九州大学学術情報リポジトリ
Kyushu University Institutional Repository
胎生魚ウミタナゴ科魚類3種の交尾行動と社会構造に 関する研究
櫻井, 真
https://doi.org/10.11501/3135131
出版情報:Kyushu University, 1997, 博士(農学), 論文博士 バージョン:
第5章 日本産ウミタナゴ科魚類の交尾生態の比較
胎生魚であるウミタナゴ科魚類は, 日本にはウミタナゴDitrema属2種,
オキタナゴNeoditrema 属1種の2属3種が分布する。 ウミタナゴDitrema temminckiは北海道南部から九州中部の岩礁域やガラモ場に, アオタナゴD.
viridisは青森県以南から九州中部の内湾域のアマモ場を中心に, オキタナゴ Neoditrema ransonnetiは北海道南部から九州中部の外洋性の岩礁域を中心
に生息する。 オキタナゴは中-表層性でカラヌス目, アミ目等の動物プラン クトンを摂餌するなど, 海底付近を遊泳し底棲の小型動物を捕食するウミタ ナゴ属の2種と生態が異なる。
本研究では, これら3種について九州沿岸の各地, 福岡県津屋崎町(ウミ タナゴ), 長崎県壱岐郡芦辺町(アオタナゴ), 大分県南海部郡上浦町(オ キタナゴ)で標本採集と潜水観察を行った。 これにより, ウミタナゴ科3種 の交尾期, 交尾期の雄の二次性徴, 体長組成や年齢, 当歳のYOY個体と1歳
以上のOlder個体のサイズ差, さらに, 野外における交尾期の生息場所, 社 会構造, 求愛・交尾行動, 婚姻形態, YOY個体とOlder個体問の求愛・交尾 行動の有無, 摂餌頻度を明らかにした。 以上の結果をまとめたものをTable 15に示す。
本章では, これらの結果を3種間で比較して, 共通点と相違点を検討する ことにより, 胎生魚ウミタナゴ科魚類の野外での交尾生態について考察す る。 さらに, 他の胎生魚の交尾生態と比較して胎生魚の交尾生態の共通点を 検討する。
ヤ 4
。ー与4、・
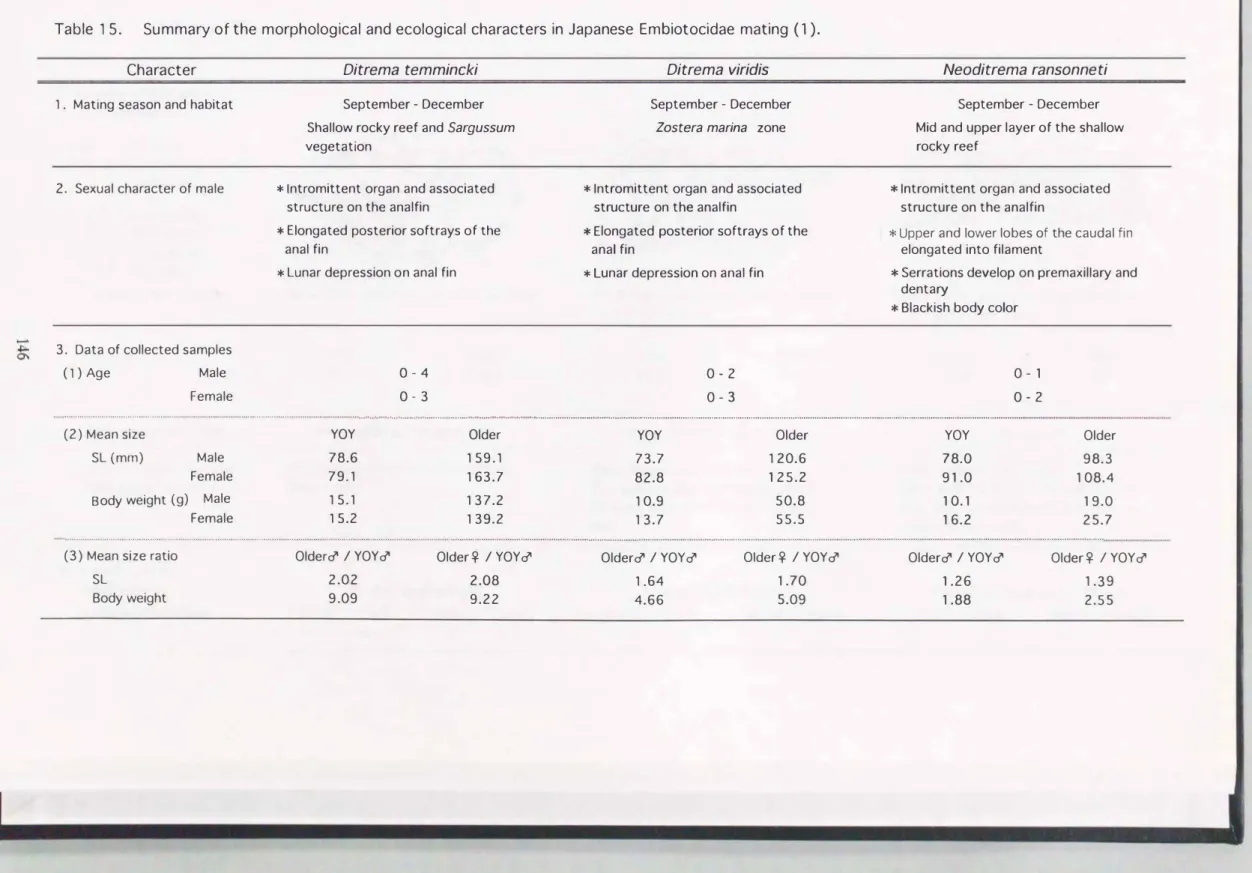
Table 15. Summary of the morphological and ecological characters in Japanese Embiotocidae mating (1).
Character 1. Mating season and habitat
2. Sexual character of male
3. Data of collected samples
(1) Age Male
Female
Ditrema temmincki
September - Oecember Shallow rocky reef and Sargussum vegetatlon
* Intromittent organ and associated structure on the analfin
* Elongated posterior softrays of the anal fin
* Lunar depression on anal fin
0-4 0-3
Ditrema viridis
September - Oecember Zostera marina zone
* Intromittent organ and associated structure on the analfin
* Elongated posterior softrays of the anal fin
* Lunar depression on anal fin
0・2 0・3
Neoditrema ransonneti
September - Oecember Mid and upper layer of the shallow rocky reef
* Intromittent organ and associated structure on the analfin
* Upper and lower lobes of the caudal fin elongated into filament
* Serrations develop on premaxillary and dentary
* Blackish body color
0-1 0・2
••••...••....•...•...•••...•••...• e ・・・・・・・・・・ ・・. ... ー・・ ・ ・・・ ・・・・・4・・ ・・・・...・・・・..・ ・ ・・・ ・ ー ・... ー ・・・・・・・・・・・・・・・.... ‘・・・...・・...‘...
