新規 B 型肝炎治療薬の創出を目的とした
レポーターHBV 感染系の構築と化合物スクリーニング
2018
原田圭輔
Establishment of Novel Hepatitis B Virus Reporter System for High-throughput Chemical Compound Screening
2018
Keisuke Harada
新規 B型肝炎治療薬の創出を目的とした
レポーターHBV 感染系の構築と化合物スクリーニング
原田 圭輔
Hepatitis B Virus (HBV)感染によって生じるB型肝炎は,肝臓が線維化し肝 硬変・肝がんへと進行する慢性疾患である。現在は B 型肝炎治療薬としてペグ 化インターフェロン (pegylated interferon, Peg-IFN) 又は核酸アナログが用い られるが,共に体内からの HBV 排除効率は低率であり,新たな作用機序の抗 HBV薬の開発が望まれている。新規抗ウイルス薬の開発及びその標的探索には,
ウイルスの in vitro 培養系と,迅速かつ簡便に化合物等をスクリーニング可能 な high-throughput screening (HTS)評価系の構築が必須である。2012 年に HBV の侵入受容体として sodium taurocholate cotransporting polypeptide (NTCP)が同定されて以降,培養細胞を用いたHBVのin vitro感染系の構築は 急速に発展した。一方,一般的なHBV複製評価法は依然としてReal-time PCR
法やELISAといった煩雑で時間がかかる手法であり,効率的に化合物ライブラ
リーをスクリーニングする新規評価系の構築は急務である。そこで本研究では,
新規抗 HBV薬の開発に向けて,HBV 複製を迅速かつ簡便に評価可能なレポー ターHBV感染系の構築に取り組んだ。まず,HBVゲノムにNanoLuc (NL)遺伝 子を挿入する事で,レポーターウイルスであるHBV/NLを作製し,そのライフ サイクルと既存 HBV 阻害薬への感受性を評価した。次に,HBV/NL 感染系を 用いて化合物ライブラリーのスクリーニングを実施し,HBV複製を阻害する化 合物を同定した。最後に,HBV 複製阻害化合物として同定した KX2-391 の作 用機序解析を実施した。
1. レポーターHBVの作製とそのライフサイクルの検討
HBVゲノムのcore及びpol領域にNLを挿入したHBV/NLは,野生型HBV と同様に肝細胞に感染し,HBV RNA 量と相関して NL を発現した。HBV/NL 感染系における既知の HBV 侵入及び転写阻害剤処置がレポーター活性を低下 させる事,HBV/NL 感染においても HBV cccDNA 形成が確認された事から,
HBV/NL感染系はHBVの侵入~cccDNA形成~HBV RNA転写を迅速・簡便に 評価可能であることが示された。
2. HBV/NLを用いたスクリーニング系の構築とその実施
HBV/NL感染系を用いた化合物のHTSを実施するために,HBV高感受性細
胞株の樹立を試みた。NTCP強制発現HepG2細胞の限界希釈法によるクローニ ングとHBV/NLによる選抜の結果,HBV感受性の高いHepG2/NTCP#22を取 得した。HepG2/NTCP#22及びprimary human hepatocyte (PHH)であるPXB 細胞のHBV/NL感染系を用いて化合物ライブラリー (1,827化合物)のスクリー ニングを実施した結果,細胞毒性のない濃度域でHBV複製を阻害する化合物と してKX2-391を同定した。
3. KX2-391の抗HBV作用メカニズム解析
Time of addition assay及びpromoter assayの結果から,KX2-391はHBV RNA の転写を阻害することが明らかになった。また,KX2-391 は Src 阻害と tubulin polymerization 阻害の 2 つの作用を持つ dual inhibitor であるが,
siRNA 及び阻害剤処置の結果から,tubulin polymerization 阻害が HBV 複製 阻害に重要であることが判明した。さらに,KX2-391 及びその他の tubulin polymerization 阻害剤は,肝臓特異的転写因子である HNF4A を mRNA レベ ルで減少させることによって,HBV RNA 転写を阻害することが明らかになっ た。
本研究において構築した HBV/NL 感染系は,HBV 複製の侵入から転写まで をレポーター活性を指標に迅速・簡便に評価可能であることが示された。化合物
及びsiRNA等のスクリーニングも可能であることから,HBV/NL感染系は今後
の新規抗 HBV 薬及びその標的分子の探索への寄与が大きく期待される。また,
本研究において同定したKX2-391 は,これまでに報告のない様式でHBV転写 を特異的に阻害した。KX2-391 の作用機序のさらに詳細な解析は,副作用の少 ない新規抗HBV薬の開発に繋がりうる。
Establishment of Novel Hepatitis B Virus Reporter System for High-throughput Chemical Compound Screening
Keisuke Harada
Hepatitis B virus (HBV) is a leading cause of liver disease including liver fibrosis, cirrhosis and hepatocellular carcinoma. Current therapies are limited to pegylated interferon alpha (PEG-IFN) and/or nucleos(t)ide analogs, however, these are only partially effective. Therefore, the development of anti- HBV drugs that target the different steps of virus replication is needed to eradicate HBV. In general, the establishment of High-throughput screening (HTS) system using a simple in vitro culture of the target virus facilitates the development of anti-virus agents. Recently, the identification of sodium taurocholate cotransporting polypeptide (NTCP) as a functional receptor for HBV has allowed the development of a variable HBV culture system. However, studies of virus infection/replication at the molecular level using wild-type HBV are labor-intensive and time-consuming. To overcome these problems, we constructed a novel recombinant HBV encoding NanoLuc (NL) as a reporter gene to monitor the infection/replication of the HBV. In this study, we showed that reporter-HBV (HBV/NL) is highly sensitive for monitoring the early stages of the HBV replication cycle. In addition, screening of anti- HBV agents using HBV/NL identified an anti-HBV compound that suppresses transcription from the HBV precore promoter.
1. Construction of HBV/NL plasmids and characterization of recombinant viruses
We constructed HBV/NL bearing the NL gene, which replaced with a portion of core and pol ORF. NL activity of HBV/NL in virus-infected cells represents the amount of cellular HBV RNA. In addition, well-known HBV inhibitors, such as human hepatitis B immunoglobulin (HBIG) or interferon β, inhibited the expression of NL in infected cells. Furthermore, covalently closed circular DNA (cccDNA) was detected in HBV/NL infected cells. These results suggest that HBV/NL can be used to monitor HBV replication between the virus entry and transcription steps by detecting NL activity.
2. High-throughput screening of anti-HBV drugs using recombinant HBV/NL A stable HepG2 cell line expressing sodium taurocholate cotransporting polypeptide (NTCP) was established by end-point dilution. The HepG2/NTCP#22 was highly susceptible to HBV infection. To identify anti- HBV drugs, we screened a library of 1,827 US Food and Drug Administration approved compounds using HBV/NL infection system, and KX2-391, a known SRC kinase and tubulin polymerization inhibitor, was identified as a new candidate HBV inhibitor.
