examining the fossil and Recent specimens for comparison of Saxidomus.
References
Akamatsu, M. 1984. On the so-called Shishinai fauna from the Ishikari Hills, Hokkaido. Annual Report of the Historical Museum of Hokkaido 12: 1–33. (in Japanese)
Amano, K. in press. Molluscan fossils from the Kitakanegasawa Formation in Aomori Prefecture, Japan, with remarks on the northern limit of the Tsushima Warm Current during the late Pliocene. Bulletin of the National Museum of Nature and Science, Series C (Geology & Paleontology) 46.
Amano, K. & Hikida, Y. 1999. Evolutionary history of the Cenozoic bivalve genus Kaneharaia (Veneridae). Paleontological Research 3: 249–258.
Amano, K. & Kiel, S. 2007. Fossil vesicomyid bivalves from the North Pacific region. The Veliger 49: 270– 293.
Amano, K. & Ogihara, R. 2012. Taxonomy of large Nuttallia (Bivalvia: Psammobiidae) in the northwestern Pacific, with remarks on the evolution of the genus. Paleontological Research 16: 146–158.
Amano, K., Khudik, V. D. & Narita, K. 1996. An early Middle Miocene warm-water molluscan fauna in southwestern Sakhalin −Molluscs from the Aushinskaya Formation around Novoselovo. Transactions and Proceedings of the Palaeontological Society of Japan (184): 651–661.
Amano, K., Miyajima, Y., Jenkins, R. G. & Kiel, S. 2019. The Miocene to Recent biogeographic history of vesicomyid bivalves in Japan, with two new records of the family. The Nautilus 133: 48–56.
Arkhipova, A. D., Brutman, N. Y., Zhidkova, L. S., Ivan’shina, L. P., Kafanov, A. I., Korobkov, A. I., Kuznetzova, V. N., Moiseeva, A. I., Popova, L. A., Pronina, I. G., Remizovsky, V. I., Runeva, N. P., Savizky, V. O., Sal’nikov, B. A., Utkina, A. I., Shainyan, S. Kh. & Schmidt, O. I. 1992 [1994]. Reference section of Paleogene Neogene deposits of South-East Sakhalin (Makarov section). 358 pp., 68 pls. All-Russia Petroleum Scientific-Research Geological-Exploration Institute, St.-Petersburg. (in All-Russian with English summary)
Baba, K. 1990. Molluscan Fossil Assemblages of the Kazusa Group, South Kwanto, Central Japan. 445 pp. Keio-Yochisha, Tokyo. (in Japanese with English description of new taxa)
Barinov, K. B. 1992. Miocene and Pliocene Mollusca. In: Gladenkov, Yu. B. & Devjatkin, E. V. (eds), Detailed Division of the Neogene of Kamchatka, pp. 27–36, pls 1–15. Transactions of the Geological Institute, 478, Nauka, Moscow. (in Russian)
Brauns, D. 1881. Geology of the environs of Tokio. Memoir of the Department of Sciences, Tokio Daigaku 4: 1–82, pls 1–8.
Chen, J., Li, Q., Kong, L. & Yu, H. 2011a. How DNA barcodes complement taxonomy and explore species diversity: The case study of a poorly understood marine fauna. PLoS ONE 6: e21326.
Chen, J., Li, Q., Kong, L. & Zheng, X. 2011b. Molecular phylogeny of venus clams (Mollusca, Bivalvia, Veneridae) with emphasis on the systematic position of taxa along the coast mainland China. Zoologica Scripta 40: 260–271.
Chinzei, K. 1963. Notes on historical change of Neogene molluscan assemblages in Northeast Japan. Fossils (Palaeontological Society of Japan) (5): 20–26. (in Japanese)
Chinzei, K. 1978. Neogene molluscan faunas in the Japanese islands: An ecologic and zoogeographic synthesis. The Veliger 21: 155–170.