(2) Mean size
SL (mm) Male Female Body weight (g) Male
Female
YOY
78.6 79.1 15.1 15.2
Older YOY
159.1 73.7
163.7 82.8
137.2 10.9
139.2 13.7
Older YOY Older
120.6 78.0 98.3
125.2 91.0 108.4
50.8 10.1 19.0
55.5 16.2 25.7
...・・ー...・...ー・・・・ー・・・・・ ・ ...・・・・・・・ ・・・・・・・・‘・・ ・・ー・-・・・ー...ー...ー..ー・・・4・・・...0-...・・・・...・・・・・・・・・・・・・・・・・・
(3) Mean size ratio Older♂/ YOY♂ Older平/YOY♂ Older♂/YOY♂ Older平/YOY♂ Older♂/YOY♂ Older平/YOY♂
SL 2.02 2.08 1.64 1.70 1.26 1.39
Body weight 9.09 9.22 4.66 5.09 1.88 2.55
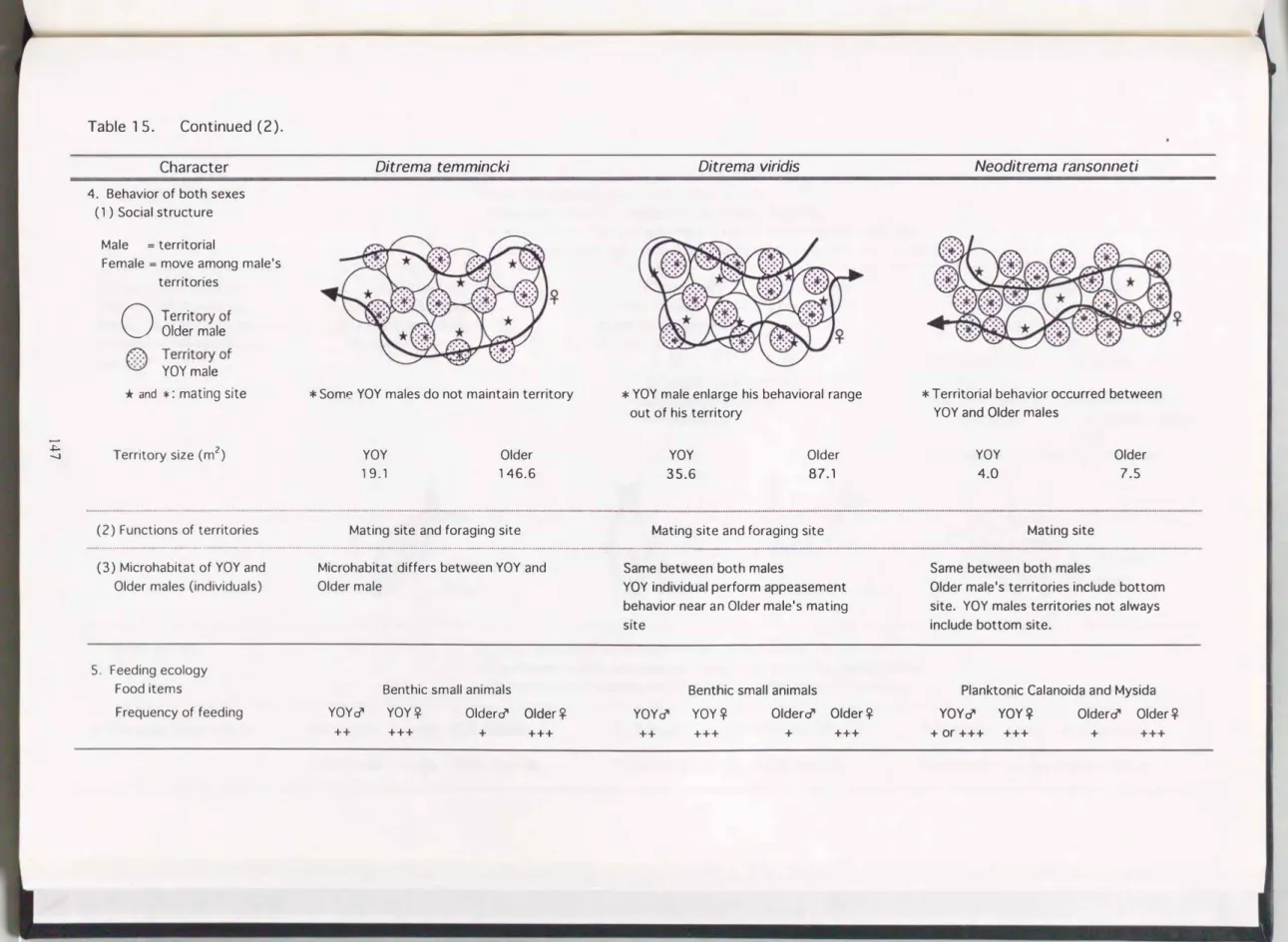
Table 1 5. Continued (2).
Character Ditrema temmincki Ditrema viridis Neoditrema ransonneti
4. Behavior of both sexes (1 ) Social structure
Male = territorial
Female = move among male's terntones
OM
Older male 平。
T州仰offYOY male
* and *: matl ng slte * YOY male enlarge his behavioral range
out of his territory
* T erritorial behavior occurred between YOY and Older males
* Some YOY males do not maintain territory
】hHJ可
Territory size (m2) YOY
4.0
Older 7.5 Older
146.6
YOY 35.6
Older 87.1 YOY
19.1
(2) Functions of territories Mating site and foraging site Mating site and foraging site Mating site
...ー ・ ・・ ・ ・・・・・・・・ー ・・・・・ ・・・ ・ ...・4・・ ・.... ・・ ・・・・・・・・・ ・・・・・・・・・・ー・ ・ ... ... ・ ・ ...・ ・・・...
(3) Microhabitat of YOY and Microhabitat differs between YOY and Same between both males Same between both males
Older males (individuals) Older male YOY individual perform appeasement Older male's territories include bottom
behavior near an Older male's mating site. YOY males territories not always
slte include bottom site.
5. Feeding ecology
Food items Benthic 5mall animals Benthic 5mall animal5 Planktonic Calanoida and Mysida
Frequency of feeding YOY♂ YOY平 Older♂ Older平 YOY♂ YOY字 Older♂ Older平 YOY♂ YOY♀ Older♂ Older平
++ +++ + +++ ++ +++ + +++ + or +++ +++ + +++
Table 1 5. Continued (3).
Character Ditrema temmincki Ditrema viridis Neoditrema ransonneti
6. Courtship and mating behavior
These characters are same among three species
* Pair formation after a sequence of courtship behavior.
* Frequency of mating is quite lower than frequent courtship behavior.
* Mating occurs by a pair within 2-3 sec. at the mating site in the territory.
A sequence of courtship behavior and copulation,
and differences of courtship behavior within and among specles.
-hF∞
<Older and YOY males>
(1 ) Head standing display (2) zigzag swimming (3) copulation
<Older male> <YOY male>
(1 ) Head standing (1 ) Feeding with display and presentation female(s)
of the body I
(2)
守守
biting at the mating site (3) zigzag swimming ( 4) copulation( 4 )Copulation in the mid water column
* Streaking like behavior by a YOY male
牟
<Older and YOY ma剖les>
(υ, ) Courtship dance (
(3) Chasing each (3) Circling other
( 4) Copulation near the bottom
Courtship dance Head standing
display
Head standing
display Feeding with female(s)
7. Mating system Same for three species = multiple mating by both sexes (Promiscuity)
* Female visit male's territories and mate males during the mating season.
* Male mate females which visit his territory during the mating season.
8. Size assortative mating YOY male __. YOY female Older male -一惨 Older female
YOY male --t惨YOY female Older male 一一砂 Older female
YOY male
ミ
YOY female Oldermale -一惨 Older female1. 交尾期と交尾期の生息場所
本研究ではまず, 日本産ウミタナゴ科3種の交尾期について検討した。 そ の結果, 津屋崎町のウミタナゴ, 芦辺町のアオタナゴ, 上浦町のオキタナゴ のいずれも交尾期は9・12月であることが判明した。 従来の報告では, 神奈川 県産(Ishii, 1959 ; Hayase and Tanaka, 1980a)でウミタナゴ, アオタナゴ,
オキタナゴの3種について, 福岡県産(Nakazono et al., 1981)と長崎県産 (水江, 1961b, 1964)でウミタナゴについて, 交尾期が9・11, 12月であるこ とが報告されている。 本研究の結果はこれら既往の知見と一致しており, 関 東以西におけるウミタナゴ科魚類の交尾期はほぼ同じであることが明らかに なった。
次に, 交尾期の生息場所についてみると, 交尾期には, ウミタナゴでは浅 場の岩礁域やガラモ場, アオタナゴではアマモ場, オキタナゴでも浅場の
中ー表層に出現し, 3種とも交尾期には接岸する点で共通していた。
北米西岸に生息する本科魚類は, 潮間帯の潮だまりに出現するものから水 深100 m以深の砂泥底から捕獲される種, さらには, 河川に生息するものま で認められ, 生息環境が多様性に富むことが知られている(Tarp, 1952;
Eschmeyer et a1., 1983; Baltz, 1984)。
これら北米産ウミタナゴ科魚類では出産期に浅場に移動する例が多いが
(Shrode et al., 1983; Eschmeyer et al., 1983; Baltz, 1984) , 交尾に伴う移動 や交尾期の生息場所が判明している種は少ない。 通常は藻場や岩礁などに生 息するCymatogaster aggregataは出産後間もなく交尾するが, 出産と交尾の 時期には浅場に移動する(Wiebe, 1968)。 一方, Embiotoca jacksoni・で は, 周年kelp bed の岩礁に縄張りを維持しており, その中に交尾場所が確
保されている(Hixon, 1981)ため, 交尾に伴う生息場所の変化は無いと推 測される。 通常潮だまりや浅場の岩礁に生息するMicrometrus minimusで は, 求愛行動も同様の場所で観察されることが知られている(Warner and
Harlan, 1982)。 しかしながら, 今回の日本産の3種以外で交尾期と交尾期以 外の生息場所を詳細に比較した例は少なく, 交尾期に接岸する日本産本科魚 類の習性が, ウミタナゴ科で一般的なものか否かは今後の検討を要する。
2. 二次性 徴
ウミタナゴ属2種の雄では, 交尾期には啓鰭鰭膜上に腺様体が発達する,