3. The mechanism of action analysis of anti-HBV agent, KX2-391
Time of addition assay and promoter assay revealed that KX2-391 suppressed HBV RNA transcription HBV DNA. In addition, not SRC inhibition but tubulin polymerization inhibition was linked to the anti-HBV activity of KX2-391. Furthermore, KX2-391 and other tubulin polymerization inhibitors decreased HNF4A, that affects HBV transcription, at mRNA level.
These results suggest that the inhibition of tubulin polymerization by KX2- 391 suppressed HBV by inhibiting the transcription of the gene encoding a precore protein.
In conclusion, we successfully produced a recombinant HBV expressing NL for evaluation of the HBV early life cycle, from entry to transcription. We also showed the utility of this system by screening of chemical agents that affect HBV infection and replication. Furthermore, the results of the present study suggest that tubulin polymerization inhibitors such as KX2-391 use a novel mechanism to suppress HBV transcription. Further study on the antiviral mechanism of tubulin inhibitors may aid the development of antiviral therapies.
略語表 本論文中に用いた略語は以下の通りである。
ADV Adefovir
ARP-1 Actin-related protein 1
Av Average
CA Capsid
cccDNA Covalently closed circular DNA CC50 50% cytotoxic concentration COUP-TF1 COUP Transcription Factor 1 CsCl Cesium chloride
C/EBP CCAAT-enhancer-binding protein DMEM Dulbecco's modified Eagle's medium EC50 50% effective concentration
EGF Epidermal growth factor
ELISA Enzyme-Linked ImmunoSorbent Assay
ETV Entecavir
FBS Fetal bovine serum
FDA US Food and Drug Administration HBV Hepatitis B Virus
HBIG Human hepatitis B immunoglobulin HCV Hepatitis C Virus
HIV-1 Human immunodeficiency virus type-1 HNF1 Hepatocyte Nuclear Factor 1
HNF3 Hepatocyte Nuclear Factor 3
HNF4A Hepatocyte Nuclear Factor 4 alpha HTS High-throughput screening
LAM Lamivudine
MRP1 Multidrug resistance protein 1
NA Nuculeoside analogue
NL NanoLuc
NTCP Sodium taurocholate cotransporting polypeptide PEG Polyethylene glycol
Peg-IFN Pegylated interferon
pgRNA Pre genome RNA
PHH Primary human hepatocyte PLB Passive Lysis Buffer
pol Polymerase
PPAR Peroxisome proliferator-activated receptor PXR Pregnane X Receptor
rcDNA Relaxed circular DNA RXR Retinoid X receptor
SCID Severe combined immunodeficiency
SD Standard deviation
sgRNA Subgenomic RNA
TAF Tenofovir alafenamide
TDF Tenofovir disoproxil fumarate
Tet Tetracycline
U Unit
本論文は、学術雑誌に掲載された以下の論文を基礎とするものである。
(1) Novel reporter system to monitor early stages of the hepatitis B virus life cycle.
Cancer Science, 106, 1616–1624 (2015).
Hironori Nishitsuji, Saneyuki Ujino, Yuko Shimizu, Keisuke Harada, Jing Zhang, Masaya Sugiyama, Masashi Mizokami, Kunitada Shimotohno
(2) Identification of KX2-391 as an inhibitor of HBV transcription by a recombinant HBV-based screening assay
Antiviral Research, 144, 138–146 (2017).
Keisuke Harada, Hironori Nishitsuji, Saneyuki Ujino, Kunitada Shimotohno
(3) Investigating the hepatitis B virus life cycle using engineered reporter hepatitis B viruses
Cancer Science, 109, 241–249 (2018).
Hironori Nishitsuji*, Keisuke Harada*, Saneyuki Ujino*, Jing Zhang, Michinori Kohara, Masaya Sugiyama, Masashi Mizokami,
Kunitada Shimotohno
*These three authors contributed equally to this work.
目次
第1章 緒論 ……….
第2章 レポーターHBVの作製とそのライフサイクルの検討 .………..
第1節 レポーターHBVの構築 ……….………..
第2節 HBV/NLライフサイクルの検討 ………..………..
第3節 考察 ………..
第3章 HBV/NLを用いたスクリーニング系の構築とその実施 ……….
第1節 HBV高感受性NTCP強制発現HepG2細胞株の樹立 ……...
第2節 HBV/NLを用いた化合物ライブラリーの
スクリーニング ………..
第3節 KX2-391の抗HBV活性 ………..
第4節 考察 ………..
第4章 KX2-391の抗HBV作用メカニズム解析 ……….…....
第1節 KX2-391が標的とするHBV複製機序の同定 ………..
第2節 KX2-391によるHBV RNA転写阻害のメカニズム解析 ……
第3節 Tubulin polymerization阻害の
HBV promoter活性への影響 ………..
第4節 考察 ………..
第5章 総括 ………..
第6章 実験の部 ………..
参考文献 ……….….