Chinzei, K. 1986. Faunal succession and geographic distribution of Neogene molluscan faunas in Japan. Palaeontological Society of Japan, Special Papers (29): 17–32.
Chinzei, K. & Iwasaki, Y. 1967. Paleoecology of shallow sea molluscan faunae in the Neogene deposits of Northeast Honshu, Japan. Transactions and Proceedings of the Palaeontological Society of Japan (67): 93–113.
from Baja California to Northern Perú. 1258 pp. Santa Barbara Museum of Natural History, Santa Barbara.
Coan, E. V., Valentich-Scott, P. & Bernard, F. R. 2000. Bivalve Seashells of Western North America. 764 pp. Santa Barbara Museum of Natural History, Santa Barbara.
Dickerson, R. E. 1914. New molluscan species from the Martinez Eocene of southern California. University of California, Bulletin of the Department of Geology 8: 299–304, pl. 29.
Fujie, T. 1958. Fossil molluscs from the Pleistocene Shishinai Formation. 1. Fossil Pelecypoda. Cenozoic Research 28: 663–679. (in Japanese)
Fujiyama, I. 1982. Shells from the Quaternary. In: Fujiyama, I., Hamada, T. & Yamagiwa, N. (eds), Illustrated Encyclopedia of the Japanese Fossils for Students, pp. 338–362. Hokuryukan, Tokyo. (in Japanese, title translated herein)
Gladenkov, Yu. B., Gladikova, V. M., Kafanov A. I., Konova, L. N., Krishtofovich, L. V., Sinelnikova, V. N. & Popov, S. V. 1984. Marine molluscs. In: Menner, V. V. (ed.), Atlas of Fauna and Flora of the Neogene Sediments of the Far East. Tochilinski Section of Western Kamchatka, pp. 152–251, 307–324, pls 27–65. Nauka, Moscow. (in Russian)
Golikov, A. N. & Scarlato, O. A. 1967. Molluscs of the Possiet Bay (the Sea of Japan) and their ecology. Transactions of the Institute of Zoology, Academy of Sciences of USSR 42: 5–154. (in Russian) Habe, T. 1951. Genera of Japanese Shells. Pelecypoda. Vol. 2. 186 pp. Privately published, Kyoto. (in
Japanese)
Habe, T. 1977. Systematics of Mollusca in Japan: Bivalvia and Scaphopoda. 372 pp. Zukan-no-Hokuryukan, Tokyo. (in Japanese)
Habe, T. & Ito, K. 1965. Shells of the World in Colour. Vol. 1. 176 pp., 56 pls. Hoikusha, Osaka. (in Japanese) Habe, T. & Kosuge, S. 1967. The Standard Book of Japanese Shells in Color. Vol. 3. 223 pp., 64 pls. Hoikusha,
Osaka. (in Japanese)
Hatai, K. & Nisiyama, S. 1952. Checklist of Japanese Tertiary marine Mollusca. Science Reports of the Tohoku University, Second Series, Special Volume (3): 1–464.
Higo, S., Callomon, P. & Goto, Y. 1999. Catalogue and Bibliography of the Marine Shell-bearing Mollusca of Japan. 749 pp. Elle Scientific Publication, Yao.
Holbourn, A., Kuhnt, W., Lyle, M., Schneider, L., Romero, O. & Andersen, N. 2014. Middle Miocene climate cooling linked to intensification of eastern equatorial Pacific upwelling. Geology 42: 19–22.
Horikoshi, M. 1987. Distinctions in the hinge structure between Akebiconcha and Calyptogena, with special reference to “subumbonal pit” and “cardinal ligament”. Venus (Japanese Journal of Malacology) 45: 245–257.
Huber, M. 2010. Compendium of Bivalves. A Full-Color Guide to the World’s Marine Bivalves. A Status on Bivalvia after 250 Years of Research. 901 pp. Conchbooks, Hackenheim.