轡鰭軟条の後端が伸張する, 管鰭基部に半円状のくぼみが現れるというこ次 性徴が見られた。 一方, オキタナゴでは交尾期の雄に, 体色が全体に黒色が 強くなる, 尾鰭上下末端が伸張する, 前上顎骨・歯骨に鋸歯状突起が発達す るという二次性徴が認められた。 しかし, 啓鰭には鰭膜上に腺様体が現れる ものの, 轡鰭末端は伸張せず, 啓鰭基部の半円状のくぼみも顕著ではなかっ た。
北米産のウミタナゴ科では, 交尾期の雄の轡鰭鰭膜上に腺様体が発達する ことが幾つかの種で知られており(Wiebe, 1968; Warner and Harlan, 1982;
OeMartini, 1988) , 本研究のウミタナゴ属とオキタナゴの轡鰭鰭膜上の腺様 体は北米産のものと同様で、あった。 ところが, オキタナゴの二次性徴のう ち, 尾鰭末端の伸張と顎骨・歯骨の鋸歯状突起は他のウミタナゴ科では知ら れておらず, 本種に特徴的な形質である。 鋸歯状突起は雄同士の闘争に使わ れると考えられ, 後述のように社会構造も雄の闘争が激しいことを示唆して いる。
3-(1). 採集標本の年齢
交尾期間中に採集された標本を比較すると, ウミタナゴとアオタナゴの雌 雄では当歳からし2, 3, 4歳の個体が採集された。 オキタナゴではほとんど
が当歳でし2歳の個体はごく少数であった。 オキタナゴでは潜水観察におい ても出現個体はYOY個体が大部分を占め, 多くが2回目の交尾期以前, すな わち1歳の秋の交尾期以前に死亡するものと考えられた。 Hayase and Tanaka
(1980a)は, ウミタナゴ属2種の寿命は3-4歳であること, オキタナゴはほ とんどの個体の寿命が1歳であると報告しているが, 本研究の結果もこれと 一致した。
3-(2). 交尾期の体長組成
交尾期の3種のサイズを比較すると, ウミタナゴが最も大型になり, アオ タナゴがこれに続き, オキタナゴは最小であった。 交尾期のYOY個体の標 準体長の平均はTable 15に示すように, ウミタナゴで雄 = 78.6 mm,
雌=79.1, アオタナゴで雄=73.7, 雌=82.8, また, オキタナゴで雄=78.0,
雌=91.0であり, 交尾期のYOY個体の体長には3種間で大きな差は認められ なかった。 しかし, OIder個体では上位からウミタナゴ, アオタナゴ, オキ タナゴの}I債で, より大型・高齢の個体が出現した。 それに伴ってOIder個体 の体長の平均値も大きくなり, ウミタナゴで雄=159.1 mm (1-4歳),
隊=163.7 (1-3歳) , アオタナゴで雄=120.6 (1・2歳), 雌=125.2 (1-3歳) であった。 オキタナゴでは, Older個体は雄=98.3 (1歳), 雌=108.4 (1・2 歳)と, ほとんどが1歳で小型であった。
サイズ差に関する同様の現象は体重でも認められた。 すなわち, YOY個 体では体重に3種間で差が小さかったが, Older個体ではウミタナゴの平均体 重が最も大きく, アオタナゴ, オキタナゴがこれに続いた。
3-( 3). YOY雄とOlder個体 の体サイズ の相違
本研究では, YOY雄雄問, および, Older峰雄聞でのみ求愛・交尾が行わ れる sizeassortative matingの有無の検討が課題の一つである。 このため,
YOY雄とOlder個体閣の体サイズの差 (体サイズ比)は形態学的データとし て重要である。 そこで, YOY雄に対するOlder個体の体サイズ比を計算し た。 まず, YOY雄に対するOlder雄の平均サイズ比(Older雄/yOY雄)を求 める と , 体長比( 体重比)はウミタナゴで2.02(9.09)で最も大きく, アオ
タナゴで1.64( 4.66) , オキタナゴでは1.26( 1.88)と 小さくなった。 次に YOY雄に対するOlder雌の平均サイズ比(Older雌/yOY雄)を求めると , 体 長比(体重比)はウミタナゴで2.08(9.22) , アオタナゴで1.70( 5.09) , オ キタナゴでは1.39( 2.55)と最も小さくなった。 本章8節で述べる ように,
size assortative matingはウミタナゴ属の2種で認められたが, オキタナゴで は行われなかった。 すなわち, YOY個体とOlder個体のサイズ差が小さい場 合にはsizeassorta tive ma tingが崩れることが示唆された。
4・( 1 ). 社会構造
ウミタナゴ科魚類の交尾期の社会構造は北米に多種が生息する にも関わら ず, 従来不明であった。 本研究により, 日本産ウミタナゴ科3種の交尾期の
社会構造は, 基本的に, 雄=縄張り形成, 雌=移動型で共通している事が明 らかになった。
縄張りの平均面積は, ウミタナゴではOlder雄=146.6 m2, YOY雄=19.1,
アオタナゴではOlder雄=87.1, YOY雄=35.6, オキタナゴではOlder雄=
7ふ YOY雄=4.0であった。 Older雄がYOY雄よりも広い面積の縄張りを維 持する点は3種で共通していたが, ウミタナゴ属に比べてオキタナゴでは縄 張りはかなり狭かった。
Older雄とYOY雄の関係を3種間で比較する。 ウミタナゴとアオタナゴで はOlder雄とYOY雄の縄張りの分布は重複しており, 両者間で縄張り行動は 認められなかった。 このように縄張りの分布が重複する現象は, 胎生魚のカ サゴ雄でも知られている(藤田, 私信)。 カサゴ雄の縄張り行動は体長の近 い個体聞でのみ見られ, 体長が大きく異なる雄閣では縄張りが重複してお り, 本研究のウミタナゴ属と類似していた。 これに対して, オキタナゴでは Older雄とYOY雄の縄張りの分布は重複せず, 両者間で縄張り行動が認めら れた。
Older雄とYOY雄で縄張り維持の強さを比較すると, Older雄では3種いず れもが交尾期を通じて縄張りを維持し, その中に留まって縄張りを防衛し た。 YOY雄では, ウミタナゴで縄張りを持つ個体と持たない個体が出現す ること, アオタナゴで縄張り雄も縄張り外へと行動範囲を広げること, オキ タナゴで雌の出現個体数が少ないときには縄張り活動が不活発になるという 現象が見られた。 このように, YOY雄ではOlder雄ほど縄張りを強く保持し ない傾向が共通して認められた。
次に, 3種間, Older雄とYOY雄問で縄張り行動の頻度を比較する。 ウミ タナゴとアオタナゴでは, Older雄の縄張りは隣接して形成されていたが,
縄張り境界線付近で闘争行動の観察される頻度はアオタナゴではウミタナゴ に比べて低かった。 また, これら2種のYOY雄の縄張りは岩礁やアマモ場に 散在しており, 必ずしも隣接していなかった。 さらに, 侵入雄に対する排除 行動はしばしば観察されたが, 隣接する縄張り雄同士の闘争行動はOlder雄
ほと、顕著ではなかった。 これは, ウミタナゴではYOY雄とOlder雄問で、 microhabitatが異なり棲み分けしているために, YOY雄が縄張りを形成でき る場所が限られること, アオタナゴでは, mating siteとなるアマモ場中の砂 地の分布が, 縄張りの分布を制限していることに由来すると考えられる。 こ れに対してオキタナゴでは, 浅場の中ー表層に密集して縄張りが形成され縄
張り行動が頻繁に観察された。 オキタナゴでは縄張り雄聞の競争が激しいた めに, 鋸歯状突起や尾鰭末端の伸長という二次性徴が発達したと推測され る。
最後に, 他科の胎生魚の社会構造と比較する。 胎生魚の交尾期の野外にお ける社会構造は, カサゴ目フサカサゴ科のメバルSebastes inermis
(Shinomiya and Ezaki, 1991) , カサゴSebastiscus marmora tus (藤田, 私 信)で知られる。 また, カサゴ目カジカ科のアナハゼPseudoblennius
percoidesは, 交尾後, 受精卵をカイメン内に産卵する体内受精魚であり交 尾期の社会構造が研究されている(四宮, 1985)。 これらの種の観察結果で は, メバルやアナハゼ、の社会構造は基本的に, 雄=縄張り形成, 雌=移動型 であった。 一方, 定住性の強いカサゴでは, 周年雄は縄張りを維持し, 雌も ある程度排他的な行動圏を持つ。 さらに, 中米淡水産胎生魚である
Poecillidaeの1種, Poeciliopsis occidentalisを水槽内で飼育すると, 大型の 雄は数日間ではあるが縄張りを維持し, 縄張り内とその周辺の雌に求愛行動 を示す(Constantz, 1975)ことから, 同種の交尾の際の社会構造は緩やか な, 雄=縄張り形成, 雌=移動型であることが示唆される。 以上の知見よ り, 本研究のウミタナゴ科の結果は, メバル, アナハゼや淡水産の
Poecillidaeと類似していると考えられた。
4-(2) . 縄張りの機能
北米産ウミタナゴ科の1種Embiotoωjacksoni・の雄が周年維持する縄張り では, 野外実験が行われている(H以00, 1981)。 それによると, 実験的に 縄張りの一部を覆い隠して摂餌できなくすると, 覆い隠されていない場所で の摂餌頻度の増加や, 縄張りの拡大が見られた。 また, 食性の近い種を縄張 りから強く排除する行動も観察されたことから, この縄張りは餌資源防衛縄
張りであると考えられた。 一方で, 縄張り内に交尾場所となる物陰(mating site )が存在することから, E. jacksoniの縄張りは摂餌場所と求愛・交尾場 所の2つの機能を持っと結論している。
本研究の3種においても, 縄張り内に必ずmatingsite が認められたことか ら, 3種の縄張りは,(i)求愛・交尾場所として機能すると考えられた。 さら に, ウミタナゴとアオタナゴでは, Older雌がOlder雄の縄張り内で頻繁に 摂餌し, その雌に対して雄が求愛を行ったことから, (ii)摂餌場所としての 機能を持つことが推測された。 しかし, ウミタナゴでは縄張りを持たない YOY雄が出現したり, アオタナゴでは縄張り外に行動範囲を広げるYOY雄 が観察されたことから, YOY雄の縄張りはOlder雄に較べて摂餌場所として の機能が低いことが推測された。 これに対し, オキタナゴは直径1-4 mの狭