1
6 7 10 13
15 16
17 19 21
24 25 27
29 32 35 37 49
第1章 緒論
肝細胞の炎症によって生じる肝炎は,急性肝炎と慢性肝炎に大別される。急性 肝炎は一部の重症化・劇症化・慢性化の症例を除いて 1 か月程度で治癒する疾 患であり,予後は良好である。一方,慢性肝炎は肝細胞の持続的な炎症によって 肝臓が線維化し,肝硬変・肝がんへと進行する疾患である。慢性肝炎の約 90%
は,C 型肝炎ウ イルス (Hepatitis C Virus, HCV)又は B 型肝炎ウイルス (Hepatitis B Virus, HBV)といった肝炎ウイルスの感染が原因である。B型肝炎 の原因ウイルスである HBV は,全世界で約 4 億人,日本でも約130 万人が感 染状態にあると報告されている (Fig. 1) 1)。HBV は血液接触によって伝播する が,感染時期によって経過が異なる。乳幼児期に母親から垂直感染した場合には,
約90%が無症候性キャリアとなり,10%が慢性肝炎へと進行する2, 3)。日本では 1985年にB 型肝炎母子感染防止事業が開始され,妊婦のHBV抗体検査と陽性 者へのワクチン投与が実施されたことから,母子感染による新規感染は著しく 減少した4)。一方,成人期に性交渉・注射器の回し打ちによって水平感染した場
合は,約80%は不顕性の一過性感染の後自然治癒し,残り20%が急性肝炎を発
症する。日本においては,B型急性肝炎からの慢性肝炎移行率は1%程度であっ たが,近年慢性化しやすい欧米型HBV (10%が慢性化)の感染が増加傾向となっ ており,水平感染でのB型慢性肝炎の流行が危惧されている5)。
Figure 1. Geographical distribution of major hepatitis B virus genotypes and worldwide frequency of chronic hepatitis B virus infection. HBVはA~Jの10のgenotypeに分類される。Genotype Aは北アメ リカ,ヨーロッパ北部及びアフリカに,genotype B及びCはアジアに,genotype Dは地中海地方,中東 及びインドに多くみられる。図中では,HBV陽性者が>8%以上の国をHigh, 2~7%の国をModerate, <1%
の国をLowと分類した。
Lancet 2014; 384: 2053–63
現在の慢性HBV感染治療は,ペグ化インターフェロン (pegylated interferon, Peg-IFN)又は核酸アナログの投与が基本である6)。Peg-IFNはHBV DNA抑制 作用と免疫賦活作用を持ち7, 8),約10%の患者においてHBVが体内から排除さ れドラッグフリーの状態となるが9),一方,60%以上の患者では治療効果が認め られない。また,インフルエンザ様の副作用や,Peg-IFN 注射のための週 1回 の通院等のデメリットも大きい。核酸アナログは,HBVの逆転写酵素を阻害す る薬剤で,現在日本ではlamivudine (LAM), adefovir (ADV), entecavir (ETV), tenofovir disoproxil fumarate (TDF)及びtenofovir alafenamide (TAF)の5剤 が承認されている10-12)。核酸アナログはHBVの複製を強力に阻害するが,持続 感染したHBVを排除することができず,投薬中止時には高率で肝炎が再発する ため,生涯の服用が必要となる13, 14)。近年,同様に持続感染が問題視されてい たHCV慢性肝炎において,異なるメカニズムの抗HCV薬を併用することで完 治が達成された 15)。新規メカニズムの抗 HBV 薬の開発は,多剤併用療法を発 展させ,HBVの根治に繋がりうるとして期待されている。
HBVのライフサイクル及び核酸アナログの作用点をFig. 2に示した16-18)。不 完全二重鎖 DNA (relaxed circular DNA, rcDNA)を遺伝情報として持つ HBV は,envelope protein (HBs)を介して肝細胞表面に結合する。その後エンドサイ トーシスを介して細胞質に侵入し,カプシドの崩壊 (脱殻)を経て核内へ移行す る。核へ移行したrcDNAは,宿主のDNA修復機構を利用して完全二重鎖構造 のcovalently closed circular DNA (cccDNA)へ変換される。cccDNAはヒスト ン蛋白質と結合してmini-chromosome構造を形成し,肝細胞中で非常に安定化 することから,HBV複製及び持続感染の起点となる19)。cccDNAを鋳型にして pre genome RNA (pgRNA)及びsubgenomic RNA (sgRNA, mRNAs)が転写さ れ,pgRNAからpolymerase (pol)及びcapsid protein (core, HBc)が,sgRNA からHBs及びx遺伝子蛋白質 (HBx)がそれぞれ翻訳される。pgRNAはpol及 び HBcと結合した後にカプシド化され,カプシド中で rcDNAへと逆転写され る。rcDNAを内包したnucleocapsidは,小胞体-ゴルジ体領域においてHBsと アセンブリーし細胞外へ放出されるか,再度核内移行して cccDNA を再形成す る (cccDNA amplification)。核酸アナログ (nuculeoside analogue, NA)は逆転 写過程を阻害しHBV複製を抑制するが,cccDNA減少作用は少ない13)。核酸ア ナログと併用可能な,新たな作用点を持つ抗HBV薬の開発が望まれている。
Figure 2. HBV life cycle and nucleoside analogues targets.
新規抗ウイルス薬の開発及びその標的探索には,ウイルスの in vitro 培養系 と , 迅 速 か つ 簡 便 に 化 合 物 等 を ス ク リ ー ニ ン グ 可 能 な high-throughput screening (HTS)評価系の構築が必須である。一方,HBVは感染を許容する宿主 細胞の範囲が狭く,侵入に用いる受容体も同定されていなかったため,近年まで in vitro培養系での感染/複製の評価が困難であった。HBV感染を許容する細胞 株として,human hepatoma cell lineのHepaRGが使用されてきたが20, 21),
HepaRG はその分化状態が HBV 複製に影響することから,一貫性や再現性に
乏しい。また,ヒト肝細胞 (primary human hepatocyte, PHH)を移植した urokinase-type plasminogen activator transgenic/SCID マウスから単離した PXB 細胞もHBV感染実験に使用可能であるが22),高額かつ供給量の問題から 使用に制限がかかる。Tetracycline 添加に よって HBV 発現が誘導される HepAD38 細胞も HBV 複製評価には有用であるが 23),侵入から cccDNA 形成 までの過程を評価できず用途が限定される。2012 年にHBV 侵入受容体として sodium taurocholate cotransporting polypeptide (NTCP)が同定されて以降24), 様々な HBV in vitro 培養系が急速に発展した。HBV感染非許容細胞であった HuH7細胞やHepG2細胞等のhepatocarcinoma cell lineへのNTCP強制発現 は,HBV感染及び複製を可能にし24),実験系の幅が大きく広がった。
一方,HBV複製の迅速かつ簡便な評価系の構築は課題として残る。一般的な HBV 複製の評価法は,HBV DNA 又はRNAの Real-time PCR法による定量
や,ウイルス蛋白質のELISAによる検出であり,大規模な評価を実施するには 非常に煩雑で時間がかかる手法である。効率的に化合物ライブラリーをスクリ ーニングする新規評価系の構築は急務である。
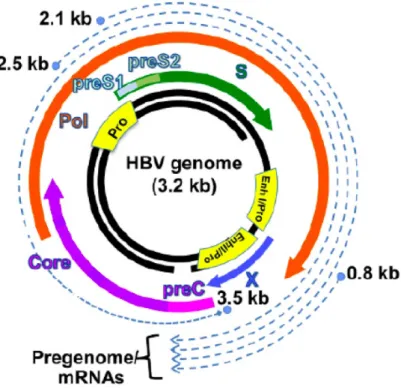
ウイルスの感染/複製評価には,ウイルスゲノムへの外来遺伝子の挿入が有効 である。Human immunodeficiency virus type-1 (HIV-1)やHCVは複製能を維 持したままレポーター又はマーカー遺伝子を挿入し,レポーター活性で複製を 評価することが可能であった 25-27)。一方,HBV はゲノムサイズが3.2 kb と小 さく,カプシド中へパッケージングされる長さにも制限がある 28)。また,ゲノ ム中のウイルス蛋白質をコードした領域がオーバーラップし,さらに,ウイルス 複製に重要なcis-elementが複数存在することから (Fig. 3),ゲノムへの外来遺 伝子の挿入が困難であった。HBV ゲノムへの外来遺伝子 (HIV-1 Tat, green fluorescent protein, DsRed 等)の挿入は複数報告されているものの28-30),既存 のレポーターHBVはウイルス産生の効率が悪く,またレポーター活性も低いた め,HTSを実施するには十分でない。
Figure 3. Schematic of the HBV genome and relative location of virus RNA and proteins on the genome.