International Commission on Zoological Nomenclature. 1999. International Code of Zoological Nomenclature, Fourth Edition. 306 pp. The International Trust for Zoological Nomenclature, London.
Ishii, H. 1987. Late Pleistocene and Holocene bivalve mollusks of Osaka City and adjacent area. Special Publications from the Osaka Museum of Natural History 19: 1–36, pls 1–20. (in Japanese with English preface)
Itoigawa, J., Shibata, H. & Nishimoto, H. 1974. Molluscan fossils of the Mizunami Group. Bulletin of the Mizunami Fossil Museum (1): 43–203, pls 1–63. (in Japanese)
Iwai, T. 1965. The geological and paleontological studies in the marginal area of the Tsugaru Basin, Aomori Prefecture, Japan. Bulletin of the Faculty of Education, Hirosaki University 15: 1–68, pls 1–20. Iwai, T. & Shiobara, T. 1968. Pleistocene Mollusca from Kamikita-gun, Aomori Prefecture, Japan. Bulletin
of the Faculty of Education, Hirosaki University 20: 1–7, pls 1–3.
Iwasaki, Y. 1970. The Shiobara-type molluscan fauna. Journal of the Faculty of Science, University of Tokyo, Sec. 2 17: 351–444, pls 1–7.
Kamada, Y. 1962. Tertiary Mollusca from the Joban Coal-field, Japan. Palaeontological Society of Japan, Special Papers (8): 1–187, pls 1–21.
Kamada, Y. 1972. Tertiary Mollusca from the Joban coal-field. 1. Paleogene Iwaki Formation. Pg-5. Atlas of Japanese Fossils 19 (109): 2 maps + 2 figs + explanation. (in Japanese with English title)
Kaseno, Y. & Matsuura, N. 1965. Pliocene shells from the Omma Formation around Kanazawa City, Japan. Science Report of the Kanazawa University 10: 27–62, pls 1–20.
Keen, M. 1969. Superfamily Veneracea Rafinesque, 1815. In: Moore, R. C. (ed.), Treatise of Paleontology, Part N, Mollusca 6, Bivalvia, Volume 2, pp. N670–N690. The University of Kanzas, Lawrence. Kinoshita, T. & Isahaya, T. 1934. Catalogue of the shell bearing molluscs from Hokkaido, no. 1. Scientific
Japanese)
Kira, T. 1959. Coloured Illustrations of the Shells of Japan. Enlarged and Revised Edition. 239 pp. Hoikusha, Osaka. (in Japanese)
Kira, T. 1962. Shells of Western Pacific in Color, vol. I. 224 pp., 72 pls. Hoikusha, Osaka.
Krishtofovich, L. V. 1969. Molluscs of Miocene deposits of eastern Kamchatka. Trudy Vsesoiuznogo Nauchno-Issledovatel’skogo, Geologo-Razvedochnogo Instituta (VNIGRI): Seriya Paleontologiya 4: 172–211, pls 1–8. (in Russian, title translated herein)
Kurihara, Y. 2010. Middle and Late Miocene marine Bivalvia from the northern Kanto region, central Japan. National Museum of Nature and Science Monographs (41): 1–87.
Kuroda, T., Habe, T. & Oyama, K. 1971. The Sea Shells of Sagami Bay. 741 pp. (in Japanese) + 489 pp. (in English), 121 pls. Maruzen, Tokyo.
Kuwahara, Y. & Taru, M. 2012. Account of the 2011 summer meeting of the Society for the Study of Molluscan Diversity in Hokkaido Prefecture (the first part). Molluscan Diversity 3: 121–133. (in Japanese)
Lischke, D. E. 1871. Japanische Meers Conchylien. 184 pp. Theodor Fischer, Cassel.
Lutaenko, K. A. & Noseworthy, R. G. 2012. Catalogue of the Living Bivalvia of the Continental Coast of the Sea of Japan (East Sea). 247 pp. Dalnauka, Vladivostok.