い縄張りを持ち, しかもプランクトン食性である。 プランクトンの供給は潮 流に影響され不安定である。 従って, オキタナゴではOlder雄, YOY雄いず れも, 摂餌場所よりも求愛・交尾場所としての機能の方が強いと考えられ る。
4・(3). Older雄と Y OY雄の出現場所
本研究のウミタナゴでは, Older雄は岩礁域に広く分布するのに対し,
YOY雄は岩礁域内の海藻や岩などの物陰に多く出現して両者で microhabitat が異なった。 Older個体とYOY個体でmicrohabitatが異なる現象は, giant
kelp (Macrocystis pyrifera)が繁茂する岩礁に生息する北米産ウミタナゴ 科, E. jacksoni, E. lateralis, Hypsurus caryi, Rhacochilus toxotes, Damalicthys
vacαで知られている(Ebeling and Laur, 1985)。 岩礁やガラモ場に生息す るウミタナゴでは giantkelpに生息する北米産と同様の結果が得られた。
アオタナゴではOlder雄とYOY雄は同じアマモ場に出現したがmatingsite
は必ず異なった。 さらに, Older雄のmating site付近でOlder雄とYOY個体 が出会った場合には, YOY個体はOlder雄からの攻撃を緩和する宥和行動を 示した。 これらの行動により, 両者はアマモ場内で共存しているものと推測 された。
オキタナゴでは, Older雄は必ず海底を合む縄張りを形成したが, YOY雄 の縄張りには海底を合むものと合まないものがあった。 両者は同じ岩礁域の 中-表層に出現し, 両者間で縄張り行動が見られた。 オキタナゴではウミタ ナゴ属と異なり, 交尾期のOlder雄とYOY雄の出現場所が同じで, 両者が対 等に振る舞うという特徴が認められた。
5. 摂餌頻度
ウミタナゴとアオタナゴは, 底棲の小型動物を捕食した。 消化管内容物重 量指数, 腹腔内脂肪量指数, 摂餌頻度の調査の結果, Older雄はOlder雌や YOY個体に較べて交尾期間中の摂餌量が少ないことが示唆された。 交尾期 間中のOlder雄にとって, 雄と配偶してより多くの子孫を残すために, 縄張 り維持や求愛・交尾を行うことは, 摂餌に多くの時間を費やすよりも重要だ と考えられる。 これに対してYOY雄は, 翌年以降にも交尾期を迎えるため に, 当歳の交尾期には摂餌も行い栄養を蓄えることが重要だと推測される。
アオタナゴのYOY雄では, 摂餌しながらmating siteヘ雌を誘導するという feeding with female(s)が観察されたが, この行動は雌に求愛しつつも摂餌で きるために, 雄にとっては有効な方法であると考えられる。
動物プランクトンを捕食するオキタナゴでは, 摂餌は餌となるプランクト ンが出現する場合に限られた。 この状況下においてもOlder雄の摂餌頻度は 低く, ウミタナゴ属と類似していた。 しかし, YOY雄では, 頻繁に摂餌す る個体と, 専ら求愛を行う個体が出現した。 YOY雄は交尾期とはいえ摂餌
が必要であるが, 餌となるプランクトンが出現する際には雌が出現する場合 が多いために, YOY雄の行動に2型が生じたものと考えられる。
一方, 雄ではウミタナゴ, アオタナゴ, オキタナゴいずれにおいても, 摂 餌頻度が高いことが示唆された。 ウミタナゴ科の雌にとっては交尾期の後に 迎える妊娠のために, 交尾に費やすコストを低く押さえて摂餌し, 栄養を蓄 えることが重要であると考えられる。
6. 求愛・交尾行 動
求愛・交尾行動に関しては, (i)雄による一連の求愛行動の後にぺアを形成 する, (ii)求愛行動に比べて交尾の頻度が低い, (iii)縄張り内のmatingsite でベアで交尾する, (iv)交尾は2-3秒の短時間であるという点が3種で共通し ていた。
しかし, 3種閣で詳細に比較すると以下のような相違が認められた。 ま ず, ウミタナゴ属の2種ではともにhead standing displayが見られたが, ウ ミタナゴではこの際の体色の変化が著しく倒立の角度が海底に対して45-75 度であるのに対し, アオタナゴでは体色の変化は顕著ではなく倒立の角度が
海底に対して約90度であった。 さらに, アオタナゴでは雄が雌をmatingsite に誘導した際に砂質底をついばむbitingat the mating site を行ったが, この 行動はウミタナゴでは見られなかった。 アオタナゴ雄聞でも, Older 雄は専 らhead standing displayやpresentation of the bodyにより求愛するが,
YOY雄ではfeeding with female(s) によって雌をmating siteヘ導くという求 愛方法の相違が認められた。
これに対し, オキタナゴではcourtshipdance により求愛を行い, 求愛行 動がウミタナゴ属と異なったo さらに, オキタナゴ雄間でも, 縄張りが海底 を合む場合にはchasing each otherを行い海底の物陰で交尾を行うが, 海底
を含まない場合には circlingを行った後に中層域で交尾するという行動の2型 が認められた。 中層域で交尾する場合には, Older雌雄の交尾の瞬間にYOY 雄がその聞に割り込もうとする streaking様行動が観察された。 この
streaking様行動はオキタナゴに特異なものであった。
最後に本研究の結果を他種と比較する。 北米産ウミタナゴでは, 求愛・交 尾行動に関する報告は非常に少ない。 c. aggregataでHubbs(1917) ,
Wiebe (1968)とShaw and Allen (1977)により記載されているだけであ る。 C. aggrega taは主に海岸線近くの浅場に生息するが( Eschmeyer et a1.,
1983; Baltz, 1984) , 同種の求愛行動は, 第4章で述べたようにオキタナゴの ものと類似していた。 次に, カサゴ自のメバル, カサゴ, アナハゼの野外に おける求愛・交尾行動(Shinomiya and Ezaki, 1991; Fujita and Kohda, 1996;
四宮, 1985)と比較すると, これらは体側誇示などの一連の求愛行動の後に ベアを形成する, 求愛行動に比べて交尾の頻度が低い, 交尾は短時間の内に 行われるという点でウミタナゴ科と共通していた。 また, Poecillidae では,
水槽内で求愛・交尾行動が観察されているが(Constantz, 1975; Schlupp et a1., 1991) , やはり, 繰り返し求愛を行った後にベアで交尾する点で本研 究のウミタナゴ科と一致した。
7. 婚姻形態
本研究では, 交尾は少数例しか観察されず, 同一の雌が2回以上交尾する 例は観察できなかった。 しかしながら, 交尾期の雌雄の行動パターンを検討 すると, 雄は縄張りに来遊する複数の雌に求愛すること, 雌は複数の縄張り を移動すること, ベアによる交尾が行われるもののその結びつきは弱く, 交 尾が終了するとペアは解消するという点が3種で共通していた。 また, 北米 産ウミタナゴ科の既往の知見においても, C. aggregataでは酵素の多型の生
化学的分析結果より, 一腹胎仔の父性に多型が存在すること(Darling et a1.,
1980) , M. minimusではある1尾の雌が複数の雄と交尾すること(Warner and Harlan, 1982)が報告されている。 以上の報告を合わせて判断すると,
本研究の日本産ウミタナゴ科では1交尾期間中に,(i)雌は複数の雄と交尾す る,(ii)雄は縄張りに来遊する複数の雌と交尾するという, 縄張り訪問型 (乱婚型=雄雄いずれも複婚)の婚姻形態(桑村,1988)を持つことが示唆 された。
他の胎生・交尾型魚類で野外の婚姻形態が明らかになっている例は少ない が, メバルやアナハゼではやはり, ウミタナゴ同様の縄張り訪問型の乱婚の 婚姻形態を持つことが雄雄の行動から示唆される(Shinomiya and Ezaki,
1991;四宮,1985)。 カサゴでは, 縄張り雄が雌の行動圏を訪問して複数雌と 求愛すること(藤田, 私信)や, 雌雄いずれもが複数の配偶者と交尾するこ と(Fujita and Kohda, 1996) から, 緩やかな一夫多妻的な婚姻形態を持つ ことが示唆されている(藤田, 私信)。 さらに, 淡水産のPoecillidae の1種
Gam busia affinisでも, 一腹の胎仔には酵素の多型が見られることから, 雌 は複数の雄と交尾することが明らかとなっている(Greene and Brown,
1991)。 本研究の結果は雌雄が複数の配偶者と交尾する点で, これら胎生 魚・体内受精魚の婚姻形態と類似していた。
8. size a s s ortative m ating
行動生態学的視座では, より多く自己の子孫を残すために繁殖の際にどの 様な配偶者を選んでいるか(配偶者選択)を検討することが, 重要な研究課 題の一つである(Krebs and Davis, 1987, 1991)。 幾つかの条件を満たした 場合には配偶者選択の結果, 同サイズの雌雄間で配偶する size assorta tive mating (= homogamy for size)が起きることが予想、されている(Ridley,
1983)。 すなわち, (i)大型の雌はより多くの卵を生産する, (ii)大きい雄ほ ど配偶相手を巡る競争で有利である, (iii)交尾期間が長いの3条件で‘ある。 ゥ ミタナゴ科魚類においては, (i)は多くの種で一般的に認められており
(Baltz, 1984;棲井, 未発表), (iii)も本研究の3種, および北米産(Wiebe,
1968; Wamer and Harlan, 1982)で知られている。 従って, 本科魚類はサイ ズ差が 大きいYOY個体とOlder個体の聞で, 求愛・ 交尾が行われるか否かと いうsize assortative matingの有無を検討することにより, 残る(ii)の条件で ある配偶者選択における雄聞の競争の状況が検討できる格好の材料の一つで ある。