HBVゲノムはcore promoter, S1 promoter, S2 promoter, X promoterの4つのpromoterを持つ。Core promoterから3.5 kbのpre genome RNA,S1及びS2 promoterからは2.1~2.5 kbのHBs mRNA,X promoter からは 0.8 kb の HBx mRNA が転写される。Enhancer I と Enhancer II はそれぞれ core promoter及びx promoterの活性を制御する。
本研究では,新規抗 HBV薬の開発に向けて,HBV複製を迅速かつ簡便に評 価可能なレポーターHBV 感染系の構築に取り組んだ。HBV ゲノムへ外来遺伝 子を挿入する際のサイズ制限,及びレポーター活性の高さを考慮して,レポータ ーとしてNanoLucを選抜し,レポーターHBV (HBV/NL)を作製した。まず,第 2章においては,HBVゲノムへのNanoLuc遺伝子の挿入と,作製したHBV/NL のライフサイクル及び既存HBV阻害薬への感受性を評価した。次に,第3章に おいて,HBV/NL 感染系を用いて化合物ライブラリーのスクリーニングを実施 し,HBV複製を阻害する化合物を選抜した。そして第4章では,化合物スクリ ーニングで見出したHBV複製を阻害する化合物について,作用メカニズム解析 を実施し,その抗HBV作用を明らかにした。
第2章
レポーターHBVの作製とそのライフサイクルの検討
レポーター遺伝子を挿入したウイルスを用いて感染/複製評価を実施するため には,高力価ウイルスの取得と高いレポーター活性が必要である。一方,外来遺 伝子を HBVゲノムに挿入する際に,ゲノムサイズが 3.7 kbを超えるとカプシ ドへのパッケージに不具合が生じ,ウイルス産生が極端に低下することが知ら れている28)。したがって,サイズが171アミノ酸と非常に小さく,firefly又は renilla luciferase よりレポーター活性が約 150 倍高い NanoLuc (NL)を 31), HBVゲノムへ挿入する遺伝子として選抜した。
本章では,HBVゲノムのcore及びpol領域を欠損させ,NL遺伝子を挿入し,
レポーターHBV プラスミドを作製した。作製したレポーターHBV プラスミド と,欠損したcore及び polを補完するためのヘルパープラスミドを細胞にコト ランスフェクションすることで,レポーターHBVを産生させた。産生したレポ ーターHBVを用いて,ライフサイクル及びHBV阻害剤感受性を評価し,HBV 複製評価に使用可能であるかを検討した。また,レポーターHBVゲノムサイズ が産生ウイルス量に与える影響を検討した。
第1節 レポーターHBVの構築
野生型 HBVゲノムプラスミド (pHBV)のcore及び pol領域の一部 (core の 192 塩基以降と pol の 287 塩基以前)を欠損させ,NanoLuc (NL)遺伝子を挿入 し,レポーターHBVゲノムプラスミド (pHBV/NL)を構築した (Fig. 4a)。pHBV 及びpHBV/NLのHBVゲノム長はそれぞれ3.2 kb, 3.3 kbである。各種HBV ゲノムプラスミドをHepG2細胞にトランスフェクションし,ウイルスRNAの 発現を確認した。HBVゲノムからは,3.5 kbのpgRNA,2.1-2.4 kbのHB RNA 及び0.8 kbのX gene RNAが転写される (Fig. 3)。pHBVを発現させた細胞は,
想定通りに3.5 kbのpgRNA及び2.1-2.4 kbのHBs RNAを発現した (Fig. 4b)。
露光時間を延長することで,0.8 kbのX gene RNAの発現も確認された (data not shown)。pHBV/NLを発現させた細胞も同サイズの3種のRNAを発現した ことから (Fig. 4b),pHBV/NL からの RNA転写が野生型と同様であることが 示された。
次に,pHBV/NL を用いたレポーターHBV (HBV/NL)の産生を実施した。
pHBV/NL はcore 及びpol を欠損するため,カプシド形成及び逆転写が不能で ある。よって,欠損した core及び polの発現を補完するために,pHBV/NL と ヘルパーHBVゲノムプラスミド (pHBV-D)の共発現を検討した。pHBV-Dは全 てのHBV蛋白質を発現するが,encapsidation signalに変異導入 (CTGTGCC to CTATGTC)されているため,pgRNA がカプシドへ封入されない。したがっ て,pHBV/NLとpHBV-Dの共発現で産生したHBV粒子中には,pHBV/NLが 選択的に取り込まれる。HepG2 細胞に pHBV/NL と pHBV をコトランスフェ クションすることで,HBV/NL を産生させた (Fig. 4c)。上清中に産生した
HBV/NL の粒子性状を,陽性対照である野生型 HBV と比較検討した。野生型
HBVは,pHBVをHepG2細胞にトランスフェクションすることで取得した。
トランスフェクションから 4 日後の培養上清を回収し,polyethylene glycol
(PEG)を用いて濃縮した後,超遠心によって精製した。精製した上清をCsCl密
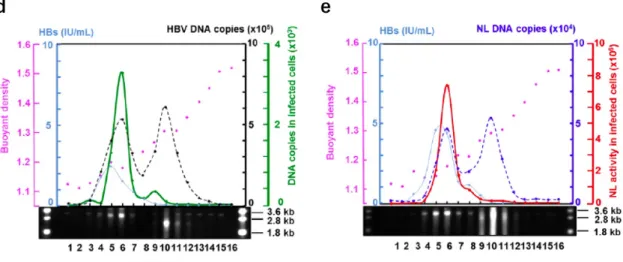
度勾配遠心によって分離し,各フラクションの性状解析を実施した (Figs. 4d and 4e)。各フラクションをSouthern blot法で解析すると,フラクション 5-7 及びフラクション9-11にHBV DNAが検出された。また,ELISAによってHBs 抗原を測定したところ,フラクション 5-7 に HBs 抗原が検出された。さらに,
各フラクションをNTCP強制発現HepG2細胞 (HepG2/NTCP)に感染させ,感 染細胞中の HBV DNA 量及びNL活性をそれぞれ定量PCR 及びルシフェラー ゼアッセイで測定したところ,フラクション 6を感染させた細胞で HBV DNA (野生型HBV)又はNL活性 (HBV/NL)がそれぞれ検出された。したがって,フ ラクション6が感染性HBV/NLであり,感染細胞中でNLを発現することが示
唆された。また,野生型HBV とHBV/NLは同等の粒子密度を持つことが確認 された。陰性対象としてpHBV-Dをトランスフェクションした上清を解析した ところ,HBs抗原は検出されたものの,HBV DNAは検出されず,pHBV-D単 独では HBV 粒子が産生しないことが示された (Fig. 4f)。続いて,レポーター HBV ゲノムサイズが産生ウイルス量に与える影響を検討した。pHBV の core 領域を欠損させ,NL 遺伝子を挿入することで pHVB/NL+pol を作製した。
pHBV/NLとpHBV/NL+polのHBVゲノム長はそれぞれ3.3 kb, 3.7 kbである (Fig. 4g)。pHBV/NL 及びpHBV/NL+pol をそれぞれpHBV-DとHepG2細胞 にコトランスフェクションすることで,HBV/NL 及び HBV/NL+pol を調製し た。HBV/NL及びHBV/NL+polをHepG2/NTCP細胞に感染させ,感染細胞中 のHBV RNA及びNL活性を定量したところ,HBV/NL+polはHBV/NLの30%
程度の感染性であった (Figs. 4h and 4i)。これらの結果より,HBV粒子産生に はHBVゲノム長のサイズ制限があり,ゲノム長の延長はHBV粒子産生を低下 させることが確認された。
Figure 4. Construction of HBV/NL plasmids and characterization of recombinant viruses. (a) Pregenomic structure of the reporter hepatitis B virus (HBV). Pregenomes of wild-type and reporter HBV, and putative ORFs of the virus are shown. The indicated sizes (kb) are of the pregenomes. A
stretch of “A”s indicates a poly A tail of the putative pre genome RNA. A lariat rope with “E” indicates an encapsidation signal and “X” on that indicates defect of encapsidation. The NL gene is inserted into the genome so as to be translated from its own initiator methionine. (b) Comparative analysis of virus RNA in transfected cells. Six days after transfection, 30 µg RNA from wild pHBV, pHBV/NL and pHBV/NL+pol transfected cells was analyzed by northern blot using digoxigenin (DIG) HBV RNA probes covering the full-length HBV genome. The gel was stained with ethidium bromide to visualize ribosomal RNA. The efficiency of transfection was similar (data not shown). (c) The reporter plasmid cannot produce PreCore and Pol, and the helper plasmid