Masuda, K., Hayasaka, S. & Noda, H. 1983. Shell bearing molluscs in Sendai Bay, northeast Honshu, Japan. Saito Ho-on Kai Museum of Natural History, Research Bulletin (51): 1–29, pls 1–5.
Matsubara, T. 2013. The valid author of Ezocallista (Bivalvia: Veneridae) revisited. Venus 71: 227–231. Matsubara, T., Kurita, H. & Matsuo, H. 2010. Eocene Mollusca from the Tainohata Formation in Kobe City,
Southwest Japan. Paleontological Research 14: 119–144.
Matsukuma, A. 2000. Family Veneridae. In: Okutani, T. (ed.), Marine Mollusks in Japan, pp. 1002–1019. Tokai University Press, Tokyo. (in Japanese and English).
Matsukuma, A. 2017. Family Veneridae. In: Okutani, T. (ed.), Marine Mollusks in Japan, The Second Edition, 559–561 (atlas), 1241–1250 (text). Tokai University Press, Hiratsuka. (in Japanese and English). Matsuura, N. 1977. Molluscan fossils from the Late Pleistocene marine terrace deposits of Hokuriku region,
Japan Sea side of Central Japan. Science Reports of Kanazawa University 22: 117–162.
Matsuura, N. 2009. Fossils of Ishikawa Prefecture (New Edition). 273 pp. Hokkoku Shimbun, Kanazawa. (in Japanese)
Michinoku Amateur Paleontologists’ Club. 1991. Fossil from the Moniwa Formation II. Fossil Shells. 32 pp. Michinoku Amateur Paleontologists’ Club, Sendai. (in Japanese, title translated)
Min, D. 2004. Mollusks in Korea. 566 pp. Min Molluscan Research Institute, Seoul. (in Korean)
Nagamori, H. 2000. Molluscan Fossils from the Togakushi Village 1. 41 pp. Togakushi Fossil Museum, Nagano. (in Japanese)
Nakagawa, T. 1998. Miocene molluscan fauna and paleoenvironment in the Niu Mountains, Fukui Prefecture, central Japan. Science Reports of the Institute of Geoscience, University of Tsukuba, Sec. B 19: 61–185. Nakagawa, T. 2018. Fossils from Oguroi, Takahama Town, Oi-gun, Fukui Prefecture. 78 pp. Fukui City
Museum of Natural History, Fukui. (in Japanese)
Nakao, K. 1993. Distribution of molluscan associations of the Pleistocene Toriyamahama Formation in the Miyazaki Plain. The Quaternary Research 32: 157–170. (in Japanese with English abstract)
Nakashima, R. 2002. Geographic distribution of the late Cenozoic bivalve Fortipecten in the northwestern Pacific. Palaeogeography, Palaeoclimatology, Palaeoecology 186: 261–274.
Nemoto, N. & Akimoto, Y. 1990. Marine molluscan fauna of Joban coast, Fukushima Prefecture, Pacific coast of northeastern Honshu. Research Report of Taira Geological Society (18): 3–50. (in Japanese with English abstract)
Nemoto, N. & O’Hara, S. 2001. Molluscan assemblages and sedimentary environments of the Shiramizu Group in northern area of the Joban district. Research Report of Taira Geological Society (23): 21–51. (in Japanese with English abstract)
Nemoto, N. & O’Hara, S. 2003. Molluscan fossils and sedimentary environments of the middle Miocene Kubota Formation in the Tanagura District. Research Report of Taira Geological Society (24): 3–32. (in Japanese with English abstract)
Nishimura, S. 1981. Sea and Life of the Earth: An Introduction to Marine Biogeography. 284 pp. Kaimei-sha, Tokyo. (in Japanese)
Noda, H. 1972. Geological significance of Saxidomus purpuratus (Sowerby) from the Pliocene Haneji Formation in Okinawa-jima. Professor J. Iwai Memorial Volume: 683–690, pl. 4.