そこで本研究では, ウミタナゴとオキタナゴではRidley(1983)の手法に 基づいて, また, アオタナゴでは雄が遭遇した雌に対して採った行動を観察 することにより size assortative matingの有無の検討を行った。 その結果,
ウミタナゴ属では求愛・交尾行動は専らOlder雌雄, YOY雌雄聞でのみ観察 されて size assortative matingが認められた。 一方, オキタナゴではOlder雄 はOlder雌に対してのみ求愛するが, YOY雄はYOY雌とOlder雌の両者に求 愛して size assorta tive ma tingが崩れていることが分かった。
北米産ウミタナゴ科における知見では, M. minimus (Warner and Harlan,
1982) , C. aggregata (DeMartini, 1988)ではYOY雌雄, Older雌雄問だ、け で求愛・交尾が行われるが, Brachyistius frenatusではYOY雄がYOY雌と Older般の両者に求愛することが知られている(DeMartini, 1988) 。 これら の研究では, size assortative matingと交尾期の社会構造の関係は不明で・あっ た。 本研究により, Older�援を巡って競争の起こらないウミタナゴやアオタ ナゴではYOY雄とOlder雄問で縄張り行動が認められずに両者の縄張りが重
複すること, 一方, オキタナゴ、ではOlder離の獲得を巡ってYOY雄とOlder 雄の問で競争が起こり, 両者間で前述のような顕著な縄張り行動が認められ ることが明らかになった。 つまり, 社会構造におけるYOY雄とOlder雄の
競争(縄張り行動)の有無がsize assortative matingの有無と一致すること が分かつた。
第6章 要約
胎生の生殖様式は, 硬骨魚類の様々な分類群で認められる。 しかしなが ら胎生魚の求愛・交尾行動, その際の社会構造, 婚姻形態に関する知見は 卵生魚類に比べて少ない。 そこで本研究は日本産ウミタナゴ科魚類2属3種,
ウミタナゴ属ウミタナゴDitrema temmincki, アオタナゴD. viridis, オキタ ナゴ属オキタナゴNeoditrema ransonneti について, 野外での交尾生態を明 らかにすることを目的とした。 1980年から1987年の聞に, 交尾期の標本採集 と潜水観察を福岡県, 長崎県, 大分県の九州各地で、行った。 これにより, 二 次性徴, 年齢組成, 当歳のYOY個体と1歳以上のOlder個体のサイズ差, 生 息場所, 社会構造, 摂餌頻度, 求愛・交尾行動, 婚姻形態, size assorta tive
mating (同じサイズグループの雌雄による配偶)の有無を明らかにした。 得 られた結果は以下のとおりである。
1. 交尾期 と交尾期の生息場所
本研究の3種の交尾期はいずれも9・12月であった。 この時期には浅場に移 動, 接岸する習性が共通して見られた。 すなわち, 交尾期にウミタナゴでは 岩礁域やガラモ場の浅場, アオタナゴではアマモ場, オキタナゴでは岩礁域 の中ー表層に出現した。
2. 二次性徴
交尾期のウミタナゴとアオタナゴの雄では, 轡鰭鰭膜上に腺様体が発達す る, 啓鰭軟条後端が伸長する, 轡鰭基部に半円状のくぼみが顕著になるとい う共通した二次性徴が認められた。 一方, オキタナゴの雄では, 腎鰭鰭膜上
に腺様体が発達する他にも, 婚姻色が現れる, 尾鰭軟条の上下末端が伸長す る, 前上顎骨と歯骨に鋸歯状突起が発達するという二次性徴が認められた。
オキタナゴにおけるこれらの形質の発達は, 雄聞の競争が激しいことを示唆 すると考えられた。
3. 年齢組成, YOY雄とOlder個体聞の体 長差
交尾期の出現個体は, 当歳のYOY個体と1歳以上のOlder個体の2つのサイ ズ、グループに区分された。 YOY個体の平均体長(SL mm)は, ウミタナゴ
=78.6 (雄),79.1 (雌) ;アオタナゴ=73.7,82.8 ;オキタナゴ=78.0,91.0で あった。 Older個体の平均体長は, ウミタナゴ=159.1,163.7 ;アオタナゴ
=120.6,125.2 ;オキタナゴ=98.3,108.4であった。
Older雄の年齢, および, Older雄とYOY雄聞のサイズ比(Older$釘YOY 雄を標準体長, 体重の平均値から計算)をみると, ウミタナゴでは Older雄 の年齢は 1・4 歳でサイズ比は標準体長で 2.02(体重で9.09), アオタナゴで は1・2歳でサイズ比は1.64(4.66) , オキタナゴでは1歳でサイズ比は1.26 (1.88)であった。 次にOlder雌の年齢, および, Older隊とYOY雄聞のサ イズ比(Older雌/yOY雄を標準体長, 体重の平均値から計算)をみると,
ウミタナゴではOlder雌の年齢は1-3歳でサイズ比は2.08(9.22) , アオタナ ゴでは1-3歳でサイズ比は1.70(5.09) , オキタナゴでは1・2歳でサイズ比は 1.39 (2.55)であった。
オキタナゴにおいてはOlder雌雄の個体数は少なく, 出現個体のほとんど は当歳のYOYであった。 従って本種では, 大部分の個体は1歳の秋に迎える 2回目の交尾期より前に死亡し 一部の個体だけが複数回の交尾期を経験す ると考えられた。
4. 社会 構造, 縄張りの機能, Older雄とYOY雄の関係
本研究の3種は交尾期には, 雄=縄張り形成, 雌=移動型の社会構造を持 つことで共通していた。 しかし, ウミタナゴ属の2種ではOlder雄とYOY雄 の縄張りが重複しており, 両者間で縄張り行動はみられないが, オキタナゴ では両者の縄張りは重複せず, Older雄とYOY雄閣で顕著な縄張り行動が認 められるという相違があった。
縄張りは3種共に求愛・交尾場所として機能すると考えられたが, ウミタ ナゴ属の2種では摂餌場所としての機能も併せ持っと推測された。
ウミタナゴではOlder雄とYOY雄で岩礁内のmicrohabitat が異なった。 ア オタナゴではYOY個体が lateral displayや摂餌という宥和行動を示して Older雄からの攻撃を緩和していた。 これによりOlder雄とYOY個体はアマ
モ場に共存できると推測された。 オキタナゴでは岩礁の中-表層でOlder雄と YOY雄が対等に振る振る舞い, ウミタナゴ属と異なった。
5. 摂餌頻度
ウミタナゴとアオタナゴは底楼の小型無脊椎動物を, オキタナゴは動物プ ランクトンを餌としていたが, 交尾期のYOY . Older個体問, 雌雄聞の摂餌 頻度に関しては多くの点で共通点が認められた。 すなわち, Older雄は専ら 縄張り行動と求愛・交尾を行い摂餌頻度は低いが, 雌では頻繁に摂餌して栄 養を蓄え, 交尾後の懐妊に備えていると考えられた。 一方, YOY雄では,
ウミタナゴ属で雌同様に摂餌頻度が高く, 翌年以降の交尾に備えていると推 測されたが, オキタナゴでは摂餌頻度が高い個体と低い個体が観察され, 必
要量を摂餌していると考えられた。
6. 求愛・交尾行 動
求愛・交尾行動に関しては, 一連の求愛の後にベアを形成する, 縄張り内 のmating siteで短時間で交尾する, 求愛行動は頻繁に観察されるが交尾の
頻度が低いという点で3種は共通していた。 これらの特徴は, 他の胎生・交 尾型魚類の求愛・交尾行動とも共通していた。 ウミタナゴ属2種の求愛行動 はほぼ同じであったが, head standing displayやbiting at the maging siteな
どで相違がみられた。 また, アオタナゴのYOY雄では head standing display の替わりに, 摂餌しつつ 雌をmating siteへ誘導する場合が多いという傾向 が認められた。 一方, オキタナゴでは一連の求愛行動がウミタナゴ属と異 なった。 さらに, 中層でOlder峰雄のベアが交尾する際にYOY雄が割り込む
streaking様行動が観察された。
7. 婚姻形態
日本産ウミタナゴ科3種の婚姻形態は, 交尾期の社会構造, 求愛・交尾行 動の観察結果より, 1交尾期間中に 雌雄が複数の異性と配偶するという縄張 り訪問型(乱婚型)で共通していることが示唆された。 これは, 胎生・交尾 型魚類の既知の知見と類似していた。
8. size a s sortative m ating
ウミタナゴ, アオタナゴのYOY · Older個体, およびオキタナゴのOlder 個体では, size assortative matingが観察された。 しかし, オキタナゴの YOY雄はYOY 雌とOlder雌の両者に求愛し, size assortative matingが崩れ
ていた。 ウミタナゴ属ではOlder雌獲得を巡ってYOY 雄とOlder雄が競争せ ず, この両者間では縄張り行動は認められなかった。 一方, Older雄とYOY
雄閣のサイズ差が最も小さいオキタナゴでは, Older雌獲得を巡って両者間 で縄張り行動が観察された。 すなわち, 社会構造におけるYOY雄とOlder雄 の競争の有無がsize assortative matingの有無と一致することが明らかと なった。
第7章 謝辞
本研究を取りまとめるに当たり, 終始懇切な御指導と御校闘を頂いた九州 大学農学部教授中園明信博士に深謝の意を表したい。 本研究を行う機会を与 えて下さった九州大学名誉教授塚原博博士に厚く御礼申し上げる。 九州大学 名誉教授奥田武男博士からは, 研究遂行に関して多大なる御援助を頂いた。