ontaining 2 mutations in the encapsidation signal (CTGTGCC to CTATGTC) expresses all HBV proteins. E: Encapsidation signal. (d, e and f) Cesium chloride (CsCl) density equilibrated centrifugation of wild-type hepatitis B virus (HBV) and recombinant HBV expressing NanoLuc (HBV ⁄ NL). Supernatants of HepG2 cells transfected with pHBV (d), pHBV ⁄ NL plus pHBV-D (e), and pHBV-D (f) were centrifuged at 150,000 g for 50 hr in a CsCl gradient. An aliquot from the top of the centrifuged tube was collected. Hepatitis B surface antigen (HBsAg; blue) in 10% of each aliquot was quantified by automated ELISA. The amount of NL DNA (dotted blue line) (e) and HBV DNA (dotted black line) (d, f) in 10% of each fraction was measured by quantitative PCR using a primer set for the NL gene and HBV DNA, respectively. Infectivity of 25% of each fraction in d and e was assayed by infecting 2×105 HepG2 ⁄ NTCP cells and measuring HBV DNA by quantitative PCR for wild HBV (green) (d) and NL activity for HBV ⁄ NL (red) (e) of 10% of the total cell lysate. Pink dots denote buoyant density. Southern blot analysis of each fraction was carried out by probing with digoxigenin-labeled HBV DNA. (g,h and i) Comparative analysis of the amount of viruses produced from transfected cells. Pregenomic structure of the HBV/NL and HBV/NL+pol (g). Six days after transfection by wild pHBV, pHBV/NL and pHBV/NL+pol, virus fractions from culture medium were harvested. One-tenth of the volume of culture medium was used to infect HepG2/NTCP. The amount of virus RNA and NL activity in cell lysates in (h) and (i), respectively, were measured after 6 days of infection. HBV, hepatitis B virus; NL, NanoLuc.
第2節 HBV/NLライフサイクルの検討
HBV/NL は野生型 HBV と同等の粒子密度を持つ感染性粒子であり,感染細
胞で NLを発現することが確認されたが,HBV/NLをスクリーニングで用いる ためには感染後の動態を確認する必要がある。したがって,野生型 HBV と
HBV/NLの感染後のライフサイクルを比較検討した。HepG2細胞へのトランス
フェクションで産生した野生型HBV又はHBV/NLを濃縮・精製し,HBV感染 を受容する PXB 細胞 22)に感染させた。感染細胞を経日的に回収し,定量 PCR でHBV RNA及びHBV DNA量を,ルシフェラーゼアッセイでNL活性をそれ
ぞれ測定した。野生型 HBV を感染させた PXB 細胞は感染 3 日後から HBV RNAを発現し,HBV RNA量は経日的に増加した (Fig. 5a)。HBV/NL感染細 胞でも同様にHBV RNA量は経日的に増加し,さらに,細胞のNL活性はHBV RNA量と相関した (Fig. 5b)。以上から,HBV/NLは野生型HBVと同様に感染
細胞内でHBV RNAを発現し,その発現はNL活性でモニターできることが示
唆された。また,HBV/NL を HepG2/NTCP 細胞に感染させた場合も,同様に 細胞内HBV RNA量とNL活性の相関が確認された (Fig. 5c)。次に,HBV/NL 感染細胞において,HBV 複製の起点であり,HBV RNA 転写の鋳型となる cccDNA 19)が形成されるかを検討した。NTCP 強制発現 HuH7 細胞に野生型 HBV又はHBV/NLを感染させ,感染4日後の細胞からHirt DNAを抽出した
32)。Southern blot法によってHBV DNAを検出したところ,2.2 kbにcccDNA のバンドが出現した (Fig. 5d)。また,2.2 kbのバンドは,HBV DNAをsingle digestionするXbaI処置によって3.2 kbのlinear DNAにシフトすることから,
HBV cccDNA であることが確認された。以上から,HBV/NL 感染でも野生型
HBV感染と同様に,HBV cccDNAを形成することが示された。
最後に,HBV/NL感染におけるHBV阻害剤の効果を検証した。HBV阻害剤 として,HBV侵入阻害剤作用が確認されるhuman hepatitis B immunoglobulin (HBIG) 33)及びHeparin 34),HBV転写阻害作用が確認されるIFN-β 7, 8)を評価 した。HBV/NL の HepG2/NTCP 感染系において各 HBV 阻害剤を評価したと ころ,全ての HBV阻害剤はその容量依存的に NL活性を阻害した (Fig. 5e)。
以上から,HBV/NL 感染系において HBV の侵入阻害剤及び転写阻害剤を評価 可能であることが示された。
Figure 5. Activity of NL in HBV ⁄ NL-infected cells reflects HBV RNA levels. (a) PXB cells (2×
105/well) were infected with approximately 2×107 copies wild-type HBV and HBV RNA (blue) and DNA (purple) in 10% of the cell lysates measured by quantitative RT-PCR and quantitative PCR, respectively, at the indicated days after infection. (b, c) PXB or HepG2 ⁄ NTCP cells (2×105/well) were infected with approximately 2×107 HBV ⁄ NL and NL activity (orange), HBV RNA (blue), and DNA (purple) in 10% of cell lysates measured at the indicated days after infection. The data shown are indicative of three independent experiments. (d) Southern blot analysis of covalently closed circular DNA (cccDNA) fractions obtained from 1×107 HuH7 ⁄ NTCP cells infected with approximately 1000 equivalent copies of wild HBV or HBV ⁄ NL DNA per cell. The cccDNA fraction was prepared by the method in materials and methods. Digestion with XbaI, which has a single cutting site in the genomes, converts the cccDNA to a single linear form. (e) Effect of known anti-HBV agents on HBV/NL infection. HepG2/NTCP cells were infected with HBV/NL in the presence of HBIG, heparin and IFN-β at the doses indicated, as well as 2% DMSO and 4% PEG8000. The level of HBV
replication was determined by reporter activity 6 days after infection. HBIG is an antibody that neutralizes HBV infection and heparin is an inhibitor for enveloped viruses.