Noda, H. & Masuda, K. 1968. On the Early Miyagian marine fauna from the Ojika Peninsula, Miyagi Prefecture, Japan. Saito Ho-on Kai Museum, Research Bulletin (37): 1–9, pl. 1.
Noda, H., Kikuchi, Y. & Nikaido, A. 1993. Molluscan fossils from the Pliocene Kume Formation in Ibaraki Prefecture, Northeastern Kanto, Japan. Science Reports of the Institute of Geoscience, the University of Tsukuba, Section B, Geological Science 14: 115–204.
Noda, H., Watanabe, R. & Kikuchi, Y. 1995. Pliocene marine molluscan fauna from Hitachi Formation in the northern part of Ibaraki Prefecture, Japan. Science Reports of the Institute of Geoscience, the University of Tsukuba, Section B, Geological Science 16: 39–93.
Noda, H., Kikuchi, Y. & Nikaido, A. 2004. 5. Molluscan fossils from the Pliocene Kume Formation in Kanasago-machi, Ibaraki Prefecture, Northern Kanto, Japan. Monographs of Ibaraki Nature Museum 1: 54–112. (in Japanese)
Nomura, S. 1935. Miocene Mollusca from Nishi-Tugaru District, Aomori-Ken, Northeast Honsyu, Japan. Saito Ho-on Kai Museum, Research Bulletin (6): 19–74, pls 2–8.
Nomura, S. 1938. Molluscan fossils from the Tatunokuti Shell Bed exposed at Goroku Cliff in the western border of Sendai. Science Reports of the Tohoku Imperial University, Second Series 19: 235–275, pls 33–36.
Nomura, S. 1940. Molluscan fauna of the Moniwa Shell Beds exposed along the Natori-Gawa in the vicinity of Sendai, Miyagi Prefecture. Science Reports of the Tohoku Imperial University, Second Series 21: 1–46, pls 1–3.
Nomura, S. & Hatai, K. 1936. Fossils from the Tanagura Beds in the vicinity of the Town Tanagura, Hukusima-Ken, Northeast Honsyu, Japan. Saito Ho-on Kai Museum of Natural History, Research Bulletin (10): 109–155, pls 13–17.
Nomura, S. & Hatai, K. 1939. On some fossils from Akita Prefecture, Northeast Honsyu, Japan. Japanese Journal of Geology and Geography 16: 39–64.
Ogasawara, K. 1994. Neogene paleogeography and marine climate of the Japanese Islands based on shallow-marine molluscs. Palaeogeography, Palaeoclimatology, Palaeoecology 108: 335–351.
Ogasawara, K., Masuda, K. & Matoba, Y. 1986. Neogene and Quaternary Molluscs from the Akita Oil-field, Japan. 310 pp. Commemorative Association of Professor Taisuke Takayasu’s Retirement and Supporters’ Foundation of Mineral Industry Museum, Mining College, Akita University, Akita. (in Japanese) Ohara, S. 1968. Geological Atlas of the Chiba Prefecture. No. 5, Bivalvia. 91 pp. Chiba-Ken Chigaku Kyôiku
Kenkyu-Kai [Research Group of Earth Science of Chiba Prefecture], Chiba. (in Japanese)
Okamoto, K. & Ibaraki, M. 1988. Early Pleistocene fossil Mollusca from the Sea of Japan off Yamaguchi Prefecture, Southwest Japan. Saito Ho-on Kai, Special Publication (2): 523–532, pls 1, 2.
Okutani, T., Tagawa, M. & Horikawa, H. 1989. Bivalves from Continental Shelf and Slope around Japan. The Intensive Research of Unexploited Fishery Resources on Continental Slopes. 190 pp. Japan Fisheries Resource Conservation Association, Tokyo. (in Japanese and English)
Otuka, Y. 1941. On the fauna of the Neogene between Honzyo and Kurosawajiri. Journal of the Japanese Association of Petroleum Technologists 9: 85–107. (in Japanese)
Oyama, K. 1973. Revision of Matajiro Yokoyama’s type Mollusca from the Tertiary and Quaternary of the Kanto Area. Palaeontological Society of Japan, Special Papers (17): 1–148, pls 1–57.