心より御礼申し上げる。 また, 筆者が鹿児島大学在学中に研究の機会を与え て下さった鹿児島大学水産学部名誉教授税所俊郎博士に深謝の意を表した い。 鹿児島大学水産学部助教授四宮明彦博士からは, 野外調査の手法につい て手ほどきを受けると共に交尾型魚類の繁殖生態について多くの御教示を頂 いた。 心より御礼申し上げる。 本研究で用いた標本・観察結果の一部は西日 本技術開発株式会社松本豊隆氏に提供して頂いた。 厚く御礼申し上げる。 大 阪市立大学理学部大学院生藤田弘氏にはカサゴの交尾生態について御教示を 受けると共に議論を交わして頂いた。 感謝の意を表したい。 本研究は野外調 査の際に以下の方々に御協力を頂くと共に, 大変御世話になった。 九州大学 農学部付属水産実験所教授古市政幸博士をはじめとする同実験所の職員・学 生の方々, 大分県南海部郡上浦町福泉健二氏と御家族の方々, 向上浦町浅海 井部落の方々, 同上浦町福治部落の方々, 長崎県壱岐郡芦辺町民宿高野の御 家族の方々, 同芦辺町諸吉南触の方々。 これら多くの方々に心より御礼申し 上げる。 本研究を通じて, 九州大学農学部助教授川口栄男博士, 同助手望岡 典隆博士をはじめとする九州大学農学部水産学第二講座の職員・学生の皆様 から暖かい励ましと御援助を頂いた。 九州大学理学部研究生小早川みどり博 士, 千葉県立中央博物館須之部友基博士からは多くの御助言と励ましを頂い た。 これらの方々に心より御礼申し上げる。
第8章 参考文献
Abe, Y. 1969. Systematics and biology of the two species of embiotocid fishes referred to the genus Ditrema in Japan. Japan. J. Ichthyol., 15(3): 105- 121.
Basolo, A. L. 1990a. Female preference for male sword length in the green swordtail, Xiphophorus helleri (Pisces: Poeciliidae). Anim. Behav., 40:
332-338.
Basolo, A. L. 1990b. Female preference predates the evolution of the sword in swordtail fish. Science., 250: 808-810.
Baltz, D. M. 1984. Life history variation among female surfperches (Perciformes: _Embiotocidae). Env. Biol. Fish., 10(3): 159-171.
Bisazza, A. and G. Marin. 1991. Male size and female mate choice in the eastern mosquitofish (Gambusia hoJbrooki: Poeciliidae). Copeia, 1991:
730-735.
Breder, C. M. and D. E. Rosen. 1966. Modes of reproduction in日shes. The Natural History Press, Garden City, New York. pp. 941.
Brembach, M. 1976. Anatomischce Beitrage zur Systematik
lebendgebarender Halbschnabler (Hemirhamphidae, Pisces). Z. Zoo1.
Syst. Evolut. Forsch., 14: 169-177.
Chernyayev, Zh. A. 1971. Some data in the reproduction and development of the cottid fish (Comephorus dybowskii Korotneff). J. Ichthyol., 11: 706- 716.
Chernyayev, Zh. A. 1974. Morphological and ecological features of the “Big Golomyanka" or Baikal oiトfish (Comephorus baicaJensis). J.lchthyol.,
14: 856-868.
Cohen, D. M. and J. P. Wourms. 1976. Microbrotula randalJi, a new
viviparous Ophidioid fish from Samoa and New Hebrides, whose embryos bear trophotaeninae. Proc. Biol. Soc. Wash., 89: 81・98.
Cohen, D. M., R. H. Rosenblatt, and H. G. Moser. 1990. Biology and
description of a bythitid fish from deep-sea thermal vents in the tropical eastern Pacific. Deep-sea Res., Vol. 37 (2): 267・283.
Constantぉ, G. D. 1975. Behavioral ecology of mating in the male gila
topminnow, Poeciliopsis occidentalis (Cyprinodontiformes: Poeciliidae).
Ecology, 56: pp. 966岨973.
Darling, J. D. S., M. L. Noble, and E. Shaw. 1980. Reproductive strategies in the surfperches. 1. Multiple insemination in natural populations of the shiner perch, Cymatogaster aggregata. Evolution, 34(2): 271-277.
Dawkins, R. 1989. 利己的な遺伝子(日高敏隆, 岸由二, 羽田節子, 垂水雄 二 訳) . 紀ノ園屋書店, 東京, pp. 548.
DeMartini, E. E. 1988. Size-assortative courtship and competition in two Embiotocid fishes. Copeia, 1988: 336-344.
Drucker, E.G. and J. S. Jensen. 1991. Functional analysis of a specialized prey processing behavior: winnowing by surfperches (Teleostei:
Embiotocidae). J. Morph. 210(3), 267・287.
deVlaming, V., D. Baltz, S. Anderson, R. Fitzgerald, G. Delahunty , and M.
Barkley. 1983. Aspects of embryo nutrition and excretion among
viviparous embiotocid teleosts: Potential endocrine involvements. Comp.
Biochem. Phisiol., 76A(1): 189・198.
Dobbs, G. H. 1975. Scanning electron microscopy of intraovarian embryos of the viviparous teleost, Micrometrus minimus (Gibbons), (Perciformes:
Embiotocidae). J. Fish. Biol., 7: 209-214.
Ebeling, A. W. and R. N. Bray. 1976. Day versus night activity of reef fishes in a kelp forest off Santa Barbara, Califomia. Fish. Bul1., 74(4): 703・717.
EbelingヲA. W. and D. R. Laur. 1985. The influence of plant cover on surfperch abundance at an offshore temperate reef. Env. Biol. Fish.,
12(3): 169・179.
Ebeling, A. W. and D. R. Laur. 1986. Foraging in the surfperches: resource partitioning or individualistic responses? Env. Biol. Fish., 16(1・3): 123・
133.
Echeverria, T. W. 1986. Sexual dimorphism in four species of rockfish genus Sebastes (S∞rpaenidae). Env. Biol. Fish., 15(3): 181-190.
Eldridge, M. B., J. A. Whipple, M. J. Bowers, B. M. Jarvis, and J. Gold. 1991.
Reproductive perfo口nance of yellowtail rockfish, Sebastes flavidus. Env.
Biol. Fish., 30: 91・102.
Eschmeyer, W. N., E. S. Herald and H. Hammann. 1983. A field guide to Pacific coast fishes of north America. Houghton Mifflin Company,
Boston, pp.336, 48 pl.
Fujita, H. and M. Kohda. 1996. Male mating effort in the viviparous
scorpionfish, Sebastiscus marmoratus. Japan. J. Ichthyol., 43(3): 24-255.