第3節 考察
本章の目的は、HBVゲノムにレポーター遺伝子であるNLを挿入することで pHBV/NL を構築することと,調製したHBV/NLがHBV 複製をNL活性で評 価可能であるかを検証することであった。
第1 節において,pHBV/NLとpHBV-Dをコトランスフェクションすること によって感染性レポーターHBV の HBV/NL が産生し,感染細胞において NL を発現することを確認した。pHBV/NL と pHBV のコトランスフェクションに
よるHBV/NLの産生も検討したが,産生ウイルスを感染させた場合の感染細胞
における NL 活性は pHBV-D を用いた場合の 50%程度であった (data not shown)。pHBV-Dはencapsidation signalへの変異導入によりHBVゲノムが 粒子 中 へ取 り 込 ま れ な い が ,pHBV は HBV ゲノ ム の取り 込み にお い て
pHBV/NLと競合するため,産生粒子中のpHBV/NLゲノムの割合が低下し,感
染時 の NL 活 性が低下 す ると 考察 され る。 よって ,HBV/NL の産 生に は
pHBV/NL と pHBV-D のコトランスフェクションが最適であると示された。
CsCl 密度勾配遠心による産生粒子性状の検討において,感染性を持たないが
HBV DNAを含むフラクションが確認された。このフラクションはHBs抗原も
検出されないことから,HBV ゲノムを包括したカプシドが HBs を纏わずに放 出されたものであると考えられ,同様の結果は他のグループからも報告されて いる35)。また,これまでに複数のレポーターHBVに関する報告がなされている が,いずれも粒子産生量が少ない点と十分なレポーター活性が得られない点が 課題であった28-30)。一方,HBV/NLは野生型HBVと同様の濃縮・精製効率で,
感染細胞における十分なNL発現が認められた。その原因として,NLが非常に 高いレポーター活性を持つため 31),少ない感染量でも十分なレポーター活性を 得られることが考えられる。また,HBV/NL+polの実験から分かるように,HBV ゲノム長は感染性粒子の産生に大きく影響する。これらの結果より,レポーター 活性が非常に高く,ゲノム長も小さいNLの使用は,ゲノム長サイズ制限の厳し いレポーターHBV構築において,最も適切な選択であると考えられた。
第2節では,HBV/NLの感染後のライフサイクルを検討した。HBV/NLのラ イフサイクル検証において,HBV cccDNAの形成を確認することは非常に重要 である。HBV 感染細胞において全長の HBVゲノムが宿主 DNA に組み込まれ ることは稀であり,cccDNA が持続感染及びゲノム複製に寄与する 19)。また,
HBV cccDNA からの HBV RNA の転写は様々な宿主因子が関与する複雑な過 程であり,新規創薬標的の存在も期待される 36-39)。第 2 節における実験から,
HBV/NL感染細胞において,野生型HBVと同様のHBV RNAの経日的増加と,
HBV cccDNAの産生が確認された。このことは,HBV/NL感染においてHBV
ゲノムが感染後に核内でcccDNAへと変換され,その後HBV RNAを産生し続 けることを示唆している。また,感染細胞中のHBV RNA量はNL活性と相関 し,その活性はHBV侵入阻害剤及び転写阻害剤によって低下した。これらの結
果より,HBV/NLは感染細胞中レポーター活性を測定することで,HBV感染の
侵入〜cccDNA形成〜HBV RNA転写までを評価できることが明らかになった。
一方,HBV/NL感染細胞ではHBV DNAの経日的増加は確認されない。このこ とは,HBV/NLがcore及びpolを欠損するため,HBV RNAのカプシドへの取 り込み及びその後の逆転写によるHBV DNA形成が進行しないことが原因であ ると考えられる。したがって,Fig. 6 に示すように,HBV/NL感染で進行する ライフサイクルはやはり侵入からHBV RNA転写までである。
Figure 6. Schematic of the HBV/NL life cycle. HBV/NL enters into cells through virus receptors including NTCP. Capsid-associated relaxed circular DNA (rcDNA) is uncoated and converted to covalently closed circular DNA (cccDNA) in the nucleus. cccDNA functions as a template for mRNA transcription. NanoLuc is expressed from the pre genome RNA.
第3章
HBV/NLを用いたスクリーニング系の構築とその実施
第2章における検討にて,HBV/NL感染細胞におけるNL活性はHBV RNA 量と相関し,NL活性でHBVの侵入〜cccDNA形成〜HBV RNA転写までをモ ニターできることを確認した。NL活性測定の手順は,細胞可溶化と基質試薬の 添加のみである事から,非常に迅速・簡便でありHTSに適している。一方,レ ポーターHBV感染系にてHTSを実施するためには,HBV感染感受性の高い細 胞株の樹立も必要である。HepG2細胞へのNTCP強制発現はHBV感染感受性 を増強することが知られており,感受性の高いクローンの選抜にHBV/NL感染 系は最適である。
本章では,HSTを実施するために,HBV感染感受性の高いHepG2/NTCP細 胞株の樹立を実施した。また,新規抗HBV薬の探索を目的として,HepG2/NTCP 細胞株へのHBV/NL感染系を用いた,化合物ライブラリーのスクリーニングを 実施した。さらに,化合物スクリーニングでNL活性低下を示した化合物につい て,実際にHBV複製阻害作用があるかを検証した。
第1節 HBV高感受性NTCP強制発現HepG2細胞株の樹立
Human NTCPの強制発現によって,HBV感染が可能となる細胞株を選抜し
た。子宮頸がん細胞株であるHeLa細胞,肝癌細胞株であるHepG2細胞,HuH7 細胞,HuH6 細胞及び PLC/PRF/5 細胞,不死化肝細胞株である PH5CH 細胞
40)及びHuS細胞41)に,NTCPを一過性発現させた。その後HBV/NLを感染さ せ,6日後のNL活性を測定したところ,NTCP発現によってHepG2細胞及び HuH7細胞のHBV感染感受性が大きく増強した (Fig. 7a)。HuH7細胞は細胞 増殖が遅かったことから,HBV感染感受性が高く細胞の増殖及び接着性が良好
であった HepG2細胞を選抜し,HBV高感受性細胞株の樹立を目的としたクロ
ーニングを開始した。NTCP一過性発現HepG2細胞を限界希釈し,G418を用 いて薬剤耐性セレクションすることで,複数の HepG2/NTCP クローンを取得 した 。各 クロ ーン に HBV/NL を 感染 させ,NL 活性を測定したところ , HepG2/NTCP#22が最も高いHBV感受性を示した (Fig. 7b)。Western blot解 析の結果,HepG2/NTCP#22はNTCPを高発現する事も確認された (Fig. 7c)。
HBV/NL 感 染 実 験 の 再 現 性 及 び 細 胞 増 殖 も 良 好 で あ っ た こ と か ら , HepG2/NTCP#22を化合物スクリーニングに用いた。
Figure 7. Selection of hepatitis B virus susceptible cells using hepatitis B virus/NanoLuc. (a) Analysis of human hepatocyte-derived cell lines and HeLa cells for hepatitis B virus/NanoLuc (HBV/NL) infectivity. HeLa, HepG2, HuH7 and PLC/PRF/5 were obtained from ATCC. PH5CH and HuS cells are described elsewhere 40, 41). Each cell line (-) and NTCP transduced cell line (+) was infected with HBV/NL, and NL activity was measured 6 days after infection. (b) A single cell clone of NTCP-myc gene transduced HepG2 cells was obtained by end-point dilution. After 6 days of HBV/NL infection to each cell clone, NL activity was measured. (c) Western blot of NTCP-myc using the anti-Myc antibody and of b-actin. Plural bands reflect modification in NTCP.