Ozaki, H. 1958. Stratigraphical and paleontological studies on the Neogene and Pleistocene formations of the Tyoshi district. Bulletin of the National Science Museum, New Series, 4: 1–182, pls 1–24.
Prothero, D. R. 1994. The Eocene-Oligocene Transition. Paradise Lost. 291 pp. Columbia University Press, New York.
Qi, Z. 2004. Family Veneridae. In: Qi, Z. (ed.), Seashells of China, pp. 297–315, pls 162–171. China Ocean Press, Beijing.
Qi, Z., Ma, X., Wang, Z., Lin, G., Xu, F., Dong, Z., Li, F. & Lu, R. 1989. Mollusca of Huanghai and Bohai. 309 pp., 13 pls. Agricultural Publishing House, Beijing. (in Chinese)
Sakagami, S., Takano, N., Sasaki, A., Nishikage, T., lehido, Y., Ozeki, S., Shinohara, H., Tanaka, R., Shimokawabe, H., Takahashi, Y., Takeshita, Y. & Hayashi, T. 1966. Fossils from the Tomikawa Formation of Kamiiso, Oshima Peninsula, Hokkaido. I. Molluscs etc. Journal of the Hokkaido University of Education, Series B 17: 78–93. (in Japanese).
Sakanoue, H. 1998. Commentary on the Sakanoue Molluscan Fossils Collection. Molluscan Fossils from the Miocene of Izumo. 62 pp. Shimane Prefectural Sanbe Open Field Museum, Ota City. (in Japanese) Sakanoue, H. 2000. Miocene molluscan fossils from the Omori Formation, Sugesawa, Izumo City, Southwest
Japan. Science Report of the Toyohashi Museum of Natural History (10): 13–21. (in Japanese with English abstract)
Sakanoue, H. & Itoigawa, J. 1997. Miocene molluscan fossils from Noshira, Matsue City, Southwest Honshu Japan. Science Report of the Toyohashi Museum of Natural History (7): 1–10. (in Japanese with English abstract)
Sato, T. & Matsushima, Y. 2000. Multivariate analysis of shell morphology of fossil Saxidomus purpurata (Bivalvia: Veneridae) and its paleobiological implications. Fossils (Palaeontological Society of Japan) 67: 19–31. (in Japanese with English abstract)
Sawada, Y. 1962. The geology and paleontology of the Setana and Kuromatsunai areas in Southwest Hokkaido. Memoirs of the Muroran Institute of Technology 4: 1–110, pls 1–8.
Scarlato, O. A. 1981. Bivalve molluscs from the middle latitude of the western part of the Pacific Ocean. Taxonomic Monograph of Fauna USSR 126: 1–479. (in Russian)
Shimamoto, M. 1986. Shell Microstructure of the Veneridae (Bivalvia) and its Phylogenetic Implications. Science Reports of the Tohoku University, Second Series 56: 1–39, pls 1–20.
Sinelnikova, V. N. 1997. Molluscan descriptions. In: Gladenkov, Yu. B. (ed.), Lower Paleogene of Western Kamchatka, pp. 250–301, pls 18–36. Publishing office GEOS, Moscow. (in Russian with English title). Sowerby, G. B. II. 1852. Monograph of the genus Tapes. Thesaurus Conchyliorum, or Monographs of the
Genera of Shells 2: 678–699, pls 145–150.
Takayasu, K. 1980. Fossils from Nange, Matsue City ̶ Molluscan fossils from various localities in Shimane Prefecture, Part 1 ̶. Memoirs of the Faculty of Science, Shimane University 14: 133–145, pls 1–6. Taki, I. & Oyama, K. 1954. Matajiro Yokoyama’s the Pliocene and later faunas from the Kwanto region in
Japan. Palaeontological Society of Japan, Special Papers (2): 1–68, pls 1–49.