Gardiner, D. M. 1978a. The origin and fate of spermatophores in the
viviparous teleost Cシmatogaster aggregata (Percifo口nes: Embiotocidae).
J. Morph., 155: 157・172.
Gardiner, D. M. 1978b. Cyclic changes in fine structure of the epithelium lining the ovary of the viviparous teleost, Cシmatogaster aggregata (Perciformes: Embiotocidae). J. Morph., 156: 367・380.
Girard, C. F. 1855. Notice upon the viYÎparous fishes inhabiting the Pacific
∞ast of North America, with an enumeration of the species observed.
U. S. A. Proc. Acad. Nat. Sci. Philadelphia., 7: 318-323.
Greene, J. M. and K. L. Brown. 1991. Demographic and genetic
characteristics of multiply inseminated female mosQuitofish (Gambusia
affinis). Copeia, 1991: 434-444.
Gunn, J. S. and R. E. Thresher. 1991. Viviparity and the reproductive ecology of clinid fishes (Clinidae) from temperate Australian waters. Env. Biol.
Fish., 31: 323・344.
Haines, S. E. and J. L. Gould. 1994. Female platys prefer long tails. Nature.,
370: 512.
Hayase, S. and S. Tanaka. 1980a. Growth and reproduction of three species of embiotocid fishes in the Zostera marina belt of Odawa Bay. Nippon Suisan Gakkaishi, 46(9): 1089-1096.
Hayase, S. and S. Tanaka. 1980b. Habitat and distribution of three species of embiotocid fishes in the Zostera marina belt of Odawa Bay. Nippon Suisan Gakkaishi, 46(8): 955-962.
Hayase, S. and S. Tanaka. 198Oc. Feeding ecology of three species of embiotocid fishes in the Zostera marina belt of Odawa Bay. Nippon Suisan Gakkaishi, 46(12): 1469・1476.
Hayase, S. and S. Tanaka. 1981. Inter-specific relations surrounding the three species of embiotocid fishes in the Zostera marina belt of Odawa Bay.
Nippon Suisan Gakkaishi, 47(10): 1317・1322.
Helvey, M. 1982. First observation of courtship behavior in rockfish, genus Sebastes. Copeia, 1982: 763-770.
Hixon, M. A. 1980. Competitive interactions between Califomia reef fishes of the genus EmbiotoC8. Ecology, 61(4): 918-931.
Hixon, M. A. 1981. An experimental analysis of territoriality in the Califomia
reef fish Embíotoca jacksoni (Embiotocidae). Copeia, 1981(3): 653-665.
Hobson, E. S. and J. R. Chess. 1976. Trophic interactions among fishes and
zooplankters near shore at Santa Catalina Island, Califomia. Fish. Bull.,
74(3): 567・598.
Hogarth, P. J. 1976. 胎生(磯野直秀 訳) . 朝倉書店, 東京, pp. 102.
Holbrook, S. J. and R. J. Schmitt. 1988. The combined effects of predation risk and food reward on patch selection. Ecology, 69(1): 125・134.
Hubbs, C. L. 1917. The breeding habits of the viviparus perch, Cシmatogaster.
Copeia, 47: 72・74.
Hubbs, C. L. 1921. The ecology and ife-history of Amphígonopterus aurora
and of other viviparous perches of Califomia. Biol. Bull., 40(4): 181-209.
Igarashi, T. 1961. Histological and cytologi cal changes in the ovary of a viviparous teleost, Neoditrema ransonneti Steindachner during gestation. Bull. Fac. Fish., Hokkaido Univ., 12(3): 181-188, 5 pl.
Igarachi, T. 1962. Morphological changes of the embryo of a viviparous
teleost, Neoditrema ransonneti Steindachner during gestation. Bull.
Fac. Fish., Hokkaido Univ., 13(2): 47・52, 2 pl.
Ishii, S. 1957. Seasonal changes in the ovary and testis of the viviparous teleost, Ditrema temÏnincki. Annot. Zool. Japon., 30(4): 204-210.
Krebs, J. R. and N. B. Davis. 1987. 行動生態学(山岸哲, 厳佐庸 訳)
.
蒼樹書房, 東京, pp. 454.
Krebs, J. R. and N. B. Davis. 1991. 進化から見た行動生態学(山岸哲, 厳佐 庸 監訳) . 蒼樹書房, 東京, pp. 578.
Krejsa, R. J. 1964. Reproductive behavior and sexual dimorphism in the manacled sculpin, Synchírus gíJJibean. Copeia, 1964: 448-450.
Kristofferson, R., S. Broberg, and M. Pekkarinen. 1973. Histology and
physiology of embryotrophe fo口nation, embryonic nutrition and growth
in the eel pout,
Zoarces viviparus (L.).Ann. Zool. Fennici., 10: 467・477.
桑村哲夫. 1988. 魚の子育てと社会 :誰が子育てをすべきか. 海鳴社, 東京.
pp. 136.
益田一・尼岡邦夫・荒賀忠一・ 上野輝禰・吉野哲夫 編. 1984. 日本産魚類 大図鑑. 東海大学出版会, 東京. pp.448, 370 pl.
Mead, G. W., E. Bertelsen, and D. M. Cohen. 1964. Reproduction among deep-sea fishes. Deep-sea Res., 11: 569-596.
Miller, R. R. 1979. Ecology, habits, and relationships of the Middle American cuatro ojos,
Anableps dowi(Pisces: Anablepidae). Copeia, 1979: 82-91.
Miller, R. R. and J. M. Fitzsimons. 1971.
Ameωsplendens,a new genus and species of goodeid fish from western Mexico with remarks on the
classification of the Goodeidae. Copeia, 1971: 1-13.
Mizue,
K.1961a. Studies on
Ditrema temmincki-
1:about the seasonal cycle of mature testis and the spermatogenesis. Rec. Oceanogr. Works Japan (Special Number 5): 67・78, 4 pl.
水江一弘. 1961b. ウミタナゴの研究- III:ウミタナゴの卵巣の成熟並びに季 節的循環に関する研究. 長崎大学水産学部研究報告,11: 1・18.
Mi却e, K. 1962. Studies on marine viviparous teleost, Ditrema temmincki
Bleeker - II: on the annulus of otolith and the growth. Rec. Oceanogr.
Works Japan (Special Number 6): 73唱79, 5 pl.
水江一弘. 1964. ウミタナゴの研究ーIV: ウミタナゴ ・カサゴ, およびメバ ノレにおける卵母細胞の起原について. 長崎大学水産学部研究報告,17: 1-
6, 2 pl.
Mohsen, T. 1962. Un nouveau genre d'Hemirhaphides: Grecarchopterus
nov. gen., base sur des characteres particuliers du systeme urogenital.
BuIl. Aquatic. Biol., 3: 109・120.
中坊徹次(編) . 1993. 日本産魚類検索:全種の同定. 東海大学出版会, 東 京. pp. 1474.
Nakazono, A., Y. Tateda, and H. Tsukahara. 1981. Mating habits of the surfperch, Ditrema temmincki. Japan. J. Ichthyol., 28(2): 122・128.
NeIson, G. 1975. Anatomy of the male urogenitaI organs of Goodea
atripinnis and Characodon JateraJis (Atheriniformes: Cyprinidontidei),
and G. atripinnis courtship. Copeia, 1975: 475-482.
Nelson, J. S. 1994. Fishes of the world. John Wiley and Sons, Inc. New York. pp. 6∞.
大島正満. 1955. 本邦産ウミタナゴに就いて. 動物学雑誌, 64(12): 375-379.
Penrith, M. L. 1969. The systemacics of the fishes of the family Clinidae in Southem Africa. Ann. S. Afr. Mus., 55: 1帽121.
Reznick, D. N. and B. Braun. 1987. Fat cycling in the mosquitofish (Gambusia affinis): fat storage as a reproductive adaptation. Oecologia, 73: 401-413.
Ridley, M. 1983. The explanation of organic diversity: the comparative method and adaptations for mating. Clarendon Press, Oxford, pp.272.
Rosenblatt, R. H. and L. R. Taylor. 1971. The pacific species of the clinid fish tribe Starksiini. Pacific Science, 25: 436・463.
Ryan, M. J. and B. Causey. 1989. “Altemative" mating behavior in the swordtails Xiphophorus nigrensis and Xiphophorus pygmaeus (Pisces:
Poeciliidae). Behav. Ecol. Sociobiol., 24: 341-348.
桜井 真・中園明信. 1990. 水槽内でのウミタナゴの出産と出生後の若魚の 形態変化. Japan. J. Ichthyol., 37(3): 302-307.
Sakurai, M. and A. Nakazono. 1995. Twilight migrations of the temperate Japanese surfperch Neoditrema ransonneti (Embiotocidae). Japan. J.
Ichthyol., 42(3/4): 261・267.
棲井 真・松本豊隆・中園明信. 1996. オキタナゴの交尾生態. 水産増殖、
44(4): 395・405.