第2節 HBV/NLを用いた化合物ライブラリーのスクリーニング
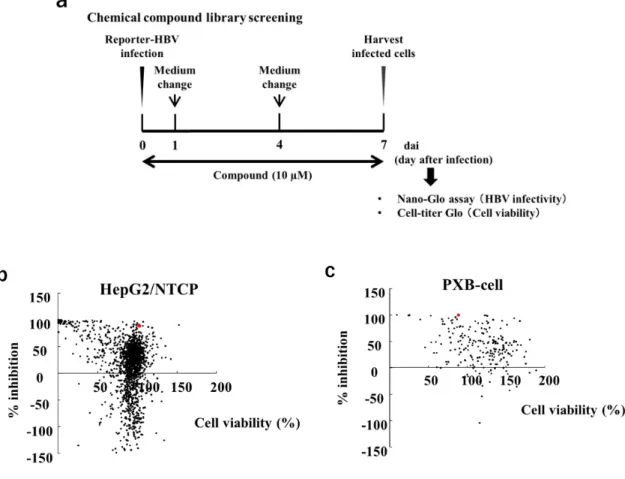
構築したHBV/NL感染系を用いて,化合物ライブラリーのスクリーニングを 実施した。化合物ライブラリーは,US Food and Drug Administration (FDA) 承認薬ライブラリーの 1,827 化合物を用いた。一次スクリーニングでは,感染 細胞にHepG2/NTCP#22を用いた。HepG2/NTCP#22を96 well plateに播種 し,翌日に上清を取り除いて HBV/NL 及び 10 µMの化合物を含む培地を添加 した。HBV/NL感染1および4日後に化合物培地を交換し,感染7日後に細胞 を回収してNL活性及び細胞生存率を測定した (Fig. 8a)。HTSアッセイ系の精 度を示す指標として,データのばらつき及びシグナルの強度から計算されるZ’ - factorがある。Z’ =1-(3xSD100%+3xSD0%)/(Av100%-Av0%)で算出され,Z’ ≧0.5が 求められるが42),本試験系はZ’ =0.773であり良好であった。また,陽性対象と して評価したHeparinや,HBV侵入阻害作用が報告されるCyclosporin A 43)処 置によってNL活性が低下した事から (data not shown),本スクリーニングの 有効性が確認された。一次スクリーニングで1,827化合物を評価した結果,NL
活性を40%以上低下させ,細胞生存率が80%以上であった化合物は218化合物 であった (Fig. 8b)。
癌化細胞株は,正常細胞と比較して様々な宿主性因子の発現が転写レベルで 異なるため,癌化細胞株でスクリーニングした化合物は正常細胞で作用を示さ ないことがある。したがって,二次スクリーニングではマウスで作製した primary human hepatocyte (PHH)であるPXB細胞22)を用いた。PXB細胞は HBV 感染及び複製が認められる primary hepatocyte であり,HBV 感染時の HBIG 処置で感染阻害も確認される化合物評価が可能な細胞である 44)。一次ス クリーニングと同様のスケジュールで,218 化合物の二次スクリーニングを実 施したところ,31化合物がNL活性を85%以上阻害し,細胞生存率も80%以上 を維持した (Fig. 8c)。それらの内,Athenex 社が抗癌剤として開発中の KX2- 391は,細胞毒性なしにNL発現を強力に阻害した (Figs. 8b and 8c, red dot)。
Figure 8. High-throughput screening of anti-HBV drugs using recombinant HBV/NL. (a) Scheme for experiments b and c. HepG2/NTCP or PXB cells were infected with approximately 1000 equivalent copies of HBV/NL DNA per cell in the presence of 10 µM of the test compound. At 1 and 4 days after infection, infected cells were washed with PBS. At 7 days after infection, NanoLuc activity and cell
viability were measured using the Nano-Glo Luciferase Assay System and CellTiter-Glo Luminescent Cell Viability Assay Kit. (b) Then, 1,827 compounds were tested using HepG2/NTCP cells and plotted based on the percentage of HBV/NL inhibition (Y-axis) and cell viability (X-axis), normalized to a DMSO control. Red dot indicates KX2-391. (c) Next, 218 compounds were selected from 1,827 compounds as candidate anti-HBV inhibitors and tested for anti-HBV activity using PXB cells. Each data point represents the average of two biological replicates per compound, normalized to a DMSO control.