Tsuchida, E. & Kurozumi, T. 1996. Fauna of marine mollusks of the sea around Otsuchi Bay, Iwate Prefecture. (6) Bivalvia-3. Otsuchi Marine Research Center Report (21): 1–31. (in Japanese)
Weaver, D. W. 1969. Geology of the North Channel Islands [California]. 200 pp. American Association of Petroleum Geologists and Society of Economic Paleontologists and Mineralogists, Pacific Sections, Special Publication.
Xu, F. & Zhang, S. 2008. An Illustrated Bivalvia Mollusca Fauna of China Seas. 336 pp. Science Publishing House, Beijing. (in Chinese)
Yabe, A., Ogasawara, K. & Uemura, K. 1995. Stratigraphic relationship between the Paleogene and Neogene rocks in the Tono Area of lwaki, Northeast Honshu, Japan. Memoirs of the National Science Museum, Tokyo (28): 31–46. (in Japanese with English abstract)
Yamamoto, G. & Habe, T. 1959. Fauna of shell-bearing mollusks in Mutsu Bay. Lamellibranchia (2). Bulletin of the Marine Biological Station of Asamushi (9): 85–122, pls 6–14.
Yamazaki, T. & Kashio, S. 2011. Molluscan Fauna of Akkeshi Marine Station Field Science Center for Northern Biosphere, Hokkaido University. Part 1. Intertidal Areas. 62 pp. Conchological Club of Northern Regions, Hokkaido University, Hakodate. (in Japanese)
Yokoyama, M. 1920. Fossils from Miura Peninsula and its immediate north. Journal of the College of Science, Imperial University of Tokyo 39: 1–193, pls 1–20.
Yokoyama, M. 1922. Fossils from upper Musashino of Kazusa and Shimosa. Journal of the College of Science, Imperial University of Tokyo 44: 1–200, pls 1–17.
Zhang, S., Zhang, J., Chen, Z. & Xu, F. 2016. Mollusks of the Yellow Sea and Bohai Sea. 421 pp. Science Publishing House, Beijing. (in Chinese)
Zhao, R., Cheng, J. & Zhao, D. 1982. Marine Molluscs of Dalian. 167 pp., 22 pls. China Ocean Publishing House, Beijing. (in Chinese, title translated herein)
北西太平洋のウチムラサキガイ属(二枚貝綱,マルスダレガイ科)の 進化と冷水域適応 天野和孝・根本 潤 要 約 マルスダレガイ科の Saxidomus ウチムラサキガイ属に知られてきた殻頂下洞を Ezocallista エゾワスレガ イ属にも確認した。この発見は両属が Callista マツヤマワスレガイ属よりも近縁であることを支持してい る。現生の Saxidomus purpurata ウチムラサキは日本周辺では温帯水域に生息しているが,北海道礼文島, 厚岸湾,中国の渤海,ロシアのポシェット湾,アムール湾,ピーター大帝湾などの冷水域にも生息してい る。本種の冷温帯水域からの最古の記録は宮城県仙台市の最上部中新統∼最下部鮮新統竜の口層からの化 石である。このことは,ウチムラサキは中新世末期∼鮮新世初期に冷水域に適応進化したことを示唆して いる。


![Fig. 5. Fossil shells of Saxidomus. A–D, F–H. Saxidomus purpurata (Sowerby) from the Tatsunokuchi Formation at Goroku misidentified as Macrocallista brevisiphonata (Carpenter) by Nomura (1938); SM 2277 [= NMNS PM 18148]; A, hinge of left valve; B, hin](https://thumb-ap.123doks.com/thumbv2/123deta/6835068.1168781/10.773.110.666.199.889/saxidomus-saxidomus-tatsunokuchi-formation-misidentified-macrocallista-brevisiphonata-carpenter.webp)