Schlupp, 1., J. Parzefall, and M. Schartl. 1991. Male mate choice in mixed bisexual / unisexual breeding complexes of PoeciJia (Teleostei:
Poeciliidae). Ethology, 88: 215・222.
Schultz E. T. 1993. Sexual size dimorphism at birth in Micrometrus minimus (Embiotocidae): A prenatal cost of reproduction. Copeia, 1993(2): 456・
463.
Shaw, E. and J. Allen. 1977. Reproductive behavior in the female shiner perch Cシmatogaster aggregata. Marine Biology, 40: 81-86.
四宮明彦. 1985. 海産カジカ科3種の生殖生理および繁殖生態に関する研究.
北海道大学水産学部, 博士論文., pp.145, 22 pl.
Shinomiya, A. and O. Ezaki. 1991. Mating habits of the rockfish Sebastes inermis. Env. Biol. Fish., 30: 15・22.
Shrode, J. B., L. J. Purcell, and J. S. Stephens, Jr. 1983. Ontogeny of thermal preference in four species of viviparous fishes (Embiotocidae). Env.
Biol. Fish., 9(1): 71・76.
Smith, C. L., C. S. Rand, B. Schaeffer, and J. W. Atz. 1975. Latimeria, the living coelacanth is ovoviviparous. Science, 190: 1105-1106.
Soin, S. G., 1968. Some features in the development of the blenny (Zoarces viviparus L.) in relation to viviparity. Probl. Ichthyol., 8: 222-229.
Suarez, S. S. 1975. The reproductive biology of OgiJbia cayorum, a viviparous brotulid fish. Bull. Mar. Sci., 25(2): 143・173.
Tarp, F. H. 1952. A revision of the family Embiotocidae (The surfperches).
Fish. Bull. Calif. Dep. Fish and Game, 88: 1-99.
Thresher, R. E. 1984. Reproduction in reef fishes. T. F. H. Publications,
Neptune City, New Jersey. pp.399.
Tumer, C. L. 1936. The absorptive processes in the embryos of ParabrotuJa dentiens, a viviparous deep-sea brotulid fish. J. Morph., 59: 313-325.
Turner, C. L. 1937. The trophotaeniae of the Goodeidae, a family of viviparous cyprinodont fishes. J. Morph., 61: 495・523.
Tumer, C. L. 1938. Histological and cytological changes in the ovary of
。matogaster aggregatus during gestation. J. Morph., 62: 351・373.
Tumer, C. L. 1940a. Pericardial sac, trophotaeniae, and alimentary tract in embryos of goodeid fishes. J. Morph., 67: 271・289.
Tumer, C. L. 1940b. Adaptations for viviparity in jenynsiid fishes. J.
Morph., 67: 291・297.
Turner, C. L. 194Oc. Follicular pseudoplacenta andωt modifications in anablepid fishes. J. Morph., 67: 91・105.
Tumer, C. L. 194Od. Pseudoamnion, pseudochorion, and follicular pseudoplacenta in poecillid fishes. J. Morph., 67: 59-89.
Tumer, C. L. 1946. Male secondary sexual characters of Dinematichthys iJuocoeteoides. Copeia, 1946: 92-96.
内田恵太郎. 1938. 胎生魚ウミタナゴの生活史. Zoo1. Mag. (Japan), 50(4):
194.
Veith, W. J. 1980. Viviparity and embryonic adaptations in the teleost αlnus superciJiosus. Can. J. Zoo1., 58: 1-12.
Wamer, R. R. and R. K. Harlan. 1982. Sperm competition and sperm storage as determinants of sexual dimorphium in the dwarf surfperch,
Micrometrus minimus. Evolution, 36(1): 44・55.
Webb, P. W. and J. R. Brett. 1972. Respiratory adaptations of prenatal
young in the ovary of two species of viviparous seaperch, Rhacochi1us vacca and Embiotoca lateraJis. J. Fish. Res. Board Can., 29(11): 1525・
1542.
Wiebe, J. P. 1968. The reproductive cycle of the viviparous seaperch,
αmatogaster aggregata Gibbons. Can. J. Zool., 46: 1221-1234.
Wourms, J. P. 1981. Viviparity: The maternal-fetal relationship in fishes.
Amer. Zool., 21: 473-515.
Wourms, J. P. and O. Bayne. 1973. Development of the viviparous brotulid fish, Dinematichthys iJuocoeteoides. Copeia, 1973: 32-40.
Wourms, J. P. and D. Cohen. 1975. Trophotaeniae, embryonic adaptations in the viviparous ophidioid fish, OJigopus Jonghursti : a study of museum specimens. J. Morph., 147: 385-401.
第9章 SUMMARY
Studies on Courtship and Mating Behavior, and Social Structure During Mating Season of Three Species of Viviparous Japanese
Surfperches
by
Makoto Sakurai
Viviparity occurs in various families of osteichthyan fishes.
However, little attention has been given regarding to mating behavior, social structure, and mating system of viviparous fishes. The purpose of this study was to clarify the mating ecology of three species of Japanese surfperch, Ditrema
temmincki, D. viridis, and Neoditrema ransonneti, by means of specimen collection and underwater observation. The study was carried out at Fukuoka, Nagasaki and Oita Prefectures in Kyushu,
Japan from 1980 to 1987. The results obtained are as follows.
1. Mating season and habitat
The mating season of three species was from September to December. During this time period, surfperches moved to
shallow water in the following pattern: D. temmincki -- rocky reef and Sargussum vegetation; D. viridis -- Zostera marina zone;
N. ransonneti -- mid- and upper layer of the rocky reef.
2. Secondary sexual character
In males of D. temmincki and D. viridis, secondary sexual characters commonly appeared on the anal fin. The characters were an intromittent organ and associated structure, elongated posterior softrays, and a lunar depression at the basal region of the anal fin. In N. ransonneti males, secondary sexual characters appeared in body color, upper and lower lobes of the caudal fin,
and serrations of the premaxillary and dentary, in addition to the intromittent organ on the anal fin. Development of these
characters implied that competition among males was conspicuous in this species.
3. Mean standard length of YOY and Older individuals, age
composition and difference of body size between YOY and Older individuals
Two size groups were recognized during the mating season;
young of the year (YOY) and age greater than one year (Older) groups.
Mean standard length (rnm) of YOY fish were: 78.6 (male) and 79.1 (female) for D. temmincki ; 73.7 and 82.8 for D. viridis;
78.0 and 91.0 for N. ransonneti. Mean standard length of Older fish were: 159.1 and 163.7 for D. temmincki; 120.6 and 125.2 for D. viridis; 98.3 and 108.4 for N. ransonneti.
Age composition of Older males and size ratios (average proportional value of Older male to YOY male = Older male / YOY male: calculated by standard length (SL) and body weight
(BW))
were: 1・4 years and 2.02 in SL (9.09 in
BW)
for D. temmincki; 1・2 years and 1.64 (4.66) for D. viridis ; one year and 1.26 (1.88) for N. ransonne ti. Age composition of Older females and size ratios (average proportional value of Older female to YOY male = Older female / YOY male) were: 1・3 years and 2.08 (9.22) for α
temmincki ; 1-3 years and 1.70 (5.09) for D. viridis ; 1・2 years
and 1.39 (2.55) for N. ransonneti. For the latter species, few Older males and females were observed and most were only one year old, therefore, it was suspected that YOY males and females would die before the mating season of next year.
4. Social structure, function of territory, and relationship between Older and YOY males
During mating season, males of all three species maintained territories in which females moved among freely. Ditrema and N ransonneti differed in territorial behavior between YOY and Older males. In both species of Ditrema, territories of YOY and Older males overlapped and territorial behavior was not observed. In N. ransonneti, territories did not overlap and frequent territorial behavior was observed between males.
Territories were considered to function as courtship and mating sites in all three species. Territories also functioned as
feeding areas in D. temmincki and D. viridis.
Microhabitat of D. temmincki differed between Older and YOY fish. In D. viridis, YOY fish displayed appeasement behavior
toward Older males. This behavior suggests a strategy by which both Older males and YOY fish coexisted within the Z ostera marina zone. In N. ransonneti, Older and YOY fish coexisted on the shallow rocky reef but appeasement behavior was not
observed.
5. Frequency of feeding
Although D. temmincki and D. viridis fed on benthic small
invertebrates and N. ransonneti fed on zooplankton, some
features of the frequency of feeding were common among these species. Among Older fish, males devoted time to territorial and courtship behavior. Therefore, the frequency of feeding of Older males was lower than that of Older females. Higher feeding frequency by females may be because energy is directed towards reproductive investment of pregnancy.
In YOY fishes, a difference in feeding activity was found
between Ditrema and N. ransonneti. In Ditrema, feeding frequency of YOY males and females was higher than in Older fish. This may be because they needed to invest energy to increase reproductive success in the following year. N.
ransonneti fed only when zooplankton were present. YOY females fed frequently, like Ditrema., however, in YOY males the