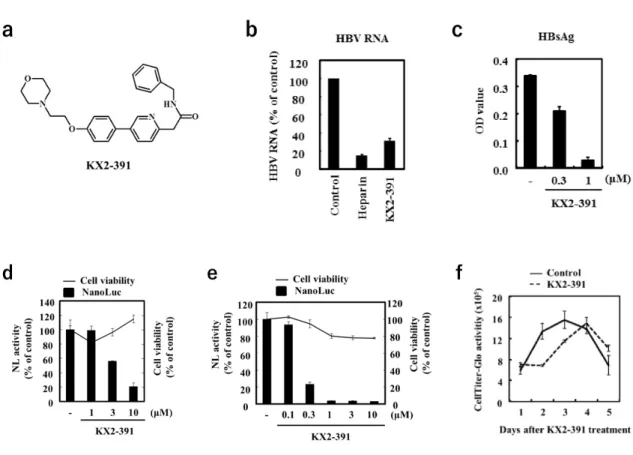
第3節 KX2-391の抗HBV活性
KX2-391は,ATP非競合性Src阻害作用とtubulin polymerization阻害作用 を持つdual inhibitorである (Fig. 9a)。KX2-391は2つの作用によって癌細胞 の異常増殖を阻害する事から,前立腺癌や日光角化症の治療薬として臨床試験 が実施されている45-47)。HepG2/NTCP#22及びPXB細胞のHBV/NL感染系に おいてNL活性を低下させたKX2-391 に関して,その抗HBV活性をより詳細 に検証した。まずは,野生型HBV感染に対して抗HBV活性を発揮するか検証 した。10 µMのKX2-391処置下で野生型HBVをHepG2/NTCP#22に感染さ せ,4日後の感染細胞内HBV RNA量を定量したところ,KX2-391はHBV RNA 量を低下させた (Fig. 9b)。また,野生型HBVをPXB細胞に感染させ,5日後 の培養上清中のHBs抗原量をELISAによって定量したところ,KX2-391は上 清に産出されるHBs抗原量を低下させた (Fig. 9c)。以上の結果から,KX2-391
はHBV/NLのみに作用するのではなく,野生型HBV感染の複製を阻害するこ
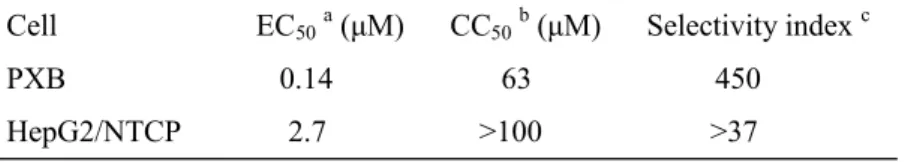
とが示唆された。続いて,KX2-391の50% cytotoxic concentration (CC50)及び 50% effective concentration (EC50)を算出した。3倍公比で段階希釈したKX2- 391処置下で,HepG2/NTCP#22又はPXB細胞にHBV/NLを感染させ,細胞 生存率と NL 活性を評価した。HepG2/NTCP#22 への HBV/NL 感染系におい て,KX2-391は濃度依存的にHBV/NLを阻害し,EC50=2.7 µM, CC50>100 µM であり,活性と細胞毒性の乖離を示すselectivity index (CC50/EC50)は>37であ った (Fig. 9d)。また,PXB 細胞への HBV/NL 感染系では,EC50=0.14 µM, CC50=63 µMであり,selectivity indexは450であった (Fig. 9e)。以上の結果 から,KX2-391 は細胞毒性のない濃度で HBV 複製を強力に阻害する化合物で あることが示された (Table 1)。一方,HBV感染時のようにconfluentな状態で はKX2-391の細胞毒性は認められないが,subconfluentな状態のHepG2細胞 に対してKX2-391は細胞増殖抑制作用を示した (Fig. 9f)。
Figure 9. KX2-391 inhibits HBV replication. (a) The compound structure of KX2-391. (b) HepG2/NTCP cells were infected with 100 equivalent copies of HBV wild per cell in the presence of 100 U/ml heparin and 10 µM KX2-391. At 1 day after infection, infected cells were washed with PBS and the level of HBV mRNA was determined by qRT-PCR at 5 days after infection. Heparin is a general inhibitor for enveloped viruses including HBV. (c) PXB cells were infected with 100 equivalent copies of HBV wild per cell in the presence of 0.3 µM or 1 µM KX2-391. At 1 day after infection, infected cells were washed with PBS, and at 5 days after infection, the level of HBsAg in the supernatant was determined by HBsAg ELISA. (d and e) HepG2/NTCP (d) or PXB (e) cells were infected with HBV/NL, approximately 1000 equivalent copies of HBV/NL DNA per cell, in the presence of KX2-391. DMSO was used as a negative control. At 1 and 4 days after infection, infected cells were washed with PBS. At 5 days after infection, NanoLuc activity and cell viability were measured using the Nano-Glo Luciferase Assay System and CellTiter-Glo Luminescent Cell Viability Assay Kit. (f) HepG2 cells were seeded at 2×104 cells/well in a 96-well plate under subconfluent conditions. Cells were treated with 1 µM KX2-391 and cell viability was determined by CellTiter- Glo Luminescent Cell Viability Assay Kit at each day after KX2-391 treatment.
Table 1. Anti-HBV activity and toxicity of KX2-391
第4節 考察
本章の目的は、HBV/NL を活用して HBV 高感受性細胞をクローニングし,
化合物スクリーニングを実施して化合物を選抜することであった。
第 1 節において,NTCP 高発現 HepG2 細胞クローンの選抜を実施した。近 年,NTCPがHBV感染の受容体として同定されたが24),その他のco-receptor の存在やHBV感染に必要な宿主因子等は明らかではない。また,human NTCP を発現させたマウス肝細胞へのHBV感染は成立しないが,マウス肝細胞とヒト 肝細胞の融合細胞には HBV 感染が成立するという報告から,NTCP 以外にも HBV 感染に必須の宿主因子が存在することが示唆されている 48)。HBV/NL 感 染実験の結果,HepG2/NTCP#22 が最も高い感染感受性を示したが,他のクロ ーンと比較してNTCP発現量が高い訳ではない。この事から,HepG2/NTCP#22 は NTCP以外の宿主因子に関連して,他のクローンより HBV感染感受性が優 れている可能性がある。HepG2細胞はクローナリティが低く、再クローン化に よってその細胞の性質が変化する事も知られている。したがって,HBV感染感 受性の高いHepG2細胞クローンの選抜には,NTCP発現量を比較するのみでな く,実際のHBV感染性試験の実施が重要であり,感染性を迅速・簡便に評価可
能なHBV/NLを用いた選抜は適切であると考えられた。
第2節において,実際にHBV/NL感染系を用いて化合物のスクリーニングを 実施した。一次及び二次スクリーニングにおいてNL活性を指標に31化合物を 選抜したが,その中には既にHBV複製阻害作用が報告されているCyclosporin A 43)やIrbesartan 49)が含まれた。また,選抜化合物のKX2-391も野生型HBV 感染に対して強い阻害を示した事から,HBV/NL 感染系によるスクリーニング の 有 効 性 が 確 認 さ れ た 。 さ ら に , 実 際 に 選 抜 31 化 合 物 に つ い て HepG2/NTCP#22 の野生型 HBV 感染系で評価したところ,18 化合物が HBV RNA を大きく減少させた事から,野生型HBV感染系との大きな相関も認めら
Cell EC50a (µM) CC50b (µM) Selectivity index c PXB 0.14 63 450 HepG2/NTCP 2.7 >100 >37 a EC50: half effective concentration
b CC50: half cytotoxic concentration c Selectivity index: CC50/EC50